

Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety

Abstract
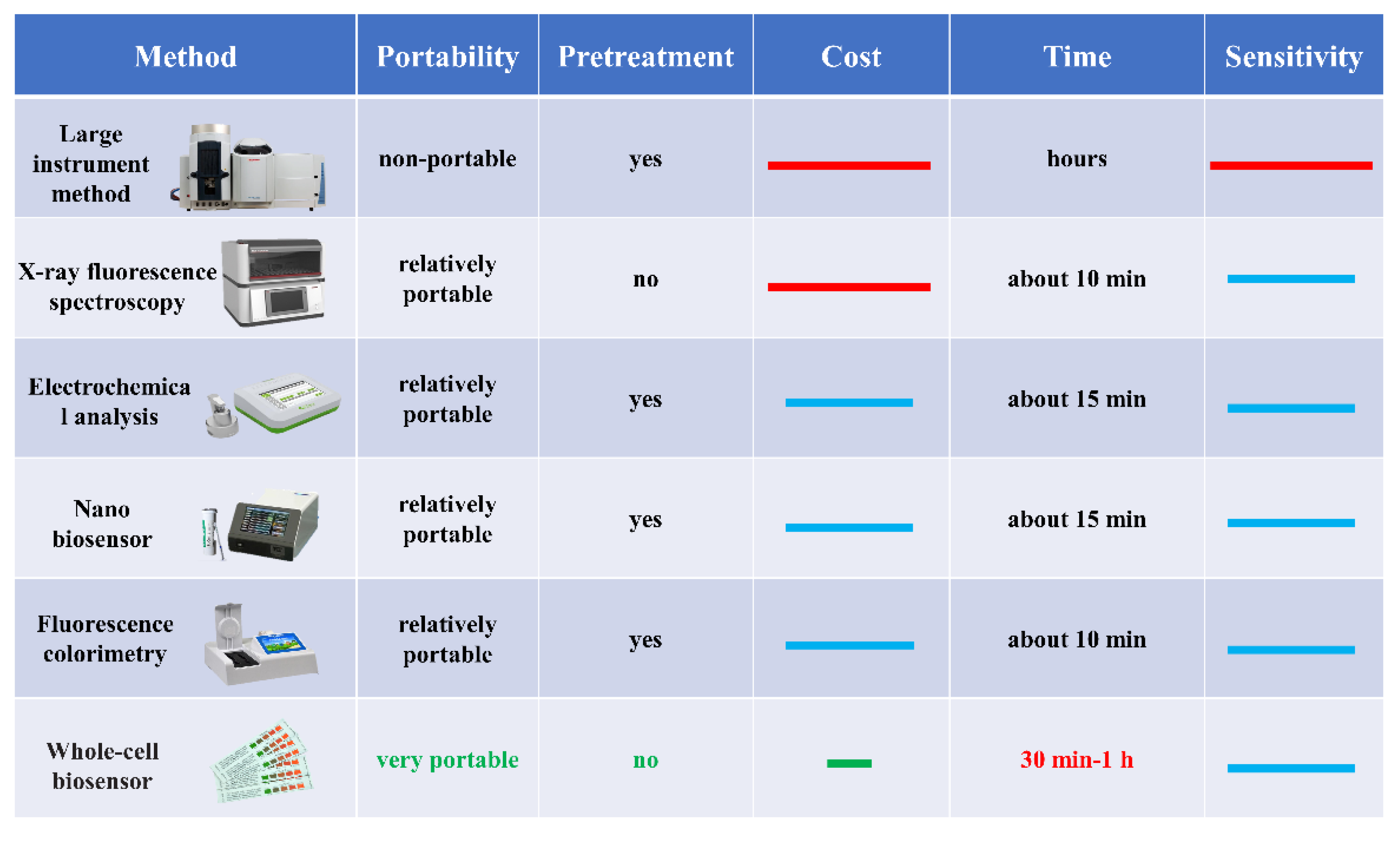
1. Introduction
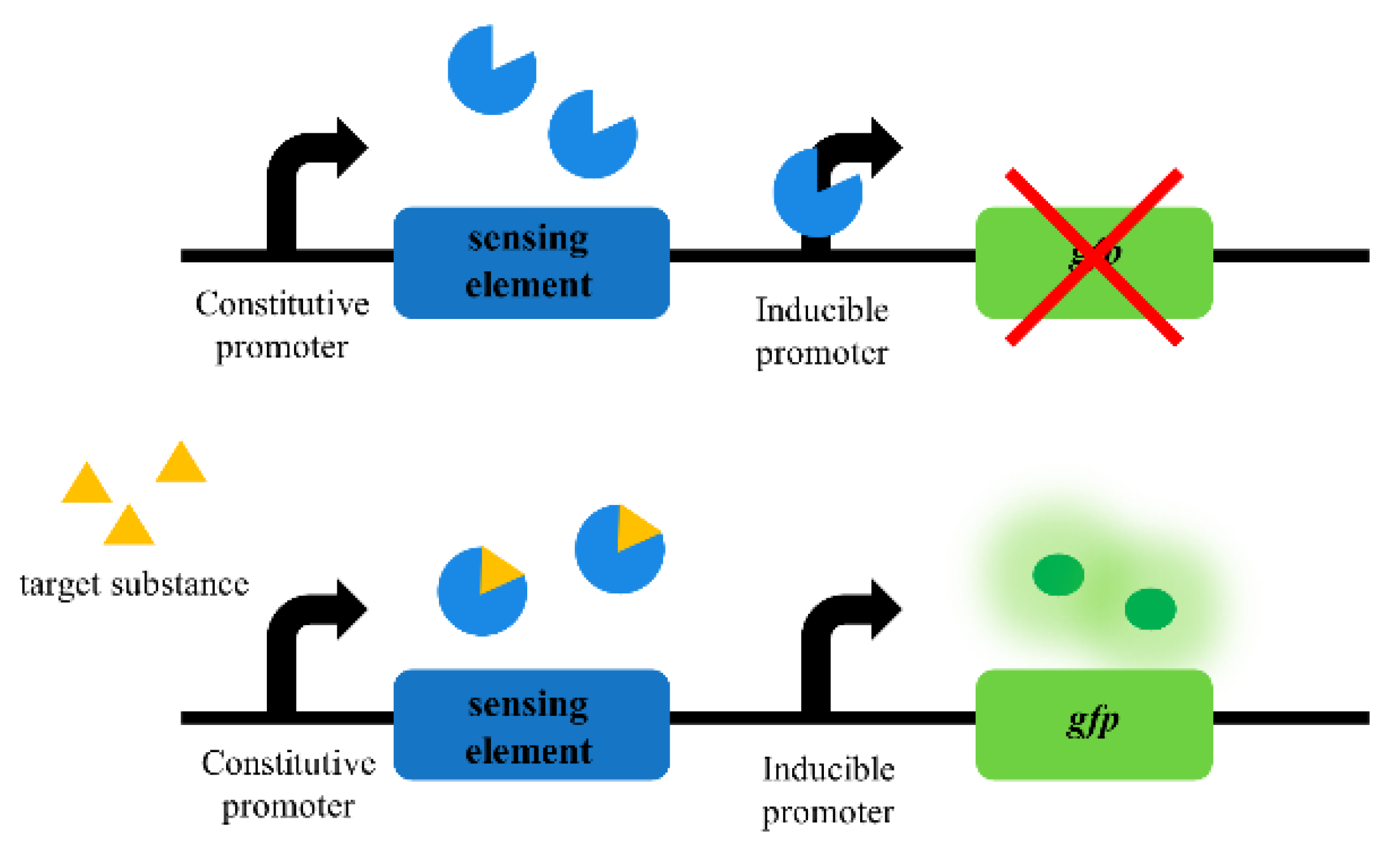
2. Sensing Elements for Whole-Cell Biosensors in Synthetic Biology
2.1. Transcription Factor
2.2. Riboswitch
3. Reporting Elements for Whole-Cell Biosensors in Synthetic Biology
3.1. Luciferase
3.2. Fluorescent Protein
3.3. Fluorescent Aptamer
3.4. Microbial Pigment Reporting Element
3.5. Gas Reporting Element
3.6. Magnetosome
3.7. Ice Nucleation Protein
3.8. Curli Protein
4. Genetic Circuits for Food Safety Detection Based on Synthetic Biology
4.1. Signal Amplification
4.1.1. Amplified Signal Based on Transcription Factor Superposition
4.1.2. Signal Amplification Based on a Positive Feedback Loop
4.1.3. Realize Signal Amplification through a Series of Signals
4.1.4. Plasmid Copy Number Riboswitch-Based Signal Amplification
4.1.5. The Signal Amplification System Is Constructed Using the Combination of Several Methods
4.2. Logical Calculation (Logic Gate)
4.2.1. AND Gate
4.2.2. OR Gate
4.2.3. NOT Gate
4.2.4. NOR Gate
4.3. Memory Element
5. Food Safety Testing System and Commercial Presentation Form Based on Synthetic Biology
5.1. Microbial Cell Sensing System
5.2. Cell-Like Sensing System
5.3. Commercial Presentation Form
6. Summary and Outlook
6.1. Challenges for Synthetic Biological Whole-Cell Biosensors
6.2. Development Direction of Synthetic Biological Whole-Cell Biosensors
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, S.; Han, D.; Peng, Y.; Zhou, H.; Ning, B.; Gao, Z. Research progress of food safety rapid detection technology. J. Food Saf. Qual. 2019, 10, 5575–5581. [Google Scholar]
- Zhao, X.; Lin, C.W.; Wang, J.; Oh, D.H. Advances in Rapid Detection Methods for Foodborne Pathogens. J. Microbiol. Biotechnol. 2014, 24, 297–312. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Tian, J.; Wu, N. Design of the Whole-cell Biosensor and its Application in Environmental Monitoring. Curr. Biotechnol. 2018, 8, 369–375. [Google Scholar]
- Zhao, G. Synthetic Biology: Unsealing the Convergence Era of Life Science Research. Bull. Chin. Acad. Sci. 2018, 33, 1135–1149. [Google Scholar]
- Rathnayake, I.V.N.; Megharaj, M.; Naidu, R. Green fluorescent protein based whole cell bacterial biosensor for the detection of bioavailable heavy metals in soil environment. Environ. Technol. Innov. 2021, 23, 101785. [Google Scholar] [CrossRef]
- Shi, S.; Choi, Y.W.; Zhao, H.; Tan, M.H.; Ang, E.L. Discovery and engineering of a 1-butanol biosensor in Saccharomyces cerevisiae. Bioresour. Technol. 2017, 245, 1343–1351. [Google Scholar] [CrossRef]
