
A Low-Activity Polymorphic Variant of Human NEIL2 DNA Glycosylase


Abstract
:1. Introduction
2. Results
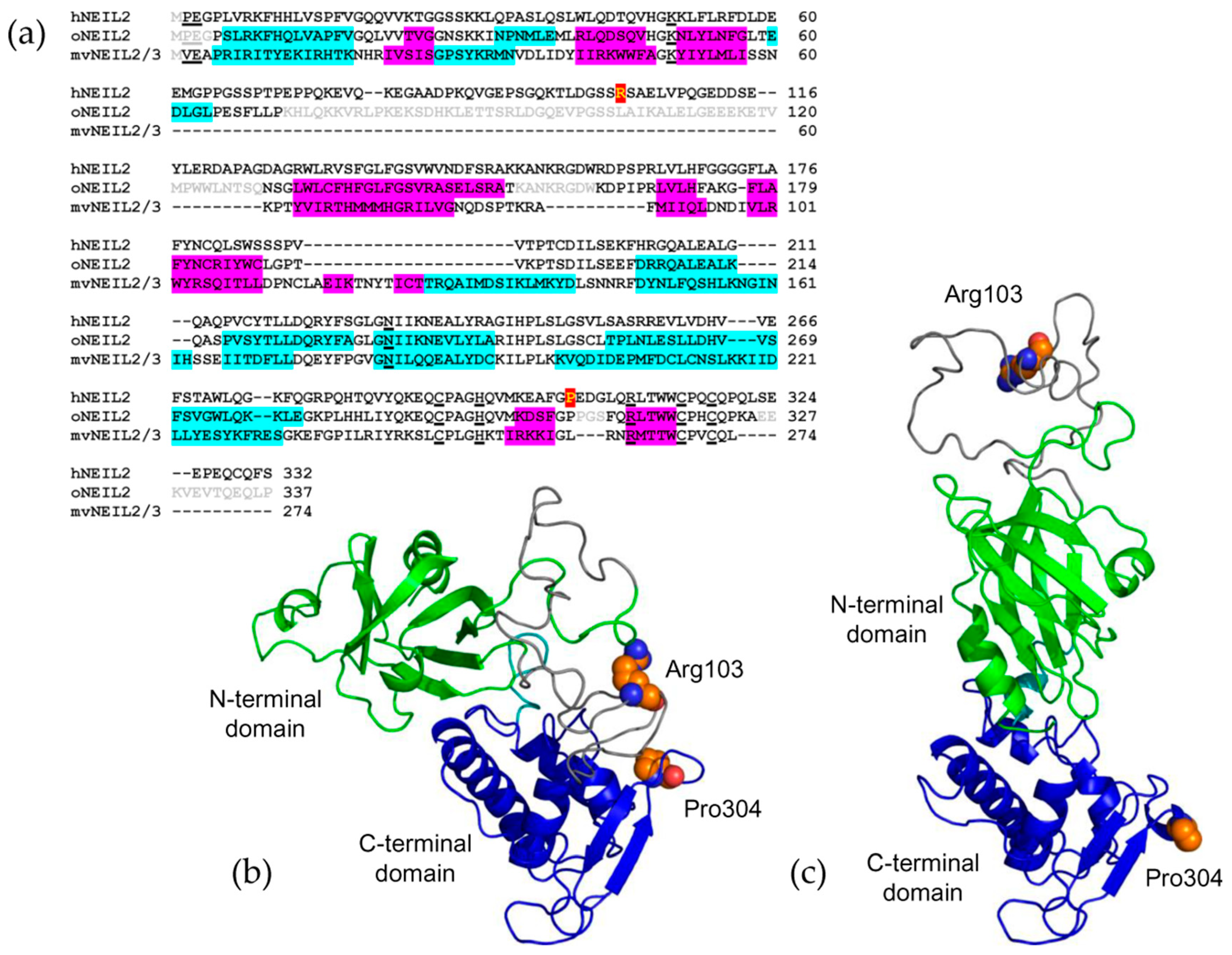
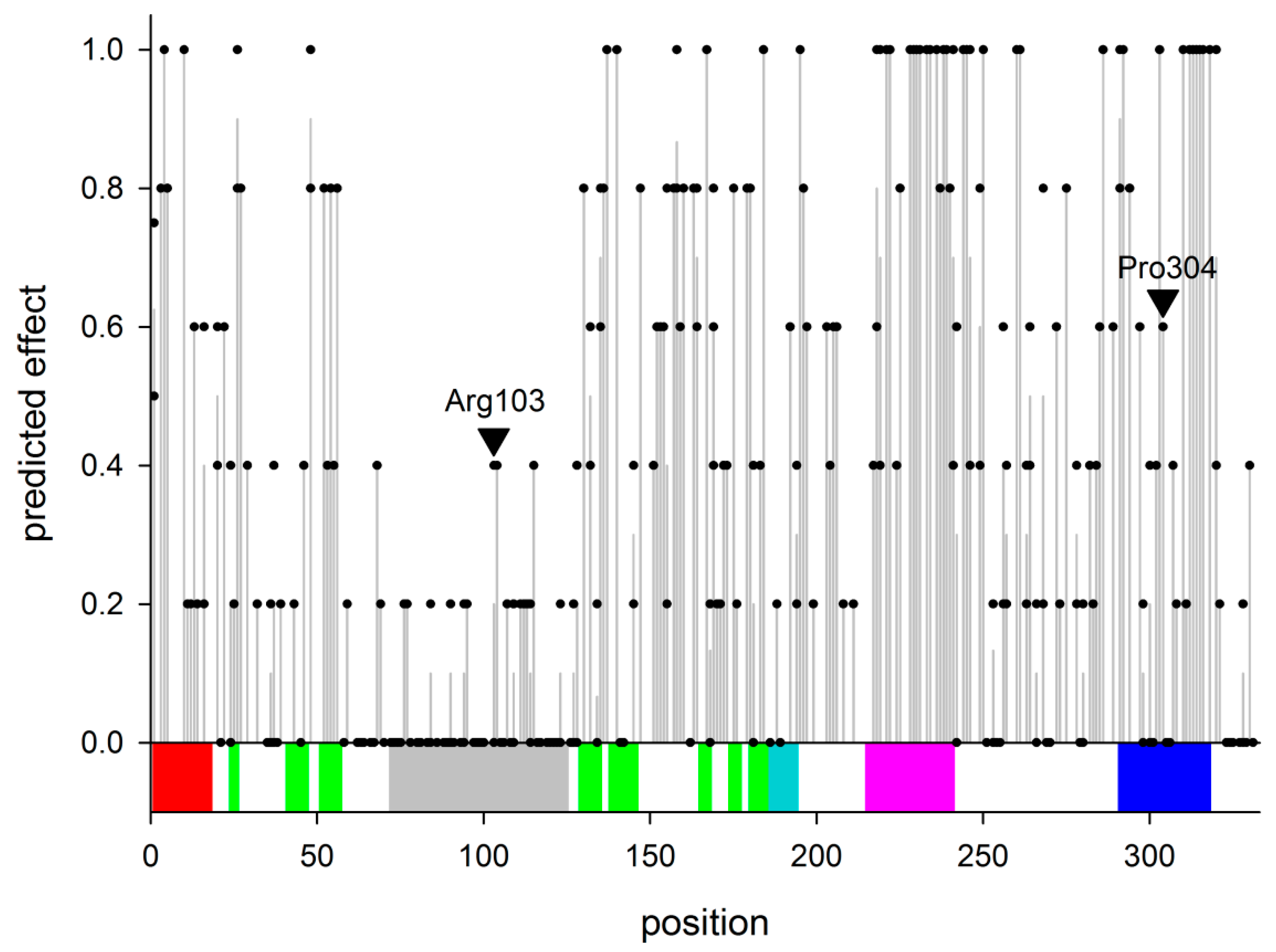
2.1. Non-Synonymous Polymorphisms in hNEIL2
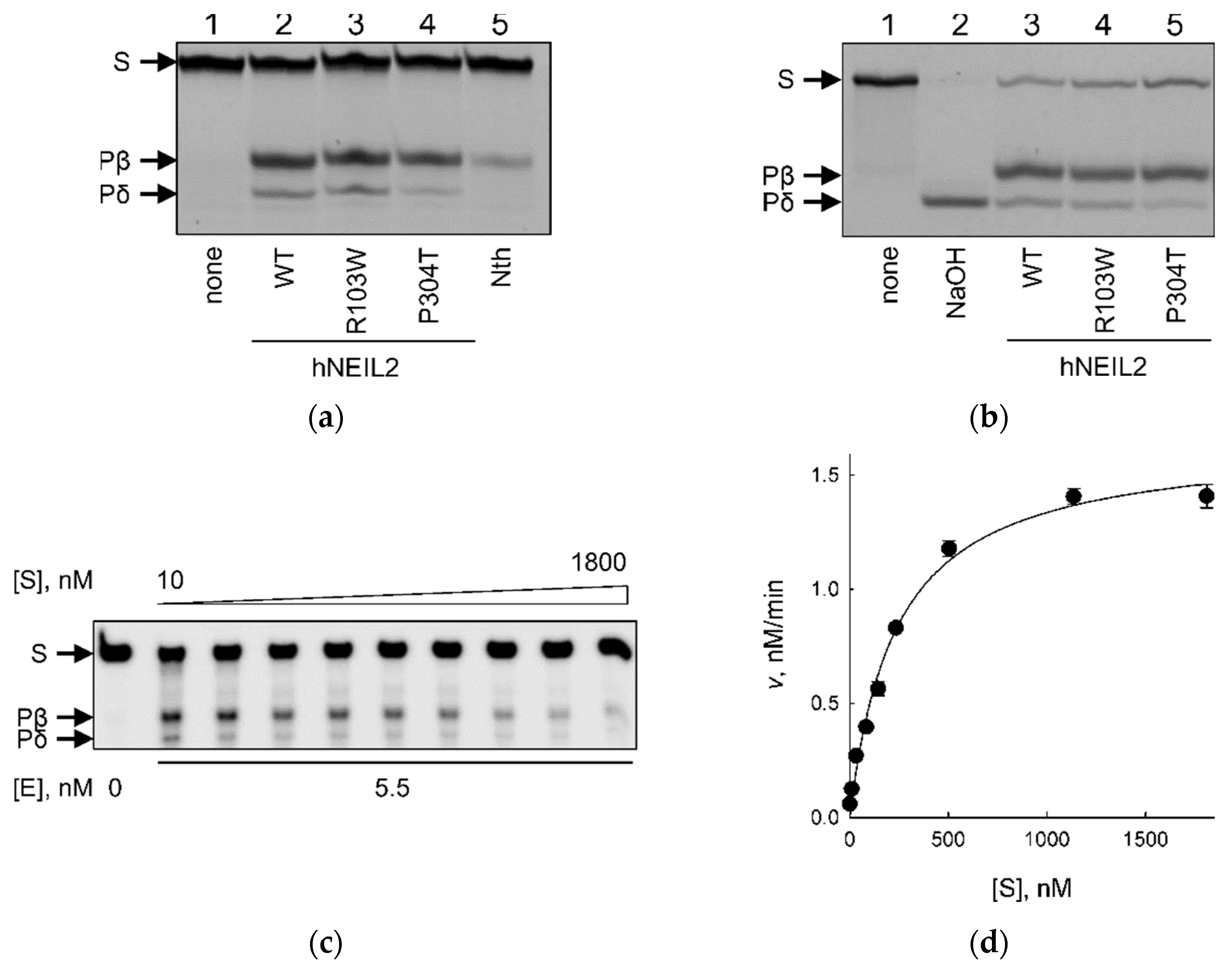
2.2. Activity of hNEIL2 R103W and P304T on Duplex DNA Substrates
2.3. hNEIL2 R103W and P304T Show Reduced Binding to DNA
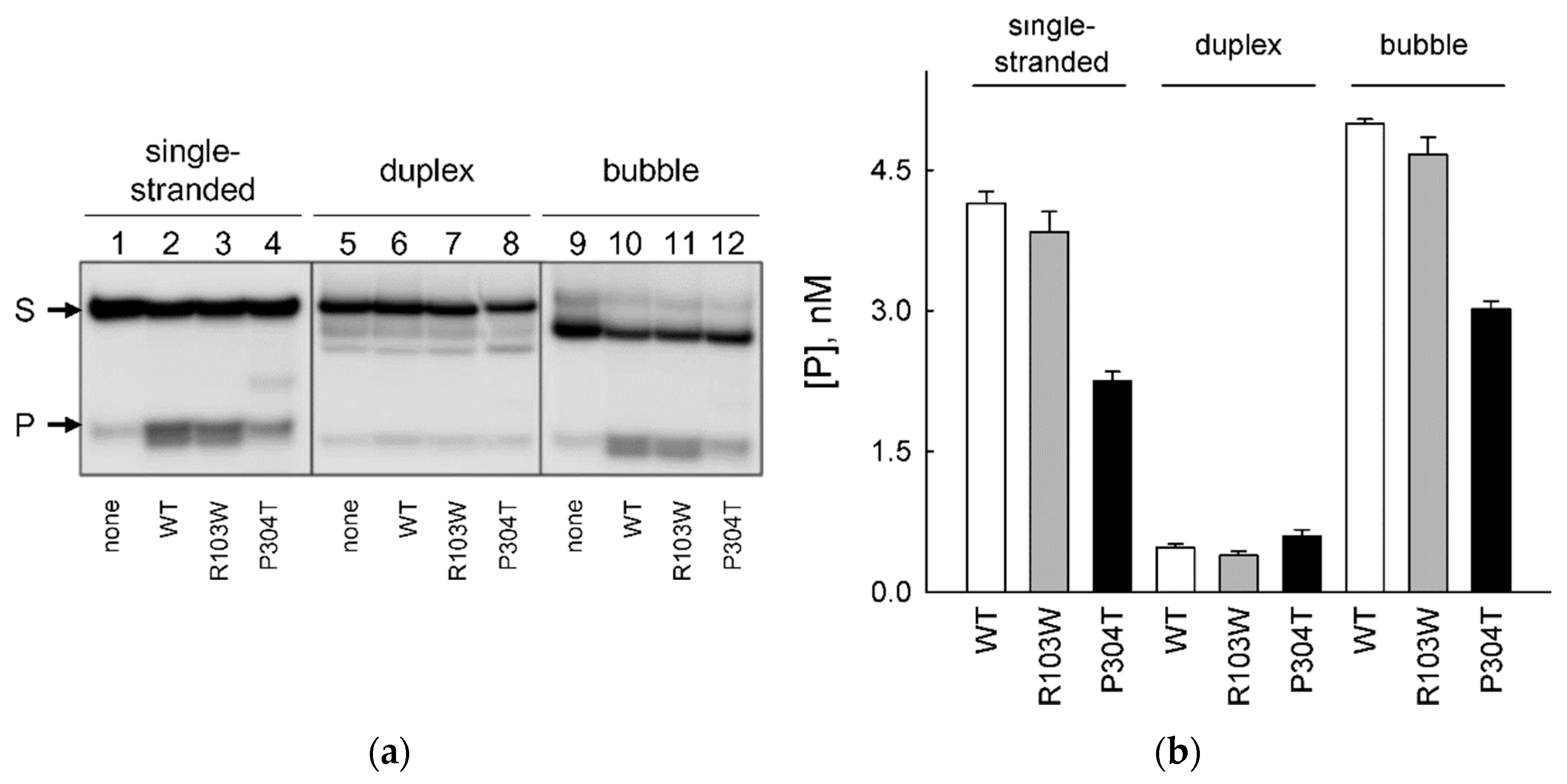
2.4. Activity of hNEIL2 R103W and P304T on Bubble DNA Substrates
3. Discussion
4. Materials and Methods
4.1. Enzymes and Oligonucleotides
4.2. hNEIL2 Purification
4.3. Enzyme Activity and Kinetics Measurements
4.4. Electrophoretic Mobiliy Shift Assay
4.5. In Silico Analysis of the Polymorphism Effect
4.6. Homology Modeling
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Friedberg, E.C.; Walker, G.C.; Siede, W.; Wood, R.D.; Schultz, R.A.; Ellenberger, T. DNA Repair and Mutagenesis; ASM Press: Washington, DC, USA, 2006; 1118p. [Google Scholar]
- Beard, W.A.; Horton, J.K.; Prasad, R.; Wilson, S.H. Eukaryotic base excision repair: New approaches shine light on mechanism. Annu. Rev. Biochem. 2019, 88, 137–162. [Google Scholar] [CrossRef] [PubMed]
- Caldecott, K.W. Mammalian DNA base excision repair: Dancing in the moonlight. DNA Repair 2020, 93, 102921. [Google Scholar] [CrossRef] [PubMed]
- Stivers, J.T.; Jiang, Y.L. A mechanistic perspective on the chemistry of DNA repair glycosylases. Chem. Rev. 2003, 103, 2729–2760. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.L.; Sundheim, O.; Tainer, J.A. DNA base damage recognition and removal: New twists and grooves. Mutat. Res. 2005, 577, 55–76. [Google Scholar] [CrossRef]
- Hitomi, K.; Iwai, S.; Tainer, J.A. The intricate structural chemistry of base excision repair machinery: Implications for DNA damage recognition, removal, and repair. DNA Repair 2007, 6, 410–428. [Google Scholar] [CrossRef]
- Zharkov, D.O. Base excision DNA repair. Cell. Mol. Life Sci. 2008, 65, 1544–1565. [Google Scholar] [CrossRef]
- Hazra, T.K.; Kow, Y.W.; Hatahet, Z.; Imhoff, B.; Boldogh, I.; Mokkapati, S.K.; Mitra, S.; Izumi, T. Identification and characterization of a novel human DNA glycosylase for repair of cytosine-derived lesions. J. Biol. Chem. 2002, 277, 30417–30420. [Google Scholar] [CrossRef] [Green Version]
