
The Circadian Clock, Nutritional Signals and Reproduction: A Close Relationship
 , , ,
, , ,  , ,
, , 
Abstract
1. Introduction
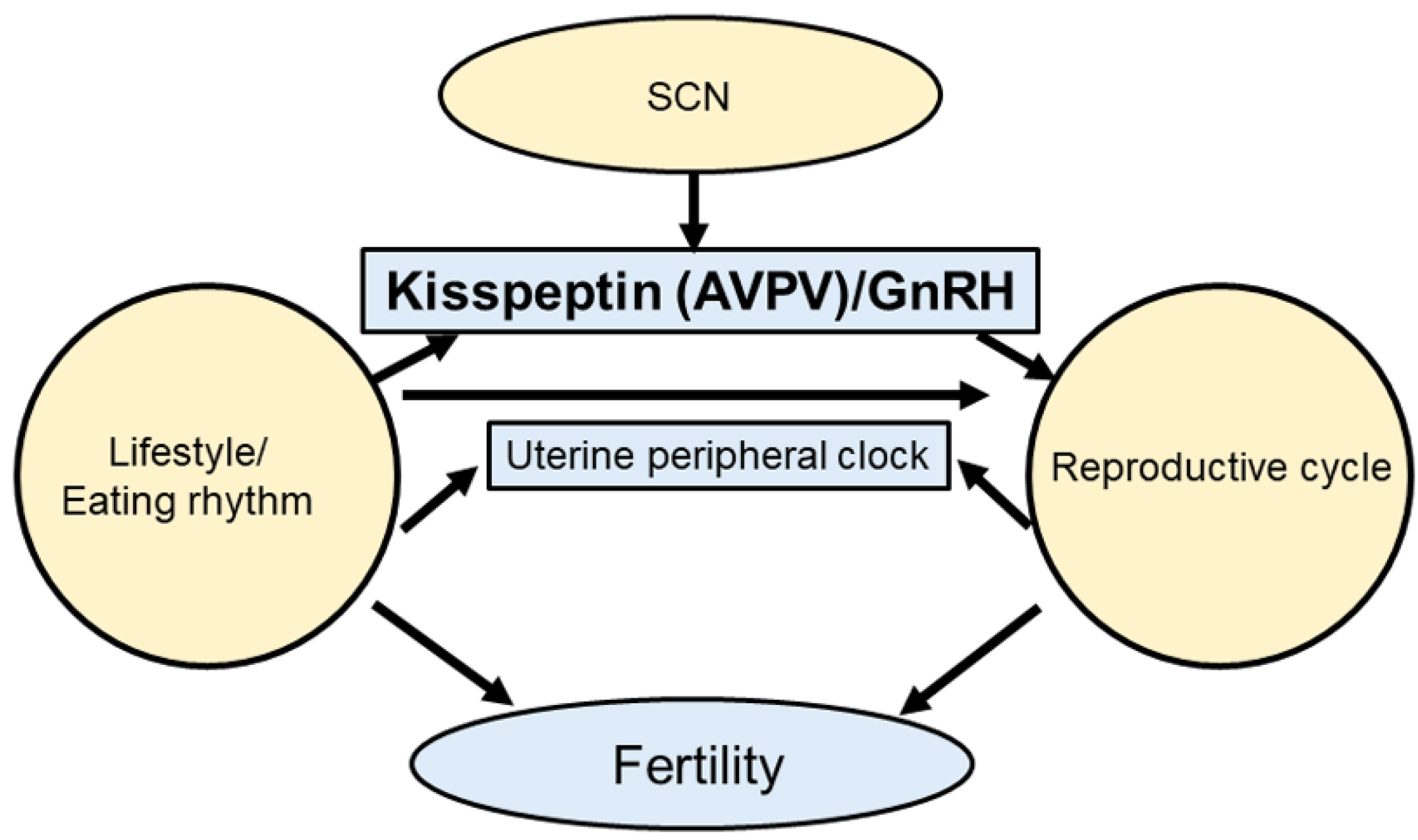
2. Effects of the Circadian Clock on the Hypothalamic–Pituitary–Gonadal (HPG) Axis and Reproduction
2.1. Circadian Clock Regulation of the HPG Axis
2.2. Ovarian Circadian Clock
2.3. Endometrial Circadian Clock
2.4. Animal Studies on the Circadian Clock and Reproductive Function

{kind=link}
{kind=link}
| Mutant Mice | Phenotypes of Reproduction | References |
|---|---|---|
| Conventional Bmal1 KO | Delayed puberty; females have longer estrous cycles; infertile | [5,76] |
| Gonadotrope Bmal1 KO | Irregular estrous cycle; fertile | [7,77] |
| Granulosa cell Bmal1 KO (GCKO) | Normal ovarian morphology and a typical estrous cycle; fertile | [7] |
| Ovarian steroidogenic cells Bmal1 KO | Typical puberty; early pregnancy loss; infertile | [73] |
| Theca cell Bmal1 KO | Fewer offsprings and increased mating failure; regular estrous cycle; subfertile | [7] |
| Uterine Bmal1 KO | Reducing placental vascularization and causing fetal mortality within the uterus; subfertile | [75] |
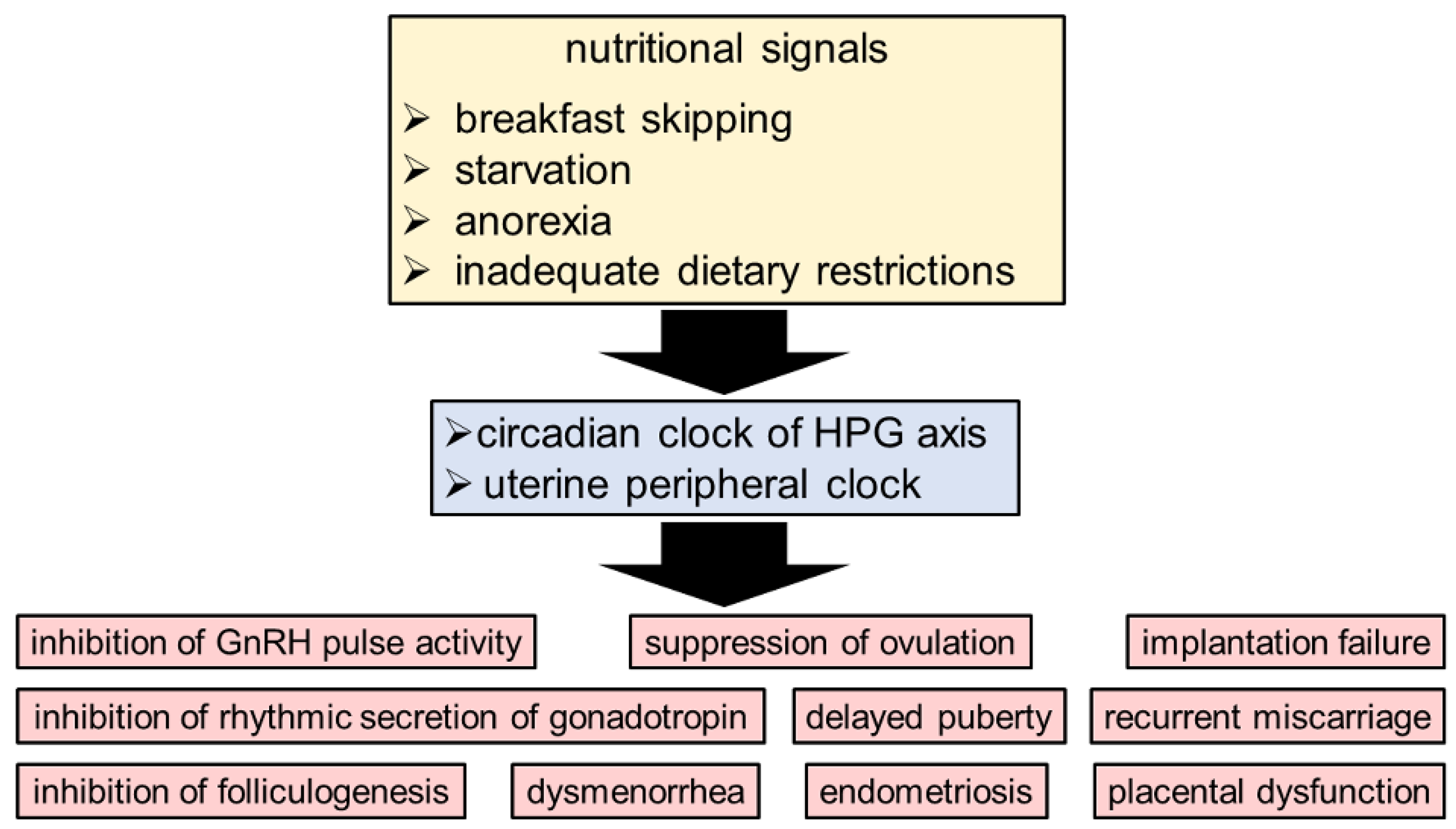
3. The Circadian Clock System as a Link between Nutritional Signals and Reproduction
4. The Circadian Clock and Puberty
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ADHOGD | adolescent dietary habit-induced obstetric and gynecologic disease |
| Bmal1 | brain and muscle Arnt-like protein-1 |
| Clock | circadian locomotor output cycles kaput |
| Cry1 | cryptochrome circadian regulator 1 |
| Cry2 | cryptochrome circadian regulator 2 |
| Dbp | albumin D-binding protein |
| FSH | follicle-stimulating hormone |
| GnRH | gonadotropin-releasing hormone |
| HDP | hypertensive disorders of pregnancy |
| HPG | hypothalamic–pituitary–gonadal |
| LH | luteinizing hormone |
| PCOS | polycystic ovary syndrome |
| Per1 | period circadian regulator 1 |
| Per2 | period circadian regulator 2 |
| Per3 | period circadian regulator 3 |
| NR1D1 | nuclear receptor subfamily 1, group D, member 1 |
References
- Mahoney, M.M. Shift work, jet lag and female reproduction. Int. J. Endocrinol. 2010, 2010, 813764. [Google Scholar] [CrossRef] [PubMed]
- Summa, K.C.; Vitaterna, M.H.; Turek, F.W. Environmental perturbation of the circadian clock disrupts pregnancy in the mouse. PLoS ONE 2012, 7, e37668. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.H.; Olson, S.L.; Turek, F.W.; Levine, J.E.; Horton, T.H.; Takahashi, J.S. Circadian clock mutation disrupts estrous cyclicity and maintenance of pregnancy. Curr. Biol. 2004, 14, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Bittman, E.L. Circadian Function in Multiple Cell Types Is Necessary for Proper Timing of the Preovulatory LH Surge. J. Biol. Rhythm. 2019, 34, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Boden, M.J.; Varcoe, T.J.; Voultsios, A.; Kennaway, D.J. Reproductive biology of female Bmal1 null mice. Reproduction 2010, 139, 1077–1090. [Google Scholar] [CrossRef] [PubMed]
- Sellix, M.T. Clocks underneath: The role of peripheral clocks in the timing of female reproductive physiology. Front. Endocrinol. 2013, 4, 91. [Google Scholar] [CrossRef]
- Mereness, A.L.; Murphy, Z.C.; Forrestel, A.C.; Butler, S.; Ko, C.; Richards, J.S.; Sellix, M.T. Conditional Deletion of Bmal1 in Ovarian Theca Cells Disrupts Ovulation in Female Mice. Endocrinology 2016, 157, 913–927. [Google Scholar] [CrossRef]
- Xu, J.; Li, Y.; Wang, Y.; Xu, Y.; Zhou, C. Loss of Bmal1 decreases oocyte fertilization, early embryo development and implantation potential in female mice. Zygote 2016, 24, 760–767. [Google Scholar] [CrossRef]
- Perez, S.; Murias, L.; Fernandez-Plaza, C.; Diaz, I.; Gonzalez, C.; Otero, J.; Diaz, E. Evidence for clock genes circadian rhythms in human full-term placenta. Syst. Biol. Reprod. Med. 2015, 61, 360–366. [Google Scholar] [CrossRef]
- Muter, J.; Lucas, E.S.; Chan, Y.W.; Brighton, P.J.; Moore, J.D.; Lacey, L.; Quenby, S.; Lam, E.W.; Brosens, J.J. The clock protein period 2 synchronizes mitotic expansion and decidual transformation of human endometrial stromal cells. FASEB J. 2015, 29, 1603–1614. [Google Scholar] [CrossRef]
- Ratajczak, C.K.; Herzog, E.D.; Muglia, L.J. Clock gene expression in gravid uterus and extra-embryonic tissues during late gestation in the mouse. Reprod. Fertil. Dev. 2010, 22, 743–750. [Google Scholar] [CrossRef]
- Kennaway, D.J.; Boden, M.J.; Varcoe, T.J. Circadian rhythms and fertility. Mol. Cell. Endocrinol. 2012, 349, 56–61. [Google Scholar] [CrossRef]
- Sen, A.; Hoffmann, H.M. Role of core circadian clock genes in hormone release and target tissue sensitivity in the reproductive axis. Mol. Cell. Endocrinol. 2020, 501, 110655. [Google Scholar] [CrossRef]