- Tao, H.-C.; Peng, Z.-W.; Li, P.-S.; Yu, T.-A.; Su, J. Optimizing cadmium and mercury specificity of CadR-based E-coli biosensors by redesign of CadR. Biotechnol. Lett. 2013, 35, 1253–1258. [Google Scholar] [CrossRef]
- Mendoza, J.I.; Soncini, F.C.; Checa, S.K. Engineering of a Au-sensor to develop a Hg-specific, sensitive and robust whole-cell biosensor for on-site water monitoring. Chem. Commun. 2020, 56, 6590–6593. [Google Scholar] [CrossRef]
- Kasey, C.M.; Zerrad, M.; Li, Y.; Cropp, T.A.; Williams, G.J. Development of Transcription Factor-Based Designer Macrolide Biosensors for Metabolic Engineering and Synthetic Biology. ACS Synth. Biol. 2018, 7, 227–239. [Google Scholar] [CrossRef]
- Chong, H.; Ching, C.B. Development of Colorimetric-Based Whole-Cell Biosensor for Organophosphorus Compounds by Engineering Transcription Regulator DmpR. ACS Synth. Biol. 2016, 5, 1290–1298. [Google Scholar] [CrossRef]
- Chang, H.-J.; Mayonove, P.; Zavala, A.; De Visch, A.; Minard, P.; Cohen-Gonsaud, M.; Bonnet, J. A Modular Receptor Platform To Expand the Sensing Repertoire of Bacteria. ACS Synth. Biol. 2018, 7, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-D.; Wu, Y.-N.; Wang, T.-M.; Zhang, C.; Xing, X.-H. Maltose Utilization as a Novel Selection Strategy for Continuous Evolution of Microbes with Enhanced Metabolite Production. ACS Synth. Biol. 2017, 6, 2326–2338. [Google Scholar] [CrossRef] [PubMed]
- Jia, X.; Ma, Y.; Bu, R.; Zhao, T.; Wu, K. Directed evolution of a transcription factor PbrR to improve lead selectivity and reduce zinc interference through dual selection. AMB Express 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Hallberg, Z.F.; Su, Y.; Kitto, R.Z.; Hammond, M.C. Engineering and In Vivo Applications of Riboswitches. In Annual Review of Biochemistry; Kornberg, R.D., Ed.; Annual Reviews: Palo Alto, CA, USA, 2017; Volume 86, pp. 515–539. [Google Scholar]
- Wang, X.; Wei, W.; Zhao, J. Using a Riboswitch Sensor to Detect Co2+/Ni2+ Transport in E. coli. Front. Chem. 2021, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.; Bronstein, L.; Diemer, J.; Koeppl, H.; Suess, B. ROC’n’Ribo: Characterizing a Riboswitching Expression System by Modeling Single-Cell Data. ACS Synth. Biol. 2017, 6, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Jang, S.; Im, D.-K.; Kang, T.J.; Oh, M.-K.; Jung, G.Y. Artificial Caprolactam-Specific Riboswitch as an Intracellular Metabolite Sensor. ACS Synth. Biol. 2019, 8, 1276–1283. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Xiu, Y.; Kang, T.J.; Lee, S.-H.; Koffas, M.A.G.; Jung, G.Y. Development of Artificial Riboswitches for Monitoring of Naringenin In Vivo. ACS Synth. Biol. 2017, 6, 2077–2085. [Google Scholar] [CrossRef]
