

From Immunosenescence to Aging Types—Establishing Reference Intervals for Immune Age Biomarkers by Centile Estimation

 , ,
, ,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
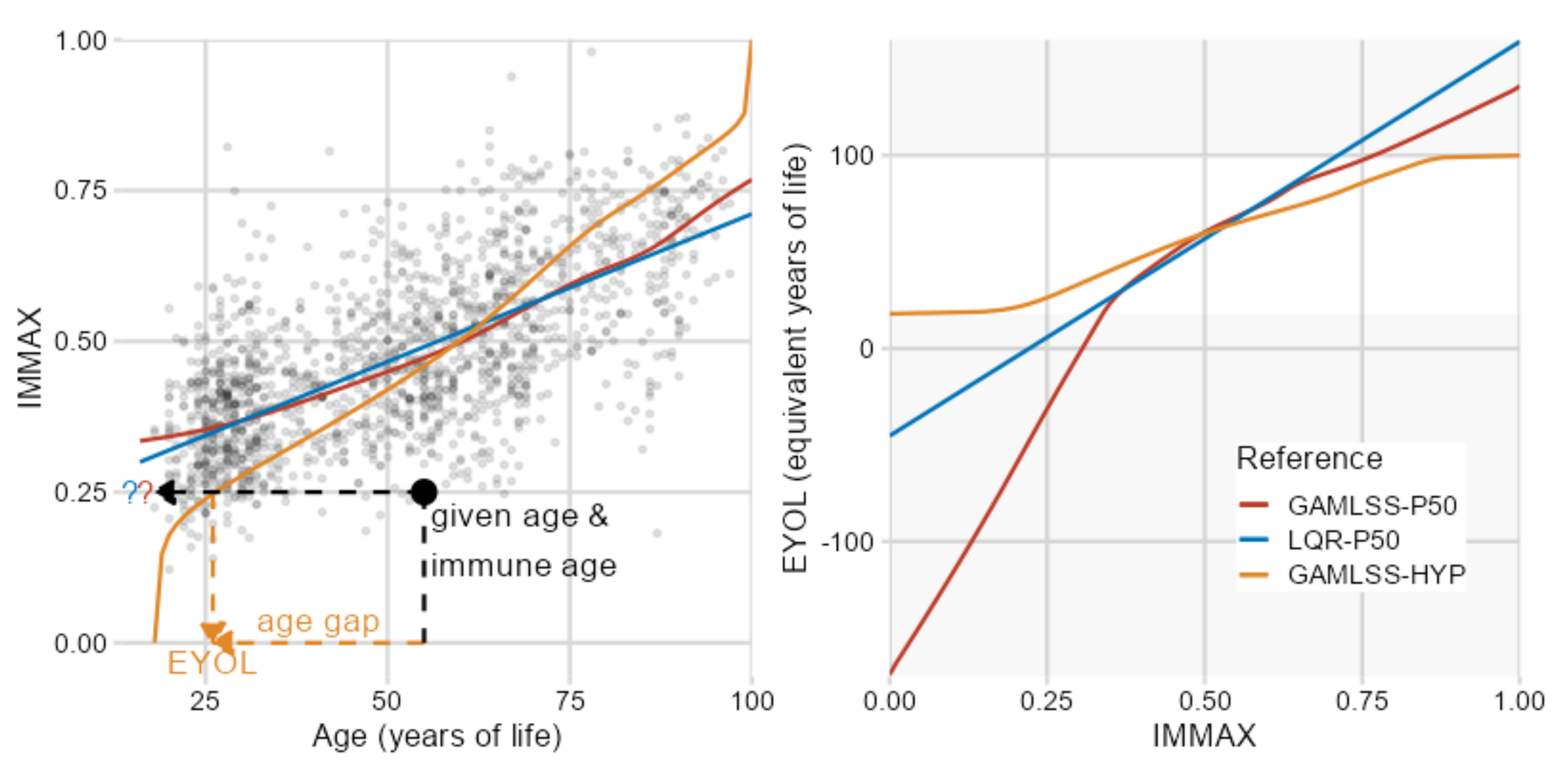
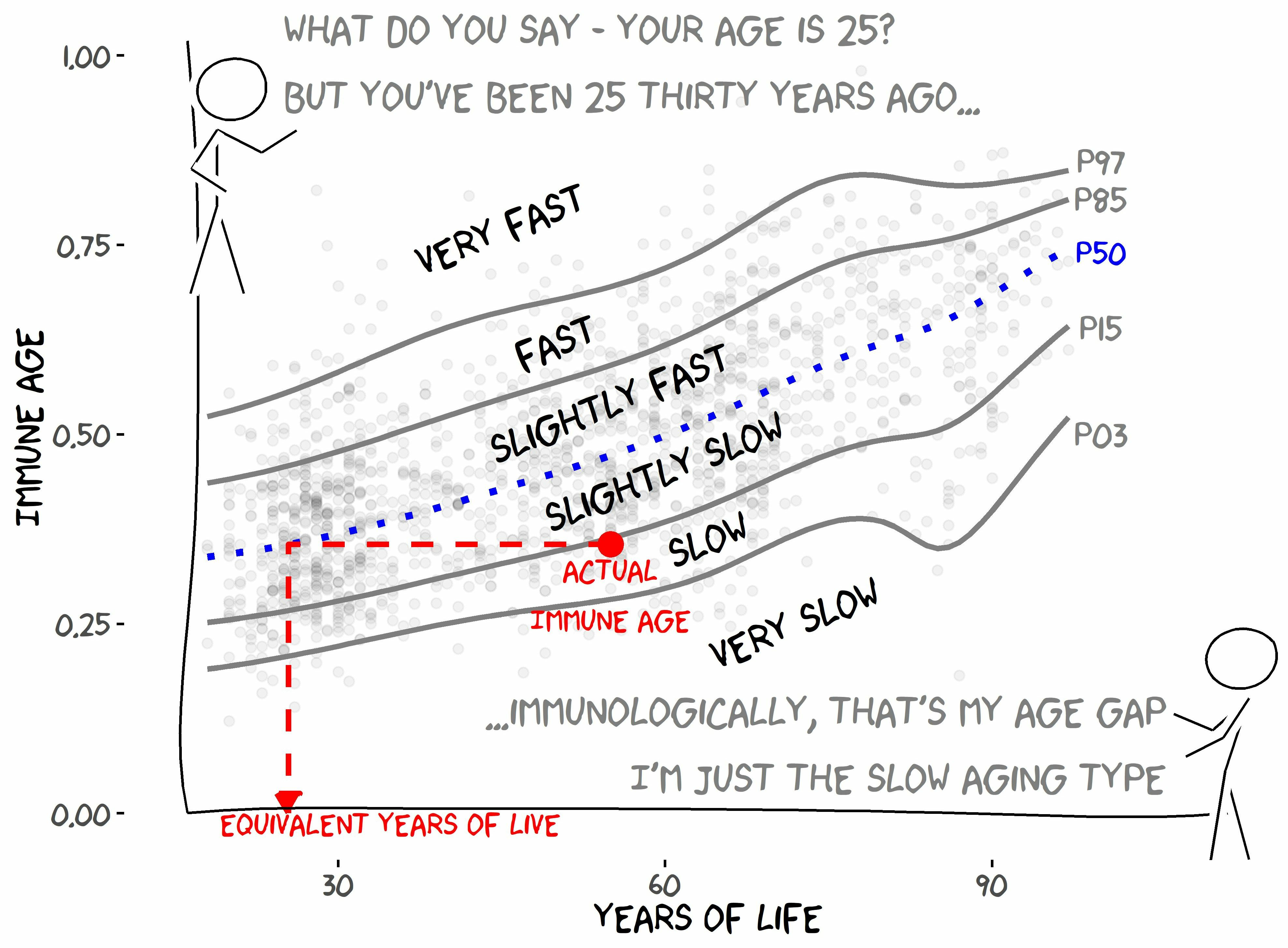
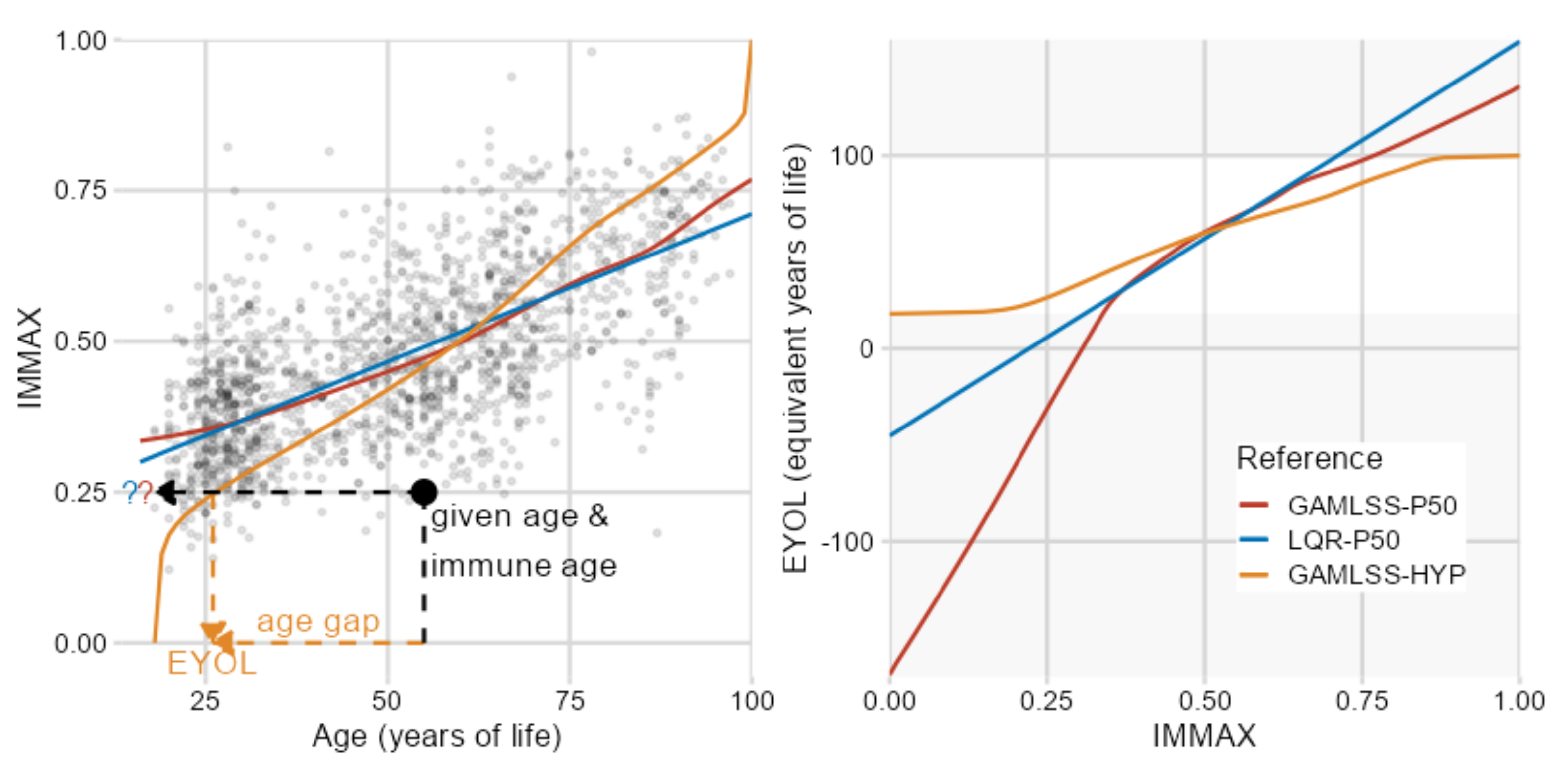
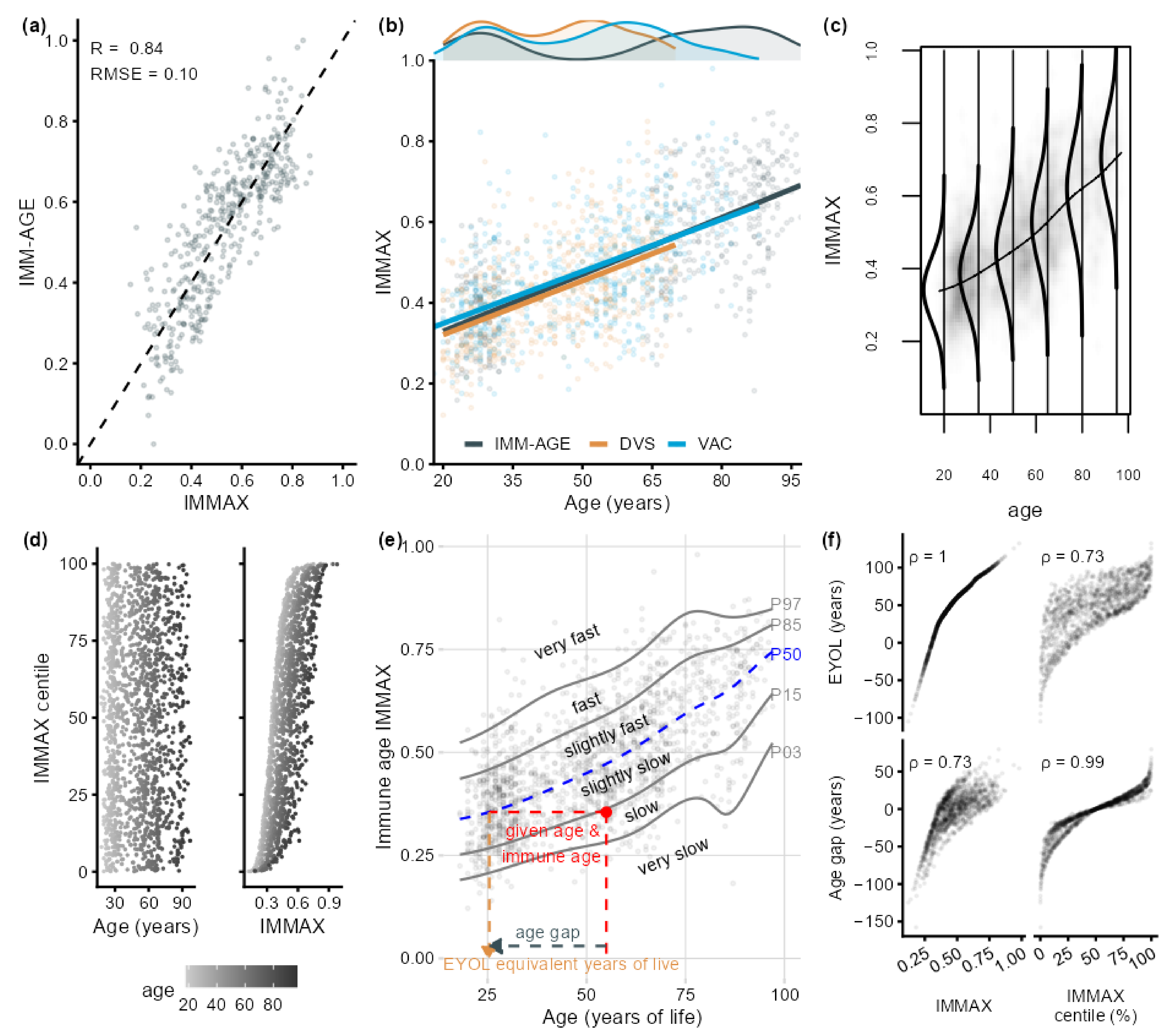
2.1. IMMAX Centile Estimation, Immunological Aging Types, and Age Gap
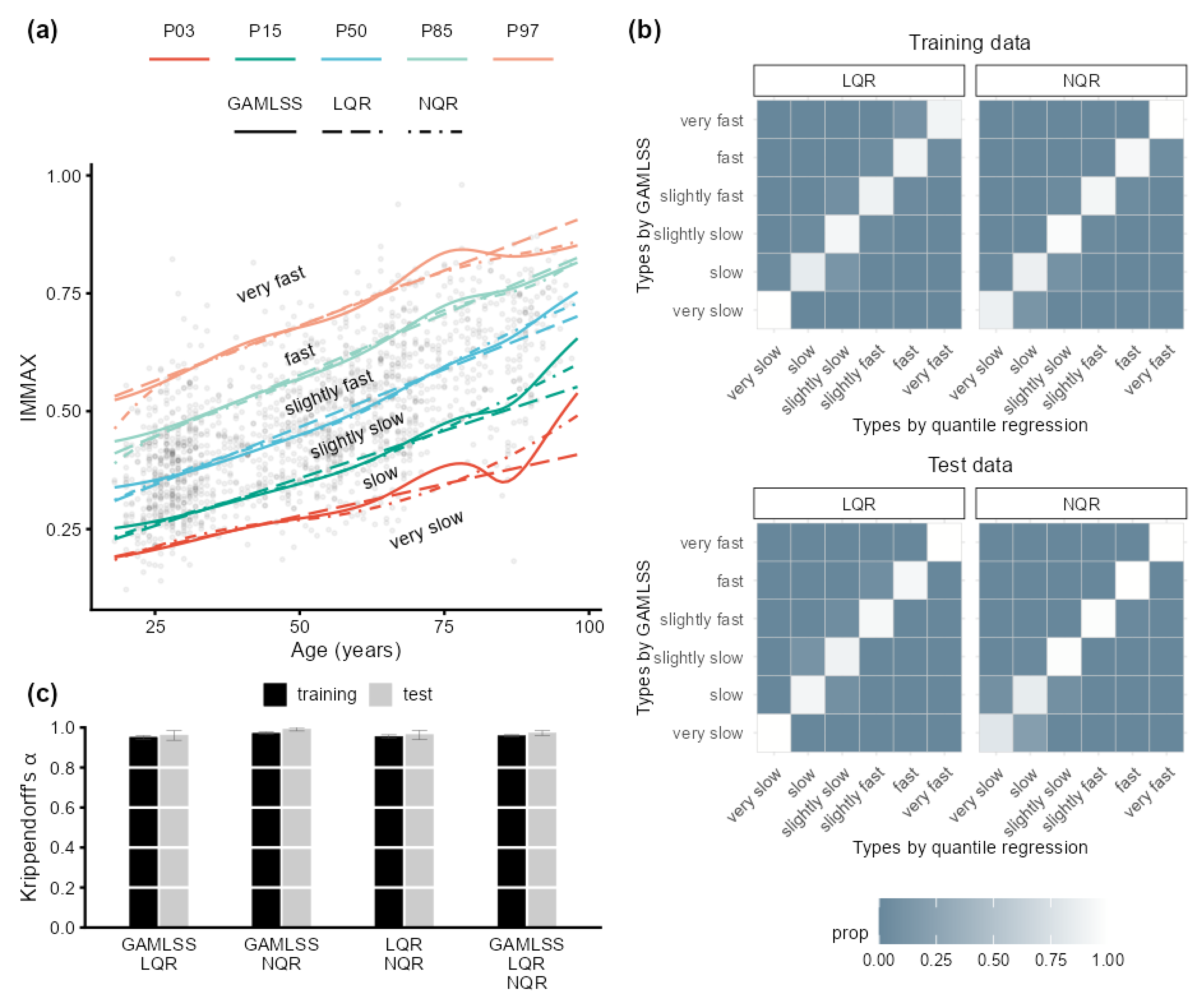
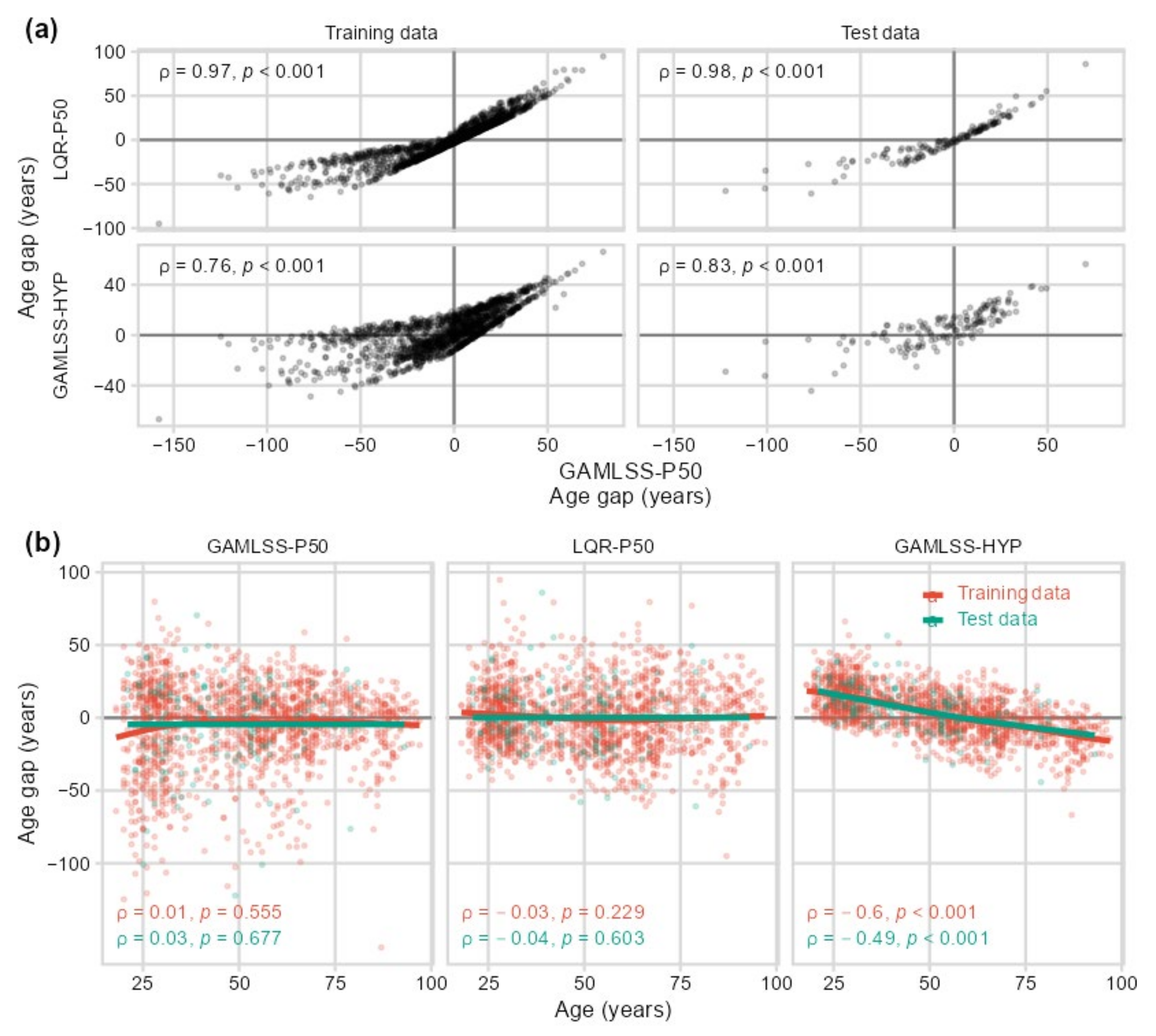
2.2. Sensitivity to Centile Estimation Modeling Strategy
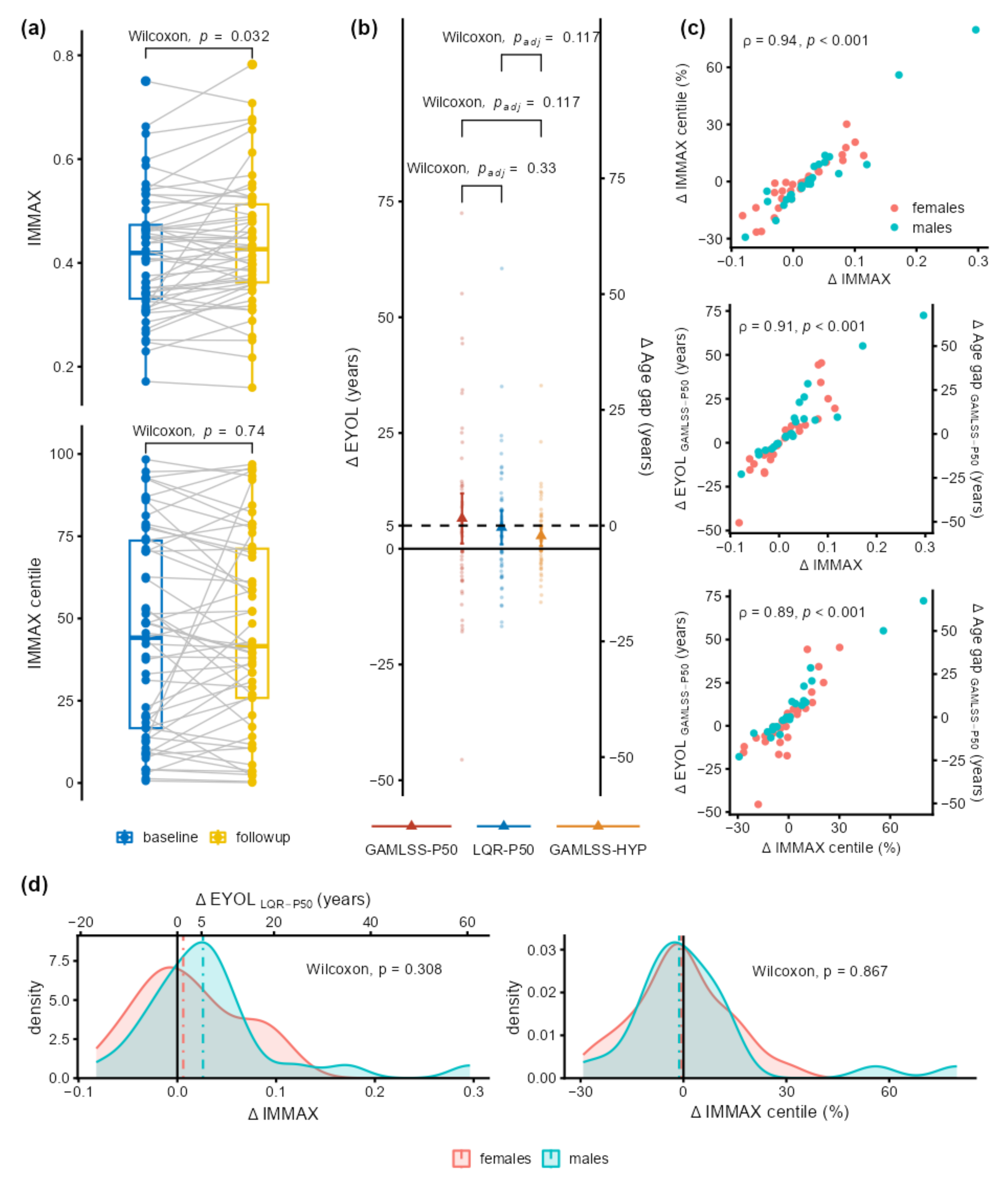
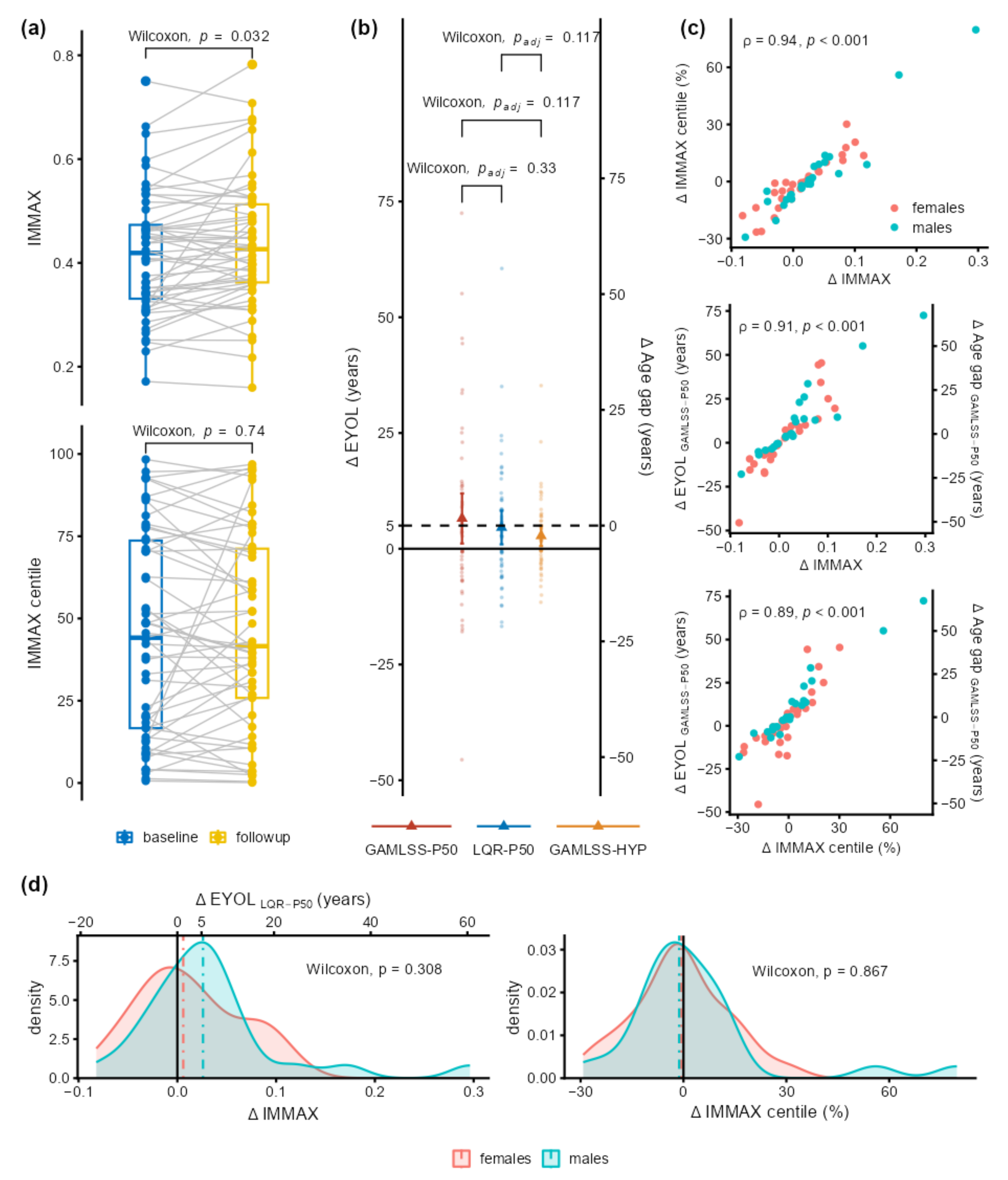
2.3. Application to Longitudinal Data from the Dortmund Vital Study
3. Discussion
4. Materials and Methods
4.1. Datasets
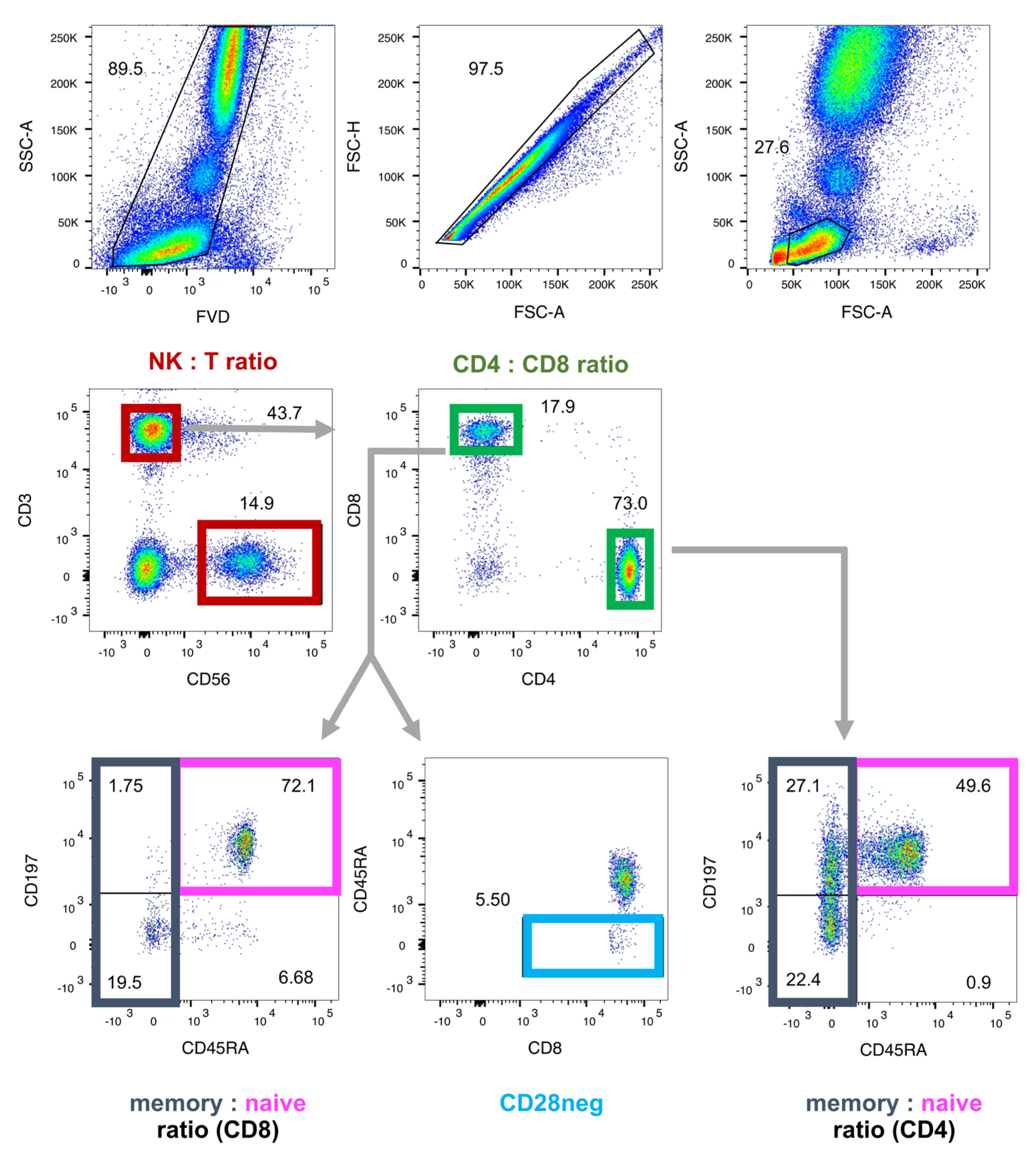
4.2. Molecular Biomarkers of Immunosenescence by Flow Cytometry
4.3. Data Analysis and Statistics
4.4. Preliminary Longitudinal Data from the Dortmund Vital Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Training Data N = 1580 | Test Data N = 150 |
|---|---|---|
| Age (years) | 51 ± 20 (18, 97) | 45 ± 16 (21, 93) |
| IMMAX | 0.47 ± 0.14 (0.12, 0.98) | 0.44 ± 0.13 (0.17, 0.85) |
| log (%memory/%naive) CD8 | −1.51 ± 0.77 (−8.17, 0.87) | −1.44 ± 0.68 (−4.23, 0.03) |
| log (%memory/%naive) CD4 | 1.17 ± 0.60 (−1.22, 4.32) | 1.11 ± 0.50 (0.06, 2.84) |
| logit (%CD8 CD28neg) | 0.55 ± 1.40 (−4.23, 13.81) | 0.31 ± 1.16 (−2.30, 4.48) |
| log (%NK/%T) | 0.57 ± 0.84 (−2.08, 5.43) | 0.43 ± 0.73 (−1.40, 3.42) |
| log (%CD4/%CD8) | −1.05 ± 1.09 (−5.26, 1.94) | −1.28 ± 1.00 (−3.80, 1.07) |
| Characteristic | N = 53 | 95% CI |
|---|---|---|
| Female sex | 28 (53%) | 39%, 66% |
| Age at baseline (years) | 44 ± 12 | 40.9, 47.4 |
| ΔIMMAX | 0.02 ± 0.06 | 0.005, 0.040 |