- Pan, X.; Taylor, M.J.; Cohen, E.; Hanna, N.; Mota, S. Circadian Clock, Time-Restricted Feeding and Reproduction. Int. J. Mol. Sci. 2020, 21, 831. [Google Scholar] [CrossRef]
- Karman, B.N.; Tischkau, S.A. Circadian clock gene expression in the ovary: Effects of luteinizing hormone. Biol. Reprod. 2006, 75, 624–632. [Google Scholar] [CrossRef]
- Nakamura, T.J.; Sellix, M.T.; Kudo, T.; Nakao, N.; Yoshimura, T.; Ebihara, S.; Colwell, C.S.; Block, G.D. Influence of the estrous cycle on clock gene expression in reproductive tissues: Effects of fluctuating ovarian steroid hormone levels. Steroids 2010, 75, 203–212. [Google Scholar] [CrossRef]
- Yaw, A.M.; Duong, T.V.; Nguyen, D.; Hoffmann, H.M. Circadian rhythms in the mouse reproductive axis during the estrous cycle and pregnancy. J. Neurosci. Res. 2021, 99, 294–308. [Google Scholar] [CrossRef]
- Mieda, M.; Okamoto, H.; Sakurai, T. Manipulating the Cellular Circadian Period of Arginine Vasopressin Neurons Alters the Behavioral Circadian Period. Curr. Biol. 2016, 26, 2535–2542. [Google Scholar] [CrossRef]
- Sun, W.; Li, S.X.; Wang, G.; Dong, S.; Jiang, Y.; Spruyt, K.; Ling, J.; Zhu, Q.; Lee, T.M.; Jiang, F. Association of Sleep and Circadian Activity Rhythm with Emotional Face Processing among 12-month-old Infants. Sci. Rep. 2018, 8, 3200. [Google Scholar] [CrossRef]
- Ikeda, Y.; Kumagai, H.; Skach, A.; Sato, M.; Yanagisawa, M. Modulation of circadian glucocorticoid oscillation via adrenal opioid-CXCR7 signaling alters emotional behavior. Cell 2013, 155, 1323–1336. [Google Scholar] [CrossRef]
- Page, A.J.; Christie, S.; Symonds, E.; Li, H. Circadian regulation of appetite and time restricted feeding. Physiol. Behav. 2020, 220, 112873. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.; Morris, C.J.; Shea, S.A. The internal circadian clock increases hunger and appetite in the evening independent of food intake and other behaviors. Obesity 2013, 21, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Baker, F.C.; Driver, H.S. Circadian rhythms, sleep, and the menstrual cycle. Sleep Med. 2007, 8, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.C.; Whelan, E.A.; Lividoti Hibert, E.N.; Spiegelman, D.; Schernhammer, E.S.; Rich-Edwards, J.W. Rotating shift work and menstrual cycle characteristics. Epidemiology 2011, 22, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Stocker, L.J.; Macklon, N.S.; Cheong, Y.C.; Bewley, S.J. Influence of shift work on early reproductive outcomes: A systematic review and meta-analysis. Obstet. Gynecol. 2014, 124, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Bonde, J.P.; Jorgensen, K.T.; Bonzini, M.; Palmer, K.T. Miscarriage and occupational activity: A systematic review and meta-analysis regarding shift work, working hours, lifting, standing, and physical workload. Scand. J. Work Environ. Health 2013, 39, 325–334. [Google Scholar] [CrossRef]
- Grajewski, B.; Whelan, E.A.; Lawson, C.C.; Hein, M.J.; Waters, M.A.; Anderson, J.L.; MacDonald, L.A.; Mertens, C.J.; Tseng, C.Y.; Cassinelli, R.T., II; et al. Miscarriage among flight attendants. Epidemiology 2015, 26, 192–203. [Google Scholar] [CrossRef]
- Suzumori, N.; Ebara, T.; Matsuki, T.; Yamada, Y.; Kato, S.; Omori, T.; Saitoh, S.; Kamijima, M.; Sugiura-Ogasawara, M.; Japan Environment & Children’s Study Group. Effects of long working hours and shift work during pregnancy on obstetric and perinatal outcomes: A large prospective cohort study-Japan Environment and Children’s Study. Birth 2020, 47, 67–79. [Google Scholar] [CrossRef]
- Patil, D.; Enquobahrie, D.A.; Peckham, T.; Seixas, N.; Hajat, A. Retrospective cohort study of the association between maternal employment precarity and infant low birth weight in women in the USA. BMJ Open 2020, 10, e029584. [Google Scholar] [CrossRef]
- Xie, Y.; Tang, Q.; Chen, G.; Xie, M.; Yu, S.; Zhao, J.; Chen, L. New Insights into the Circadian Rhythm and Its Related Diseases. Front. Physiol. 2019, 10, 682. [Google Scholar] [CrossRef]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of circadian rhythms in health and disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef]
- Abbas, S.; Ahmed, I.; Kudo, T.; Iqbal, M.; Lee, Y.J.; Fujiwara, T.; Ohkuma, M. A heavy metal tolerant novel bacterium, Bacillus malikii sp. nov., isolated from tannery effluent wastewater. Antonie Leeuwenhoek 2015, 108, 1319–1330. [Google Scholar] [CrossRef]
- Miller, B.H.; Olson, S.L.; Levine, J.E.; Turek, F.W.; Horton, T.H.; Takahashi, J.S. Vasopressin regulation of the proestrous luteinizing hormone surge in wild-type and Clock mutant mice. Biol. Reprod. 2006, 75, 778–784. [Google Scholar] [CrossRef]