- Xiu, Y.; Jang, S.; Jones, J.A.; Zill, N.A.; Linhardt, R.J.; Yuan, Q.; Jung, G.Y.; Koffas, M.A.G. Naringenin-responsive riboswitch-based fluorescent biosensor module for Escherichia coli co-cultures. Biotechnol. Bioeng. 2017, 114, 2235–2244. [Google Scholar] [CrossRef]
- Villa, J.K.; Su, Y.; Contreras, L.M.; Hammond, M.C. Synthetic Biology of Small RNAs and Riboswitches. Microbiol. Spectr. 2018, 6, 1. [Google Scholar]
- Gong, S.; Wang, Y.; Wang, Z.; Zhang, W. Computational Methods for Modeling Aptamers and Designing Riboswitches. Int. J. Mol. Sci. 2017, 18, 2442. [Google Scholar] [CrossRef]
- Shaner, N.C.; Steinbach, P.A.; Tsien, R.Y. A guide to choosing fluorescent proteins. Nat. Methods 2005, 2, 905–909. [Google Scholar] [CrossRef]
- Ilgu, M.; Ray, J.; Bendickson, L.; Wang, T.; Geraskin, I.M.; Kraus, G.A.; Nilsen-Hamilton, M. Light-up and FRET aptamer reporters; evaluating their applications for imaging transcription in eukaryotic cells. Methods 2016, 98, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Strack, R.L.; Svensen, N.; Jaffrey, S.R. Plug-and-Play Fluorophores Extend the Spectral Properties of Spinach. J. Am. Chem. Soc. 2014, 136, 1198–1201. [Google Scholar] [CrossRef]
- Dolgosheina, E.V.; Jeng, S.C.Y.; Panchapakesan, S.S.S.; Cojocaru, R.; Chen, P.S.K.; Wilson, P.D.; Hawkins, N.; Wiggins, P.A.; Unrau, P.J. RNA Mango Aptamer-Fluorophore: A Bright, High-Affinity Complex for RNA Labeling and Tracking. ACS Chem. Biol. 2014, 9, 2412–2420. [Google Scholar] [CrossRef]
- Filonov, G.S.; Moon, J.D.; Svensen, N.; Jaffrey, S.R. Broccoli: Rapid Selection of an RNA Mimic of Green Fluorescent Protein by Fluorescence-Based Selection and Directed Evolution. J. Am. Chem. Soc. 2014, 136, 16299–16308. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Yoshioka, D.; Inoue, K.; Takaichi, S.; Maeda, I. Evaluation of colors in green mutants isolated from purple bacteria as a host for colorimetric whole-cell biosensors. Appl. Microbiol. Biotechnol. 2007, 76, 1043–1050. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Munoz, V.H.; Cabrera-Valladares, N.; Bolivar, F.; Gosset, G.; Martinez, A. Optimum melanin production using recombinant Escherichia coli. J. Appl. Microbiol. 2006, 101, 1002–1008. [Google Scholar] [CrossRef]
