

Effects of Myostatin on Nuclear Morphology at the Myotendinous Junction

 ,
,
Abstract
1. Introduction
2. Results
2.1. Regeneration Processes on the Tendon Side of the MTJ
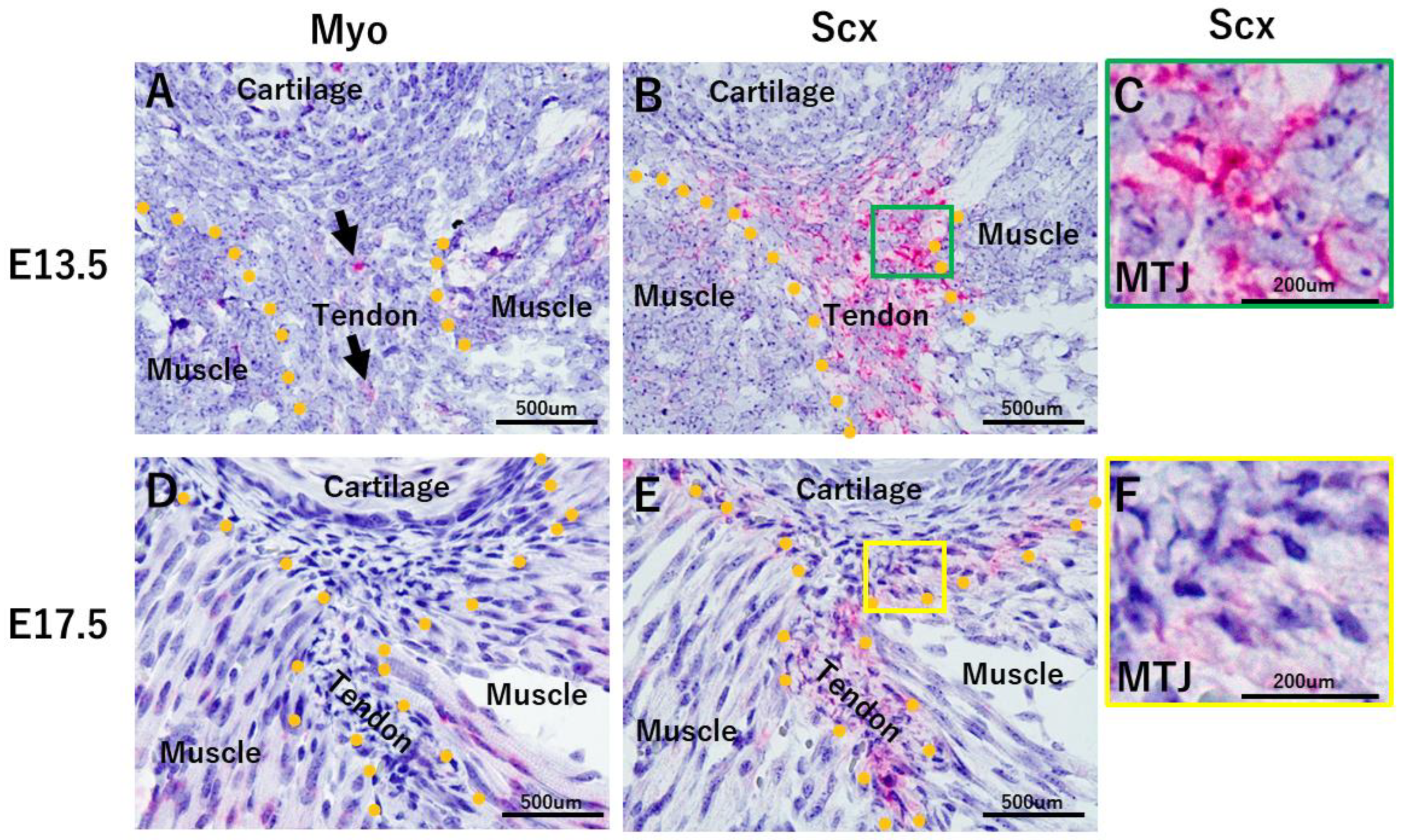
2.2. Developmental Processes on the Tendon Side of the MTJ
2.3. Preparation of the Pseudo MTJ model (Part 1)
2.4. Preparation of a Pseudo MTJ Model (Part 2)
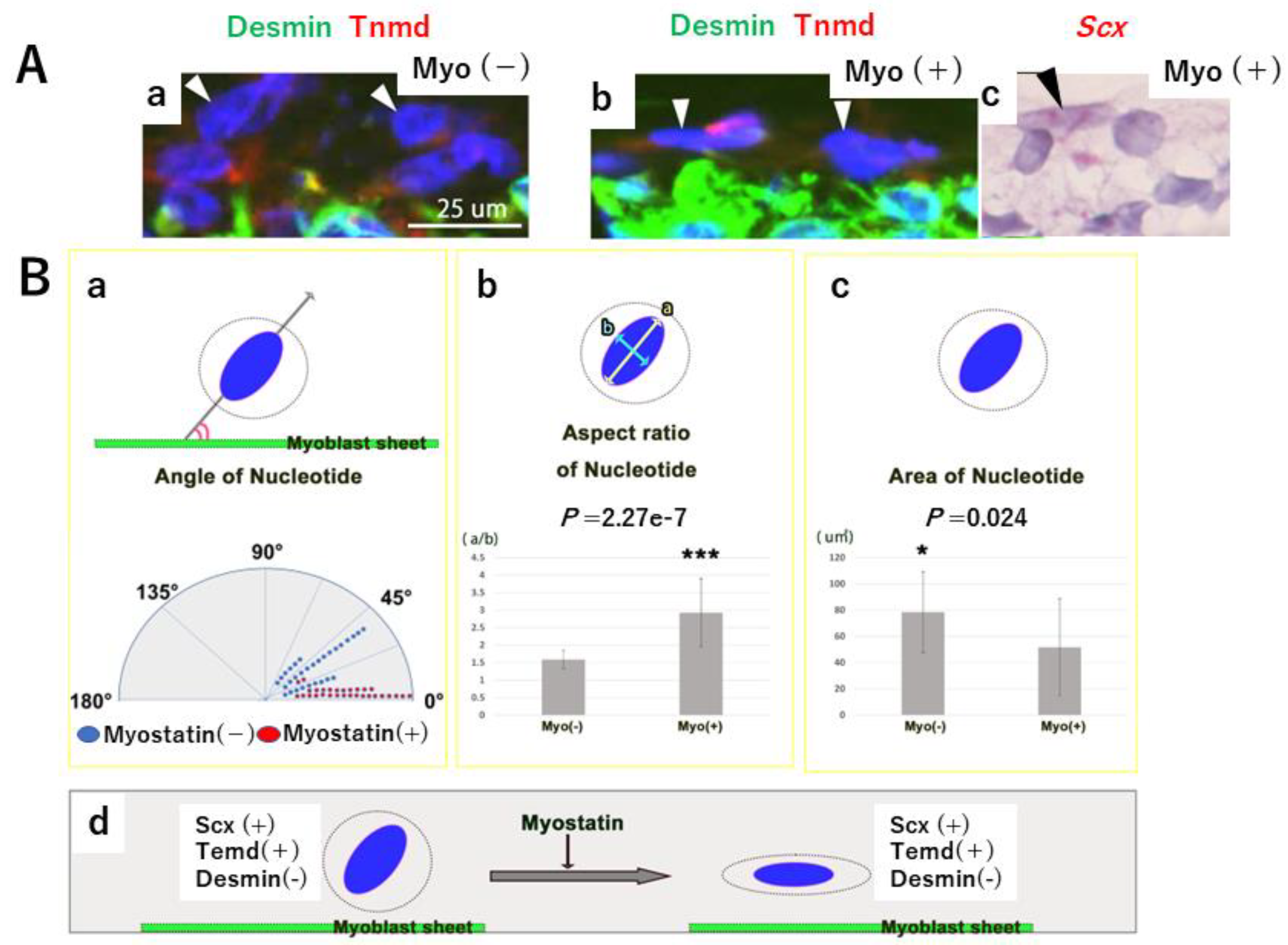
2.5. Effects of Myostatin Addition on the Pseudo MTJ Model
3. Discussion
4. Materials and Methods
4.1. Achilles Tendon Injury Model
4.2. Preparation of the Mouse Myoblast, Mesenchymal Cell Line
4.3. Preparation of Cell Sheets and Culture Conditions for the Pseudo MTJ Model
- (1)
- Myoblast sheet only (as a control) (C2C12);
- (2)
- A layer of collagen gel (CellmatrixⓇ; Nitta-gelatin Co., Osaka, Japan) on top of myoblasts (C2C12-Gel);
- (3)
- A well-stirred mixture of collagen gel and NIH3T3 cells overlaid on myoblasts (C2C12-Gel 3T3),
4.4. Histochemical Analysis (In Vitro)
4.5. In Situ Hybridization
4.6. Reverse Transcription-Polymerase Chain Reaction
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charvet, B.; Ruggiero, F.; Le Guellec, D. The development of the myotendinous junction. A review. Muscles Ligaments Tendons J. 2012, 2, 53–63. [Google Scholar]
- Mohassel, P.; Foley, A.R.; Bönnemann, C.G. Extracellular matrix-driven congenital muscular dystrophies. Matrix Biol. 2018, 71–72, 188–204. [Google Scholar] [CrossRef]
- Petrany, M.J.; Swoboda, C.O.; Sun, C.; Chetal, K.; Chen, X.; Weirauch, M.T.; Salomonis, N.; Millay, D.P. Single-nucleus RNA-seq identifies transcriptional heterogeneity in multinucleated skeletal myofibers. Nat. Commun. 2020, 11, 6374. [Google Scholar] [CrossRef]
- Jakobsen, J.R.; Krogsgaard, M.R. The myotendinous junction-A vulnerable companion in sports. A narrative review. Front. Physiol. 2021, 12, 635561. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, R.; Yamamoto, M.; Jeong, J.; Hinata, N.; Katori, Y.; Chang, W.-J.; Abe, S. Switching of Sox9 expression during musculoskeletal system development. Sci. Rep. 2020, 10, 8425. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Rhee, S.-K.; Osonoi, M.; Nakamura, T.; Cho, B.H.; Murakami, G.; Ide, Y. Expression of intermediate filaments at muscle insertions in human fetuses. J. Anat. 2010, 217, 167–173. [Google Scholar] [CrossRef]