- Pendergast, J.S.; Oda, G.A.; Niswender, K.D.; Yamazaki, S. Period determination in the food-entrainable and methamphetamine-sensitive circadian oscillator(s). Proc. Natl. Acad. Sci. USA 2012, 109, 14218–14223. [Google Scholar] [CrossRef]
- De Bundel, D.; Gangarossa, G.; Biever, A.; Bonnefont, X.; Valjent, E. Cognitive dysfunction, elevated anxiety, and reduced cocaine response in circadian clock-deficient cryptochrome knockout mice. Front. Behav. Neurosci. 2013, 7, 152. [Google Scholar] [CrossRef]
- Mohawk, J.A.; Green, C.B.; Takahashi, J.S. Central and peripheral circadian clocks in mammals. Annu. Rev. Neurosci. 2012, 35, 445–462. [Google Scholar] [CrossRef]
- Whitmore, D.; Foulkes, N.S.; Sassone-Corsi, P. Light acts directly on organs and cells in culture to set the vertebrate circadian clock. Nature 2000, 404, 87–91. [Google Scholar] [CrossRef]
- Bass, J. Circadian topology of metabolism. Nature 2012, 491, 348–356. [Google Scholar] [CrossRef]
- Moore, R.Y.; Speh, J.C.; Leak, R.K. Suprachiasmatic nucleus organization. Cell Tissue Res. 2002, 309, 89–98. [Google Scholar] [CrossRef]
- Putteeraj, M.; Soga, T.; Ubuka, T.; Parhar, I.S. A “Timed” Kiss Is Essential for Reproduction: Lessons from Mammalian Studies. Front. Endocrinol. 2016, 7, 121. [Google Scholar] [CrossRef]
- Robertson, J.L.; Clifton, D.K.; de la Iglesia, H.O.; Steiner, R.A.; Kauffman, A.S. Circadian regulation of Kiss1 neurons: Implications for timing the preovulatory gonadotropin-releasing hormone/luteinizing hormone surge. Endocrinology 2009, 150, 3664–3671. [Google Scholar] [CrossRef] [PubMed]
- Christian, C.A.; Moenter, S.M. The neurobiology of preovulatory and estradiol-induced gonadotropin-releasing hormone surges. Endocr. Rev. 2010, 31, 544–577. [Google Scholar] [CrossRef] [PubMed]
- Neumann, A.M.; Schmidt, C.X.; Brockmann, R.M.; Oster, H. Circadian regulation of endocrine systems. Auton. Neurosci. 2019, 216, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Sen, A.; Sellix, M.T. The Circadian Timing System and Environmental Circadian Disruption: From Follicles to Fertility. Endocrinology 2016, 157, 3366–3373. [Google Scholar] [CrossRef] [PubMed]
- Van der Vinne, V.; Tachinardi, P.; Riede, S.J.; Akkerman, J.; Scheepe, J.; Daan, S.; Hut, R.A. Maximising survival by shifting the daily timing of activity. Ecol. Lett. 2019, 22, 2097–2102. [Google Scholar] [CrossRef]
- Smarr, B.L.; Morris, E.; de la Iglesia, H.O. The dorsomedial suprachiasmatic nucleus times circadian expression of Kiss1 and the luteinizing hormone surge. Endocrinology 2012, 153, 2839–2850. [Google Scholar] [CrossRef]
- Chassard, D.; Bur, I.; Poirel, V.J.; Mendoza, J.; Simonneaux, V. Evidence for a Putative Circadian Kiss-Clock in the Hypothalamic AVPV in Female Mice. Endocrinology 2015, 156, 2999–3011. [Google Scholar] [CrossRef]
- Comninos, A.N.; Wall, M.B.; Demetriou, L.; Shah, A.J.; Clarke, S.A.; Narayanaswamy, S.; Nesbitt, A.; Izzi-Engbeaya, C.; Prague, J.K.; Abbara, A.; et al. Kisspeptin modulates sexual and emotional brain processing in humans. J. Clin. Investig. 2017, 127, 709–719. [Google Scholar] [CrossRef]
- Dror, T.; Franks, J.; Kauffman, A.S. Analysis of multiple positive feedback paradigms demonstrates a complete absence of LH surges and GnRH activation in mice lacking kisspeptin signaling. Biol. Reprod. 2013, 88, 146. [Google Scholar] [CrossRef]
- Saedi, S.; Khoradmehr, A.; Mohammad Reza, J.S.; Tamadon, A. The role of neuropeptides and neurotransmitters on kisspeptin/kiss1r-signaling in female reproduction. J. Chem. Neuroanat. 2018, 92, 71–82. [Google Scholar] [CrossRef]
- Kunimura, Y.; Iwata, K.; Ishigami, A.; Ozawa, H. Age-related alterations in hypothalamic kisspeptin, neurokinin B, and dynorphin neurons and in pulsatile LH release in female and male rats. Neurobiol. Aging. 2017, 50, 30–38. [Google Scholar] [CrossRef]
- Garcia, J.P.; Guerriero, K.A.; Keen, K.L.; Kenealy, B.P.; Seminara, S.B.; Terasawa, E. Kisspeptin and Neurokinin B Signaling Network Underlies the Pubertal Increase in GnRH Release in Female Rhesus Monkeys. Endocrinology 2017, 158, 3269–3280. [Google Scholar] [CrossRef]
- Qiu, J.; Nestor, C.C.; Zhang, C.; Padilla, S.L.; Palmiter, R.D.; Kelly, M.J.; Ronnekleiv, O.K. High-frequency stimulation-induced peptide release synchronizes arcuate kisspeptin neurons and excites GnRH neurons. eLife 2016, 5, e16246. [Google Scholar] [CrossRef]
- Kalil, B.; Ribeiro, A.B.; Leite, C.M.; Uchoa, E.T.; Carolino, R.O.; Cardoso, T.S.; Elias, L.L.; Rodrigues, J.A.; Plant, T.M.; Poletini, M.O.; et al. The Increase in Signaling by Kisspeptin Neurons in the Preoptic Area and Associated Changes in Clock Gene Expression That Trigger the LH Surge in Female Rats Are Dependent on the Facilitatory Action of a Noradrenaline Input. Endocrinology 2016, 157, 323–335. [Google Scholar] [CrossRef]