| ΔIMMAX centile (%) | 1.1 ± 18 | −3.87, 6.12 |
| ΔEYOL (years) | ||
| GAMLSS-P50 | 6.5 ± 20 | 1.13, 11.9 |
| LQR-P50 | 4.6 ± 13 | 0.949, 8.22 |
| GAMLSS-HYP | 2.8 ± 8.0 | 0.584, 4.98 |
| ΔAge gap (years) | ||
| GAMLSS-P50 | 1.5 ± 20 | −3.87, 6.94 |
| LQR-P50 | −0.42 ± 13 | −4.05, 3.22 |
| GAMLSS-HYP | −2.2 ± 8.0 | −4.42, −0.019 |
References
- Rutledge, J.; Oh, H.; Wyss-Coray, T. Measuring biological age using omics data. Nat. Rev. Genet. 2022, 23, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.A. The Aging Immune System: Primer and Prospectus. Science 1996, 273, 70–74. [Google Scholar] [CrossRef]
- Fulop, T.; Larbi, A.; Dupuis, G.; Le Page, A.; Frost, E.H.; Cohen, A.A.; Witkowski, J.M.; Franceschi, C. Immunosenescence and Inflamm-Aging As Two Sides of the Same Coin: Friends or Foes? Front. Immunol. 2018, 8, 1960. [Google Scholar] [CrossRef] [PubMed]
- Gayoso, I.; Sanchez-Correa, B.; Campos, C.; Alonso, C.; Pera, A.; Casado, J.G.; Morgado, S.; Tarazona, R.; Solana, R. Immunosenescence of human natural killer cells. J. Innate Immun. 2011, 3, 337–343. [Google Scholar] [CrossRef]
- Ligotti, M.E.; Aiello, A.; Accardi, G.; Aprile, S.; Bonura, F.; Bulati, M.; Gervasi, F.; Giammanco, G.M.; Pojero, F.; Zareian, N.; et al. Analysis of T and NK cell subsets in the Sicilian population from young to supercentenarian: The role of age and gender. Clin. Exp. Immunol. 2021, 205, 198–212. [Google Scholar] [CrossRef]
- Pangrazzi, L.; Weinberger, B. T cells, aging and senescence. Exp. Gerontol. 2020, 134, 110887. [Google Scholar] [CrossRef] [PubMed]
- Huff, W.X.; Kwon, J.H.; Henriquez, M.; Fetcko, K.; Dey, M. The Evolving Role of CD8+CD28− Immunosenescent T Cells in Cancer Immunology. Int. J. Mol. Sci. 2019, 20, 2810. [Google Scholar] [CrossRef]