- Poulter, S.; Carlton, T.M.; Su, X.; Spring, D.R.; Salmond, G.P.C. Engineering of new prodigiosin-based biosensors of Serratia for facile detection of short-chain N-acyl homoserine lactone quorum-sensing molecules. Environ. Microbiol. Rep. 2010, 2, 322–328. [Google Scholar] [CrossRef]
- Mueller, M.; Auslaender, S.; Auslaender, D.; Kemmer, C.; Fussenegger, M. A novel reporter system for bacterial and mammalian cells based on the non-ribosomal peptide indigoidine. Metab. Eng. 2012, 14, 325–335. [Google Scholar] [CrossRef]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef]
- Fulk, E.M.; Gao, X.; Lu, L.C.; Redeker, K.R.; Masiello, C.A.; Silberg, J.J. Nondestructive Chemical Sensing within Bulk Soil Using 1000 Biosensors Per Gram of Matrix. ACS Synth. Biol. 2022, 11, 2372–2383. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, M.; Du, R.; Chi, J.; He, X.; Xie, Z.; Huang, K.; Luo, Y.; Xu, W. A gas reporting whole-cell microbial biosensor system for rapid on-site detection of mercury contamination in soils. Biosens. Bioelectron. 2020, 170, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.-Y.; Masiello, C.A.; Del Valle, I.; Gao, X.; Bennett, G.N.; Silberg, J.J. Ratiometric Gas Reporting: A Nondisruptive Approach To Monitor Gene Expression in Soils. ACS Synth. Biol. 2018, 7, 903–911. [Google Scholar] [CrossRef]
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef]
- Kolinko, I.; Lohsse, A.; Borg, S.; Raschdorf, O.; Jogler, C.; Tu, Q.; Posfai, M.; Tompa, E.; Plitzko, J.M.; Brachmann, A.; et al. Biosynthesis of magnetic nanostructures in a foreign organism by transfer of bacterial magnetosome gene clusters. Nat. Nanotechnol. 2014, 9, 193–197. [Google Scholar] [CrossRef]
- Yan, L.; Da, H.; Zhang, S.; López, V.M.; Wang, W. Bacterial magnetosome and its potential application. Microbiol. Res. 2017, 203, 19–28. [Google Scholar] [CrossRef]
- Himmelreich, U.; Dresselaers, T. Cell labeling and tracking for experimental models using Magnetic Resonance Imaging. Methods 2009, 48, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.H.; Arias, J.L.; Nicolas, J.; Couvreur, P. Magnetic Nanoparticles: Design and Characterization, Toxicity and Biocompatibility, Pharmaceutical and Biomedical Applications. Chem. Rev. 2012, 112, 5818–5878. [Google Scholar] [CrossRef]
- Alphandéry, E.; Guyot, F.; Chebbi, I. Preparation of chains of magnetosomes, isolated from Magnetospirillum magneticum strain AMB-1 magnetotactic bacteria, yielding efficient treatment of tumors using magnetic hyperthermia. Int. J. Pharm. 2012, 434, 444–452. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, Y.; Ding, X.; Liu, Y.; Shen, F.; Zhang, X.; Deng, S.; Xiao, H.; Yang, G.; Peng, H. Magnetotactic bacteria: Promising biosorbents for heavy metals. Appl. Microbiol. Biotechnol. 2012, 95, 1097–1104. [Google Scholar] [CrossRef]