- Adams, C.; Stroberg, W.; DeFazio, R.A.; Schnell, S.; Moenter, S.M. Gonadotropin-Releasing Hormone (GnRH) Neuron Excitability Is Regulated by Estradiol Feedback and Kisspeptin. J. Neurosci. 2018, 38, 1249–1263. [Google Scholar] [CrossRef]
- Williams, W.P., III; Jarjisian, S.G.; Mikkelsen, J.D.; Kriegsfeld, L.J. Circadian control of kisspeptin and a gated GnRH response mediate the preovulatory luteinizing hormone surge. Endocrinology 2011, 152, 595–606. [Google Scholar] [CrossRef]
- Smith, J.T.; Acohido, B.V.; Clifton, D.K.; Steiner, R.A. KiSS-1 neurones are direct targets for leptin in the ob/ob mouse. J. Neuroendocrinol. 2006, 18, 298–303. [Google Scholar] [CrossRef]
- Navarro, V.M.; Castellano, J.M.; McConkey, S.M.; Pineda, R.; Ruiz-Pino, F.; Pinilla, L.; Clifton, D.K.; Tena-Sempere, M.; Steiner, R.A. Interactions between kisspeptin and neurokinin B in the control of GnRH secretion in the female rat. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E202–E210. [Google Scholar] [CrossRef]
- Laposky, A.D.; Bradley, M.A.; Williams, D.L.; Bass, J.; Turek, F.W. Sleep-wake regulation is altered in leptin-resistant (db/db) genetically obese and diabetic mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R2059–R2066. [Google Scholar] [CrossRef]
- Sutton, G.M.; Centanni, A.V.; Butler, A.A. Protein malnutrition during pregnancy in C57BL/6J mice results in offspring with altered circadian physiology before obesity. Endocrinology 2010, 151, 1570–1580. [Google Scholar] [CrossRef]
- Ando, H.; Kumazaki, M.; Motosugi, Y.; Ushijima, K.; Maekawa, T.; Ishikawa, E.; Fujimura, A. Impairment of peripheral circadian clocks precedes metabolic abnormalities in ob/ob mice. Endocrinology 2011, 152, 1347–1354. [Google Scholar] [CrossRef] [PubMed]
- Tonsfeldt, K.J.; Goodall, C.P.; Latham, K.L.; Chappell, P.E. Oestrogen induces rhythmic expression of the Kisspeptin-1 receptor GPR54 in hypothalamic gonadotrophin-releasing hormone-secreting GT1-7 cells. J. Neuroendocrinol. 2011, 23, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Fahrenkrug, J.; Georg, B.; Hannibal, J.; Hindersson, P.; Gras, S. Diurnal rhythmicity of the clock genes Per1 and Per2 in the rat ovary. Endocrinology 2006, 147, 3769–3776. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, T.; Sellix, M.; Pezuk, P.; Menaker, M. Timing of the ovarian circadian clock is regulated by gonadotropins. Endocrinology 2009, 150, 4338–4347. [Google Scholar] [CrossRef] [PubMed]
- Mereness, A.L.; Murphy, Z.C.; Sellix, M.T. Developmental programming by androgen affects the circadian timing system in female mice. Biol. Reprod. 2015, 92, 88. [Google Scholar] [CrossRef]
- Lv, S.; Wang, N.; Ma, J.; Li, W.P.; Chen, Z.J.; Zhang, C. Impaired decidualization caused by downregulation of circadian clock gene BMAL1 contributes to human recurrent miscarriage dagger. Biol. Reprod. 2019, 101, 138–147. [Google Scholar] [CrossRef]
- Kovanen, L.; Saarikoski, S.T.; Aromaa, A.; Lonnqvist, J.; Partonen, T. ARNTL (BMAL1) and NPAS2 gene variants contribute to fertility and seasonality. PLoS ONE 2010, 5, e10007. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, N.; Bao, H.; Jiang, Y.; Yang, N.; Wu, K.; Wu, J.; Wang, H.; Kong, S.; Zhang, Y. Circadian gene PER1 senses progesterone signal during human endometrial decidualization. J. Endocrinol. 2019, 243, 229–242. [Google Scholar] [CrossRef]
- Seron-Ferre, M.; Valenzuela, G.J.; Torres-Farfan, C. Circadian clocks during embryonic and fetal development. Birth Defects Res. C Embryo Today 2007, 81, 204–214. [Google Scholar] [CrossRef]
- Akiyama, S.; Ohta, H.; Watanabe, S.; Moriya, T.; Hariu, A.; Nakahata, N.; Chisaka, H.; Matsuda, T.; Kimura, Y.; Tsuchiya, S.; et al. The uterus sustains stable biological clock during pregnancy. Tohoku J. Exp. Med. 2010, 221, 287–298. [Google Scholar] [CrossRef]
- Papacleovoulou, G.; Nikolova, V.; Oduwole, O.; Chambers, J.; Vazquez-Lopez, M.; Jansen, E.; Nicolaides, K.; Parker, M.; Williamson, C. Gestational disruptions in metabolic rhythmicity of the liver, muscle, and placenta affect fetal size. FASEB J. 2017, 31, 1698–1708. [Google Scholar] [CrossRef]
- Alvarez, J.D.; Hansen, A.; Ord, T.; Bebas, P.; Chappell, P.E.; Giebultowicz, J.M.; Williams, C.; Moss, S.; Sehgal, A. The circadian clock protein BMAL1 is necessary for fertility and proper testosterone production in mice. J. Biol. Rhythm. 2008, 23, 26–36. [Google Scholar] [CrossRef]
- Liu, Y.; Johnson, B.P.; Shen, A.L.; Wallisser, J.A.; Krentz, K.J.; Moran, S.M.; Sullivan, R.; Glover, E.; Parlow, A.F.; Drinkwater, N.R.; et al. Loss of BMAL1 in ovarian steroidogenic cells results in implantation failure in female mice. Proc. Natl. Acad. Sci. USA 2014, 111, 14295–14300. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, S.; Xu, W.; Ying, J.; Qu, Y.; Jiang, X.; Zhang, A.; Yue, Y.; Zhou, R.; Ruan, T.; et al. Critical Roles of the Circadian Transcription Factor BMAL1 in Reproductive Endocrinology and Fertility. Front. Endocrinol. 2022, 13, 818272. [Google Scholar] [CrossRef]