- Zhang, H.; Weyand, C.M.; Goronzy, J.J. Hallmarks of the aging T-cell system. FEBS J. 2021, 288, 7123–7142. [Google Scholar] [CrossRef]
- Fagnoni, F.F.; Vescovini, R.; Passeri, G.; Bologna, G.; Pedrazzoni, M.; Lavagetto, G.; Casti, A.; Franceschi, C.; Passeri, M.; Sansoni, P. Shortage of circulating naive CD8+ T cells provides new insights on immunodeficiency in aging. Blood 2000, 95, 2860–2868. [Google Scholar] [CrossRef]
- Garrido-Rodríguez, V.; Herrero-Fernández, I.; Castro, M.J.; Castillo, A.; Rosado-Sánchez, I.; Galvá, M.I.; Ramos, R.; Olivas-Martínez, I.; Bulnes-Ramos, Á.; Cañizares, J.; et al. Immunological features beyond CD4/CD8 ratio values in older individuals. Aging 2021, 13, 13443–13459. [Google Scholar] [CrossRef]
- Alpert, A.; Pickman, Y.; Leipold, M.; Rosenberg-Hasson, Y.; Ji, X.; Gaujoux, R.; Rabani, H.; Starosvetsky, E.; Kveler, K.; Schaffert, S.; et al. A clinically meaningful metric of immune age derived from high-dimensional longitudinal monitoring. Nat. Med. 2019, 25, 487–495. [Google Scholar] [CrossRef]
- Sayed, N.; Huang, Y.; Nguyen, K.; Krejciova-Rajaniemi, Z.; Grawe, A.P.; Gao, T.; Tibshirani, R.; Hastie, T.; Alpert, A.; Cui, L.; et al. An inflammatory aging clock (iAge) based on deep learning tracks multimorbidity, immunosenescence, frailty and cardiovascular aging. Nat. Aging 2021, 1, 598–615. [Google Scholar] [CrossRef]
- Rizzo, L.B.; Swardfager, W.; Maurya, P.K.; Graiff, M.Z.; Pedrini, M.; Asevedo, E.; Cassinelli, A.C.; Bauer, M.E.; Cordeiro, Q.; Scott, J.; et al. An immunological age index in bipolar disorder: A confirmatory factor analysis of putative immunosenescence markers and associations with clinical characteristics. Int. J. Methods Psychiatr. Res. 2018, 27, e1614. [Google Scholar] [CrossRef]
