

Consequences of COVID-19 on Adipose Tissue Signatures
 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Clinical and Metabolic Characteristics of Study Cohort
2.2. Smaller Average Adipocyte Size in VAT and SAT of COVID-19 Infected Patients
2.3. History of COVID-19 Infection Is Associated with a Distinct AT Gene Expression Signature That Is Fat Depot-Specific
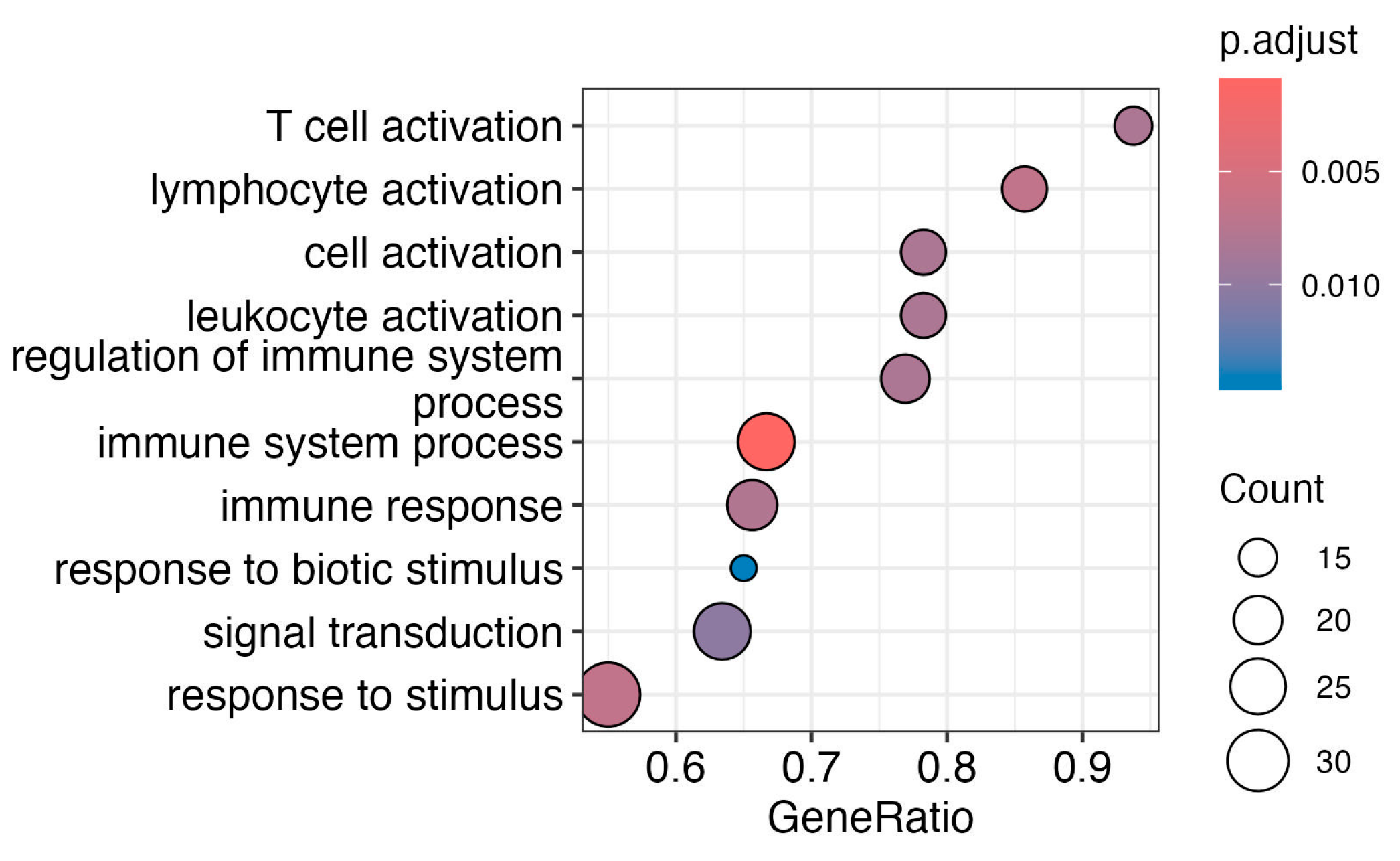
2.4. Subcutaneous Adipose Tissue Immune System Pathways Are Affected by Vaccination
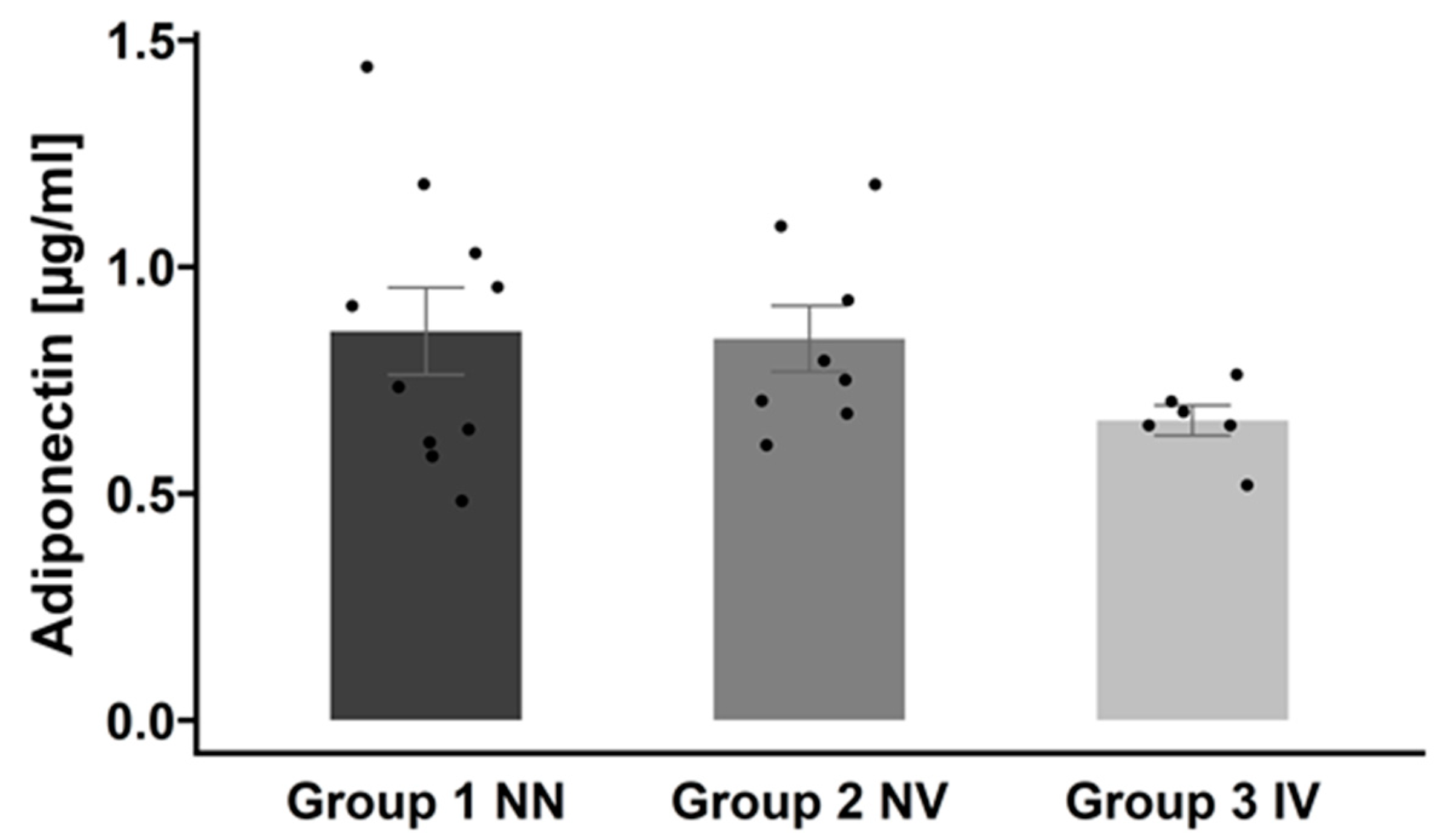
2.5. Effects of COVID-19 History on Serum Protein Profile
3. Discussion
Limitations
4. Materials and Methods
4.1. Baseline Characteristics of the Subjects
4.2. SARS-CoV-2 Antibody Detection
4.3. Viral RNA Detection in Adipose Tissue
4.4. Characterization of Adipose Tissue Samples
4.5. RNA-Sequencing of Adipose Tissue
4.6. Serum Proteomics Profiling
4.7. Serum Adiponectin Measurement
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reiterer, M.; Rajan, M.; Gómez-Banoy, N.; Lau, J.D.; Gomez-Escobar, L.G.; Ma, L.; Gilani, A.; Alvarez-Mulett, S.; Sholle, E.T.; Chandar, V.; et al. Hyperglycemia in Acute COVID-19 Is Characterized by Insulin Resistance and Adipose Tissue Infectivity by SARS-CoV-2. Cell Metab. 2021, 33, 2174–2188.e5. [Google Scholar] [CrossRef]
- Zickler, M.; Stanelle-Bertram, S.; Ehret, S.; Heinrich, F.; Lange, P.; Schaumburg, B.; Kouassi, N.M.; Beck, S.; Jaeckstein, M.Y.; Mann, O.; et al. Replication of SARS-CoV-2 in Adipose Tissue Determines Organ and Systemic Lipid Metabolism in Hamsters and Humans. Cell Metab. 2022, 34, 1–2. [Google Scholar] [CrossRef]
- Palaiodimos, L.; Kokkinidis, D.G.; Li, W.; Karamanis, D.; Ognibene, J.; Arora, S.; Southern, W.N.; Mantzoros, C.S. Severe Obesity, Increasing Age and Male Sex Are Independently Associated with Worse in-Hospital Outcomes, and Higher in-Hospital Mortality, in a Cohort of Patients with COVID-19 in the Bronx, New York. Metabolism 2020, 108, 154262. [Google Scholar] [CrossRef]
- Price-Haywood, E.G.; Burton, J.; Fort, D.; Seoane, L. Hospitalization and Mortality among Black Patients and White Patients with COVID-19. N. Engl. J. Med. 2020, 382, 2534–2543. [Google Scholar] [CrossRef] [PubMed]
- Drucker, D.J. Diabetes, Obesity, Metabolism, and SARS-CoV-2 Infection: The End of the Beginning. Cell Metab. 2021, 33, 479–498. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Zheng, K.I.; Wang, X.B.; Sun, Q.F.; Pan, K.H.; Wang, T.Y.; Chen, Y.P.; Targher, G.; Byrne, C.D.; George, J.; et al. Obesity Is a Risk Factor for Greater COVID-19 Severity. Diabetes Care 2020, 43, E72–E74. [Google Scholar] [CrossRef] [PubMed]
- Kassir, R. Risk of COVID-19 for Patients with Obesity. Obes. Rev. 2020, 21, e13034. [Google Scholar] [CrossRef] [PubMed]