- Ono, M.; Toyoda, N.; Kagami, K.; Hosono, T.; Matsumoto, T.; Horike, S.I.; Yamazaki, R.; Nakamura, M.; Mizumoto, Y.; Fujiwara, T.; et al. Uterine Deletion of Bmal1 Impairs Placental Vascularization and Induces Intrauterine Fetal Death in Mice. Int. J. Mol. Sci. 2022, 23, 7637. [Google Scholar] [CrossRef]
- Ratajczak, C.K.; Boehle, K.L.; Muglia, L.J. Impaired steroidogenesis and implantation failure in Bmal1−/− mice. Endocrinology 2009, 150, 1879–1885. [Google Scholar] [CrossRef]
- Chu, A.; Zhu, L.; Blum, I.D.; Mai, O.; Leliavski, A.; Fahrenkrug, J.; Oster, H.; Boehm, U.; Storch, K.F. Global but not gonadotrope-specific disruption of Bmal1 abolishes the luteinizing hormone surge without affecting ovulation. Endocrinology 2013, 154, 2924–2935. [Google Scholar] [CrossRef]
- Pilorz, V.; Steinlechner, S. Low reproductive success in Per1 and Per2 mutant mouse females due to accelerated ageing? Reproduction 2008, 135, 559–568. [Google Scholar] [CrossRef]
- Zheng, B.; Albrecht, U.; Kaasik, K.; Sage, M.; Lu, W.; Vaishnav, S.; Li, Q.; Sun, Z.S.; Eichele, G.; Bradley, A.; et al. Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock. Cell 2001, 105, 683–694. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, C.; Li, Y.; Jiang, H.; Yang, P.; Tang, J.; Xu, Y.; Wang, H.; He, Y. Loss-of-function mutations with circadian rhythm regulator Per1/Per2 lead to premature ovarian insufficiency dagger. Biol. Reprod. 2019, 100, 1066–1072. [Google Scholar] [CrossRef]
- Vitaterna, M.H.; King, D.P.; Chang, A.M.; Kornhauser, J.M.; Lowrey, P.L.; McDonald, J.D.; Dove, W.F.; Pinto, L.H.; Turek, F.W.; Takahashi, J.S. Mutagenesis and mapping of a mouse gene, Clock, essential for circadian behavior. Science 1994, 264, 719–725. [Google Scholar] [CrossRef] [PubMed]
- Turek, F.W.; Joshu, C.; Kohsaka, A.; Lin, E.; Ivanova, G.; McDearmon, E.; Laposky, A.; Losee-Olson, S.; Easton, A.; Jensen, D.R.; et al. Obesity and metabolic syndrome in circadian Clock mutant mice. Science 2005, 308, 1043–1045. [Google Scholar] [CrossRef]
- Lamia, K.A.; Storch, K.F.; Weitz, C.J. Physiological significance of a peripheral tissue circadian clock. Proc. Natl. Acad. Sci. USA 2008, 105, 15172–15177. [Google Scholar] [CrossRef] [PubMed]
- Barclay, J.L.; Shostak, A.; Leliavski, A.; Tsang, A.H.; Johren, O.; Muller-Fielitz, H.; Landgraf, D.; Naujokat, N.; van der Horst, G.T.; Oster, H. High-fat diet-induced hyperinsulinemia and tissue-specific insulin resistance in Cry-deficient mice. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E1053–E1063. [Google Scholar] [CrossRef]
- Hatori, M.; Vollmers, C.; Zarrinpar, A.; DiTacchio, L.; Bushong, E.A.; Gill, S.; Leblanc, M.; Chaix, A.; Joens, M.; Fitzpatrick, J.A.; et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012, 15, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Sherman, H.; Genzer, Y.; Cohen, R.; Chapnik, N.; Madar, Z.; Froy, O. Timed high-fat diet resets circadian metabolism and prevents obesity. FASEB J. 2012, 26, 3493–3502. [Google Scholar] [CrossRef]
- Chaix, A.; Lin, T.; Le, H.D.; Chang, M.W.; Panda, S. Time-Restricted Feeding Prevents Obesity and Metabolic Syndrome in Mice Lacking a Circadian Clock. Cell Metab. 2019, 29, 303–319.e4. [Google Scholar] [CrossRef]
- Arble, D.M.; Bass, J.; Laposky, A.D.; Vitaterna, M.H.; Turek, F.W. Circadian timing of food intake contributes to weight gain. Obesity 2009, 17, 2100–2102. [Google Scholar] [CrossRef]
- Dolatshad, H.; Campbell, E.A.; O’Hara, L.; Maywood, E.S.; Hastings, M.H.; Johnson, M.H. Developmental and reproductive performance in circadian mutant mice. Hum. Reprod. 2006, 21, 68–79. [Google Scholar] [CrossRef]
- Wagner, G.C.; Johnston, J.D.; Tournier, B.B.; Ebling, F.J.; Hazlerigg, D.G. Melatonin induces gene-specific effects on rhythmic mRNA expression in the pars tuberalis of the Siberian hamster (Phodopus sungorus). Eur. J. Neurosci. 2007, 25, 485–490. [Google Scholar] [CrossRef]
- Kosonsiriluk, S.; Mauro, L.J.; Chaiworakul, V.; Chaiseha, Y.; El Halawani, M.E. Photoreceptive oscillators within neurons of the premammillary nucleus (PMM) and seasonal reproduction in temperate zone birds. Gen. Comp. Endocrinol. 2013, 190, 149–155. [Google Scholar] [CrossRef]
- Yoshimura, T.; Yasuo, S.; Watanabe, M.; Iigo, M.; Yamamura, T.; Hirunagi, K.; Ebihara, S. Light-induced hormone conversion of T4 to T3 regulates photoperiodic response of gonads in birds. Nature 2003, 426, 178–181. [Google Scholar] [CrossRef]