- Ramasubramanian, R.; Meier, H.C.S.; Vivek, S.; Klopack, E.; Crimmins, E.M.; Faul, J.; Nikolich-Žugich, J.; Thyagarajan, B. Evaluation of T-cell aging-related immune phenotypes in the context of biological aging and multimorbidity in the Health and Retirement Study. Immun. Ageing 2022, 19, 33. [Google Scholar] [CrossRef]
- Guerville, F.; De Souto Barreto, P.; Ader, I.; Andrieu, S.; Casteilla, L.; Dray, C.; Fazilleau, N.; Guyonnet, S.; Langin, D.; Liblau, R.; et al. Revisiting the Hallmarks of Aging to Identify Markers of Biological Age. J. Prev. Alzheimer’s Dis. 2020, 7, 56–64. [Google Scholar] [CrossRef]
- Frasca, D.; Diaz, A.; Romero, M.; Garcia, D.; Blomberg, B.B. B Cell Immunosenescence. Annu. Rev. Cell Dev. Biol. 2020, 36, 551–574. [Google Scholar] [CrossRef]
- Xu, W.; Wong, G.; Hwang, Y.Y.; Larbi, A. The untwining of immunosenescence and aging. Semin. Immunopathol. 2020, 42, 559–572. [Google Scholar] [CrossRef]
- Pawelec, G. The human immunosenescence phenotype: Does it exist? Semin. Immunopathol. 2020, 42, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.A.; Bentley, C.; Hazeldine, J.; Acharjee, A.; Nahman, O.; Shen-Orr, S.S.; Lord, J.M.; Duggal, N.A. Investigating the potential of a prematurely aged immune phenotype in severely injured patients as predictor of risk of sepsis. Immun. Ageing 2022, 19, 60. [Google Scholar] [CrossRef] [PubMed]
- Bröde, P.; Claus, M.; Gajewski, P.D.; Getzmann, S.; Golka, K.; Hengstler, J.G.; Wascher, E.; Watzl, C. Calibrating a Comprehensive Immune Age Metric to Analyze the Cross Sectional Age-Related Decline in Cardiorespiratory Fitness. Biology 2022, 11, 1576. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, P.D.; Rieker, J.A.; Athanassiou, G.; Bröde, P.; Claus, M.; Golka, K.; Hengstler, J.G.; Kleinsorge, T.; Nitsche, M.A.; Reinders, J.; et al. A Systematic Analysis of Biological, Sociodemographic, Psychosocial, and Lifestyle Factors Contributing to Work Ability Across the Working Life Span: Cross-sectional Study. JMIR Form. Res. 2023, 7, e40818. [Google Scholar] [CrossRef] [PubMed]
- Claus, M.; Bröde, P.; Urlaub, D.; Wolfsdorff, N.; Watzl, C. Investigation of the relationship between Immune Age and Vaccination against SARS-CoV-2. Eur. J. Immunol. 2022, 52, 168. [Google Scholar]
- Bafei, S.E.C.; Shen, C. Biomarkers selection and mathematical modeling in biological age estimation. NPJ Aging 2023, 9, 13. [Google Scholar] [CrossRef]
- Verschoor, C.P.; Belsky, D.W.; Andrew, M.K.; Haynes, L.; Loeb, M.; Pawelec, G.; McElhaney, J.E.; Kuchel, G.A. Advanced biological age is associated with improved antibody responses in older high-dose influenza vaccine recipients over four consecutive seasons. Immun. Ageing 2022, 19, 39. [Google Scholar] [CrossRef]
- Bröde, P.; Fiala, D.; Blazejczyk, K.; Holmér, I.; Jendritzky, G.; Kampmann, B.; Tinz, B.; Havenith, G. Deriving the operational procedure for the Universal Thermal Climate Index (UTCI). Int. J. Biometeorol. 2012, 56, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Ahadi, S.; Zhou, W.; Schüssler-Fiorenza Rose, S.M.; Sailani, M.R.; Contrepois, K.; Avina, M.; Ashland, M.; Brunet, A.; Snyder, M. Personal aging markers and ageotypes revealed by deep longitudinal profiling. Nat. Med. 2020, 26, 83–90. [Google Scholar] [CrossRef]
- WHO Multicentre Growth Reference Study Group; de Onis, M. WHO Child Growth Standards based on length/height, weight and age. Acta Paediatr. 2006, 95, 76–85. [Google Scholar] [CrossRef]
