
Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation?



Abstract
1. Introduction
2. Cytokines
2.1. Interleukin-1 Beta
2.2. Soluble Interleukin-2 Receptor
2.3. Interleukin-3
2.4. Interleukin-4
2.5. Interleukin-5
2.6. Interleukin-6
2.7. Interleukin-8
2.8. Interleukin-9
2.9. Interleukin-10
2.10. Interleukin-12
2.11. Interleukin-13
2.12. Interleukin-17A
2.13. Interleukin-18
2.14. Interferon-Gamma
2.15. CCL2
2.16. CCL3
2.17. CCL5
2.18. CCL11
2.19. Tumor Necrosis Factor
2.20. Soluble TNF Receptor 2
2.21. Transforming Growth Factor-Beta
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Depression. Available online: https://www.who.int/news-room/fact-sheets/detail/depression (accessed on 29 November 2022).
- Mathers, C.D.; Loncar, D. Projections of Global Mortality and Burden of Disease from 2002 to 2030. PLoS Med. 2006, 3, e442. [Google Scholar] [CrossRef] [PubMed]
- Santomauro, D.F.; Herrera, A.M.M.; Shadid, J.; Zheng, P.; Ashbaugh, C.; Pigott, D.M.; Abbafati, C.; Adolph, C.; Amlag, J.O.; Aravkin, A.Y.; et al. Global Prevalence and Burden of Depressive and Anxiety Disorders in 204 Countries and Territories in 2020 Due to the COVID-19 Pandemic. Lancet 2021, 398, 1700–1712. [Google Scholar] [CrossRef] [PubMed]
- Kola, L.; Kohrt, B.A.; Hanlon, C.; Naslund, J.A.; Sikander, S.; Balaji, M.; Benjet, C.; Cheung, E.Y.L.; Eaton, J.; Gonsalves, P.; et al. COVID-19 Mental Health Impact and Responses in Low-Income and Middle-Income Countries: Reimagining Global Mental Health. Lancet Psychiatry 2021, 8, 535–550. [Google Scholar] [CrossRef] [PubMed]
- Bains, N.; Abdijadid, S. Major Depressive Disorder. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Nobis, A.; Zalewski, D.; Waszkiewicz, N. Peripheral Markers of Depression. J. Clin. Med. 2020, 9, 3793. [Google Scholar] [CrossRef]
- Lang, U.E.; Borgwardt, S. Molecular Mechanisms of Depression: Perspectives on New Treatment Strategies. Cell. Physiol. Biochem. 2013, 31, 761–777. [Google Scholar] [CrossRef]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and Depression: A Review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- MÉNARD, C.; HODES, G.E.; RUSSO, S.J. Pathogenesis of Depression: Insights from Human and Rodent Studies. Neuroscience 2016, 321, 138–162. [Google Scholar] [CrossRef]
- Li, Z.; Ruan, M.; Chen, J.; Fang, Y. Major Depressive Disorder: Advances in Neuroscience Research and Translational Applications. Neurosci. Bull. 2021, 37, 863–880. [Google Scholar] [CrossRef]
- Cuijpers, P.; Quero, S.; Dowrick, C.; Arroll, B. Psychological Treatment of Depression in Primary Care: Recent Developments. Curr. Psychiatry Rep. 2019, 21, 129. [Google Scholar] [CrossRef]
- Kverno, K.S.; Mangano, E. Treatment-Resistant Depression: Approaches to Treatment. J. Psychosoc. Nurs. Ment. Health Serv. 2021, 59, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Siwek, M.; Sowa-Kućma, M.; Dudek, D.; Styczeń, K.; Szewczyk, B.; Kotarska, K.; Misztak, P.; Pilc, A.; Wolak, M.; Nowak, G. Oxidative Stress Markers in Affective Disorders. Pharmacol. Rep. 2013, 65, 1558–1571. [Google Scholar] [CrossRef] [PubMed]
- Farooq, R.K.; Asghar, K.; Kanwal, S.; Zulqernain, A. Role of Inflammatory Cytokines in Depression: Focus on Interleukin-1β (Review). Biomed. Rep. 2017, 6, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wei, Y.B.; Strawbridge, R.; Bao, Y.; Chang, S.; Shi, L.; Que, J.; Gadad, B.S.; Trivedi, M.H.; Kelsoe, J.R.; et al. Peripheral Cytokine Levels and Response to Antidepressant Treatment in Depression: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2020, 25, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Haapakoski, R.; Mathieu, J.; Ebmeier, K.P.; Alenius, H.; Kivimäki, M. Cumulative Meta-Analysis of Interleukins 6 and 1β, Tumour Necrosis Factor α and C-Reactive Protein in Patients with Major Depressive Disorder. Brain Behav. Immun. 2015, 49, 206–215. [Google Scholar] [CrossRef]
- Miller, A.H.; Maletic, V.; Raison, C.L. Inflammation and Its Discontents: The Role of Cytokines in the Pathophysiology of Major Depression. Biol. Psychiatry 2009, 65, 732–741. [Google Scholar] [CrossRef]
- Blume, J.; Douglas, S.D.; Evans, D.L. Immune Suppression and Immune Activation in Depression. Brain Behav. Immun. 2011, 25, 221–229. [Google Scholar] [CrossRef]
- Maes, M.; Carvalho, A.F. The Compensatory Immune-Regulatory Reflex System (CIRS) in Depression and Bipolar Disorder. Mol. Neurobiol. 2018, 55, 8885–8903. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Derry, H.M.; Fagundes, C.P. Inflammation: Depression Fans the Flames and Feasts on the Heat. Am. J. Psychiatry 2015, 172, 1075–1091. [Google Scholar] [CrossRef]
- Hazeltine, D.B.; Polokowski, A.R.; Reigada, L.C. Inflammatory Cytokines, but Not Dietary Patterns, Are Related to Somatic Symptoms of Depression in a Sample of Women. Front. Psychiatry 2022, 13. [Google Scholar] [CrossRef]
- Miller, A.H. Mechanisms of Cytokine-Induced Behavioral Changes: Psychoneuroimmunology at the Translational Interface. Brain Behav. Immun. 2009, 23, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Müller, N.; Ackenheil, M. Psychoneuroimmunology and the Cytokine Action in the CNS: Implications for Psychiatric Disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 1998, 22, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.J.; Wang, J.; Ando, T. Effects of Cytokines on Cerebral Neurotransmission. Comparison with the Effects of Stress. Adv. Exp. Med. Biol. 1999, 461, 117–127. [Google Scholar] [CrossRef]
- Dantzer, R.; Wollman, E.; Vitkovic, L.; Yirmiya, R. Cytokines and Depression: Fortuitous or Causative Association? Mol. Psychiatry 1999, 4, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Licinio, J.; Wong, M.L. The Role of Inflammatory Mediators in the Biology of Major Depression: Central Nervous System Cytokines Modulate the Biological Substrate of Depressive Symptoms, Regulate Stress-Responsive Systems, and Contribute to Neurotoxicity and Neuroprotection. Mol. Psychiatry 1999, 4, 317–327. [Google Scholar] [CrossRef]
- Tsai, S.-J. Effects of Interleukin-1beta Polymorphisms on Brain Function and Behavior in Healthy and Psychiatric Disease Conditions. Cytokine Growth Factor Rev. 2017, 37, 89–97. [Google Scholar] [CrossRef]
- Simpson, R.J.; Hammacher, A.; Smith, D.K.; Matthews, J.M.; Ward, L.D. Interleukin-6: Structure-Function Relationships. Protein Sci. 1997, 6, 929–955. [Google Scholar] [CrossRef]
- Lotrich, F.E. Inflammatory Cytokine-Associated Depression. Brain Res. 2015, 1617, 113–125. [Google Scholar] [CrossRef]
- Beurel, E.; Lowell, J.A. Th17 Cells in Depression. Brain Behav. Immun. 2018, 69, 28–34. [Google Scholar] [CrossRef]
- Neupane, S.P.; Virtej, A.; Myhren, L.E.; Bull, V.H. Biomarkers Common for Inflammatory Periodontal Disease and Depression: A Systematic Review. Brain Behav. Immun. Health 2022, 21, 100450. [Google Scholar] [CrossRef]
- Yui, S.; Sasayama, D.; Yamaguchi, M.; Washizuka, S. Altered Levels of Salivary Cytokines in Patients with Major Depressive Disorder. Clin. Neurol. Neurosurg. 2022, 221, 107390. [Google Scholar] [CrossRef]