- Yamamoto, M.; Shinomiya, T.; Kishi, A.; Yamane, S.; Umezawa, T.; Ide, Y.; Abe, S. Desmin and nerve terminal expression during embryonic development of the lateral pterygoid muscle in mice. Arch. Oral Biol. 2014, 59, 871–879. [Google Scholar] [CrossRef]
- Edom-Vovard, F.; Schuler, B.; Bonnin, M.-A.; Teillet, M.-A.; Duprez, D. Fgf4 Positively Regulates scleraxis and Tenascin Expression in Chick Limb Tendons. Dev. Biol. 2002, 247, 351–366. [Google Scholar] [CrossRef] [PubMed]
- De Lima, J.E.; Blavet, C.; Bonnin, M.-A.; Hirsinger, E.; Comai, G.; Yvernogeau, L.; Delfini, M.-C.; Bellenger, L.; Mella, S.; Nassari, S.; et al. Unexpected contribution of fibroblasts to muscle lineage as a mechanism for limb muscle patterning. Nat. Commun. 2021, 12, 3851. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, W.; Kraft-Sheleg, O.; Zaffryar-Eilot, S.; Melamed, S.; Sun, C.; Millay, D.P.; Hasson, P. Fibroblast fusion to the muscle fiber regulates myotendinous junction formation. Nat. Commun. 2021, 12, 3852. [Google Scholar] [CrossRef]
- Yan, R.; Zhang, H.; Ma, Y.; Lin, R.; Zhou, B.; Zhang, T.; Fan, C.; Zhang, Y.; Wang, Z.; Fang, T.; et al. Discovery of Muscle-Tendon Progenitor Subpopulation in Human Myotendinous Junction at Single-Cell Resolution. Research 2022, 2022, 9760390. [Google Scholar] [CrossRef]
- McPherron, A.C.; Lawler, A.M.; Lee, S.J. Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member. Nature 1997, 387, 83–90. [Google Scholar] [CrossRef]
- McCroskery, S.; Thomas, M.; Maxwell, L.; Sharma, M.; Kambadur, R. Myostatin negatively regulates satellite cell activation and self-renewal. J. Cell Biol. 2003, 162, 1135–1147. [Google Scholar] [CrossRef]
- Mendias, C.L.; Bakhurin, K.I.; Faulkner, J.A. Tendons of myostatin-deficient mice are small, brittle, and hypocellular. Proc. Natl. Acad. Sci. USA 2008, 105, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Mendias, C.L.; Lynch, E.B.; Gumucio, J.P.; Flood, M.D.; Rittman, D.S.; Van Pelt, D.W.; Roche, S.M.; Davis, C.S. Changes in skeletal muscle and tendon structure and function following genetic inactivation of myostatin in rats. J. Physiol. 2015, 593, 2037–2052. [Google Scholar] [CrossRef] [PubMed]