- Rigby, R.A.; Stasinopoulos, D.M. Generalized additive models for location, scale and shape. J. R. Stat. Soc. Ser. C 2005, 54, 507–554. [Google Scholar] [CrossRef]
- Koenker, R.W.; D’Orey, V. Computing Regression Quantiles. J. R. Stat. Soc. Ser. C 1987, 36, 383–393. [Google Scholar] [CrossRef]
- Muggeo, V.M.R.; Torretta, F.; Eilers, P.H.C.; Sciandra, M.; Attanasio, M. Multiple smoothing parameters selection in additive regression quantiles. Stat. Model. 2021, 21, 428–448. [Google Scholar] [CrossRef]
- Gajewski, P.D.; Getzmann, S.; Bröde, P.; Burke, M.; Cadenas, C.; Capellino, S.; Claus, M.; Genç, E.; Golka, K.; Hengstler, J.G.; et al. Impact of Biological and Lifestyle Factors on Cognitive Aging and Work Ability in the Dortmund Vital Study: Protocol of an Interdisciplinary, Cross-sectional, and Longitudinal Study. JMIR Res. Protoc. 2022, 11, e32352. [Google Scholar] [CrossRef]
- Hayes, A.F.; Krippendorff, K. Answering the Call for a Standard Reliability Measure for Coding Data. Commun. Methods Meas. 2007, 1, 77–89. [Google Scholar] [CrossRef]
- Pawelec, G. Hallmarks of human “immunosenescence”: Adaptation or dysregulation? Immun. Ageing 2012, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Ogrodnik, M.; Gladyshev, V.N. The meaning of adaptation in aging: Insights from cellular senescence, epigenetic clocks and stem cell alterations. Nat. Aging 2023, 3, 766–775. [Google Scholar] [CrossRef]
- Bröde, P.; Claus, M.; Urlaub, D.; Wolfsdorff, N.; Watzl, C. Immune age: The feminine side of firemen? In Proceedings of the 10th European Conference on Protective Clothing, Arnhem, The Netherlands, 9–12 May 2023; pp. 156–157. [Google Scholar]