- Byrne, M.L.; O’Brien-Simpson, N.M.; Reynolds, E.C.; Walsh, K.A.; Laughton, K.; Waloszek, J.M.; Woods, M.J.; Trinder, J.; Allen, N.B. Corrigendum to “Acute Phase Protein and Cytokine Levels in Serum and Saliva: A Comparison of Detectable Levels and Correlations in a Depressed and Healthy Adolescent Sample” [Brain Behav. Immun. 34 (2013) 164–175]. Brain Behav. Immun. 2020, 89, 696–697. [Google Scholar] [CrossRef]
- Köhler, C.A.; Freitas, T.H.; Maes, M.; de Andrade, N.Q.; Liu, C.S.; Fernandes, B.S.; Stubbs, B.; Solmi, M.; Veronese, N.; Herrmann, N.; et al. Peripheral Cytokine and Chemokine Alterations in Depression: A Meta-Analysis of 82 Studies. Acta Psychiatr. Scand. 2017, 135, 373–387. [Google Scholar] [CrossRef] [PubMed]
- Hart, B.L. Biological Basis of the Behavior of Sick Animals. Neurosci. Biobehav. Rev. 1988, 12, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Brebner, K. Synergistic Effects of Interleukin-1β, Interleukin-6, and Tumor Necrosis Factor-α Central Monoamine, Corticosterone, and Behavioral Variations. Neuropsychopharmacology 2000, 22, 566–580. [Google Scholar] [CrossRef] [PubMed]
- Ng, A.; Tam, W.W.; Zhang, M.W.; Ho, C.S.; Husain, S.F.; McIntyre, R.S.; Ho, R.C. IL-1β, IL-6, TNF- α and CRP in Elderly Patients with Depression or Alzheimer’s Disease: Systematic Review and Meta-Analysis. Sci. Rep. 2018, 8, 12050. [Google Scholar] [CrossRef]
- Ellul, P.; Boyer, L.; Groc, L.; Leboyer, M.; Fond, G. Interleukin-1 β-Targeted Treatment Strategies in Inflammatory Depression: Toward Personalized Care. Acta Psychiatr. Scand. 2016, 134, 469–484. [Google Scholar] [CrossRef]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine Regulation of Brain Cytokines After Psychological Stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef]
- Fleshner, M.; Crane, C.R. Exosomes, DAMPs and MiRNA: Features of Stress Physiology and Immune Homeostasis. Trends Immunol. 2017, 38, 768–776. [Google Scholar] [CrossRef]
- Nguyen, K.T.; Deak, T.; Owens, S.M.; Kohno, T.; Fleshner, M.; Watkins, L.R.; Maier, S.F. Exposure to Acute Stress Induces Brain Interleukin-1β Protein in the Rat. J. Neurosci. 1998, 18, 2239–2246. [Google Scholar] [CrossRef]
- Levine, J.; Barak, Y.; Chengappa, K.N.R.; Rapoport, A.; Rebey, M.; Barak, V. Cerebrospinal Cytokine Levels in Patients with Acute Depression. Neuropsychobiology 1999, 40, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Thomas, A.J.; Davis, S.; Morris, C.; Jackson, E.; Harrison, R.; O’Brien, J.T. Increase in Interleukin-1β in Late-Life Depression. AJP 2005, 162, 175–177. [Google Scholar] [CrossRef] [PubMed]
- Corwin, E.J.; Johnston, N.; Pugh, L. Symptoms of Postpartum Depression Associated with Elevated Levels of Interleukin-1 Beta During the First Month Postpartum. Biol. Res. Nurs. 2008, 10, 128–133. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, A.M.; Leal, B.; Ferreira, I.; Brás, S.; Moreira, I.; Samões, R.; Sousa, A.P.; Santos, E.; Silva, B.; Costa, P.P.; et al. Depression and Anxiety in Multiple Sclerosis Patients: The Role of Genetic Variability of Interleukin 1β. Mult. Scler. Relat. Disord. 2021, 52, 102982. [Google Scholar] [CrossRef]
- Ovaskainen, Y.; Koponen, H.; Jokelainen, J.; Keinänen-Kiukaanniemi, S.; Kumpusalo, E.; Vanhala, M. Depressive Symptomatology Is Associated with Decreased Interleukin-1 Beta and Increased Interleukin-1 Receptor Antagonist Levels in Males. Psychiatry Res. 2009, 167, 73–79. [Google Scholar] [CrossRef]
- Seil, M.; Ouaaliti, M.E.; Abdou Foumekoye, S.; Pochet, S.; Dehaye, J. Distinct Regulation by Lipopolysaccharides of the Expression of Interleukin-1β by Murine Macrophages and Salivary Glands. Innate Immun. 2012, 18, 14–24. [Google Scholar] [CrossRef]
- Clark, A.K.; Staniland, A.A.; Marchand, F.; Kaan, T.K.Y.; McMahon, S.B.; Malcangio, M. P2X7-Dependent Release of Interleukin-1 and Nociception in the Spinal Cord Following Lipopolysaccharide. J. Neurosci. 2010, 30, 573–582. [Google Scholar] [CrossRef]
- Porterfield, V.M.; Gabella, K.M.; Simmons, M.A.; Johnson, J.D. Repeated Stressor Exposure Regionally Enhances Beta-Adrenergic Receptor-Mediated Brain IL-1β Production. Brain Behav. Immun. 2012, 26, 1249–1255. [Google Scholar] [CrossRef]
- Barnard, D.F.; Gabella, K.M.; Kulp, A.C.; Parker, A.D.; Dugan, P.B.; Johnson, J.D. Sex Differences in the Regulation of Brain IL-1β in Response to Chronic Stress. Psychoneuroendocrinology 2019, 103, 203–211. [Google Scholar] [CrossRef]
- Barrientos, R.M.; Sprunger, D.B.; Campeau, S.; Higgins, E.A.; Watkins, L.R.; Rudy, J.W.; Maier, S.F. Brain-Derived Neurotrophic Factor MRNA Downregulation Produced by Social Isolation Is Blocked by Intrahippocampal Interleukin-1 Receptor Antagonist. Neuroscience 2003, 121, 847–853. [Google Scholar] [CrossRef]
- Schmid, A.W.; Lynch, M.A.; Herron, C.E. The Effects of IL-1 Receptor Antagonist on Beta Amyloid Mediated Depression of LTP in the Rat CA1 in Vivo. Hippocampus 2009, 19, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, C.; Numakawa, T.; Odaka, H.; Ooshima, Y.; Kiyama, Y.; Manabe, T.; Kunugi, H.; Iwakura, Y. IL-1 Receptor-Antagonist (IL-1Ra) Knockout Mice Show Anxiety-like Behavior by Aging. Neurosci. Lett. 2015, 599, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, S.D.; Mazzon, E.; Basile, M.S.; Cavalli, E.; Bramanti, P.; Nania, R.; Fagone, P.; Nicoletti, F.; Petralia, M.C. Upregulation of IL-1 Receptor Antagonist in a Mouse Model of Migraine. Brain Sci. 2019, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Euteneuer, F.; Schwarz, M.J.; Dannehl, K.; Hartung, A.; Westermann, S.; Rief, W. Increased Soluble Interleukin-2 Receptor Levels Are Related to Somatic but Not to Cognitive-Affective Features in Major Depression. Brain Behav. Immun. 2012, 26, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J. The IL-2-IL-2 Receptor Pathway in Health and Disease: The Role of the Soluble IL-2 Receptor. Clin. Immunol. 2020, 218, 108515. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Bosmans, E.; Suy, E.; Vandervorst, C.; DeJonckheere, C.; Raus, J. Depression-Related Disturbances in Mitogen-Induced Lymphocyte Responses and Interleukin-1 Beta and Soluble Interleukin-2 Receptor Production. Acta Psychiatr. Scand. 1991, 84, 379–386. [Google Scholar] [CrossRef]
- Al-Hakeim, H.K.; Al-Rammahi, D.A.; Al-Dujaili, A.H. IL-6, IL-18, SIL-2R, and TNFα Proinflammatory Markers in Depression and Schizophrenia Patients Who Are Free of Overt Inflammation. J. Affect. Disord. 2015, 182, 106–114. [Google Scholar] [CrossRef]
- Liu, Y.; Ho, R.C.-M.; Mak, A. Interleukin (IL)-6, Tumour Necrosis Factor Alpha (TNF-α) and Soluble Interleukin-2 Receptors (SIL-2R) Are Elevated in Patients with Major Depressive Disorder: A Meta-Analysis and Meta-Regression. J. Affect. Disord. 2012, 139, 230–239. [Google Scholar] [CrossRef]
- IL3 Interleukin 3 [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3562 (accessed on 18 December 2022).