- Hailer, M.K.; Slade, P.G.; Martin, B.D.; Rosenquist, T.A.; Sugden, K.D. Recognition of the oxidized lesions spiroiminodihydantoin and guanidinohydantoin in DNA by the mammalian base excision repair glycosylases NEIL1 and NEIL2. DNA Repair 2005, 4, 41–50. [Google Scholar] [CrossRef]
- Redrejo-Rodríguez, M.; Saint-Pierre, C.; Couve, S.; Mazouzi, A.; Ishchenko, A.A.; Gasparutto, D.; Saparbaev, M. New insights in the removal of the hydantoins, oxidation product of pyrimidines, via the base excision and nucleotide incision repair pathways. PLoS ONE 2011, 6, e21039. [Google Scholar] [CrossRef] [Green Version]
- Makasheva, K.A.; Endutkin, A.V.; Zharkov, D.O. Requirements for DNA bubble structure for efficient cleavage by helix–two-turn–helix DNA glycosylases. Mutagenesis 2020, 35, 119–128. [Google Scholar] [CrossRef]
- Dou, H.; Mitra, S.; Hazra, T.K. Repair of oxidized bases in DNA bubble structures by human DNA glycosylases NEIL1 and NEIL2. J. Biol. Chem. 2003, 278, 49679–49684. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Krishnamurthy, N.; Burrows, C.J.; David, S.S. Mutation versus repair: NEIL1 removal of hydantoin lesions in single-stranded, bulge, bubble, and duplex DNA contexts. Biochemistry 2010, 49, 1658–1666. [Google Scholar] [CrossRef] [Green Version]
- Eckenroth, B.E.; Cao, V.B.; Averill, A.M.; Dragon, J.A.; Doublié, S. Unique structural features of mammalian NEIL2 DNA glycosylase prime its activity for diverse DNA substrates and environments. Structure 2021, 29, 29–42.e4. [Google Scholar] [CrossRef] [PubMed]
- Prakash, A.; Eckenroth, B.E.; Averill, A.M.; Imamura, K.; Wallace, S.S.; Doublié, S. Structural investigation of a viral ortholog of human NEIL2/3 DNA glycosylases. DNA Repair 2013, 12, 1062–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhdanova, P.V.; Ishchenko, A.A.; Chernonosov, A.A.; Zharkov, D.O.; Koval, V.V. Dynamics and conformational changes in human NEIL2 DNA glycosylase analyzed by hydrogen/deuterium exchange mass spectrometry. J. Mol. Biol. 2022, 434, 167334. [Google Scholar] [CrossRef]
- Tate, J.G.; Bamford, S.; Jubb, H.C.; Sondka, Z.; Beare, D.M.; Bindal, N.; Boutselakis, H.; Cole, C.G.; Creatore, C.; Dawson, E.; et al. COSMIC: The Catalogue Of Somatic Mutations In Cancer. Nucleic Acids Res. 2019, 47, D941–D947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chae, Y.K.; Anker, J.F.; Carneiro, B.A.; Chandra, S.; Kaplan, J.; Kalyan, A.; Santa-Maria, C.A.; Platanias, L.C.; Giles, F.J. Genomic landscape of DNA repair genes in cancer. Oncotarget 2016, 7, 23312–23321. [Google Scholar] [CrossRef] [Green Version]