- Kruglikov, I.L.; Scherer, P.E. The Role of Adipocytes and Adipocyte-Like Cells in the Severity of COVID-19 Infections. Obesity 2020, 28, 1187–1190. [Google Scholar] [CrossRef] [PubMed]
- Saccon, T.D.; Mousovich-Neto, F.; Ludwig, R.G.; Carregari, V.C.; dos Anjos Souza, A.B.; dos Passos, A.S.C.; Martini, M.C.; Barbosa, P.P.; de Souza, G.F.; Muraro, S.P.; et al. SARS-CoV-2 Infects Adipose Tissue in a Fat Depot- and Viral Lineage-Dependent Manner. Nat. Commun. 2022, 13, 5722. [Google Scholar] [CrossRef]
- Leipzig Obesity Bio Bank (LOBB)—Helmholtz Munich. Available online: https://www.helmholtz-munich.de/en/hi-mag/cohort/leipzig-obesity-bio-bank-lobb (accessed on 30 January 2024).
- Perrotta, F.; Scialò, F.; Mallardo, M.; Signoriello, G.; D’Agnano, V.; Bianco, A.; Daniele, A.; Nigro, E. Adiponectin, Leptin, and Resistin Are Dysregulated in Patients Infected by SARS-CoV-2. Int. J. Mol. Sci. 2023, 24, 1131. [Google Scholar] [CrossRef]
- Conte, C.; Cipponeri, E.; Roden, M. Diabetes Mellitus, Energy Metabolism and COVID-19. Endocr. Rev. 2023, bnad032. [Google Scholar] [CrossRef] [PubMed]
- Bastard, L.; Rech, J.S.; Senet, P.; Soria, A.; Fellahi, S.; Vatier, C.; Georgin-Lavialle, S.; Bastard, J.P. Does Adipose Tissue Contribute to Acute Infection-Related Inflammation in COVID-19? Nutr. Metab. Cardiovasc. Dis. 2023, 33, 2527–2528. [Google Scholar] [CrossRef]
- Moser, J.; Emous, M.; Heeringa, P.; Rodenhuis-Zybert, I.A. Mechanisms and Pathophysiology of SARS-CoV-2 Infection of the Adipose Tissue. Trends Endocrinol. Metab. 2023, 34, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Basolo, A.; Poma, A.M.; Bonuccelli, D.; Proietti, A.; Macerola, E.; Ugolini, C.; Torregrossa, L.; Giannini, R.; Vignali, P.; Basolo, F.; et al. Adipose Tissue in COVID-19: Detection of SARS-CoV-2 in Adipocytes and Activation of the Interferon-Alpha Response. J. Endocrinol. Investig. 2022, 45, 1021. [Google Scholar] [CrossRef]
- Martínez-Colón, G.J.; Ratnasiri, K.; Chen, H.; Jiang, S.; Zanley, E.; Rustagi, A.; Verma, R.; Chen, H.; Andrews, J.R.; Mertz, K.D.; et al. SARS-CoV-2 Infection Drives an Inflammatory Response in Human Adipose Tissue through Infection of Adipocytes and Macrophages. Sci. Transl. Med. 2022, 14, eabm9151. [Google Scholar] [CrossRef]
- Ryan, P.M.D.; Caplice, N.M. Is Adipose Tissue a Reservoir for Viral Spread, Immune Activation, and Cytokine Amplification in Coronavirus Disease 2019? Obesity 2020, 28, 1191–1194. [Google Scholar] [CrossRef] [PubMed]
- Koethe, J.R.; Lagathu, C.; Lake, J.E.; Domingo, P.; Calmy, A.; Falutz, J.; Brown, T.T.; Capeau, J. HIV and Antiretroviral Therapy-Related Fat Alterations. Nat. Rev. Dis. Primers 2020, 6, 48. [Google Scholar] [CrossRef]
- Soares, V.C.; Dias, S.S.G.; Santos, J.C.; Azevedo-Quintanilha, I.G.; Moreira, I.B.G.; Sacramento, C.Q.; Fintelman-Rodrigues, N.; Temerozo, J.R.; da Silva, M.A.N.; Barreto-Vieira, D.F.; et al. Inhibition of the SREBP Pathway Prevents SARS-CoV-2 Replication and Inflammasome Activation. Life Sci. Alliance 2023, 6, e202302049. [Google Scholar] [CrossRef]