- Yaffe, D.; Saxel, O. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature 1977, 270, 725–727. [Google Scholar] [CrossRef] [PubMed]
- Kokabu, S.; Nakatomi, C.; Matsubara, T.; Ono, Y.; Addison, W.N.; Lowery, J.W.; Urata, M.; Hudnall, A.M.; Hitomi, S.; Nakatomi, M.; et al. The transcriptional co-repressor TLE3 regulates myogenic differentiation by repressing the activity of the MyoD transcription factor. J. Biol. Chem. 2017, 292, 12885–12894. [Google Scholar] [CrossRef]
- Katagiri, T.; Yamaguchi, A.; Komaki, M.; Abe, E.; Takahashi, N.; Ikeda, T.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic rrotein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef]
- Teboul, L.; Gaillard, D.; Staccini, L.; Inadera, H.; Amri, E.Z.; Gri Ker, E.D.F.; Chu, B.; Phillippi, J.A.; Gharaibeh, B.; Huard, J.; et al. Engineering spatial control of multiple differentiation fates within a stem cell population. Bio-materials 2011, 32, 3413–3422. [Google Scholar]
- Uemura, K.; Hayashi, M.; Itsubo, T.; Oishi, A.; Iwakawa, H.; Komatsu, M.; Uchiyama, S.; Kato, H. Myostatin promotes tenogenic differentiation of C2C12 myoblast cells through Smad3. FEBS Open Bio 2017, 7, 522–532. [Google Scholar] [CrossRef]
- Todaro, G.J.; Green, H. Quantitative Studies of the Growth of Mouse Embryo Cells in Culture and Their Development into Established Lines. J. Cell Biol. 1963, 17, 299–313. [Google Scholar] [CrossRef]
- Fu, B.; Quintero, J.; Baker, C.C. Keratinocyte growth conditions modulate telomerase expression, senescence, and immor-talization by human papillomavirus type 16 E6 and E7 oncogenes. Cancer Res. 2003, 63, 7815–7824. [Google Scholar] [PubMed]
- Barreca, A.; de Luca, M.; Del Monte, P.; Bondanza, S.; Damonte, G.; Cariola, G.; di Marco, E.; Giordano, G.; Cancedda, R.; Minuto, F. In vitro paracrine regulation of human keratinocyte growth by fibroblast-derived insulin-like growth factors. J. Cell. Physiol. 1992, 151, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Masson, G.B.; Fugère, C.; Leclerc, S.; Lefort, N.R.; Germain, L.; Guérin, S.L. Influence of a feeder layer in cell culture: Modulation of the transcription factors Sp1 and Sp3 by irradiated 3T3 cells. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3838. [Google Scholar]