- Osorio, A.; Milne, R.L.; Kuchenbaecker, K.; Vaclová, T.; Pita, G.; Alonso, R.; Peterlongo, P.; Blanco, I.; de la Hoya, M.; Duran, M.; et al. DNA glycosylases involved in base excision repair may be associated with cancer risk in BRCA1 and BRCA2 mutation carriers. PLoS Genet. 2014, 10, e1004256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamina, C.; Coassin, S.; Illig, T.; Kronenberg, F. Look beyond one’s own nose: Combination of information from publicly available sources reveals an association of GATA4 polymorphisms with plasma triglycerides. Atherosclerosis 2011, 219, 698–703. [Google Scholar] [CrossRef]
- Benítez-Buelga, C.; Baquero, J.M.; Vaclova, T.; Fernández, V.; Martín, P.; Inglada-Perez, L.; Urioste, M.; Osorio, A.; Benítez, J. Genetic variation in the NEIL2 DNA glycosylase gene is associated with oxidative DNA damage in BRCA2 mutation carriers. Oncotarget 2017, 8, 114626–114636. [Google Scholar] [CrossRef]
- Zhai, X.; Zhao, H.; Liu, Z.; Wang, L.-E.; El-Naggar, A.K.; Sturgis, E.M.; Wei, Q. Functional variants of the NEIL1 and NEIL2 genes and risk and progression of squamous cell carcinoma of the oral cavity and oropharynx. Clin. Cancer Res. 2008, 14, 4345–4352. [Google Scholar] [CrossRef] [Green Version]
- Koster, R.; Mitra, N.; D’Andrea, K.; Vardhanabhuti, S.; Chung, C.C.; Wang, Z.; Erickson, R.L.; Vaughn, D.J.; Litchfield, K.; Rahman, N.; et al. Pathway-based analysis of GWAs data identifies association of sex determination genes with susceptibility to testicular germ cell tumors. Hum. Mol. Genet. 2014, 23, 6061–6068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mou, X.; Li, T.; Wang, J.; Ali, Z.; Zhang, Y.; Chen, Z.; Deng, Y.; Li, S.; Su, E.; Jia, Q.; et al. Genetic variation of BCL2 (rs2279115), NEIL2 (rs804270), LTA (rs909253), PSCA (rs2294008) and PLCE1 (rs3765524, rs10509670) genes and their correlation to gastric cancer risk based on universal tagged arrays and Fe3O4 magnetic nanoparticles. J. Biomed. Nanotechnol. 