- da Silva Gomes Dias, S.; Soares, V.C.; Ferreira, A.C.; Sacramento, C.Q.; Fintelman-Rodrigues, N.; Temerozo, J.R.; Teixeira, L.; da Silva, M.A.N.; Barreto, E.; Mattos, M.; et al. Lipid Droplets Fuel SARS-CoV-2 Replication and Production of Inflammatory Mediators. PLoS Pathog. 2020, 16, e1009127. [Google Scholar] [CrossRef]
- D’Avila, H.; Lima, C.N.R.; Rampinelli, P.G.; Mateus, L.C.O.; de Sousa Silva, R.V.; Correa, J.R.; de Almeida, P.E. Lipid Metabolism Modulation during SARS-CoV-2 Infection: A Spotlight on Extracellular Vesicles and Therapeutic Prospects. Int. J. Mol. Sci. 2024, 25, 640. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Fos-Domenech, J.; Serra, D.; Herrero, L.; Sánchez-Infantes, D. White Adipose Tissue Dysfunction in Obesity and Aging. Biochem. Pharmacol. 2021, 192, 114723. [Google Scholar] [CrossRef]
- Roncagalli, R.; Cucchetti, M.; Jarmuzynski, N.; Grégoire, C.; Bergot, E.; Audebert, S.; Baudelet, E.; Menoita, M.G.; Joachim, A.; Durand, S.; et al. The Scaffolding Function of the RLT PR Protein Explains Its Essential Role for CD28 Co-Stimulation in Mouse and Human T Cells. J. Exp. Med. 2016, 213, 2437–2457. [Google Scholar] [CrossRef]
- Atschekzei, F.; Jacobs, R.; Wetzke, M.; Sogkas, G.; Schröder, C.; Ahrenstorf, G.; Dhingra, A.; Ott, H.; Baumann, U.; Schmidt, R.E. A Novel CARMIL2 Mutation Resulting in Combined Immunodeficiency Manifesting with Dermatitis, Fungal, and Viral Skin Infections As Well as Selective Antibody Deficiency. J. Clin. Immunol. 2019, 39, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pomar, N.; Cunill, V.; Segura-Guerrero, M.; Pol-Pol, E.; Escobar Oblitas, D.; Pons, J.; Ayestarán, I.; Pruneda, P.C.; Losada, I.; Toledo-Pons, N.; et al. Hyperinflammatory Immune Response in COVID-19: Host Genetic Factors in Pyrin Inflammasome and Immunity to Virus in a Spanish Population from Majorca Island. Biomedicines 2023, 11, 2548. [Google Scholar] [CrossRef] [PubMed]
- Mothes, R.; Pascual-Reguant, A.; Koehler, R.; Liebeskind, J.; Liebheit, A.; Bauherr, S.; Philipsen, L.; Dittmayer, C.; Laue, M.; von Manitius, R.; et al. Distinct Tissue Niches Direct Lung Immunopathology via CCL18 and CCL21 in Severe COVID-19. Nat. Commun. 2023, 14, 791. [Google Scholar] [CrossRef] [PubMed]
- Tveita, A.; Murphy, S.L.; Holter, J.C.; Kildal, A.B.; Michelsen, A.E.; Lerum, T.V.; Kaarbø, M.; Heggelund, L.; Holten, A.R.; Finbråten, A.K.; et al. High Circulating Levels of the Homeostatic Chemokines CCL19 and CCL21 Predict Mortality and Disease Severity in COVID-19. J. Infect. Dis. 2022, 226, 2150–2160. [Google Scholar] [CrossRef] [PubMed]