- Mangi, M.H.; Newland, A.C. Interleukin-3: Promises and Perspectives. Hematology 1998, 3, 55–66. [Google Scholar] [CrossRef]
- Xiu, M.H.; Lin, C.G.; Tian, L.; Tan, Y.L.; Chen, J.; Chen, S.; Tan, S.P.; Wang, Z.R.; Yang, F.D.; Chen, D.C.; et al. Increased IL-3 Serum Levels in Chronic Patients with Schizophrenia: Associated with Psychopathology. Psychiatry Res. 2015, 229, 225–229. [Google Scholar] [CrossRef]
- Gao, W.; Xu, Y.; Liang, J.; Sun, Y.; Zhang, Y.; Shan, F.; Ge, J.; Xia, Q. Comparison of Serum Cytokines Levels in Normal-Weight and Overweight Patients with First-Episode Drug-Naïve Major Depressive Disorder. Front. Endocrinol. (Lausanne) 2022, 13, 1048337. [Google Scholar] [CrossRef] [PubMed]
- Osimo, E.F.; Pillinger, T.; Rodriguez, I.M.; Khandaker, G.M.; Pariante, C.M.; Howes, O.D. Inflammatory Markers in Depression: A Meta-Analysis of Mean Differences and Variability in 5,166 Patients and 5,083 Controls. Brain Behav. Immun. 2020, 87, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Qurashi, T.A.; Bhat, G.A.; Khan, M.S.; Rasool, R.; Sameen, F.; Hassan, I.; Mudassar, S. Interleukin 4 and Interleukin 4 Receptor Alpha Gene Variants and Risk of Atopy-A Case Control Study Based Assessment. Clin. Immunol. 2021, 229, 108783. [Google Scholar] [CrossRef]
- Li, J.; Leschka, S.; Rutschow, S.; Schwimmbeck, P.L.; Husmann, L.; Noutsias, M.; Westermann, D.; Poller, W.; Zeichhardt, H.; Klingel, K.; et al. Immunomodulation by Interleukin-4 Suppresses Matrix Metalloproteinases and Improves Cardiac Function in Murine Myocarditis. Eur. J. Pharmacol. 2007, 554, 60–68. [Google Scholar] [CrossRef]
- Quarta, A.; Berneman, Z.; Ponsaerts, P. Neuroprotective Modulation of Microglia Effector Functions Following Priming with Interleukin 4 and 13: Current Limitations in Understanding Their Mode-of-Action. Brain Behav. Immun. 2020, 88, 856–866. [Google Scholar] [CrossRef]
- Zhang, J.; Rong, P.; Zhang, L.; He, H.; Zhou, T.; Fan, Y.; Mo, L.; Zhao, Q.; Han, Y.; Li, S.; et al. IL4-Driven Microglia Modulate Stress Resilience through BDNF-Dependent Neurogenesis. Sci. Adv. 2021, 7, eabb9888. [Google Scholar] [CrossRef] [PubMed]
- Wachholz, S.; Knorr, A.; Mengert, L.; Plümper, J.; Sommer, R.; Juckel, G.; Friebe, A. Interleukin-4 Is a Participant in the Regulation of Depressive-like Behavior. Behav. Brain Res. 2017, 326, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Chou, Y.-H.; Hsieh, W.-C.; Chen, L.-C.; Lirng, J.-F.; Wang, S.-J. Association between the Serotonin Transporter and Cytokines: Implications for the Pathophysiology of Bipolar Disorder. J. Affect. Disord. 2016, 191, 29–35. [Google Scholar] [CrossRef]
- Wachholz, S.; Eßlinger, M.; Manitz, M.-P.; Sommer, R.; Plümper, J.; Juckel, G.; Friebe, A. Interleukin-4−/− Mice Show a Depression-like Phenotype. Neurol. Psychiatry Brain Res. 2016, 22, 24. [Google Scholar] [CrossRef]
- Schwenger, G.T.; Sanderson, C.J. New Directions in Understanding Interleukin-5 Gene Expression. Leuk. Lymphoma 1998, 28, 443–450. [Google Scholar] [CrossRef]
- Elomaa, A.-P.; Niskanen, L.; Herzig, K.-H.; Viinamäki, H.; Hintikka, J.; Koivumaa-Honkanen, H.; Honkalampi, K.; Valkonen-Korhonen, M.; Harvima, I.T.; Lehto, S.M. Elevated Levels of Serum IL-5 Are Associated with an Increased Likelihood of Major Depressive Disorder. BMC Psychiatry 2012, 12, 2. [Google Scholar] [CrossRef] [PubMed]
- Sawada, M.; Suzumura, A.; Itoh, Y.; Marunouchi, T. Production of Interleukin-5 by Mouse Astrocytes and Microglia in Culture. Neurosci. Lett. 1993, 155, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Ringheim, G.E. Mitogenic Effects of Interleukin-5 on Microglia. Neurosci. Lett. 1995, 201, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.-Y.; Chin-Hung Chen, V.; Tzang, B.-S.; Hsieh, C.-C.; Wang, W.-K.; Weng, Y.-P.; Hsu, Y.-T.; Hsaio, H.-P.; Weng, J.-C.; Chen, Y.-L. Circulating Cytokines as Predictors of Depression in Patients with Breast Cancer. J. Psychiatr. Res. 2021, 136, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Zhou, Q.; Sun, Y.; Niu, W.; Du, J. Association of Three Single Nucleotide Polymorphisms in Interleukin 6 Gene with Risk of Chronic Obstructive Pulmonary Disease. Gene 2022, 828, 146467. [Google Scholar] [CrossRef]
- Zadka, Ł.; Dzięgiel, P.; Kulus, M.; Olajossy, M. Clinical Phenotype of Depression Affects Interleukin-6 Synthesis. J. Interferon Cytokine Res. 2017, 37, 231–245. [Google Scholar] [CrossRef]
- Li, Y.; Xie, Y.; Xu, Y.; Xian, X.; Wang, R.; Cai, L.; Li, G.; Li, Y. Interleukin-6-White Matter Network Differences Explained the Susceptibility to Depression after Stressful Life Events. J. Affect. Disord. 2022, 305, 122–132. [Google Scholar] [CrossRef]
- Hodes, G.E.; Ménard, C.; Russo, S.J. Integrating Interleukin-6 into Depression Diagnosis and Treatment. Neurobiol. Stress 2016, 4, 15–22. [Google Scholar] [CrossRef]
- Manfro, P.H.; Anselmi, L.; Barros, F.; Gonçalves, H.; Murray, J.; Oliveira, I.O.; Tovo-Rodrigues, L.; Wehrmeister, F.C.; Menezes, A.M.B.; Mondelli, V.; et al. Youth Depression and Inflammation: Cross-Sectional Network Analyses of C-Reactive Protein, Interleukin-6 and Symptoms in a Population-Based Sample. J. Psychiatr. Res. 2022, 150, 197–201. [Google Scholar] [CrossRef]
- Young, J.J.; Bruno, D.; Pomara, N. A Review of the Relationship between Proinflammatory Cytokines and Major Depressive Disorder. J. Affect. Disord. 2014, 169, 15–20. [Google Scholar] [CrossRef]
- Wang, P.; Feng, Y.; Wang, L.; Li, Y.; Fan, C.; Song, Q.; Yu, S.Y. Interleukin-6: Its Role and Mechanisms in Rescuing Depression-like Behaviors in Rat Models of Depression. Brain Behav. Immun. 2019, 82, 106–121. [Google Scholar] [CrossRef] [PubMed]
- Schumertl, T.; Lokau, J.; Rose-John, S.; Garbers, C. Function and Proteolytic Generation of the Soluble Interleukin-6 Receptor in Health and Disease. Biochim. Biophys. Acta Mol. Cell Res. 2022, 1869, 119143. [Google Scholar] [CrossRef] [PubMed]
- García-Juárez, M.; Camacho-Morales, A. Defining the Role of Anti- and Pro-Inflammatory Outcomes of Interleukin-6 in Mental Health. Neuroscience 2022, 492, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Ushiroyama, T.; Ikeda, A.; Ueki, M. Elevated Plasma Interleukin-6 (IL-6) and Soluble IL-6 Receptor Concentrations in Menopausal Women with and without Depression. Int. J. Gynaecol. Obs. 2002, 79, 51–52. [Google Scholar] [CrossRef] [PubMed]
- Kelly, K.M.; Smith, J.A.; Mezuk, B. Depression and Interleukin-6 Signaling: A Mendelian Randomization Study. Brain Behav. Immun. 2021, 95, 106–114. [Google Scholar] [CrossRef]
- Maes, M.; Meltzer, H.Y.; Bosmans, E.; Bergmans, R.; Vandoolaeghe, E.; Ranjan, R.; Desnyder, R. Increased Plasma Concentrations of Interleukin-6, Soluble Interleukin-6, Soluble Interleukin-2 and Transferrin Receptor in Major Depression. J. Affect. Disord. 1995, 34, 301–309. [Google Scholar] [CrossRef]
- Tsai, S.-J. Role of Interleukin 8 in Depression and Other Psychiatric Disorders. Prog. Neuropsychopharmacol. Biol. Psychiatry 2021, 106, 110173. [Google Scholar] [CrossRef]
- Kruse, J.L.; Olmstead, R.; Hellemann, G.; Breen, E.C.; Tye, S.J.; Brooks, J.O.; Wade, B.; Congdon, E.; Espinoza, R.; Narr, K.L.; et al. Interleukin-8 and Lower Severity of Depression in Females, but Not Males, with Treatment-Resistant Depression. J. Psychiatr. Res. 2021, 140, 350–356. [Google Scholar] [CrossRef]
- Kruse, J.L.; Vasavada, M.M.; Olmstead, R.; Hellemann, G.; Wade, B.; Breen, E.C.; Brooks, J.O.; Congdon, E.; Espinoza, R.; Narr, K.L.; et al. Depression Treatment Response to Ketamine: Sex-Specific Role of Interleukin-8, but Not Other Inflammatory Markers. Transl. Psychiatry 2021, 11, 167. [Google Scholar] [CrossRef]
- Kruse, J.L.; Olmstead, R.; Hellemann, G.; Wade, B.; Jiang, J.; Vasavada, M.M.; Brooks Iii, J.O.; Congdon, E.; Espinoza, R.; Narr, K.L.; et al. Inflammation and Depression Treatment Response to Electroconvulsive Therapy: Sex-Specific Role of Interleukin-8. Brain Behav. Immun. 2020, 89, 59–66. [Google Scholar] [CrossRef]