2015, 11, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Liu, J.; Wang, H.; Chen, X.; Cheng, Q.; Chen, H. Cervical carcinoma risk associate with genetic polymorphisms of NEIL2 gene in Chinese population and its significance as predictive biomarker. Sci. Rep. 2020, 10, 5136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cumova, A.; Vymetalkova, V.; Opattova, A.; Bouskova, V.; Pardini, B.; Kopeckova, K.; Kozevnikovova, R.; Lickova, K.; Ambrus, M.; Vodickova, L.; et al. Genetic variations in 3′UTRs of SMUG1 and NEIL2 genes modulate breast cancer risk, survival and therapy response. Mutagenesis 2021, 36, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Pardini, B.; Rosa, F.; Barone, E.; Di Gaetano, C.; Slyskova, J.; Novotny, J.; Levy, M.; Garritano, S.; Vodickova, L.; Buchler, T.; et al. Variation within 3′-UTRs of base excision repair genes and response to therapy in colorectal cancer patients: A potential modulation of microRNAs binding. Clin. Cancer Res. 2013, 19, 6044–6056. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Pang, L.; Gong, S.; Wang, X.; Hou, L. Nei endonuclease VIII-like 2 gene rs8191670 polymorphism affects the sensitivity of non-small cell lung cancer to cisplatin by binding with MiR-548a. J. Cancer 2020, 11, 4801–4809. [Google Scholar] [CrossRef]
- Wei, H.; Kamat, A.; Chen, M.; Ke, H.-L.; Chang, D.W.; Yin, J.; Grossman, H.B.; Dinney, C.P.; Wu, X. Association of polymorphisms in oxidative stress genes with clinical outcomes for bladder cancer treated with Bacillus Calmette-Guérin. PLoS ONE 2012, 7, e38533. [Google Scholar] [CrossRef]
- McGue, M.; Zhang, Y.; Miller, M.B.; Basu, S.; Vrieze, S.; Hicks, B.; Malone, S.; Oetting, W.S.; Iacono, W.G. A genome-wide association study of behavioral disinhibition. Behav. Genet. 2013, 43, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.; Zou, X.; Zhang, G.; Xiang, J.; Wang, Y.; Yang, M.; Chen, X.; Wu, J.; Guan, H. A variant in a microRNA binding site in NEIL2 3′UTR confers susceptibility to age-related cataracts. FASEB J. 2019, 33, 10469–10476. [Google Scholar] [CrossRef]
- Kinslow, C.J.; El-Zein, R.A.; Hill, C.E.; Wickliffe, J.K.; Abdel-Rahman, S.Z. Single nucleotide polymorphisms 5′ upstream the coding region of the NEIL2 gene influence gene transcription levels and alter levels of genetic damage. Genes Chromosomes Cancer 2008, 47, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, A.; Wakamiya, M.; Venkova-Canova, T.; Pandita, R.K.; Aguilera-Aguirre, L.; Sarker, A.H.; Singh, D.K.; Hosoki, K.; Wood, T.G.; Sharma, G.; et al. Neil2-null mice accumulate oxidized DNA bases in the transcriptionally active sequences of the genome and are susceptible to innate inflammation. J. Biol. Chem. 2015, 290, 24636–24648. [Google Scholar] [CrossRef] [Green Version]