- Nakao, N.; Ono, H.; Yamamura, T.; Anraku, T.; Takagi, T.; Higashi, K.; Yasuo, S.; Katou, Y.; Kageyama, S.; Uno, Y.; et al. Thyrotrophin in the pars tuberalis triggers photoperiodic response. Nature 2008, 452, 317–322. [Google Scholar] [CrossRef]
- Olcese, J.; Lozier, S.; Paradise, C. Melatonin and the circadian timing of human parturition. Reprod. Sci. 2013, 20, 168–174. [Google Scholar] [CrossRef]
- Olcese, J. Circadian clocks and pregnancy. Front. Endocrinol. 2014, 5, 123. [Google Scholar] [CrossRef]
- Sharkey, J.T.; Puttaramu, R.; Word, R.A.; Olcese, J. Melatonin synergizes with oxytocin to enhance contractility of human myometrial smooth muscle cells. J. Clin. Endocrinol. Metab. 2009, 94, 421–427. [Google Scholar] [CrossRef]
- Nakamura, T.J.; Sellix, M.T.; Menaker, M.; Block, G.D. Estrogen directly modulates circadian rhythms of PER2 expression in the uterus. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1025–E1031. [Google Scholar] [CrossRef]
- Loh, D.H.; Kuljis, D.A.; Azuma, L.; Wu, Y.; Truong, D.; Wang, H.B.; Colwell, C.S. Disrupted reproduction, estrous cycle, and circadian rhythms in female mice deficient in vasoactive intestinal peptide. J. Biol. Rhythm. 2014, 29, 355–369. [Google Scholar] [CrossRef]
- Gamble, K.L.; Resuehr, D.; Johnson, C.H. Shift work and circadian dysregulation of reproduction. Front. Endocrinol. 2013, 4, 92. [Google Scholar] [CrossRef]
- Miller, B.H.; Takahashi, J.S. Central circadian control of female reproductive function. Front. Endocrinol. 2013, 4, 195. [Google Scholar] [CrossRef]
- Ratajczak, C.K.; Asada, M.; Allen, G.C.; McMahon, D.G.; Muglia, L.M.; Smith, D.; Bhattacharyya, S.; Muglia, L.J. Generation of myometrium-specific Bmal1 knockout mice for parturition analysis. Reprod. Fertil. Dev. 2012, 24, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Andrisani, A.; Sabbadin, C.; Minardi, S.; Favaro, A.; Dona, G.; Bordin, L.; Ambrosini, G.; Armanini, D. Persistent amenorrhea and decreased DHEAS to cortisol ratio after recovery from anorexia nervosa. Gynecol. Endocrinol. 2017, 33, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Luisi, S.; Ciani, V.; Podfigurna-Stopa, A.; Lazzeri, L.; De Pascalis, F.; Meczekalski, B.; Petraglia, F. Serum anti-Mullerian hormone, inhibin B, and total inhibin levels in women with hypothalamic amenorrhea and anorexia nervosa. Gynecol. Endocrinol. 2012, 28, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Booth, P.J.; Cosgrove, J.R.; Foxcroft, G.R. Endocrine and metabolic responses to realimentation in feed-restricted prepubertal gilts: Associations among gonadotropins, metabolic hormones, glucose and uteroovarian development. J. Anim. Sci. 1996, 74, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, H.; Pasquier, A.; Mounier, A.M.; Prunier, A. Influence of feed restriction during lactation on gonadotropic hormones and ovarian development in primiparous sows. J. Anim. Sci. 1998, 76, 856–863. [Google Scholar] [CrossRef]
- Veldhuis, J.D.; Iranmanesh, A.; Evans, W.S.; Lizarralde, G.; Thorner, M.O.; Vance, M.L. Amplitude suppression of the pulsatile mode of immunoradiometric luteinizing hormone release in fasting-induced hypoandrogenemia in normal men. J. Clin. Endocrinol. Metab. 1993, 76, 587–593. [Google Scholar]
- Brito, L.F.; Barth, A.D.; Rawlings, N.C.; Wilde, R.E.; Crews, D.H.J.; Boisclair, Y.R.; Ehrhardt, R.A.; Kastelic, J.P. Effect of feed restriction during calfhood on serum concentrations of metabolic hormones, gonadotropins, testosterone, and on sexual development in bulls. Reproduction 2007, 134, 171–181. [Google Scholar] [CrossRef]
- Brito, L.F.; Barth, A.D.; Rawlings, N.C.; Wilde, R.E.; Crews, D.H.J.; Mir, P.S.; Kastelic, J.P. Effect of nutrition during calfhood and peripubertal period on serum metabolic hormones, gonadotropins and testosterone concentrations, and on sexual development in bulls. Domest. Anim. Endocrinol. 2007, 33, 1–18. [Google Scholar] [CrossRef]
- Tropp, J.; Markus, E.J. Effects of mild food deprivation on the estrous cycle of rats. Physiol. Behav. 2001, 73, 553–559. [Google Scholar] [CrossRef]
- Stephan, F.K. Phase shifts of circadian rhythms in activity entrained to food access. Physiol. Behav. 1984, 32, 663–671. [Google Scholar] [CrossRef]
- Mistlberger, R.E. Circadian food-anticipatory activity: Formal models and physiological mechanisms. Neurosci. Biobehav. Rev. 1994, 18, 171–195. [Google Scholar] [CrossRef]
- Vollmers, C.; Gill, S.; DiTacchio, L.; Pulivarthy, S.R.; Le, H.D.; Panda, S. Time of feeding and the intrinsic circadian clock drive rhythms in hepatic gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 21453–21458. [Google Scholar] [CrossRef]