- Lu, H.-F.; Chua, K.-N.; Zhang, P.-C.; Lim, W.-S.; Ramakrishna, S.; Leong, K.W.; Mao, H.-Q. Three-dimensional co-culture of rat hepatocyte spheroids and NIH/3T3 fibroblasts enhances hepatocyte functional maintenance. Acta Biomater. 2005, 1, 399–410. [Google Scholar] [CrossRef]
- Trevino, C.; Anderson, R.; Muneoka, K. 3T3 Cell Integration and Differentiative Potential during Limb Development in the Mouse. Dev. Biol. 1993, 155, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Koeck, K.S.; Salehi, S.; Humenik, M.; Scheibel, T. Processing of Continuous Non-Crosslinked Collagen Fibers for Microtissue Formation at the Muscle-Tendon Interface. Adv. Funct. Mater. 2021, 32, 21122. [Google Scholar] [CrossRef]
- Yamane, S.; Higa, K.; Umezawa, T.; Serikawa, M.; Shimazaki, J.; Abe, S. Engineered three-dimensional rabbit oral epitheli-al-mesenchymal-muscular hybrid sheets. Int. J. Oral. Sci. 2016, 8, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, T.; Higa, K.; Serikawa, M.; Yamamoto, M.; Matsunaga, S.; Shimazaki, J.; Abe, S. Proliferative activity of skeletal myoblast sheet by paracrine effects of mesenchymal stem cells. J. Oral Biosci. 2016, 58, 158–166. [Google Scholar] [CrossRef]
- Manceau, M.; Gros, J.; Savage, K.; Thomé, V.; McPherron, A.; Paterson, B.; Marcelle, C. Myostatin promotes the terminal differentiation of embryonic muscle progenitors. Genes Dev. 2008, 22, 668–681. [Google Scholar] [CrossRef]
- Chen, B.; Co, C.; Ho, C.-C. Cell shape dependent regulation of nuclear morphology. Biomaterials 2015, 67, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Venturini, V.; Pezzano, F.; Castro, F.C.; Häkkinen, H.-M.; Jiménez-Delgado, S.; Colomer-Rosell, M.; Marro, M.; Tolosa-Ramon, Q.; Paz-López, S.; Valverde, M.A.; et al. The nucleus measures shape changes for cellular proprioception to control dynamic cell behavior. Science 2020, 370, eaba2644. [Google Scholar] [CrossRef] [PubMed]