- Irwin, M.R.; Olmstead, R.; Kruse, J.; Breen, E.C.; Haque, R. Association of Interleukin-8 and Risk of Incident and Recurrent Depression in Long-Term Breast Cancer Survivors. Brain Behav. Immun. 2022, 105, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Skoog, I.; Börjesson-Hanson, A.; Blennow, K.; Zetterberg, H.; Ostling, S.; Kern, J.; Gudmundsson, P.; Marlow, T.; Rosengren, L.; et al. Higher CSF Interleukin-6 and CSF Interleukin-8 in Current Depression in Older Women. Results from a Population-Based Sample. Brain Behav. Immun. 2014, 41, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Kuzior, H.; Fiebich, B.L.; Yousif, N.M.; Saliba, S.W.; Ziegler, C.; Nickel, K.; Maier, S.J.; Süß, P.; Runge, K.; Matysik, M.; et al. Increased IL-8 Concentrations in the Cerebrospinal Fluid of Patients with Unipolar Depression. Compr. Psychiatry 2020, 102, 152196. [Google Scholar] [CrossRef]
- Kruse, J.L.; Boyle, C.C.; Olmstead, R.; Breen, E.C.; Tye, S.J.; Eisenberger, N.I.; Irwin, M.R. Interleukin-8 and Depressive Responses to an Inflammatory Challenge: Secondary Analysis of a Randomized Controlled Trial. Sci. Rep. 2022, 12, 12627. [Google Scholar] [CrossRef] [PubMed]
- Sajjad, M.U.; Blennow, K.; Knapskog, A.B.; Idland, A.-V.; Chaudhry, F.A.; Wyller, T.B.; Zetterberg, H.; Watne, L.O. Cerebrospinal Fluid Levels of Interleukin-8 in Delirium, Dementia, and Cognitively Healthy Patients. J. Alzheimers Dis. 2020, 73, 1363–1372. [Google Scholar] [CrossRef]
- Chakraborty, S.; Kubatzky, K.F.; Mitra, D.K. An Update on Interleukin-9: From Its Cellular Source and Signal Transduction to Its Role in Immunopathogenesis. Int. J. Mol. Sci. 2019, 20, 2113. [Google Scholar] [CrossRef]
- Donninelli, G.; Saraf-Sinik, I.; Mazziotti, V.; Capone, A.; Grasso, M.G.; Battistini, L.; Reynolds, R.; Magliozzi, R.; Volpe, E. Interleukin-9 Regulates Macrophage Activation in the Progressive Multiple Sclerosis Brain. J. Neuroinflamm. 2020, 17, 149. [Google Scholar] [CrossRef]
- Shelton, R.C.; Claiborne, J.; Sidoryk-Wegrzynowicz, M.; Reddy, R.; Aschner, M.; Lewis, D.A.; Mirnics, K. Altered Expression of Genes Involved in Inflammation and Apoptosis in Frontal Cortex in Major Depression. Mol. Psychiatry 2011, 16, 751–762. [Google Scholar] [CrossRef]
- Karlsson, L.; Nousiainen, N.; Scheinin, N.M.; Maksimow, M.; Salmi, M.; Lehto, S.M.; Tolvanen, M.; Lukkarinen, H.; Karlsson, H. Cytokine Profile and Maternal Depression and Anxiety Symptoms in Mid-Pregnancy—The FinnBrain Birth Cohort Study. Arch. Womens Ment. Health 2017, 20, 39–48. [Google Scholar] [CrossRef]
- Varshney, P.; Parveen, R.; Khan, M.A.; Kohli, S.; Agarwal, N.B. Increased Serum Interleukin-9 and Interleukin-1β Are Associated with Depression in Type 2 Diabetes Patients. Arq. Neuro-Psiquiatr. 2020, 78, 255–261. [Google Scholar] [CrossRef]
- Saraiva, M.; O’Garra, A. The Regulation of IL-10 Production by Immune Cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed]
- Yadav, U.; Kumar, P.; Rai, V. Interleukin-10 (IL-10) Gene Polymorphisms and Prostate Cancer Susceptibility: Evidence from a Meta-Analysis. Gene Rep. 2021, 25, 101377. [Google Scholar] [CrossRef]
- Chi, C.-H.; Huang, Y.-Y.; Ye, S.-Z.; Shao, M.-M.; Jiang, M.-X.; Yang, M.-Y.; Wu, Q.; Shao, B.; Li, X.-M. Interleukin-10 Level Is Associated with Post-Stroke Depression in Acute Ischaemic Stroke Patients. J. Affect. Disord. 2021, 293, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Wiener, C.D.; Moreira, F.P.; Portela, L.V.; Strogulski, N.R.; Lara, D.R.; da Silva, R.A.; Souza, L.D.d.M.; Jansen, K.; Oses, J.P. Interleukin-6 and Interleukin-10 in Mood Disorders: A Population-Based Study. Psychiatry Res. 2019, 273, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Al-Fadhel, S.Z.; Al-Hakeim, H.K.; Al-Dujaili, A.H.; Maes, M. IL-10 Is Associated with Increased Mu-Opioid Receptor Levels in Major Depressive Disorder. Eur. Psychiatry 2019, 57, 46–51. [Google Scholar] [CrossRef]
- Roque, S.; Correia-Neves, M.; Mesquita, A.R.; Palha, J.A.; Sousa, N. Interleukin-10: A Key Cytokine in Depression? Cardiovasc. Psychiatry Neurol. 2009, 2009, 187894. [Google Scholar] [CrossRef]
- Kubera, M.; Maes, M.; Holan, V.; Basta-Kaim, A.; Roman, A.; Shani, J. Prolonged Desipramine Treatment Increases the Production of Interleukin-10, an Anti-Inflammatory Cytokine, in C57BL/6 Mice Subjected to the Chronic Mild Stress Model of Depression. J. Affect. Disord. 2001, 63, 171–178. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Wang, Y.; He, Y.; Wang, T.; Huang, X.-H.; Zhao, C.-M.; Zhang, L.; Li, S.-W.; Wang, C.; Qu, Y.-N.; et al. A1 Astrocytes Contribute to Murine Depression-like Behavior and Cognitive Dysfunction, Which Can Be Alleviated by IL-10 or Fluorocitrate Treatment. J. Neuroinflamm. 2020, 17, 200. [Google Scholar] [CrossRef]
- IL12A Interleukin 12A [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3592 (accessed on 30 November 2022).
- IL12B Interleukin 12B [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3593 (accessed on 30 November 2022).
- Liu, J.; Cao, S.; Kim, S.; Chung, E.Y.; Homma, Y.; Guan, X.; Jimenez, V.; Ma, X. Interleukin-12: An Update on Its Immunological Activities, Signaling and Regulation of Gene Expression. Curr. Immunol. Rev. 2005, 1, 119–137. [Google Scholar] [CrossRef]
- Quinones, M.; Ahuja, S.K.; Melby, P.C.; Pate, L.; Reddick, R.L.; Ahuja, S.S. Preformed Membrane-Associated Stores of Interleukin (IL)-12 Are a Previously Unrecognized Source of Bioactive IL-12 That Is Mobilized within Minutes of Contact with an Intracellular Parasite. J. Exp. Med. 2000, 192, 507–516. [Google Scholar] [CrossRef]
- Peluso, I.; Pallone, F.; Monteleone, G. Interleukin-12 and Th1 Immune Response in Crohn’s Disease: Pathogenetic Relevance and Therapeutic Implication. World J. Gastroenterol. 2006, 12, 5606–5610. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K. Plasma Increase of Interleukin-12 and Interferon-Gamma. Pathological Significance in Autism. J. Neuroimmunol. 1996, 66, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-K.; Suh, I.-B.; Kim, H.; Han, C.-S.; Lim, C.-S.; Choi, S.-H.; Licinio, J. The Plasma Levels of Interleukin-12 in Schizophrenia, Major Depression, and Bipolar Mania: Effects of Psychotropic Drugs. Mol. Psychiatry 2002, 7, 1107–1114. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-M.; Kim, Y.-K. The Role of IL-12 and TGF-Beta1 in the Pathophysiology of Major Depressive Disorder. Int. Immunopharmacol. 2006, 6, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Sutcigil, L.; Oktenli, C.; Musabak, U.; Bozkurt, A.; Cansever, A.; Uzun, O.; Sanisoglu, S.Y.; Yesilova, Z.; Ozmenler, N.; Ozsahin, A.; et al. Pro- and Anti-Inflammatory Cytokine Balance in Major Depression: Effect of Sertraline Therapy. Clin. Dev. Immunol. 2007, 2007, 76396. [Google Scholar] [CrossRef]
- Nowak, W.; Grendas, L.N.; Sanmarco, L.M.; Estecho, I.G.; Arena, Á.R.; Eberhardt, N.; Rodante, D.E.; Aoki, M.P.; Daray, F.M.; Carrera Silva, E.A.; et al. Pro-Inflammatory Monocyte Profile in Patients with Major Depressive Disorder and Suicide Behaviour and How Ketamine Induces Anti-Inflammatory M2 Macrophages by NMDAR and MTOR. EBioMedicine 2019, 50, 290–305. [Google Scholar] [CrossRef]
- El-Tantawy, A.M.; El-Sayed, A.E.; Kora, B.A.; Amin, R.T. Psychiatric Morbidity Associated with Some Cytokines (IL-1beta, IL-12, IL-18 and TNF-Alpha) among Rheumatoid Arthritis Patients. Egypt J. Immunol. 2008, 15, 1–11. [Google Scholar]
- Schmidt, F.M.; Lichtblau, N.; Minkwitz, J.; Chittka, T.; Thormann, J.; Kirkby, K.C.; Sander, C.; Mergl, R.; Faßhauer, M.; Stumvoll, M.; et al. Cytokine Levels in Depressed and Non-Depressed Subjects, and Masking Effects of Obesity. J. Psychiatr. Res. 2014, 55, 29–34. [Google Scholar] [CrossRef]
- Hou, R.; Ye, G.; Liu, Y.; Chen, X.; Pan, M.; Zhu, F.; Fu, J.; Fu, T.; Liu, Q.; Gao, Z.; et al. Effects of SSRIs on Peripheral Inflammatory Cytokines in Patients with Generalized Anxiety Disorder. Brain Behav. Immun. 2019, 81, 105–110. [Google Scholar] [CrossRef]
- IL13 Interleukin 13 [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3596 (accessed on 30 November 2022).