- Hosono, T.; Ono, M.; Daikoku, T.; Mieda, M.; Nomura, S.; Kagami, K.; Iizuka, T.; Nakata, R.; Fujiwara, T.; Fujiwara, H.; et al. Time-Restricted Feeding Regulates Circadian Rhythm of Murine Uterine Clock. Curr. Dev. Nutr. 2021, 5, nzab064. [Google Scholar] [CrossRef]
- Fujiwara, T.; Nakata, R. Skipping breakfast is associated with reproductive dysfunction in post-adolescent female college students. Appetite 2010, 55, 714–717. [Google Scholar] [CrossRef]
- Fujiwara, T.; Ono, M.; Mieda, M.; Yoshikawa, H.; Nakata, R.; Daikoku, T.; Sekizuka-Kagami, N.; Maida, Y.; Ando, H.; Fujiwara, H. Adolescent Dietary Habit-induced Obstetric and Gynecologic Disease (ADHOGD) as a New Hypothesis-Possible Involvement of Clock System. Nutrients 2020, 12, 1294. [Google Scholar] [CrossRef]
- Fujiwara, T. Skipping breakfast is associated with dysmenorrhea in young women in Japan. Int. J. Food Sci. Nutr. 2003, 54, 505–509. [Google Scholar] [CrossRef]
- Angelin, P.; Dileep, D.; Manju, T.; Veena, M.; Pradeep, D.; Amreen, K.; Soumitra, S. Effect of Skipping Breakfast on Young Girls’ Menstruation. Ind. J. Youth Adol. Health 2017, 4, 17–20. [Google Scholar]
- Abu Helwa, H.A.; Mitaeb, A.A.; Al-Hamshri, S.; Sweileh, W.M. Prevalence of dysmenorrhea and predictors of its pain intensity among Palestinian female university students. BMC Women’s Health 2018, 18, 18. [Google Scholar] [CrossRef]
- Hu, Z.; Tang, L.; Chen, L.; Kaminga, A.C.; Xu, H. Prevalence and Risk Factors Associated with Primary Dysmenorrhea among Chinese Female University Students: A Cross-sectional Study. J. Pediatr. Adolesc. Gynecol. 2020, 33, 15–22. [Google Scholar] [CrossRef]
- Fujiwara, T.; Ono, M.; Iizuka, T.; Sekizuka-Kagami, N.; Maida, Y.; Adachi, Y.; Fujiwara, H.; Yoshikawa, H. Breakfast Skipping in Female College Students Is a Potential and Preventable Predictor of Gynecologic Disorders at Health Service Centers. Diagnostics 2020, 10, 476. [Google Scholar] [CrossRef]
- Shibata, S.; Tahara, Y.; Hirao, A. The adjustment and manipulation of biological rhythms by light, nutrition and abused drugs. Adv. Drug Deliv. Rev. 2010, 62, 918–927. [Google Scholar] [CrossRef] [PubMed]
- Bajalan, Z.; Alimoradi, Z.; Moafi, F. Nutrition as a Potential Factor of Primary Dysmenorrhea: A Systematic Review of Observational Studies. Gynecol. Obstet. Investig. 2019, 84, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Dashti, H.S.; Merino, J.; Lane, J.M.; Song, Y.; Smith, C.E.; Tanaka, T.; McKeown, N.M.; Tucker, C.; Sun, D.; Bartz, T.M.; et al. Genome-wide association study of breakfast skipping links clock regulation with food timing. Am. J. Clin. Nutr. 2019, 110, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Hanzawa, F.; Kim, D.; Sun, S.; Laurent, T.; Umeki, M.; Ikeda, S.; Mochizuki, S.; Oda, H. Delayed first active-phase meal, a breakfast-skipping model, led to increased body weight and shifted the circadian oscillation of the hepatic clock and lipid metabolism-related genes in rats fed a high-fat diet. PLoS ONE 2018, 13, e0206669. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Nakata, R.; Ono, M.; Mieda, M.; Ando, H.; Daikoku, T.; Fujiwara, H. Time Restriction of Food Intake During the Circadian Cycle Is a Possible Regulator of Reproductive Function in Postadolescent Female Rats. Curr. Dev. Nutr. 2019, 3, nzy093. [Google Scholar] [CrossRef]
- Fujiwara, T.; Nakata, R. Current problems of food intake in young women in Japan: Their influence on female reproductive function. Reprod. Med. Biol. 2004, 3, 107–114. [Google Scholar] [CrossRef]
- Simonneaux, V.; Bahougne, T.; Angelopoulou, E. Daily rhythms count for female fertility. Best Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 505–519. [Google Scholar] [CrossRef]
- Sellix, M.T.; Murphy, Z.C.; Menaker, M. Excess androgen during puberty disrupts circadian organization in female rats. Endocrinology 2013, 154, 1636–1647. [Google Scholar] [CrossRef]
- Huhtaniemi, I. Mutations along the pituitary-gonadal axis affecting sexual maturation: Novel information from transgenic and knockout mice. Mol. Cell. Endocrinol. 2006, 254–255, 84–90. [Google Scholar] [CrossRef]
- Resuehr, H.E.; Resuehr, D.; Olcese, J. Induction of mPer1 expression by GnRH in pituitary gonadotrope cells involves EGR-1. Mol. Cell. Endocrinol. 2009, 311, 120–125. [Google Scholar] [CrossRef]
- Perry, J.R.; Day, F.; Elks, C.E.; Sulem, P.; Thompson, D.J.; Ferreira, T.; He, C.; Chasman, D.I.; Esko, T.; Thorleifsson, G.; et al. Parent-of-origin-specific allelic associations among 106 genomic loci for age at menarche. Nature 2014, 514, 92–97. [Google Scholar] [CrossRef]
- Bayarri, M.J.; Rodriguez, L.; Zanuy, S.; Madrid, J.A.; Sanchez-Vazquez, F.J.; Kagawa, H.; Okuzawa, K.; Carrillo, M. Effect of photoperiod manipulation on the daily rhythms of melatonin and reproductive hormones in caged European sea bass (Dicentrarchus labrax). Gen. Comp. Endocrinol. 2004, 136, 72–81. [Google Scholar] [CrossRef]