- Sayed, I.M.; Sahan, A.Z.; Venkova, T.; Chakraborty, A.; Mukhopadhyay, D.; Bimczok, D.; Beswick, E.J.; Reyes, V.E.; Pinchuk, I.; Sahoo, D.; et al. Helicobacter pylori infection downregulates the DNA glycosylase NEIL2, resulting in increased genome damage and inflammation in gastric epithelial cells. J. Biol. Chem. 2020, 295, 11082–11098. [Google Scholar] [CrossRef]
- Sayed, I.M.; Chakraborty, A.; Abd El-Hafeez, A.A.; Sharma, A.; Sahan, A.Z.; Huang, W.J.M.; Sahoo, D.; Ghosh, P.; Hazra, T.K.; Das, S. The DNA glycosylase NEIL2 suppresses Fusobacterium-infection-induced inflammation and DNA damage in colonic epithelial cells. Cells 2020, 9, 1980. [Google Scholar] [CrossRef]
- Englander, E.W.; Ma, H. Differential modulation of base excision repair activities during brain ontogeny: Implications for repair of transcribed DNA. Mech. Ageing Dev. 2006, 127, 64–69. [Google Scholar] [CrossRef]
- Bhakat, K.K.; Hazra, T.K.; Mitra, S. Acetylation of the human DNA glycosylase NEIL2 and inhibition of its activity. Nucleic Acids Res. 2004, 32, 3033–3039. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Rajagopalan, L.; Mathura, V.S.; Rigby, S.J.; Mitra, S.; Hazra, T.K. Identification of a zinc finger domain in the human NEIL2 (Nei-like-2) protein. J. Biol. Chem. 2004, 279, 47132–47138. [Google Scholar] [CrossRef] [Green Version]
- Dey, S.; Maiti, A.K.; Hegde, M.L.; Hegde, P.M.; Boldogh, I.; Sarkar, P.S.; Abdel-Rahman, S.Z.; Sarker, A.H.; Hang, B.; Xie, J.; et al. Increased risk of lung cancer associated with a functionally impaired polymorphic variant of the human DNA glycosylase NEIL2. DNA Repair 2012, 11, 570–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shihab, H.A.; Gough, J.; Cooper, D.N.; Stenson, P.D.; Barker, G.L.A.; Edwards, K.J.; Day, I.N.M.; Gaunt, T.R. Predicting the functional, molecular, and phenotypic consequences of amino acid substitutions using hidden Markov models. Hum. Mutat. 2013, 34, 57–65. [Google Scholar] [CrossRef]