- Förster, R.; Schubel, A.; Breitfeld, D.; Kremmer, E.; Renner-Müller, I.; Wolf, E.; Lipp, M. CCR7 Coordinates the Primary Immune Response by Establishing Functional Microenvironments in Secondary Lymphoid Organs. Cell 1999, 99, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Hellmann, J.; Sansbury, B.E.; Holden, C.R.; Tang, Y.; Wong, B.; Wysoczynski, M.; Rodriguez, J.; Bhatnagar, A.; Hill, B.G.; Spite, M. CCR7 Maintains Nonresolving Lymph Node and Adipose Inflammation in Obesity. Diabetes 2016, 65, 2268–2281. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, P.S.; Wimmers, F.; Mok, C.K.P.; Perera, R.A.P.M.; Scott, M.; Hagan, T.; Sigal, N.; Feng, Y.; Bristow, L.; Tsang, O.T.Y.; et al. Systems Biological Assessment of Immunity to Mild versus Severe COVID-19 Infection in Humans. Science 2020, 369, 1210–1220. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.E.; dos Santos, C.C.; O’Gorman, D.B.; Carter, D.E.; Patterson, E.K.; Slessarev, M.; Martin, C.; Daley, M.; Miller, M.R.; Cepinskas, G.; et al. Transcriptional Profiling of Leukocytes in Critically Ill COVID19 Patients: Implications for Interferon Response and Coagulation. Intensive Care Med. Exp. 2020, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Thair, S.A.; He, Y.D.; Hasin-Brumshtein, Y.; Sakaram, S.; Pandya, R.; Toh, J.; Rawling, D.; Remmel, M.; Coyle, S.; Dalekos, G.N.; et al. Transcriptomic Similarities and Differences in Host Response between SARS-CoV-2 and Other Viral Infections. iScience 2020, 24, 101947. [Google Scholar] [CrossRef] [PubMed]
- Overmyer, K.A.; Shishkova, E.; Miller, I.J.; Balnis, J.; Bernstein, M.N.; Peters-Clarke, T.M.; Meyer, J.G.; Quan, Q.; Muehlbauer, L.K.; Trujillo, E.A.; et al. Large-Scale Multi-Omic Analysis of COVID-19 Severity. Cell Syst. 2021, 12, 23–40.e7. [Google Scholar] [CrossRef] [PubMed]
- McClain, M.T.; Constantine, F.J.; Henao, R.; Liu, Y.; Tsalik, E.L.; Burke, T.W.; Steinbrink, J.M.; Petzold, E.; Nicholson, B.P.; Rolfe, R.; et al. Dysregulated Transcriptional Responses to SARS-CoV-2 in the Periphery. Nat. Commun. 2021, 12, 1079. [Google Scholar] [CrossRef] [PubMed]
- Ng, D.L.; Granados, A.C.; Santos, Y.A.; Servellita, V.; Goldgof, G.M.; Meydan, C.; Sotomayor-Gonzalez, A.; Levine, A.G.; Balcerek, J.; Han, L.M.; et al. A Diagnostic Host Response Biosignature for COVID-19 from RNA Profiling of Nasal Swabs and Blood. Sci. Adv. 2021, 7, eabe5984. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Fong, S.; Poh, C.; Carissimo, G.; Yeo, N.K.; Amrun, S.N.; Goh, Y.S.; Lim, J.; Xu, W.; Chee, R.S.; et al. Asymptomatic COVID-19: Disease Tolerance with Efficient Anti-Viral Immunity against SARS-CoV-2. EMBO Mol. Med. 2021, 13, e14045. [Google Scholar] [CrossRef]
- Hong, R.; Lai, N.; Xiong, E.; Ouchida, R.; Sun, J.; Zhou, Y.; Tang, Y.; Hikida, M.; Tsubata, T.; Tagawa, M.; et al. Distinct Roles of BCNP1 in B-Cell Development and Activation. Int. Immunol. 2020, 32, 17–26. [Google Scholar] [CrossRef]