- Loper, J.E.; Henkels, M.D. Utilization of heterologous siderophores enhances levels of iron available to Pseudomonas putida in the rhizosphere. Appl. Environ. Microbiol. 1999, 65, 5357–5363. [Google Scholar] [CrossRef]
- Tay, P.K.R.; Nguyen, P.Q.; Joshi, N.S. A Synthetic Circuit for Mercury Bioremediation Using Self Assembling Functional Amyloids. ACS Synth. Biol. 2017, 6, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Karig, D.; Weiss, R. Signal-amplifying genetic enables in vivo observation circuit of weak promoter activation in the RhI quorum sensing system. Biotechnol. Bioeng. 2005, 89, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Sayut, D.J.; Niu, Y.; Sun, L. Construction and engineering of positive feedback loops. ACS Chem. Biol. 2006, 1, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Nistala, G.J.; Wu, K.; Rao, C.V.; Bhalerao, K.D. A modular positive feedback-based gene amplifier. J. Biol. Eng. 2010, 4, 4. [Google Scholar] [CrossRef]
- Goodson, M.S.; Bennett, A.C.; Jennewine, B.R.; Briskin, E.; Harbaugh, S.V.; Kelley-Loughnane, N. Amplifying Riboswitch Signal Output Using Cellular Wiring. ACS Synth. Biol. 2017, 6, 1440–1444. [Google Scholar] [CrossRef]
- Jang, S.; Jang, S.; Noh, M.H.; Lim, H.G.; Jung, G.Y. Novel Hybrid Input Part Using Riboswitch and Transcriptional Repressor for Signal Inverting Amplifier. ACS Synth. Biol. 2018, 7, 2199–2204. [Google Scholar] [CrossRef]
- Dwidar, M.; Yokobayashi, Y. Riboswitch Signal Amplification by Controlling Plasmid Copy Number. ACS Synth. Biol. 2019, 8, 245–250. [Google Scholar] [CrossRef]
- Kim, S.K.; Kim, S.H.; Subhadra, B.; Woo, S.-G.; Rha, E.; Kim, S.-W.; Kim, H.; Lee, D.-H.; Lee, S.-G. A Genetically Encoded Biosensor for Monitoring Isoprene Production in Engineered Escherichia coli. ACS Synth. Biol. 2018, 7, 2379–2390. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.; Volpetti, F.; Petrova, E.; French, C.; Maerkl, S.J.; Wang, B. Cascaded amplifying circuits enable ultrasensitive cellular sensors for toxic metals. Nat. Chem. Biol. 2019, 15, 540–548. [Google Scholar] [CrossRef]
- Wang, B.J.; Barahona, M.; Buck, M. A modular cell-based biosensor using engineered genetic logic circuits to detect and integrate multiple environmental signals. Biosens. Bioelectron. 2013, 40, 368–376. [Google Scholar] [CrossRef]