- Reva, B.; Antipin, Y.; Sander, C. Predicting the functional impact of protein mutations: Application to cancer genomics. Nucleic Acids Res. 2011, 39, e118. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, J.M.; Cooper, D.N.; Schuelke, M.; Seelow, D. MutationTaster2: Mutation prediction for the deep-sequencing age. Nat. Methods 2014, 11, 361–362. [Google Scholar] [CrossRef] [PubMed]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Sims, G.E.; Murphy, S.; Miller, J.R.; Chan, A.P. Predicting the functional effect of amino acid substitutions and indels. PLoS ONE 2012, 7, e46688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dodson, M.L.; Michaels, M.L.; Lloyd, R.S. Unified catalytic mechanism for DNA glycosylases. J. Biol. Chem. 1994, 269, 32709–32712. [Google Scholar] [CrossRef]
- Zharkov, D.O.; Shoham, G.; Grollman, A.P. Structural characterization of the Fpg family of DNA glycosylases. DNA Repair 2003, 2, 839–862. [Google Scholar] [CrossRef]
- Brot, F.E.; Bender, M.L. Use of the specificity constant of α-chymotrypsin. J. Am. Chem. Soc. 1969, 91, 7187–7191. [Google Scholar] [CrossRef]
- Fastrez, J.; Fersht, A.R. Mechanism of chymotrypsin. Structure, reactivity, and nonproductive binding relations. Biochemistry 1973, 12, 1067–1074. [Google Scholar] [CrossRef]
- Szeltner, Z.; Rea, D.; Juhász, T.; Renner, V.; Mucsi, Z.; Orosz, G.; Fülöp, V.; Polgár, L. Substrate-dependent competency of the catalytic triad of prolyl oligopeptidase. J. Biol. Chem. 2002, 277, 44597–44605. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Hunt, J.F. Enzymological and structural studies of the mechanism of promiscuous substrate recognition by the oxidative DNA repair enzyme AlkB. Proc. Natl Acad. Sci. USA 2009, 106, 14315–14320. [Google Scholar] [CrossRef] [Green Version]
- Smider, V.; Hwang, B.J.; Chu, G. Electrophoretic mobility shift assays to study protein binding to damaged DNA. Methods Mol. Biol. 2006, 314, 323–344. [Google Scholar] [CrossRef]
- Imamura, K.; Wallace, S.S.; Doublié, S. Structural characterization of a viral NEIL1 ortholog unliganded and bound to abasic site-containing DNA. J. Biol. Chem. 2009, 284, 26174–26183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grin, I.R.; Rieger, R.A.; Zharkov, D.O. Inactivation of NEIL2 DNA glycosylase by pyridoxal phosphate reveals a loop important for substrate binding. Biochem. Biophys. Res. Commun. 2010, 394, 100–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Lu, L.; Zhang, J.; Yue, Z.; Song, J.; Zong, S.; Liu, M.; Stovicek, O.; Gao, Y.Q.; Yi, C. Tautomerization-dependent recognition and excision of oxidation damage in base-excision DNA repair. Proc. Natl Acad. Sci. USA 2016, 113, 7792–7797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endutkin, A.V.; Yudkina, A.V.; Sidorenko, V.S.; Zharkov, D.O. Transient protein–protein complexes in base excision repair. J. Biomol. Struct. Dyn. 2019, 37, 4407–4418. [Google Scholar] [CrossRef] [PubMed]