- Claus, M.; Dychus, N.; Ebel, M.; Damaschke, J.; Maydych, V.; Wolf, O.T.; Kleinsorge, T.; Watzl, C. Measuring the immune system: A comprehensive approach for the analysis of immune functions in humans. Arch. Toxicol. 2016, 90, 2481–2495. [Google Scholar] [CrossRef]
- Stasinopoulos, M.D.; Rigby, R.A.; Bastiani, F.D. GAMLSS: A distributional regression approach. Stat. Model. 2018, 18, 248–273. [Google Scholar] [CrossRef]
- Koenker, R.W. Quantreg: Quantile Regression, R Package Version 5.95; The R Foundation: Vienna, Austria, 2023; Available online: https://CRAN.R-project.org/package=quantreg (accessed on 19 July 2023).
- Muggeo, V.M.R.; Sciandra, M.; Tomasello, A.; Calvo, S. Estimating growth charts via nonparametric quantile regression: A practical framework with application in ecology. Environ. Ecol. Stat. 2013, 20, 519–531. [Google Scholar] [CrossRef]
- Krippendorff, K. Computing Krippendorff’s Alpha Reliability; Penn Collection, Departmental Papers (ASC) 43; University of Pennsylvania: Philadelphia, PA, USA, 2011; Available online: https://repository.upenn.edu/handle/20.500.14332/2089 (accessed on 15 July 2023).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://www.R-project.org/ (accessed on 9 July 2023).

| Sample | Antigen | Clone | Fluorochrome | Company | Dilution 1/x |
|---|---|---|---|---|---|
| DVS | live/dead | zombie Yellow | Biolegend (San Diego, CA, USA) | 1000 | |
| CD3 | UCHT1 | BV510 | BD Horizon™ (Franklin Lakes, NJ, USA) | 400 | |
| CD56 | B159 | PE-CF594 | BD Pharmingen™ (Franklin Lakes, NJ, USA) | 100 | |
| CD4 | RPA-T4 | APC-H7 | BD Pharmingen™ | 100 | |
| CD8 | RPA-T8 | FITC | BD Pharmingen™ | 200 | |
| CD197 (CCR7) | 150,503 | Alexa Fluor® 647 | BD Pharmingen™ | 50 | |
| CD45RA | HI100 | Alexa Fluor® 700 | BD Pharmingen™ | 400 | |
| CD28 | CD28.2 | PerCP-Cy™ 5.5 | BD Pharmingen™ | 100 | |