- Minty, A.; Chalon, P.; Derocq, J.M.; Dumont, X.; Guillemot, J.C.; Kaghad, M.; Labit, C.; Leplatois, P.; Liauzun, P.; Miloux, B. Interleukin-13 Is a New Human Lymphokine Regulating Inflammatory and Immune Responses. Nature 1993, 362, 248–250. [Google Scholar] [CrossRef]
- Kolosowska, N.; Keuters, M.H.; Wojciechowski, S.; Keksa-Goldsteine, V.; Laine, M.; Malm, T.; Goldsteins, G.; Koistinaho, J.; Dhungana, H. Peripheral Administration of IL-13 Induces Anti-Inflammatory Microglial/Macrophage Responses and Provides Neuroprotection in Ischemic Stroke. Neurotherapeutics 2019, 16, 1304–1319. [Google Scholar] [CrossRef] [PubMed]
- Khalil, S.M.; Bernstein, I.; Kulaga, H.; Gour, N.; Rowan, N.; Lajoie, S.; Lane, A.P. Interleukin 13 (IL-13) Alters Hypoxia-Associated Genes and Upregulates CD73. Int. Forum Allergy Rhinol. 2020, 10, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Doherty, T.M.; Kastelein, R.; Menon, S.; Andrade, S.; Coffman, R.L. Modulation of Murine Macrophage Function by IL-13. J. Immunol. 1993, 151, 7151–7160. [Google Scholar] [PubMed]
- Corren, J. Role of Interleukin-13 in Asthma. Curr. Allergy Asthma Rep. 2013, 13, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Iwaszko, M.; Biały, S.; Bogunia-Kubik, K. Significance of Interleukin (IL)-4 and IL-13 in Inflammatory Arthritis. Cells 2021, 10, 3000. [Google Scholar] [CrossRef] [PubMed]
- Pavón, L.; Sandoval-López, G.; Eugenia Hernández, M.; Loría, F.; Estrada, I.; Pérez, M.; Moreno, J.; Avila, U.; Leff, P.; Antón, B.; et al. Th2 Cytokine Response in Major Depressive Disorder Patients before Treatment. J. Neuroimmunol. 2006, 172, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Matalka, K.Z. Neuroendocrine and Cytokines-Induced Responses to Minutes, Hours, and Days of Mental Stress. Neuro Endocrinol. Lett. 2003, 24, 283–292. [Google Scholar]
- Hernández, M.E.; Mendieta, D.; Martínez-Fong, D.; Loría, F.; Moreno, J.; Estrada, I.; Bojalil, R.; Pavón, L. Variations in Circulating Cytokine Levels during 52 Week Course of Treatment with SSRI for Major Depressive Disorder. Eur. Neuropsychopharmacol. 2008, 18, 917–924. [Google Scholar] [CrossRef]
- Wong, M.-L.; Dong, C.; Maestre-Mesa, J.; Licinio, J. Polymorphisms in Inflammation-Related Genes Are Associated with Susceptibility to Major Depression and Antidepressant Response. Mol. Psychiatry 2008, 13, 800–812. [Google Scholar] [CrossRef]
- Czysz, A.H.; Mason, B.L.; Li, Q.; Chin-Fatt, C.; Minhajuddin, A.; Carmody, T.; Trivedi, M.H. Comparison of Inflammatory Markers as Moderators of Depression Outcomes: A CO-MED Study. J. Affect. Disord. 2021, 295, 1066–1071. [Google Scholar] [CrossRef]
- Vai, B.; Mazza, M.G.; Cazzetta, S.; Calesella, F.; Aggio, V.; Lorenzi, C.; Zanardi, R.; Poletti, S.; Colombo, C.; Benedetti, F. Higher Interleukin 13 Differentiates Patients with a Positive History of Suicide Attempts in Major Depressive Disorder. J. Affect. Disord. Rep. 2021, 6, 100254. [Google Scholar] [CrossRef]
- Timonen, M.; Jokelainen, J.; Hakko, H.; Silvennoinen-Kassinen, S.; Meyer-Rochow, V.B.; Herva, A.; Räsänen, P. Atopy and Depression: Results from the Northern Finland 1966 Birth Cohort Study. Mol. Psychiatry 2003, 8, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Timonen, M.; Viilo, K.; Hakko, H.; Särkioja, T.; Meyer-Rochow, V.B.; Väisänen, E.; Räsänen, P. Is Seasonality of Suicides Stronger in Victims with Hospital-Treated Atopic Disorders? Psychiatry Res. 2004, 126, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Tonelli, L.H.; Stiller, J.; Rujescu, D.; Giegling, I.; Schneider, B.; Maurer, K.; Schnabel, A.; Möller, H.-J.; Chen, H.H.; Postolache, T.T. Elevated Cytokine Expression in the Orbitofrontal Cortex of Victims of Suicide. Acta Psychiatr. Scand. 2008, 117, 198–206. [Google Scholar] [CrossRef]
- IL17A Interleukin 17A [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3605 (accessed on 13 December 2022).
- Rouvier, E.; Luciani, M.F.; Mattéi, M.G.; Denizot, F.; Golstein, P. CTLA-8, Cloned from an Activated T Cell, Bearing AU-Rich Messenger RNA Instability Sequences, and Homologous to a Herpesvirus Saimiri Gene. J. Immunol. 1993, 150, 5445–5456. [Google Scholar] [PubMed]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Matusevicius, D.; Kivisäkk, P.; He, B.; Kostulas, N.; Ozenci, V.; Fredrikson, S.; Link, H. Interleukin-17 MRNA Expression in Blood and CSF Mononuclear Cells Is Augmented in Multiple Sclerosis. Mult. Scler. 1999, 5, 101–104. [Google Scholar] [CrossRef] [PubMed]
- Nothdurfter, C.; Milenkovic, V.M.; Sarubin, N.; Hilbert, S.; Manook, A.; Weigl, J.; Almeqbaali, K.; Wetzel, C.H.; Rupprecht, R.; Baghai, T.C. The Cytokine IL-17A as a Marker of Treatment Resistance in Major Depressive Disorder? Eur. J. Neurosci. 2021, 53, 172–182. [Google Scholar] [CrossRef]
- Medina-Rodriguez, E.M.; Madorma, D.; O’Connor, G.; Mason, B.L.; Han, D.; Deo, S.K.; Oppenheimer, M.; Nemeroff, C.B.; Trivedi, M.H.; Daunert, S.; et al. Identification of a Signaling Mechanism by Which the Microbiome Regulates Th17 Cell-Mediated Depressive-Like Behaviors in Mice. Am. J. Psychiatry 2020, 177, 974–990. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.A.; Gómez-Lahoz, A.M.; Orozco, A.; Lahera, G.; Diaz, D.; Ortega, M.A.; Albillos, A.; Quintero, J.; Aubá, E.; Monserrat, J.; et al. Expansion of CD4 T Lymphocytes Expressing Interleukin 17 and Tumor Necrosis Factor in Patients with Major Depressive Disorder. J. Pers. Med. 2021, 11, 220. [Google Scholar] [CrossRef]
- Min, Z.; Li, Y.; Ying, H. Blood T-Helper 17 Cells and Interleukin-17A Correlate with the Elevated Risk of Postpartum Depression and Anxiety. J. Clin. Lab. Anal. 2022, 36, e24559. [Google Scholar] [CrossRef] [PubMed]
- IL18 Interleukin 18 [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/3606 (accessed on 30 November 2022).