- Golan, G.; Zharkov, D.O.; Feinberg, H.; Fernandes, A.S.; Zaika, E.I.; Kycia, J.H.; Grollman, A.P.; Shoham, G. Structure of the uncomplexed DNA repair enzyme endonuclease VIII indicates significant interdomain flexibility. Nucleic Acids Res. 2005, 33, 5006–5016. [Google Scholar] [CrossRef] [Green Version]
- Duclos, S.; Aller, P.; Jaruga, P.; Dizdaroglu, M.; Wallace, S.S.; Doublié, S. Structural and biochemical studies of a plant formamidopyrimidine-DNA glycosylase reveal why eukaryotic Fpg glycosylases do not excise 8-oxoguanine. DNA Repair 2012, 11, 714–725. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Imamura, K.; Averill, A.M.; Wallace, S.S.; Doublié, S. Structural characterization of a mouse ortholog of human NEIL3 with a marked preference for single-stranded DNA. Structure 2013, 21, 247–256. [Google Scholar] [CrossRef] [Green Version]
- Prakash, A.; Carroll, B.L.; Sweasy, J.B.; Wallace, S.S.; Doublié, S. Genome and cancer single nucleotide polymorphisms of the human NEIL1 DNA glycosylase: Activity, structure, and the effect of editing. DNA Repair 2014, 14, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Rabbie, R.; Ferguson, P.; Wong, K.; Couturier, D.-L.; Moran, U.; Turner, C.; Emanuel, P.; Haas, K.; Saunus, J.M.; Davidson, M.R.; et al. The mutational landscape of melanoma brain metastases presenting as the first visceral site of recurrence. Br. J. Cancer 2021, 124, 156–160. [Google Scholar] [CrossRef]
- Dherin, C.; Radicella, J.P.; Dizdaroglu, M.; Boiteux, S. Excision of oxidatively damaged DNA bases by the human α-hOgg1 protein and the polymorphic α-hOgg1(Ser326Cys) protein which is frequently found in human populations. Nucleic Acids Res. 1999, 27, 4001–4007. [Google Scholar] [CrossRef] [Green Version]
- Simonelli, V.; Camerini, S.; Mazzei, F.; Van Loon, B.; Allione, A.; D’Errico, M.; Barone, F.; Minoprio, A.; Ricceri, F.; Guarrera, S.; et al. Genotype–phenotype analysis of S326C OGG1 polymorphism: A risk factor for oxidative pathologies. Free Radic. Biol. Med. 2013, 63, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Kim, S.K.; Park, H.J.; Chung, J.