- Ma, D.L.; He, H.Z.; Ma, V.P.; Chan, D.S.; Leung, K.H.; Zhong, H.J.; Lu, L.; Mergny, J.L.; Leung, C.H. Label-free sensing of pH and silver nanoparticles using an “OR” logic gate. Anal Chim Acta 2012, 733, 78–83. [Google Scholar] [CrossRef]
- Green, A.A.; Kim, J.; Ma, D.; Ilver, P.A.S.; Collins, J.J.; Yin, P. Complex cellular logic computation using ribocomputing devices. Nature 2017, 548, 117–121. [Google Scholar] [CrossRef]
- Chuang, M.C.; Windmiller, J.R.; Santhosh, P.; Valdes Ramirez, G.; Katz, E.; Wang, J. High-fidelity determination of security threats via a Boolean biocatalytic cascade. Chem. Commun. 2011, 47, 3087–3089. [Google Scholar] [CrossRef]
- Kotula, J.W.; Kerns, S.J.; Shaket, L.A.; Siraj, L.; Collins, J.J.; Way, J.C.; Silver, P.A. Programmable bacteria detect and record an environmental signal in the mammalian gut. Proc. Natl. Acad. Sci. USA 2014, 111, 4838–4843. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Nielsen, A.A.K.; Fernandez-Rodriguez, J.; McClune, C.J.; Laub, M.T.; Lu, T.K.; Voigtwill, C.A. Permanent genetic memory with > 1-byte capacity. Nat. Methods 2014, 11, 1261–1266. [Google Scholar] [CrossRef]
- Vickers, C.E. The minimal genome comes of age. Nat. Biotechnol. 2016, 34, 623–624. [Google Scholar] [CrossRef]
- DeAngelis, K.M.; Firestone, M.K.; Lindow, S.E. Sensitive whole-cell biosensor suitable for detecting a variety of N-acyl homoserine lactones in intact rhizosphere microbial communities. Appl. Environ. Microbiol. 2007, 73, 3724–3727. [Google Scholar] [CrossRef]
- Liu, P.; Huang, Q.; Chen, W. Construction and application of a zinc-specific biosensor for assessing the immobilization and bioavailability of zinc in different soils. Environ. Pollut. 2012, 164, 66–72. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, K.M.; Ji, P.S.; Firestone, M.K.; Lindow, S.E. Two novel bacterial biosensors for detection of nitrate availability in the rhizosphere. Appl. Environ. Microbiol. 2005, 71, 8537–8547. [Google Scholar] [CrossRef] [PubMed]
- Hay, A.G.; Rice, J.F.; Applegate, B.M.; Bright, N.G.; Sayler, G.S. A bioluminescent whole-cell reporter for detection of 2,4-dichlorophenoxyacetic acid and 2,4-dichlorophenol in soil. Appl. Environ. Microbiol. 2000, 66, 4589–4594. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Zhang, B.; Liu, Y.-M.; Yang, K.-Q.; Liu, S.