- Nakamura, K.; Okamura, H.; Wada, M.; Nagata, K.; Tamura, T. Endotoxin-Induced Serum Factor That Stimulates Gamma Interferon Production. Infect. Immun. 1989, 57, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Felderhoff-Mueser, U.; Schmidt, O.I.; Oberholzer, A.; Bührer, C.; Stahel, P.F. IL-18: A Key Player in Neuroinflammation and Neurodegeneration? Trends Neurosci. 2005, 28, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Zheng, D.; Liwinski, T.; Elinav, E. Inflammasome Activation and Regulation: Toward a Better Understanding of Complex Mechanisms. Cell Discov. 2020, 6, 36. [Google Scholar] [CrossRef]
- Wong, M.-L.; Inserra, A.; Lewis, M.D.; Mastronardi, C.A.; Leong, L.; Choo, J.; Kentish, S.; Xie, P.; Morrison, M.; Wesselingh, S.L.; et al. Inflammasome Signaling Affects Anxiety- and Depressive-like Behavior and Gut Microbiome Composition. Mol. Psychiatry 2016, 21, 797–805. [Google Scholar] [CrossRef]
- Song, A.-Q.; Gao, B.; Fan, J.-J.; Zhu, Y.-J.; Zhou, J.; Wang, Y.-L.; Xu, L.-Z.; Wu, W.-N. NLRP1 Inflammasome Contributes to Chronic Stress-Induced Depressive-like Behaviors in Mice. J. Neuroinflamm. 2020, 17, 178. [Google Scholar] [CrossRef]
- Fan, N.; Luo, Y.; Ou, Y.; He, H. Altered Serum Levels of TNF-α, IL-6, and IL-18 in Depressive Disorder Patients. Hum. Psychopharmacol. 2017, 32, e2588. [Google Scholar] [CrossRef]
- Kokai, M.; Kashiwamura, S.; Okamura, H.; Ohara, K.; Morita, Y. Plasma Interleukin-18 Levels in Patients with Psychiatric Disorders. J. Immunother. 2002, 25 (Suppl. S1), S68–S71. [Google Scholar] [CrossRef]
- Alcocer-Gómez, E.; de Miguel, M.; Casas-Barquero, N.; Núñez-Vasco, J.; Sánchez-Alcazar, J.A.; Fernández-Rodríguez, A.; Cordero, M.D. NLRP3 Inflammasome Is Activated in Mononuclear Blood Cells from Patients with Major Depressive Disorder. Brain Behav. Immun. 2014, 36, 111–117. [Google Scholar] [CrossRef]
- Prossin, A.R.; Koch, A.E.; Campbell, P.L.; McInnis, M.G.; Zalcman, S.S.; Zubieta, J.-K. Association of Plasma Interleukin-18 Levels with Emotion Regulation and μ-Opioid Neurotransmitter Function in Major Depression and Healthy Volunteers. Biol. Psychiatry 2011, 69, 808–812. [Google Scholar] [CrossRef]
- Witek-Janusek, L.; Albuquerque, K.; Chroniak, K.R.; Chroniak, C.; Durazo, R.; Mathews, H.L. Effect of Mindfulness Based Stress Reduction on Immune Function, Quality of Life and Coping In Women Newly Diagnosed with Early Stage Breast Cancer. Brain Behav. Immun. 2008, 22, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.L.; Farrar, W.B.; Golden-Kreutz, D.; Kutz, L.A.; MacCallum, R.; Courtney, M.E.; Glaser, R. Stress and Immune Responses after Surgical Treatment for Regional Breast Cancer. J. Natl. Cancer Inst. 1998, 90, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Andersen, B.L.; Farrar, W.B.; Golden-Kreutz, D.M.; Glaser, R.; Emery, C.F.; Crespin, T.R.; Shapiro, C.L.; Carson, W.E. Psychological, Behavioral, and Immune Changes after a Psychological Intervention: A Clinical Trial. J. Clin. Oncol. 2004, 22, 3570–3580. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Song, C.; Lin, A.-H.; Bonaccorso, S.; Kenis, G.; De Jongh, R.; Bosmans, E.; Scharpé, S. Negative Immunoregulatory Effects of Antidepressants: Inhibition of Interferon-γ and Stimulation of Interleukin-10 Secretion. Neuropsychopharmacology 1999, 20, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Pokryszko-Dragan, A.; Frydecka, I.; Kosmaczewska, A.; Ciszak, L.; Bilińska, M.; Gruszka, E.; Podemski, R.; Frydecka, D. Stimulated Peripheral Production of Interferon-Gamma Is Related to Fatigue and Depression in Multiple Sclerosis. Clin. Neurol. Neurosurg. 2012, 114, 1153–1158. [Google Scholar] [CrossRef] [PubMed]
- Mandolesi, G.; Bullitta, S.; Fresegna, D.; Gentile, A.; De Vito, F.; Dolcetti, E.; Rizzo, F.R.; Strimpakos, G.; Centonze, D.; Musella, A. Interferon-γ Causes Mood Abnormalities by Altering Cannabinoid CB1 Receptor Function in the Mouse Striatum. Neurobiol. Dis. 2017, 108, 45–53. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Daria, S.; Proma, M.A.; Shahriar, M.; Islam, S.M.A.; Bhuiyan, M.A.; Islam, M.R. Serum Interferon-Gamma Level Is Associated with Drug-Naïve Major Depressive Disorder. SAGE Open Med. 2020, 8, 2050312120974169. [Google Scholar] [CrossRef]
- Maes, M.; Scharpé, S.; Meltzer, H.Y.; Okayli, G.; Bosmans, E.; D’Hondt, P.; Vanden Bossche, B.V.; Cosyns, P. Increased Neopterin and Interferon-Gamma Secretion and Lower Availability of L-Tryptophan in Major Depression: Further Evidence for an Immune Response. Psychiatry Res. 1994, 54, 143–160. [Google Scholar] [CrossRef]
- Nam, J.-H.; Cha, B.; Park, J.-Y.; Abekura, F.; Kim, C.-H.; Kim, J.-R. Mitogen-Induced Interferon Gamma Production in Human Whole Blood: The Effect of Heat and Cations. Curr. Pharm. Biotechnol. 2019, 20, 562–572. [Google Scholar] [CrossRef]
- Zhang, J.; He, H.; Qiao, Y.; Zhou, T.; He, H.; Yi, S.; Zhang, L.; Mo, L.; Li, Y.; Jiang, W.; et al. Priming of Microglia with IFN-γ Impairs Adult Hippocampal Neurogenesis and Leads to Depression-like Behaviors and Cognitive Defects. Glia 2020, 68, 2674–2692. [Google Scholar] [CrossRef] [PubMed]
- Pusic, A.D.; Mitchell, H.M.; Kunkler, P.E.; Klauer, N.; Kraig, R.P. Spreading Depression Transiently Disrupts Myelin via Interferon-Gamma Signaling. Exp. Neurol. 2015, 264, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Oxenkrug, G.; Perianayagam, M.; Mikolich, D.; Requintina, P.; Shick, L.; Ruthazer, R.; Zucker, D.; Summergrad, P. Interferon-Gamma (+874) T/A Genotypes and Risk of IFN-Alpha-Induced Depression. J. Neural Transm. (Vienna) 2011, 118, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Mariani, N.; Everson, J.; Pariante, C.M.; Borsini, A. Modulation of Microglial Activation by Antidepressants. J. Psychopharmacol. 2022, 36, 131–150. [Google Scholar] [CrossRef] [PubMed]
- Myint, A.M.; Bondy, B.; Baghai, T.C.; Eser, D.; Nothdurfter, C.; Schüle, C.; Zill, P.; Müller, N.; Rupprecht, R.; Schwarz, M.J. Tryptophan Metabolism and Immunogenetics in Major Depression: A Role for Interferon-γ Gene. Brain Behav. Immun. 2013, 31, 128–133. [Google Scholar] [CrossRef] [PubMed]
- CCL2 C-C Motif Chemokine Ligand 2 [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/6347 (accessed on 13 December 2022).
- Bose, S.; Cho, J. Role of Chemokine CCL2 and Its Receptor CCR2 in Neurodegenerative Diseases. Arch. Pharm. Res. 2013, 36, 1039–1050. [Google Scholar] [CrossRef] [PubMed]
- Proma, M.A.; Daria, S.; Nahar, Z.; Ashraful Islam, S.M.; Bhuiyan, M.A.; Islam, M.R. Monocyte Chemoattractant Protein-1 Levels Are Associated with Major Depressive Disorder. J. Basic Clin. Physiol. Pharm. 2022, 33, 735–741. [Google Scholar] [CrossRef]
- Myung, W.; Lim, S.-W.; Woo, H.I.; Park, J.H.; Shim, S.; Lee, S.-Y.; Kim, D.K. Serum Cytokine Levels in Major Depressive Disorder and Its Role in Antidepressant Response. Psychiatry Investig. 2016, 13, 644–651. [Google Scholar] [CrossRef] [PubMed]
- Janelidze, S.; Ventorp, F.; Erhardt, S.; Hansson, O.; Minthon, L.; Flax, J.; Samuelsson, M.; Traskman-Bendz, L.; Brundin, L. Altered Chemokine Levels in the Cerebrospinal Fluid and Plasma of Suicide Attempters. Psychoneuroendocrinology 2013, 38, 853–862. [Google Scholar] [CrossRef]