- Pickett, S.J.; Deen, D.; Pyle, A.; Santibanez-Koref, M.; Hudson, G. Interactions between Nuclear and Mitochondrial SNPs and Parkinson’s Disease Risk. Mitochondrion 2022, 63, 85–88. [Google Scholar] [CrossRef]
- Meng, L.; Shen, L.; Ji, H.F. Impact of Infection on Risk of Parkinson’s Disease: A Quantitative Assessment of Case-Control and Cohort Studies. J. NeuroVirology 2019, 25, 221–228. [Google Scholar] [CrossRef]
- Pavlasova, G.; Mraz, M. The Regulation and Function of CD20: An “Enigma” of B-Cell Biology and Targeted Therapy. Haematologica 2020, 105, 1494–1506. [Google Scholar] [CrossRef]
- Yu, L.; Zhang, H.; Guan, X.; Qin, D.; Zhou, J.; Wu, X. Loss of ESRP1 Blocks Mouse Oocyte Development and Leads to Female Infertility. Development 2021, 148, dev196931. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Aguayo, V.; Jiménez-Vacas, J.M.; Sáez-Martínez, P.; Gómez-Gómez, E.; López-Cánovas, J.L.; Garrido-Sánchez, L.; Herrera-Martínez, A.D.; García-Bermejo, L.; MacÍas-González, M.; López-Miranda, J.; et al. Influence of Obesity in the MiRNome: MiR-4454, a Key Regulator of Insulin Response Via Splicing Modulation in Prostate. J. Clin. Endocrinol. Metab. 2021, 106, e469–e484. [Google Scholar] [CrossRef] [PubMed]
- Vadlamudi, Y.; Kang, S.C. Silencing ESRP1 Expression Promotes Caspase-Independent Cell Death via Nuclear Translocation of AIF in Colon Cancer Cells. Cell. Signal. 2022, 91, 110237. [Google Scholar] [CrossRef]
- Saeedifar, A.M.; Ghorban, K.; Ganji, A.; Mosayebi, G.; Gholami, M.; Dadmanesh, M.; Rouzbahani, N.H. Evaluation of Tcell Exhaustion Based on the Expression of EOMES, Tbet and Co-Inhibitory Receptors in Severe and Non-Severe COVID-19 Patients. Gene Rep. 2023, 31, 101747. [Google Scholar] [CrossRef]
- Geginat, J.; Vasco, C.; Gruarin, P.; Bonnal, R.; Rossetti, G.; Silvestri, Y.; Carelli, E.; Pulvirenti, N.; Scantamburlo, M.; Moschetti, G.; et al. Eomesodermin-Expressing Type 1 Regulatory (EOMES+ Tr1)-like T Cells: Basic Biology and Role in Immune-Mediated Diseases. Eur. J. Immunol. 2023, 53, e2149775. [Google Scholar] [CrossRef]
- Van De Veerdonk, F.L.; Janssen, N.A.F.; Grondman, I.; De Nooijer, A.H.; Koeken, V.A.C.M.; Matzaraki, V.; Boahen, C.K.; Kumar, V.; Kox, M.; Koenen, H.J.P.M.; et al. A Systems Approach to Inflammation Identifies Therapeutic Targets in SARS-CoV-2 Infection. medRxiv 2020. [Google Scholar] [CrossRef]
- Wueest, S.; Konrad, D. The Controversial Role of IL-6 in Adipose Tissue on Obesity-Induced Dysregulation of Glucose Metabolism. Am. J. Physiol. Endocrinol. Metab. 2020, 319, E607–E613. [Google Scholar] [CrossRef] [PubMed]