- Bayarri, M.J.; Zanuy, S.; Yilmaz, O.; Carrillo, M. Effects of continuous light on the reproductive system of European sea bass gauged by alterations of circadian variations during their first reproductive cycle. Chronobiol. Int. 2009, 26, 184–199. [Google Scholar] [CrossRef]
- Flynn-Evans, E.E.; Stevens, R.G.; Tabandeh, H.; Schernhammer, E.S.; Lockley, S.W. Effect of light perception on menarche in blind women. Ophthalmic Epidemiol. 2009, 16, 243–248. [Google Scholar] [CrossRef]
- De Vries, L.; Kauschansky, A.; Shohat, M.; Phillip, M. Familial central precocious puberty suggests autosomal dominant inheritance. J. Clin. Endocrinol. Metab. 2004, 89, 1794–1800. [Google Scholar] [CrossRef]
- Wehkalampi, K.; Widen, E.; Laine, T.; Palotie, A.; Dunkel, L. Patterns of inheritance of constitutional delay of growth and puberty in families of adolescent girls and boys referred to specialist pediatric care. J. Clin. Endocrinol. Metab. 2008, 93, 723–728. [Google Scholar] [CrossRef]
- Day, F.R.; Thompson, D.J.; Helgason, H.; Chasman, D.I.; Finucane, H.; Sulem, P.; Ruth, K.S.; Whalen, S.; Sarkar, A.K.; Albrecht, E.; et al. Genomic analyses identify hundreds of variants associated with age at menarche and support a role for puberty timing in cancer risk. Nat. Genet. 2017, 49, 834–841. [Google Scholar] [CrossRef]
- Livadas, S.; Chrousos, G.P. Molecular and Environmental Mechanisms Regulating Puberty Initiation: An Integrated Approach. Front. Endocrinol. 2019, 10, 828. [Google Scholar] [CrossRef] [PubMed]
- Beszterda, M.; Franski, R. Endocrine disruptor compounds in environment: As a danger for children health. Pediatr. Endocrinol. Diabetes Metab. 2018, 24, 88–95. [Google Scholar] [CrossRef]
- Fudvoye, J.; Lopez-Rodriguez, D.; Franssen, D.; Parent, A.S. Endocrine disrupters and possible contribution to pubertal changes. Best Pract. Res. Clin. Endocrinol. Metab. 2019, 33, 101300. [Google Scholar] [CrossRef]
- Sorensen, K.; Mouritsen, A.; Aksglaede, L.; Hagen, C.P.; Mogensen, S.S.; Juul, A. Recent secular trends in pubertal timing: Implications for evaluation and diagnosis of precocious puberty. Horm. Res. Paediatr. 2012, 77, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Ochiai, M.; Iida, M.; Agusa, T.; Takaguchi, K.; Fujii, S.; Nomiyama, K.; Iwata, H. Effects of 4-Hydroxy-2,3,3’,4’,5-Pentachlorobiphenyl (4-OH-CB107) on Liver Transcriptome in Rats: Implication in the Disruption of Circadian Rhythm and Fatty Acid Metabolism. Toxicol. Sci. 2018, 165, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, S.; Mirbahai, L.; Castiglioni, S.; Fent, K. Transcriptional and physiological responses induced by binary mixtures of drospirenone and progesterone in zebrafish (Danio rerio). Environ. Sci. Technol. 2014, 48, 3523–3531. [Google Scholar] [CrossRef]
- Zhao, Y.; Castiglioni, S.; Fent, K. Synthetic progestins medroxyprogesterone acetate and dydrogesterone and their binary mixtures adversely affect reproduction and lead to histological and transcriptional alterations in zebrafish (Danio rerio). Environ. Sci. Technol. 2015, 49, 4636–4645. [Google Scholar] [CrossRef]
- Wang, J.; Wang, X.; Xiong, C.; Liu, J.; Hu, B.; Zheng, L. Chronic bisphenol A exposure alters behaviors of zebrafish (Danio rerio). Environ. Pollut. 2015, 206, 275–281. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ono, M.; Ando, H.; Daikoku, T.; Fujiwara, T.; Mieda, M.; Mizumoto, Y.; Iizuka, T.; Kagami, K.; Hosono, T.; Nomura, S.; et al. The Circadian Clock, Nutritional Signals and Reproduction: A Close Relationship. Int. J. Mol. Sci. 2023, 24, 1545. https://doi.org/10.3390/ijms24021545
Ono M, Ando H, Daikoku T, Fujiwara T, Mieda M, Mizumoto Y, Iizuka T, Kagami K, Hosono T, Nomura S, et al. The Circadian Clock, Nutritional Signals and Reproduction: A Close Relationship. International Journal of Molecular Sciences. 2023; 24(2):1545. https://doi.org/10.3390/ijms24021545
Chicago/Turabian StyleOno, Masanori, Hitoshi Ando, Takiko Daikoku, Tomoko Fujiwara, Michihiro Mieda, Yasunari Mizumoto, Takashi Iizuka, Kyosuke Kagami, Takashi Hosono, Satoshi Nomura, and et al. 2023. "The Circadian Clock, Nutritional Signals and Reproduction: A Close Relationship" International Journal of Molecular Sciences 24, no. 2: 1545. https://doi.org/10.3390/ijms24021545
APA StyleOno, M., Ando, H., Daikoku, T., Fujiwara, T., Mieda, M., Mizumoto, Y., Iizuka, T., Kagami, K., Hosono, T., Nomura, S., Toyoda, N., Sekizuka-Kagami, N., Maida, Y., Kuji, N., Nishi, H., & Fujiwara, H. (2023). The Circadian Clock, Nutritional Signals and Reproduction: A Close Relationship. International Journal of Molecular Sciences, 24(2), 1545. https://doi.org/10.3390/ijms24021545