- Seo, K.; Suzuki, T.; Kobayashi, K.; Nishimura, T. Adipocytes suppress diferentiation of muscle cells in a co-culture system. Anim. Sci. J. 2019, 90, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Shima, A.; Itou, A.; Takeuchi, S. Cell fibers promote proliferation of co-cultured cells on a dish. Sci. Rep. 2020, 10, 288. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Takada, H.; Ishizuka, S.; Kitamura, K.; Jeong, J.; Sato, M.; Hinata, N.; Abe, S. Morphological association between the muscles and bones in the craniofacial region. PLoS ONE 2020, 15, e0227301. [Google Scholar] [CrossRef]
- Yamamoto, M.; Abe, S. Mechanism of muscle-tendon-bone complex development in the head. Anat. Sci. Int. 2020, 95, 165–173. [Google Scholar] [CrossRef]
- Abe, S.; Yamamoto, M. Factors Involved in Morphogenesis in the Muscle–Tendon–Bone Complex. Int. J. Mol. Sci. 2021, 22, 6365. [Google Scholar] [CrossRef]
- Ishizuka, S.; Yamamoto, M.; Hirouchi, H.; Yotsuya, M.; Ohkubo, M.; Sato, M.; Abe, S. Muscle-bone relationship in temporo-mandibular joint disorders after partial discectomy. J. Oral. Biosci. 2021, 63, 436–443. [Google Scholar] [CrossRef]
- Yamamoto, M.; Sakiyama, K.; Kitamura, K.; Yamamoto, Y.; Takagi, T.; Sekiya, S.; Watanabe, G.; Taniguchi, S.; Ogawa, Y.; Ishizuka, S.; et al. Development and Regeneration of Muscle, Tendon, and Myotendinous Junctions in Striated Skeletal Muscle. Int. J. Mol. Sci. 2022, 23, 3006. [Google Scholar] [CrossRef]
- Kanehira, C.; Yamamoto, M.; Hirouchi, H.; Ishuzuka, S.; Sakiyama, K.; Higa, K.; Murakami, G.; Abe, S. Tendinous annulus of zinn for a common origin of the extraocular rectus muscles: A histological study of the orbital apex from donated elderly cadavers. Anat. Sci. Int. 2022, 97, 369–379. [Google Scholar] [CrossRef]
- Takagi, T.; Yamamoto, M.; Sugano, A.; Kanehira, C.; Kitamura, K.; Katayama, M.; Sakai, K.; Sato, M.; Abe, S. Alteration of Oral and Perioral Soft Tissue in Mice following Incisor Tooth Extraction. Int. J. Mol. Sci. 2022, 23, 2987. [Google Scholar] [CrossRef]
- Schweitzer, R.; Chyung, J.H.; Murtaugh, L.C.; Brent, A.E.; Rosen, V.; Olson, E.N.; Lassar, A.; Tabin, C.J. Analysis of the tendon cell fate using Scleraxis, a specific marker for tendons and ligaments. Development 2001, 128, 3855–3866. [Google Scholar] [CrossRef]
- Brent, A.E.; Schweitzer, R.; Tabin, C.J. A somatic compartment of tendon progenitors. Cell 2003, 113, 235–248. [Google Scholar] [CrossRef] [PubMed]