- Jonas, M.I.; Kurylowicz, A.; Bartoszewicz, Z.; Lisik, W.; Jonas, M.; Wierzbicki, Z.; Chmura, A.; Pruszczyk, P.; Puzianowska-Kuznicka, M. Interleukins 6 and 15 Levels Are Higher in Subcutaneous Adipose Tissue, but Obesity Is Associated with Their Increased Content in Visceral Fat Depots. Int. J. Mol. Sci. 2015, 16, 25817. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Tleyjeh, I.M.; Kashour, Z.; Riaz, M.; Hassett, L.; Veiga, V.C.; Kashour, T. Efficacy and Safety of Tocilizumab in COVID-19 Patients: A Living Systematic Review and Meta-Analysis, First Update. Clin. Microbiol. Infect. 2021, 27, 1076–1082. [Google Scholar] [CrossRef]
- Ghosn, L.; Chaimani, A.; Evrenoglou, T.; Davidson, M.; Graña, C.; Schmucker, C.; Bollig, C.; Henschke, N.; Sguassero, Y.; Nejstgaard, C.H.; et al. Interleukin-6 Blocking Agents for Treating COVID-19: A Living Systematic Review. Cochrane Database Syst. Rev. 2021, 3. [Google Scholar] [CrossRef]
- Nagao, Y.; Platero, J.S.; Waheed, A.; Sly, W.S. Human Mitochondrial Carbonic Anhydrase: CDNA Cloning, Expression, Subcellular Localization, and Mapping to Chromosome 16. Proc. Natl. Acad. Sci. USA 1993, 90, 7623–7627. [Google Scholar] [CrossRef]
- Van Karnebeek, C.D.; Sly, W.S.; Ross, C.J.; Salvarinova, R.; Yaplito-Lee, J.; Santra, S.; Shyr, C.; Horvath, G.A.; Eydoux, P.; Lehman, A.M.; et al. Mitochondrial Carbonic Anhydrase VA Deficiency Resulting from CA5A Alterations Presents with Hyperammonemia in Early Childhood. Am. J. Hum. Genet. 2014, 94, 453–461. [Google Scholar] [CrossRef]
- Queen, A.; Khan, P.; Azam, A.; Hassan, M.I. Understanding the Role and Mechanism of Carbonic Anhydrase V in Obesity and Its Therapeutic Implications. Curr. Protein Pept. Sci. 2018, 19, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Bauer, A.; Nano, J.; Petrera, A.; Rathmann, W.; Herder, C.; Hauck, S.M.; Sun, B.B.; Hoyer, A.; Peters, A.; et al. Associations of Plasma Proteomics with Type 2 Diabetes and Related Traits: Results from the Longitudinal KORA S4/F4/FF4 Study. Diabetologia 2023, 66, 1655–1668. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.S. The Role of Carbonic Anhydrase in Hepatic Glucose Production. Curr. Diabetes Rev. 2018, 14, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Mercuri, L.; Papadimitriou, D.; Galdikas, A.; Roadknight, G.; Davies, J.; Glampson, B.; Mayer, E.; Hill, N.E.; Rea, R. Increase in Hypoglycaemia and Hyperglycaemia in People with Diabetes Admitted to Hospital during COVID-19 Pandemic. J. Diabetes Its Complicat. 2023, 37, 108474. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, B.; Lu, H.; Chen, Q.; Li, J.C. Identification of Novel Candidate Biomarkers for Acute Myocardial Infarction by the Olink Proteomics Platform. Clin. Chim. Acta 2023, 548, 117506. [Google Scholar] [CrossRef] [PubMed]