-H.; Ban, J.Y. Human 8-oxoguanine DNA glycosylase gene polymorphism (Ser326Cys) and cancer risk: Updated meta-analysis. Oncotarget 2017, 8, 44761–44775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuznetsov, N.A.; Kladova, O.A.; Kuznetsova, A.A.; Ishchenko, A.A.; Saparbaev, M.K.; Zharkov, D.O.; Fedorova, O.S. Conformational dynamics of DNA repair by Escherichia coli endonuclease III. J. Biol. Chem. 2015, 290, 14338–14349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—Distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Sequence, 5′→3′ | Modification (X) |
|---|---|---|
| 23OHU | CTCTCCCTTCXCTCCTTTCCTCT | OHU |
| 23U | CTCTCCCTTCXCTCCTTTCCTCT | U |
| 23F | CTCTCCCTTCXCTCCTTTCCTCT | THF |
| 23C | CTCTCCCTTCCCTCCTTTCCTCT | |
| 23comp | AGAGGAAAGGAGGGAAGGGAGAG | |
| 40C | CCTGCATGGGCGGCAGAACTCGAGGCCATCCTCACCATCC | |
| 40F | CCTGCATGGGCGGCAGAACTXGAGGCCATCCTCACCATCC | THF |
| 40bubble | GGATGGTGAGGATGGGAGGTTCCCTTGCCGCCCATGCAGG | |
| 60DHU | GTAACAGTTCCTGCATGGGCGGCATGAACXGGAGGCCCATCCTCACCATCATCACACTGG | DHU |
| 60comp | CCAGTGTGATGATGGTGAGGATGGGCCTCCGGTTCATGCCGCCCATGCAGGAACTGTTAC | |
| 60bubble | CCAGTGTGATGATGGTGAGGATGGCGAGGTTCCCTCTGCCGCCCATGCAGGAACTGTTAC |
| Enzyme | KM, nM | kcat, min−1 | kcat/KM, μM−1·min−1 |
|---|---|---|---|
| WT | 17 ± 2 | 0.060 ± 0.002 | 3.5 ± 0.4 |
| R103W | 99 ± 9 | 0.28 ± 0.01 | 2.8 ± 0.3 |
| P304T | 500 ± 30 | 0.36 ± 0.01 | 0.72 ± 0.05 |
| Enzyme | Kbindapp(THF), μM | Kbindapp(C), μM |
|---|---|---|
| WT | 1.0 ± 0.1 | 1.0 ± 0.1 |
| R103W | 10.4 ± 1.2 | 11.7 ± 0.7 |
| P304T | 6.4 ± 0.1 | 7.0 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kakhkharova, Z.I.; Zharkov, D.O.; Grin, I.R. A Low-Activity Polymorphic Variant of Human NEIL2 DNA Glycosylase. Int. J. Mol. Sci. 2022, 23, 2212. https://doi.org/10.3390/ijms23042212
Kakhkharova ZI, Zharkov DO, Grin IR. A Low-Activity Polymorphic Variant of Human NEIL2 DNA Glycosylase. International Journal of Molecular Sciences. 2022; 23(4):2212. https://doi.org/10.3390/ijms23042212
Chicago/Turabian StyleKakhkharova, Zarina I., Dmitry O. Zharkov, and Inga R. Grin. 2022. "A Low-Activity Polymorphic Variant of Human NEIL2 DNA Glycosylase" International Journal of Molecular Sciences 23, no. 4: 2212. https://doi.org/10.3390/ijms23042212
APA StyleKakhkharova, Z. I., Zharkov, D. O., & Grin, I. R. (2022). A Low-Activity Polymorphic Variant of Human NEIL2 DNA Glycosylase. International Journal of Molecular Sciences, 23(4), 2212. https://doi.org/10.3390/ijms23042212