| VAC | live/dead | Fixable Viability Dye eFluor™ 780 | ThermoFisher Scientific (Waltham, MA, USA) | 400 | |
| CD3 | UCHT1 | BV510 | BD Horizon™ | 100 | |
| CD56 | B159 | PE-Cy™ 5 | BD Pharmingen™ | 50 | |
| CD4 | RPA-T4 | BV421 | BD Horizon™ | 100 | |
| CD8 | RPA-T8 | BB515 | BD Horizon™ | 400 | |
| CD197 (CCR7) | 3D12 | PE | BD Pharmingen™ | 100 | |
| CD45RA | HI100 | Alexa Fluor® 700 | BD Pharmingen™ | 100 | |
| CD28 | CD28.2 | PerCP-Cy™ 5.5 | BD Pharmingen™ | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bröde, P.; Claus, M.; Gajewski, P.D.; Getzmann, S.; Wascher, E.; Watzl, C. From Immunosenescence to Aging Types—Establishing Reference Intervals for Immune Age Biomarkers by Centile Estimation. Int. J. Mol. Sci. 2023, 24, 13186. https://doi.org/10.3390/ijms241713186
Bröde P, Claus M, Gajewski PD, Getzmann S, Wascher E, Watzl C. From Immunosenescence to Aging Types—Establishing Reference Intervals for Immune Age Biomarkers by Centile Estimation. International Journal of Molecular Sciences. 2023; 24(17):13186. https://doi.org/10.3390/ijms241713186
Chicago/Turabian StyleBröde, Peter, Maren Claus, Patrick D. Gajewski, Stephan Getzmann, Edmund Wascher, and Carsten Watzl. 2023. "From Immunosenescence to Aging Types—Establishing Reference Intervals for Immune Age Biomarkers by Centile Estimation" International Journal of Molecular Sciences 24, no. 17: 13186. https://doi.org/10.3390/ijms241713186
APA StyleBröde, P., Claus, M., Gajewski, P. D., Getzmann, S., Wascher, E., & Watzl, C. (2023). From Immunosenescence to Aging Types—Establishing Reference Intervals for Immune Age Biomarkers by Centile Estimation. International Journal of Molecular Sciences, 24(17), 13186. https://doi.org/10.3390/ijms241713186