- Zuin, M.; Rigatelli, G.; Battisti, V.; Costola, G.; Roncon, L.; Bilato, C. Increased Risk of Acute Myocardial Infarction after COVID-19 Recovery: A Systematic Review and Meta-Analysis. Int. J. Cardiol. 2023, 372, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Ho, G.; Ali, A.; Takamatsu, Y.; Wada, R.; Masliah, E.; Hashimoto, M. Diabetes, Inflammation, and the Adiponectin Paradox: Therapeutic Targets in SARS-CoV-2. Drug Discov. Today 2021, 26, 2036. [Google Scholar] [CrossRef]
- Fasshauer, M.; Blüher, M. Adipokines in Health and Disease. Trends Pharmacol. Sci. 2015, 36, 461–470. [Google Scholar] [CrossRef]
- Choi, H.M.; Doss, H.M.; Kim, K.S. Multifaceted Physiological Roles of Adiponectin in Inflammation and Diseases. Int. J. Mol. Sci. 2020, 21, 1219. [Google Scholar] [CrossRef] [PubMed]
- Vozarova, B.; Weyer, C.; Hanson, K.; Tataranni, P.A.; Bogardus, C.; Pratley, R.E. Circulating Interleukin-6 in Relation to Adiposity, Insulin Action, and Insulin Secretion. Obes. Res. 2001, 9, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Yang, S.; Xiao, H.; Wang, M.; Ye, J.; Cao, L.; Sun, G. Role of Adiponectin in Cardiovascular Diseases Related to Glucose and Lipid Metabolism Disorders. Int. J. Mol. Sci. 2022, 23, 15627. [Google Scholar] [CrossRef] [PubMed]
- Hashizume, M.; Mihara, M. IL-6 and Lipid Metabolism. Inflamm. Regen. 2011, 31, 325–333. [Google Scholar] [CrossRef][Green Version]
- Glund, S.; Krook, A. Role of Interleukin-6 Signalling in Glucose and Lipid Metabolism. Acta Physiol. 2008, 192, 37–48. [Google Scholar] [CrossRef]
- Lehrskov, L.L.; Christensen, R.H. The Role of Interleukin-6 in Glucose Homeostasis and Lipid Metabolism. Semin. Immunopathol. 2019, 41, 491–499. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Minniti, G.; Miola, V.F.B.; Haber, J.F.D.S.; Bueno, P.C.D.S.; de Argollo Haber, L.S.; Girio, R.S.J.; Detregiachi, C.R.P.; Dall’Antonia, C.T.; Rodrigues, V.D.; et al. Organokines in COVID-19: A Systematic Review. Cells 2023, 12, 1349. [Google Scholar] [CrossRef]
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-Sensitive Obesity. Am. J. Physiol. -Endocrinol. Metab. 2010, 299, E506–E515. [Google Scholar] [CrossRef]
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2014, 37, S81–S90. [Google Scholar] [CrossRef]
- PEI Dossier Coronavirus SARS-CoV-2 and COVID-19Coronavirus and COVID-19—Paul-Ehrlich-Institut. Available online: https://www.pei.de/EN/newsroom/dossier/coronavirus/coronavirus-content.html?nn=164146&cms_pos=2 (accessed on 24 May 2023).
- Mardinoglu, A.; Heiker, J.T.; Gärtner, D.; Björnson, E.; Schön, M.R.; Flehmig, G.; Klöting, N.; Krohn, K.; Fasshauer, M.; Stumvoll, M.; et al. Extensive Weight Loss Reveals Distinct Gene Expression Changes in Human Subcutaneous and Visceral Adipose Tissue. Sci. Rep. 2015, 5, 14841. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast Universal RNA-Seq Aligner. Bioinformatics 2013, 29, 15. [Google Scholar] [CrossRef]
- Babraham Bioinformatics—FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 30 January 2024).