-J. New Intracellular Shikimic Acid Biosensor for Monitoring Shikimate Synthesis in Corynebacterium glutamicum. ACS Synth. Biol. 2018, 7, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.; Lv, X.; Wu, Y.; Li, J.; Du, G.; Ledesma-Amaro, R.; Liu, L. Engineering a Bifunctional Phr60-Rap60-Spo0A Quorum-Sensing Molecular Switch for Dynamic Fine-Tuning of Menaquinone-7 Synthesis in Bacillus subtilis. ACS Synth. Biol. 2019, 8, 1826–1837. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Cress, B.F.; Yang, Z.; Hordines, J.C., III; Zhao, S.; Jung, G.Y.; Wang, Z.; Koffas, M.A.G. Design and Characterization of Biosensors for the Screening of Modular Assembled Naringenin Biosynthetic Library in Saccharomyces cerevisiae. ACS Synth. Biol. 2019, 8, 2121–2130. [Google Scholar] [CrossRef]
- Beabout, K.; Bernhards, C.B.; Thakur, M.; Turner, K.B.; Cole, S.D.; Walper, S.A.; Chavez, J.L.; Lux, M.W. Optimization of Heavy Metal Sensors Based on Transcription Factors and Cell-Free Expression Systems. ACS Synth. Biol. 2021, 10, 3040–3054. [Google Scholar] [CrossRef] [PubMed]
- Pardee, K.; Green, A.A.; Takahashi, M.K.; Braff, D.; Lambert, G.; Lee, J.W.; Ferrante, T.; Ma, D.; Donghia, N.; Fan, M.; et al. Rapid, Low-Cost Detection of Zika Virus Using Programmable Biomolecular Components. Cell 2016, 165, 1255–1266. [Google Scholar] [CrossRef]
- Guo, M.; Wang, J.; Du, R.; Liu, Y.; Chi, J.; He, X.; Huang, K.; Luo, Y.; Xu, W. A test strip platform based on a whole-cell microbial biosensor for simultaneous on-site detection of total inorganic mercury pollutants in cosmetics without the need for predigestion. Biosens. Bioelectron. 2020, 150, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, P.Q.; Soenksen, L.R.; Donghia, N.M.; Angenent-Mari, N.M.; de Puig, H.; Huang, A.; Lee, R.; Slomovic, S.; Galbersanini, T.; Lansberry, G.; et al. Wearable materials with embedded synthetic biology sensors for biomolecule detection. Nat. Biotechnol. 2021, 39, 1366–1374. [Google Scholar] [CrossRef]
- Edwardson, T.G.W.; Mori, T.; Hilvert, D. Rational Engineering of a Designed Protein Cage for siRNA Delivery. J. Am. Chem. Soc. 2018, 140, 10439–10442. [Google Scholar] [CrossRef] [PubMed]
- Hao, N.; Lu, J.W.; Zhou, Z.; Hua, R.; Wang, K. A pH-Resolved Colorimetric Biosensor for Simultaneous Multiple Target Detection. ACS Sens. 2018, 3, 2159–2165. [Google Scholar] [CrossRef]
- Berepiki, A.; Kent, R.; Machado, L.F.M.; Dixon, N. Development of High-Performance Whole Cell Biosensors Aided by Statistical Modeling. ACS Synth. Biol. 2020, 9, 576–589. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reporting Element Type | Classification | Output Signal | Substrate Requirement | Testing System Requirement | Signal Maintenance Time | Signal Visualization |