- Simon, N.M.; McNamara, K.; Chow, C.W.; Maser, R.S.; Papakostas, G.I.; Pollack, M.H.; Nierenberg, A.A.; Fava, M.; Wong, K.K. A Detailed Examination of Cytokine Abnormalities in Major Depressive Disorder. Eur. Neuropsychopharmacol. 2008, 18, 230–233. [Google Scholar] [CrossRef]
- Zhou, Z.; Guille, C.; Ogunrinde, E.; Liu, R.; Luo, Z.; Powell, A.; Jiang, W. Increased Systemic Microbial Translocation Is Associated with Depression during Early Pregnancy. J. Psychiatr. Res. 2018, 97, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Curzytek, K.; Leśkiewicz, M. Targeting the CCL2-CCR2 Axis in Depressive Disorders. Pharm. Rep. 2021, 73, 1052–1062. [Google Scholar] [CrossRef] [PubMed]
- Marciniak, E.; Faivre, E.; Dutar, P.; Alves Pires, C.; Demeyer, D.; Caillierez, R.; Laloux, C.; Buée, L.; Blum, D.; Humez, S. The Chemokine MIP-1α/CCL3 Impairs Mouse Hippocampal Synaptic Transmission, Plasticity and Memory. Sci. Rep. 2015, 5, 15862. [Google Scholar] [CrossRef] [PubMed]
- Hoge, E.A.; Brandstetter, K.; Moshier, S.; Pollack, M.H.; Wong, K.K.; Simon, N.M. Broad Spectrum of Cytokine Abnormalities in Panic Disorder and Posttraumatic Stress Disorder. Depress. Anxiety 2009, 26, 447–455. [Google Scholar] [CrossRef]
- Merendino, R.A.; Di Pasquale, G.; De Luca, F.; Di Pasquale, L.; Ferlazzo, E.; Martino, G.; Palumbo, M.C.; Morabito, F.; Gangemi, S. Involvement of Fractalkine and Macrophage Inflammatory Protein-1 Alpha in Moderate-Severe Depression. Mediat. Inflamm. 2004, 13, 205–207. [Google Scholar] [CrossRef] [PubMed]
- Gaspersz, R.; Lamers, F.; Wittenberg, G.; Beekman, A.T.F.; van Hemert, A.M.; Schoevers, R.A.; Penninx, B.W.J.H. The Role of Anxious Distress in Immune Dysregulation in Patients with Major Depressive Disorder. Transl. Psychiatry 2017, 7, 1268. [Google Scholar] [CrossRef]
- Sundberg, I.; Rasmusson, A.J.; Ramklint, M.; Just, D.; Ekselius, L.; Cunningham, J.L. Daytime Melatonin Levels in Saliva Are Associated with Inflammatory Markers and Anxiety Disorders. Psychoneuroendocrinology 2020, 112, 104514. [Google Scholar] [CrossRef]
- Wang, X.; Fang, C.; Liu, X.; Wei, W.; Zhang, M.; Chen, S.; Shi, F. High Serum Levels of INOS and MIP-1α Are Associated with Post-Stroke Depression. Neuropsychiatr. Dis. Treat. 2021, 17, 2481–2487. [Google Scholar] [CrossRef]
- Camacho-Arroyo, I.; Flores-Ramos, M.; Mancilla-Herrera, I.; Cruz, F.M.C.; Hernández-Ruiz, J.; Diaz, G.P.; Labonne, B.F.; Del Pilar Meza-Rodríguez, M.; Gelman, P.L. Chemokine Profile in Women with Moderate to Severe Anxiety and Depression during Pregnancy. BMC Pregnancy Childbirth 2021, 21, 807. [Google Scholar] [CrossRef]
- Roomruangwong, C.; Sirivichayakul, S.; Carvalho, A.F.; Maes, M. The Uterine-Chemokine-Brain Axis: Menstrual Cycle-Associated Symptoms (MCAS) Are in Part Mediated by CCL2, CCL5, CCL11, CXCL8 and CXCL10. J. Affect. Disord. 2020, 269, 85–93. [Google Scholar] [CrossRef]
- Polacchini, A.; Girardi, D.; Falco, A.; Zanotta, N.; Comar, M.; De Carlo, N.A.; Tongiorgi, E. Distinct CCL2, CCL5, CCL11, CCL27, IL-17, IL-6, BDNF Serum Profiles Correlate to Different Job-Stress Outcomes. Neurobiol. Stress 2018, 8, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Hu, Q.; Zhang, K.; Teng, H.; Li, M.; Li, D.; Wang, J.; Du, Q.; Zhao, M. The Clock-Controlled Chemokine Contributes to Neuroinflammation-Induced Depression. FASEB J. 2020, 34, 8357–8366. [Google Scholar] [CrossRef] [PubMed]
- Małujło-Balcerska, E.; Kumor-Kisielewska, A.; Szemraj, J.; Pietras, T. Chemokine (C-C Motif) Ligand 5 (RANTES) Concentrations in the Peripheral Blood of Patients with a Depressive Disorder. Pharm. Rep. 2022, 74, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Leighton, S.P.; Nerurkar, L.; Krishnadas, R.; Johnman, C.; Graham, G.J.; Cavanagh, J. Chemokines in Depression in Health and in Inflammatory Illness: A Systematic Review and Meta-Analysis. Mol. Psychiatry 2018, 23, 48–58. [Google Scholar] [CrossRef]
- Ivanovska, M.; Abdi, Z.; Murdjeva, M.; Macedo, D.; Maes, A.; Maes, M. CCL-11 or Eotaxin-1: An Immune Marker for Ageing and Accelerated Ageing in Neuro-Psychiatric Disorders. Pharmaceuticals 2020, 13, 230. [Google Scholar] [CrossRef]
- Máchal, J.; Vašků, A.; Kincl, V.; Hlavna, M.; Bartáková, V.; Jurajda, M.; Meluzín, J. Association between Three Single Nucleotide Polymorphisms in Eotaxin (CCL 11) Gene, Hexanucleotide Repetition Upstream, Severity and Course of Coronary Atherosclerosis. J. Appl. Genet. 2012, 53, 271–278. [Google Scholar] [CrossRef]
- Teixeira, A.L.; Gama, C.S.; Rocha, N.P.; Teixeira, M.M. Revisiting the Role of Eotaxin-1/CCL11 in Psychiatric Disorders. Front. Psychiatry 2018, 9, 241. [Google Scholar] [CrossRef]
- Mohite, S.; Cordeiro, T.; Tannous, J.; Mwangi, B.; Selvaraj, S.; Soares, J.C.; Sanches, M.; Teixeira, A.L. Eotaxin-1/CCL11 Correlates with Left Superior Temporal Gyrus in Bipolar Disorder: A Preliminary Report Suggesting Accelerated Brain Aging. J. Affect. Disord. 2020, 273, 592–596. [Google Scholar] [CrossRef]
- Magalhaes, P.V.S.; Jansen, K.; Stertz, L.; Ferrari, P.; Pinheiro, R.T.; da Silva, R.A.; Kapczinski, F. Peripheral Eotaxin-1 (CCL11) Levels and Mood Disorder Diagnosis in a Population-Based Sample of Young Adults. J. Psychiatr. Res. 2014, 48, 13–15. [Google Scholar] [CrossRef]
- Grassi-Oliveira, R.; Brieztke, E.; Teixeira, A.; Pezzi, J.C.; Zanini, M.; Lopes, R.P.; Bauer, M.E. Peripheral Chemokine Levels in Women with Recurrent Major Depression with Suicidal Ideation. Braz J. Psychiatry 2012, 34, 71–75. [Google Scholar] [CrossRef]
- Ho, P.-S.; Yen, C.-H.; Chen, C.-Y.; Huang, S.-Y.; Liang, C.-S. Changes in Cytokine and Chemokine Expression Distinguish Dysthymic Disorder from Major Depression and Healthy Controls. Psychiatry Res. 2017, 248, 20–27. [Google Scholar] [CrossRef] [PubMed]
- TNF Tumor Necrosis Factor [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/7124 (accessed on 30 November 2022).
- Grimstad, Ø. Tumor Necrosis Factor and the Tenacious α. JAMA Derm. 2016, 152, 557. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Poletti, S.; Vai, B.; Mazza, M.G.; Lorenzi, C.; Brioschi, S.; Aggio, V.; Branchi, I.; Colombo, C.; Furlan, R.; et al. Higher Baseline Interleukin-1β and TNF-α Hamper Antidepressant Response in Major Depressive Disorder. Eur. Neuropsychopharmacol. 2021, 42, 35–44. [Google Scholar] [CrossRef]
- Das, R.; Emon, M.P.Z.; Shahriar, M.; Nahar, Z.; Islam, S.M.A.; Bhuiyan, M.A.; Islam, S.N.; Islam, M.R. Higher Levels of Serum IL-1β and TNF-α Are Associated with an Increased Probability of Major Depressive Disorder. Psychiatry Res. 2021, 295, 113568. [Google Scholar] [CrossRef] [PubMed]
- Bialek, K.; Czarny, P.; Watala, C.; Synowiec, E.; Wigner, P.; Bijak, M.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Preliminary Study of the Impact of Single-Nucleotide Polymorphisms of IL-1α, IL-1β and TNF-α Genes on the Occurrence, Severity and Treatment Effectiveness of the Major Depressive Disorder. Cell. Mol. Neurobiol. 2020, 40, 1049–1056. [Google Scholar] [CrossRef]
- Perry, B.I.; Upthegrove, R.; Kappelmann, N.; Jones, P.B.; Burgess, S.; Khandaker, G.M. Associations of Immunological Proteins/Traits with Schizophrenia, Major Depression and Bipolar Disorder: A Bi-Directional Two-Sample Mendelian Randomization Study. Brain Behav. Immun. 2021, 97, 176–185. [Google Scholar] [CrossRef]
- TNFRSF1B TNF Receptor Superfamily Member 1B [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/7133 (accessed on 30 November 2022).