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An Efficient General Purpose Program for Assigning Sequence Reads to Genomic Features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Leek, J.T.; Evan, J.; Parker, H.S.; Fertig, E.J.; Jaffe, A.E.; Zhang, Y.; Storey, J.D.; Collado Torres, L. Sva: Surrogate Variable Analysis 2023. doi:10.18129/B9.bioc.sva, R package version 3.50.0. Available online: https://bioconductor.org/packages/sva (accessed on 30 January 2024).
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-Tailed Prior Distributions for Sequence Count Data: Removing the Noise and Preserving Large Differences. Bioinformatics 2019, 35, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. ClusterProfiler 4.0: A Universal Enrichment Tool for Interpreting Omics Data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, R.; Sugawara, H.; Shumway, M.; on behalf of the International Nucleotide Sequence Database Collaboration. The Sequence Read Archive. Nucleic Acids Res. 2011, 39, D19–D21. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group 1 Controls (NN) | Group 2 Vaccination (NV) | Group 3 Infection/Vaccination (IV) | |
|---|---|---|---|
| n = 10 | n = 10 | n = 7 | |
| Men/women | 5/5 | 5/5 | 2/5 |
| Age [years] | 42.8 ± 10.4 | 43.0 ± 9.5 | 42.7 ± 8.9 |
| Body weight [kg] | 138.5 ± 34.3 | 141.8 ± 20.3 | 127.0 ± 21.9 |
| Height [m] | 1.7 ± 0.1 | 1.7 ± 0.1 | 1.7 ± 0.0 |
| BMI [kg/m2] | 44.1 ± 7.1 | 44.3 ± 4.9 | 43.8 ± 5.8 |
| Body fat mass [%] | 48.6 ± 9.6 | 46.9 ± 8.1 | 50.3 ± 7.8 |
| Creatinine [μmol/L] | 73.1 ± 17.6 | 76.3 ± 12.8 | 75.1 ± 10.5 |
| C-reactive protein [mg/L] | 9.5 ± 10.2 | 6.6 ± 7.1 | 4.0 ± 1.5 |
| Albumin [g/dl] | 4.5 ± 0.3 | 4.6 ± 0.2 | 4.5 ± 0.2 |
| FPG [mmol/L] | 6.6 ± 3.9 | 6.2 ± 1.6 | 4.7 ± 0.4 |
| HOMA-IR Index | 9.6 ± 9.8 | 4.8 ± 5.6 | 1.8 ± 0.9 |
| HbA1c [%] | 6.1 ± 1.4 | 6.4 ± 1.2 | 5.5 ± 0.5 |
| Total cholesterol [mmol/L] | 4.6 ± 1.3 | 4.3 ± 1.3 | 4.3 ± 0.6 |
| HDL cholesterol [mmol/L] | 1.5 ± 0.6 | 1.2 ± 0.2 | 1.3 ± 0.3 |
| LDL cholesterol [mmol/L] | 2.8 ± 1.1 | 2.8 ± 1.2 | 2.6 ± 0.7 |
| Triglycerides [mmol/L] | 1.5 ± 1.5 | 1.6 ± 0.5 | 1.4 ± 0.9 |
| Diabetes status | |||
| Normal glucose tolerance [%] | 30 | 40 | 71 |
| Type 2 diabetes [%] | 40 | 50 | 14 |
| Impaired glucose tolerance [%] | 30 | 10 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupka, S.; Hoffmann, A.; Jasaszwili, M.; Dietrich, A.; Guiu-Jurado, E.; Klöting, N.; Blüher, M. Consequences of COVID-19 on Adipose Tissue Signatures. Int. J. Mol. Sci. 2024, 25, 2908. https://doi.org/10.3390/ijms25052908
Krupka S, Hoffmann A, Jasaszwili M, Dietrich A, Guiu-Jurado E, Klöting N, Blüher M. Consequences of COVID-19 on Adipose Tissue Signatures. International Journal of Molecular Sciences. 2024; 25(5):2908. https://doi.org/10.3390/ijms25052908
Chicago/Turabian StyleKrupka, Sontje, Anne Hoffmann, Mariami Jasaszwili, Arne Dietrich, Esther Guiu-Jurado, Nora Klöting, and Matthias Blüher. 2024. "Consequences of COVID-19 on Adipose Tissue Signatures" International Journal of Molecular Sciences 25, no. 5: 2908. https://doi.org/10.3390/ijms25052908
APA StyleKrupka, S., Hoffmann, A., Jasaszwili, M., Dietrich, A., Guiu-Jurado, E., Klöting, N., & Blüher, M. (2024). Consequences of COVID-19 on Adipose Tissue Signatures. International Journal of Molecular Sciences, 25(5), 2908. https://doi.org/10.3390/ijms25052908