|---|---|---|---|---|---|---|
| Luciferase | Bacterial luciferase | fluorescence signal | endogenous/exogenous | transparent | real-time signal | no |
| Firefly luciferase | ||||||
| Aequorin | ||||||
| Fluorescent protein | Dark blue | fluorescence signal | no substrate required | transparent | real-time signal | strong signal visual |
| Blue | ||||||
| Cyan | ||||||
| Green | ||||||
| Yellow | ||||||
| Orange | ||||||
| Red | ||||||
| Dark red | ||||||
| Near infrared | ||||||
| Fluorescent aptamer | Malachite green aptamer | fluorescence signal | exogenous | transparent | real-time signal | no |
| Spinach aptamer | ||||||
| Mango aptamer | ||||||
| Broccoli aptamer | ||||||
| Microbial pigment reporting element | Carotenoid | pigment signal | endogenous/exogenous | transparent | cumulative signal | visual |
| Violacein | ||||||
| Melanin | ||||||
| Prodiginine | ||||||
| Indigoidine | ||||||
| Betaxanthin | ||||||
| Gas reporting element | N2O | gas signal | endogenous/exogenous | no gas adsorption | real-time signal | no |
| CH3Cl, CH3Br, CH3I | ||||||
| CH3SH | ||||||
| C2H4 | ||||||
| CH3CHO | ||||||
| Magnetic reporting element | Magnetosome | magnetic signal | endogenous | no | real-time signal | no |
| MagA | ||||||
| Metallothionein and Phytochelatin synthase | ||||||
| Ferritin | ||||||
| Other reporting element | Ice nucleation protein | ice nucleation protein | no substrate required | no | cumulative signal | no |
| Curli protein | bacteriophage sedimentation | no substrate required | transparent, liquid | cumulative signal | visual |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.; Chen, X.; Su, H.; Guo, M.; Liu, H. Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety. Int. J. Mol. Sci. 2023, 24, 7989. https://doi.org/10.3390/ijms24097989
Chen S, Chen X, Su H, Guo M, Liu H. Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety. International Journal of Molecular Sciences. 2023; 24(9):7989. https://doi.org/10.3390/ijms24097989
Chicago/Turabian StyleChen, Shijing, Xiaolin Chen, Hongfei Su, Mingzhang Guo, and Huilin Liu. 2023. "Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety" International Journal of Molecular Sciences 24, no. 9: 7989. https://doi.org/10.3390/ijms24097989
APA StyleChen, S., Chen, X., Su, H., Guo, M., & Liu, H. (2023). Advances in Synthetic-Biology-Based Whole-Cell Biosensors: Principles, Genetic Modules, and Applications in Food Safety. International Journal of Molecular Sciences, 24(9), 7989. https://doi.org/10.3390/ijms24097989