- Shukunami, C.; Takimoto, A.; Oro, M.; Hiraki, Y. Scleraxis positively regulates the expression of tenomodulin, a differentiation marker of tenocytes. Dev. Biol. 2006, 298, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Yoshimoto, Y.; Takimoto, A.; Watanabe, H.; Hiraki, Y.; Kondoh, G.; Shukunami, C. Scleraxis is required for maturation of tissue domains for proper integration of the musculoskeletal system. Sci. Rep. 2017, 7, 45010. [Google Scholar] [CrossRef]
- Berthet, E.; Chen, C.; Butcher, K.; Schneider, R.A.; Alliston, T.; Amirtharajah, M. Smad3 binds scleraxis and mohawk and regulates tendon matrix organization. J. Orthop. Res. 2013, 31, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Sakabe, T.; Sakai, K.; Maeda, T.; Sunaga, A.; Furuta, N.; Schweitzer, R.; Sasaki, T.; Sakai, T. Transcription factor scleraxis vitally contributes to progenitor lineage direction in wound healing of adult tendon in mice. J. Biol. Chem. 2018, 293, 5766–5780. [Google Scholar] [CrossRef]
- Cesario, J.M.; Almaidhan, A.A.; Jeong, J. Expression of forkhead box transcription factor genes Foxp1 and Foxp2 during jaw development. Gene Expr. Patterns 2016, 20, 111–119. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Assay ID | Amplicon Length | |
|---|---|---|---|
| Scleraxis | transcription factor for differentiated tendon cells. | Mm01205675_m1 | 59 |
| Tenomodulin | late differentiation marker gene for tendon and ligament tissue. | Mm00491594_m1 | 65 |
| Sex determining region Y-box9 | transcription factor for differentiated chondrocyte cells and tendon cells. | Mm00448840_m1 | 101 |
| Desmin | Type III intermediate filament in skeletal, smooth and cardiac muscle tissue. | Mm00802455_m1 | 92 |
| Paired Box 7 | Paired box transcription factor family member involved in maintaining proliferation and preventing differentiation in skeletal muscle progenitor cells. | Mm00482759_m1 | 102 |
| Myostatin | Negative regulated protein for muscle | Mm01254559_m1 | 90 |
| S18 | S13P family of ribosomal proteins used as an internal control. | Mm02601777_g1 | 76 |
| Primer | Sequence (5′–3′) | Product Size (bp) | |
|---|---|---|---|
| Scleraxis | transcription factor for differentiated tendon cells. | AGCCCAAACAGATCTGCACCTT | 139 |