- Black, R.A. Tumor Necrosis Factor-Alpha Converting Enzyme. Int. J. Biochem. Cell. Biol. 2002, 34, 1–5. [Google Scholar] [CrossRef]
- Bobińska, K.; Gałecka, E.; Szemraj, J.; Gałecki, P.; Talarowska, M. Is There a Link between TNF Gene Expression and Cognitive Deficits in Depression? Acta Biochim. Pol. 2017, 64, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Yamamori, H.; Ishima, T.; Yasuda, Y.; Fujimoto, M.; Kudo, N.; Ohi, K.; Hashimoto, K.; Takeda, M.; Hashimoto, R. Assessment of a Multi-Assay Biological Diagnostic Test for Mood Disorders in a Japanese Population. Neurosci. Lett. 2016, 612, 167–171. [Google Scholar] [CrossRef]
- Papakostas, G.I.; Shelton, R.C.; Kinrys, G.; Henry, M.E.; Bakow, B.R.; Lipkin, S.H.; Pi, B.; Thurmond, L.; Bilello, J.A. Assessment of a Multi-Assay, Serum-Based Biological Diagnostic Test for Major Depressive Disorder: A Pilot and Replication Study. Mol. Psychiatry 2013, 18, 332–339. [Google Scholar] [CrossRef]
- Grassi-Oliveira, R.; Brietzke, E.; Pezzi, J.C.; Lopes, R.P.; Teixeira, A.L.; Bauer, M.E. Increased Soluble Tumor Necrosis Factor-Alpha Receptors in Patients with Major Depressive Disorder. Psychiatry Clin. Neurosci. 2009, 63, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Diniz, B.S.; Teixeira, A.L.; Talib, L.L.; Mendonça, V.A.; Gattaz, W.F.; Forlenza, O.V. Increased Soluble TNF Receptor 2 in Antidepressant-Free Patients with Late-Life Depression. J. Psychiatr. Res. 2010, 44, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, F.M.; Koch, J.; Nowak, C.; Holdt, L.M.; Teupser, D.; Hegerl, U.; Himmerich, H. Ligands and Receptors of the TNF Superfamily Are Decreased in Major Depression and during Early Antidepressant Therapy. J. Psychiatr. Res. 2019, 119, 116–121. [Google Scholar] [CrossRef] [PubMed]
- Pillai, J.A.; Bebek, G.; Khrestian, M.; Bena, J.; Bergmann, C.C.; Bush, W.S.; Leverenz, J.B.; Bekris, L.M. TNFRSF1B Gene Variants and Related Soluble TNFR2 Levels Impact Resilience in Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 638922. [Google Scholar] [CrossRef] [PubMed]
- Hiew, L.-F.; Poon, C.-H.; You, H.-Z.; Lim, L.-W. TGF-β/Smad Signalling in Neurogenesis: Implications for Neuropsychiatric Diseases. Cells 2021, 10, 1382. [Google Scholar] [CrossRef] [PubMed]
- Qiu, A.; Zhang, H.; Wang, C.; Chong, Y.-S.; Shek, L.P.; Gluckman, P.D.; Meaney, M.J.; Fortier, M.V.; Wu, Y. Canonical TGF-β Signaling Regulates the Relationship between Prenatal Maternal Depression and Amygdala Development in Early Life. Transl. Psychiatry 2021, 11, 170. [Google Scholar] [CrossRef] [PubMed]
- Caraci, F.; Spampinato, S.F.; Morgese, M.G.; Tascedda, F.; Salluzzo, M.G.; Giambirtone, M.C.; Caruso, G.; Munafò, A.; Torrisi, S.A.; Leggio, G.M.; et al. Neurobiological Links between Depression and AD: The Role of TGF-Β1 Signaling as a New Pharmacological Target. Pharm. Res. 2018, 130, 374–384. [Google Scholar] [CrossRef]
- Grasso, M.; Caruso, G.; Godos, J.; Bonaccorso, A.; Carbone, C.; Castellano, S.; Currenti, W.; Grosso, G.; Musumeci, T.; Caraci, F. Improving Cognition with Nutraceuticals Targeting TGF-Β1 Signaling. Antioxidants 2021, 10, 1075. [Google Scholar] [CrossRef]
- Huang, C.; Zhang, F.; Li, P.; Song, C. Low-Dose IL-2 Attenuated Depression-like Behaviors and Pathological Changes through Restoring the Balances between IL-6 and TGF-β and between Th17 and Treg in a Chronic Stress-Induced Mouse Model of Depression. Int. J. Mol. Sci. 2022, 23, 13856. [Google Scholar] [CrossRef]
- Mihailova, S.; Ivanova-Genova, E.; Lukanov, T.; Stoyanova, V.; Milanova, V.; Naumova, E. A Study of TNF-α, TGF-β, IL-10, IL-6, and IFN-γ Gene Polymorphisms in Patients with Depression. J. Neuroimmunol. 2016, 293, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, A.; Cattane, N.; Malpighi, C.; Czamara, D.; Suarez, A.; Mariani, N.; Kajantie, E.; Luoni, A.; Eriksson, J.G.; Lahti, J.; et al. FoxO1, A2M, and TGF-Β1: Three Novel Genes Predicting Depression in Gene X Environment Interactions Are Identified Using Cross-Species and Cross-Tissues Transcriptomic and MiRNomic Analyses. Mol. Psychiatry 2018, 23, 2192–2208. [Google Scholar] [CrossRef] [PubMed]
- Wyller, V.B.; Nguyen, C.B.; Ludviksen, J.A.; Mollnes, T.E. Transforming Growth Factor Beta (TGF-β) in Adolescent Chronic Fatigue Syndrome. J. Transl. Med. 2017, 15, 245. [Google Scholar] [CrossRef] [PubMed]
- Vogelzangs, N.; Beekman, A.T.F.; van Reedt Dortland, A.K.B.; Schoevers, R.A.; Giltay, E.J.; de Jonge, P.; Penninx, B.W.J.H. Inflammatory and Metabolic Dysregulation and the 2-Year Course of Depressive Disorders in Antidepressant Users. Neuropsychopharmacology 2014, 39, 1624–1634. [Google Scholar] [CrossRef] [PubMed]
- Sanada, K.; Montero-Marin, J.; Barceló-Soler, A.; Ikuse, D.; Ota, M.; Hirata, A.; Yoshizawa, A.; Hatanaka, R.; Valero, M.S.; Demarzo, M.; et al. Effects of Mindfulness-Based Interventions on Biomarkers and Low-Grade Inflammation in Patients with Psychiatric Disorders: A Meta-Analytic Review. Int. J. Mol. Sci. 2020, 21, 2484. [Google Scholar] [CrossRef] [PubMed]
- Walsh, E.; Eisenlohr-Moul, T.; Baer, R. Brief Mindfulness Training Reduces Salivary IL-6 and TNF-α in Young Women with Depressive Symptomatology. J. Consult. Clin. Psychol. 2016, 84, 887–897. [Google Scholar] [CrossRef]


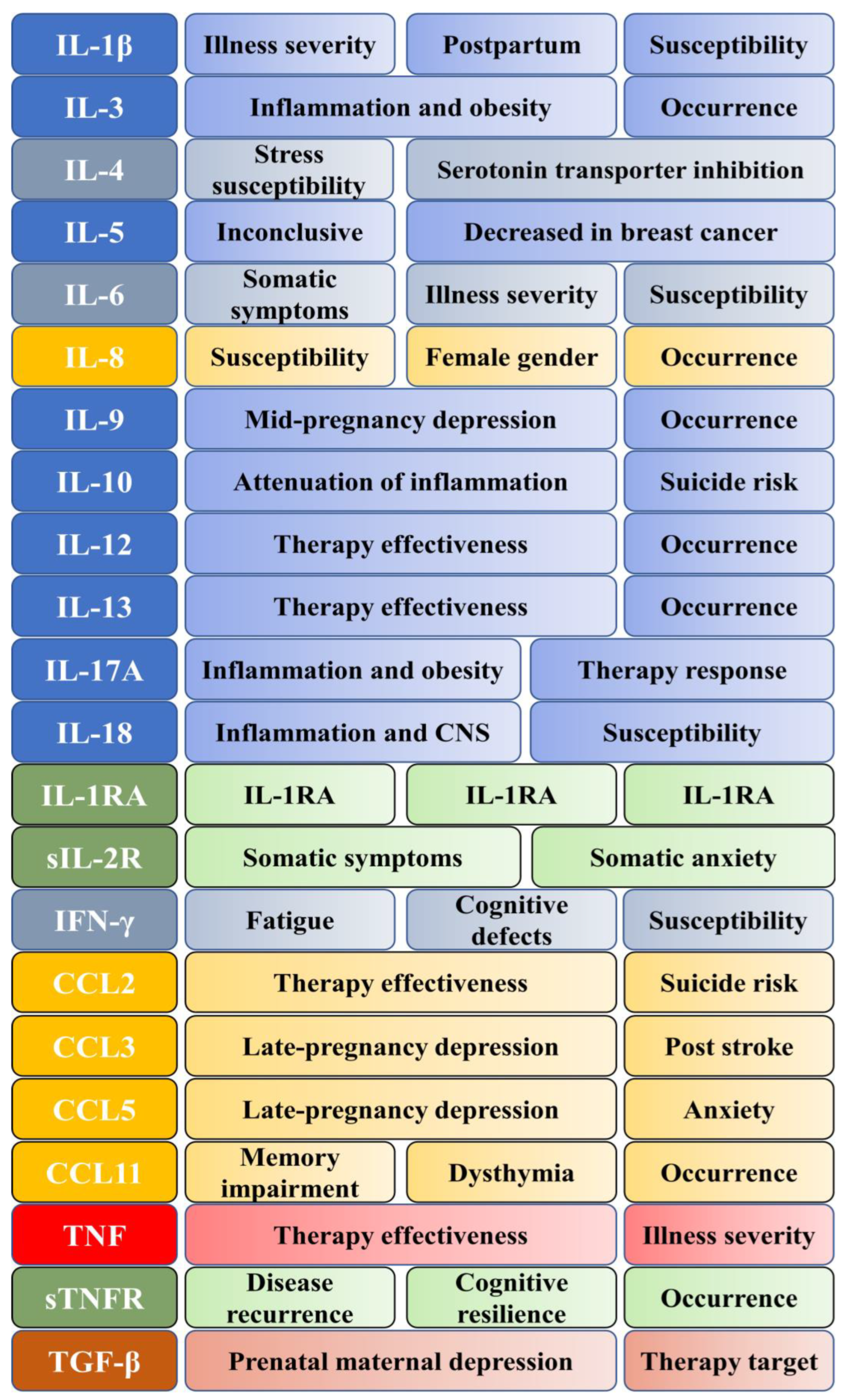{kind=link}
| Group | Cytokines | Soluble Receptors |
|---|---|---|
| Interleukins | IL-1β IL-3 IL-4 IL-5 IL-6 IL-9 IL-10 IL-12 IL-13 IL-17A IL-18 | IL-1 receptor antagonist Soluble IL-2 receptor |
| Interferons | IFN-γ | |
| Chemokines | IL-8 CCL2 CCL3 (MIP-1α) CCL5 (RANTES) CCL11 (Eotaxin-1) | |
| Lymphokines | IL-4 IL-6 IFN-γ | Soluble IL-2 receptor |
| Tumor necrosis factor superfamily | TNF | Soluble TNF receptor 2 |
| Transforming growth factor superfamily | TGF-β |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harsanyi, S.; Kupcova, I.; Danisovic, L.; Klein, M. Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation? Int. J. Mol. Sci. 2023, 24, 578. https://doi.org/10.3390/ijms24010578
Harsanyi S, Kupcova I, Danisovic L, Klein M. Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation? International Journal of Molecular Sciences. 2023; 24(1):578. https://doi.org/10.3390/ijms24010578
Chicago/Turabian StyleHarsanyi, Stefan, Ida Kupcova, Lubos Danisovic, and Martin Klein. 2023. "Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation?" International Journal of Molecular Sciences 24, no. 1: 578. https://doi.org/10.3390/ijms24010578
APA StyleHarsanyi, S., Kupcova, I., Danisovic, L., & Klein, M. (2023). Selected Biomarkers of Depression: What Are the Effects of Cytokines and Inflammation? International Journal of Molecular Sciences, 24(1), 578. https://doi.org/10.3390/ijms24010578