| CTTCCACCTTCACTAGTGGCATCA | |||
| Tenomodulin | late differentiation marker gene for tendon and ligament tissue. | ATGGGTGGTCCCACAAGTGAA | 123 |
| CTCTCATCCAGCATGGGATCAA | |||
| Sex determining region Y-box9 | transcription factor for differentiated chondrocyte cells and tendon cells. | ATCTGAAGAAGGAGAGCGAG | 263 |
| TCAGAAGTCTCCAGAGCTTG | |||
| Desmin | Type III intermediate filament in skeletal, smooth and cardiac muscle tissue. | ACCAGATCCAGTCCTACACC | 202 |
| TTGAGCAGGTCCTGGTACTC | |||
| Paired Box 7 | Paired box transcription factor family member involved in maintaining proliferation and preventing differentiation in skeletal muscle progenitor cells. | ATCCGGCCCTGTGTCATCTC | 278 |
| CACGCGGCTAATCGAACTCA | |||
| Glyceraldehyde 3-phosphate dehydrogenase | Glyceraldehyde-3-phosphate dehydrogenase, internal control. | ACCACAGTCCATGCCATCAC | 452 |
| TCCACCACCCTGTTGCTGTA | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amemiya, H.; Yamamoto, M.; Higa, K.; Watanabe, G.; Taniguchi, S.; Kitamura, K.; Jeong, J.; Yanagisawa, N.; Fukuda, K.-i.; Abe, S. Effects of Myostatin on Nuclear Morphology at the Myotendinous Junction. Int. J. Mol. Sci. 2023, 24, 6634. https://doi.org/10.3390/ijms24076634
Amemiya H, Yamamoto M, Higa K, Watanabe G, Taniguchi S, Kitamura K, Jeong J, Yanagisawa N, Fukuda K-i, Abe S. Effects of Myostatin on Nuclear Morphology at the Myotendinous Junction. International Journal of Molecular Sciences. 2023; 24(7):6634. https://doi.org/10.3390/ijms24076634
Chicago/Turabian StyleAmemiya, Hikari, Masahito Yamamoto, Kazunari Higa, Genji Watanabe, Shuichiro Taniguchi, Kei Kitamura, Juhee Jeong, Nobuaki Yanagisawa, Ken-ichi Fukuda, and Shinichi Abe. 2023. "Effects of Myostatin on Nuclear Morphology at the Myotendinous Junction" International Journal of Molecular Sciences 24, no. 7: 6634. https://doi.org/10.3390/ijms24076634
APA StyleAmemiya, H., Yamamoto, M., Higa, K., Watanabe, G., Taniguchi, S., Kitamura, K., Jeong, J., Yanagisawa, N., Fukuda, K.-i., & Abe, S. (2023). Effects of Myostatin on Nuclear Morphology at the Myotendinous Junction. International Journal of Molecular Sciences, 24(7), 6634. https://doi.org/10.3390/ijms24076634