
Genome-Wide Characterization and Expression Analysis of Pathogenesis-Related 1 (PR-1) Gene Family in Tea Plant (Camellia sinensis (L.) O. Kuntze) in Response to Blister-Blight Disease Stress


Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of CsPR-1 Gene-Family Members
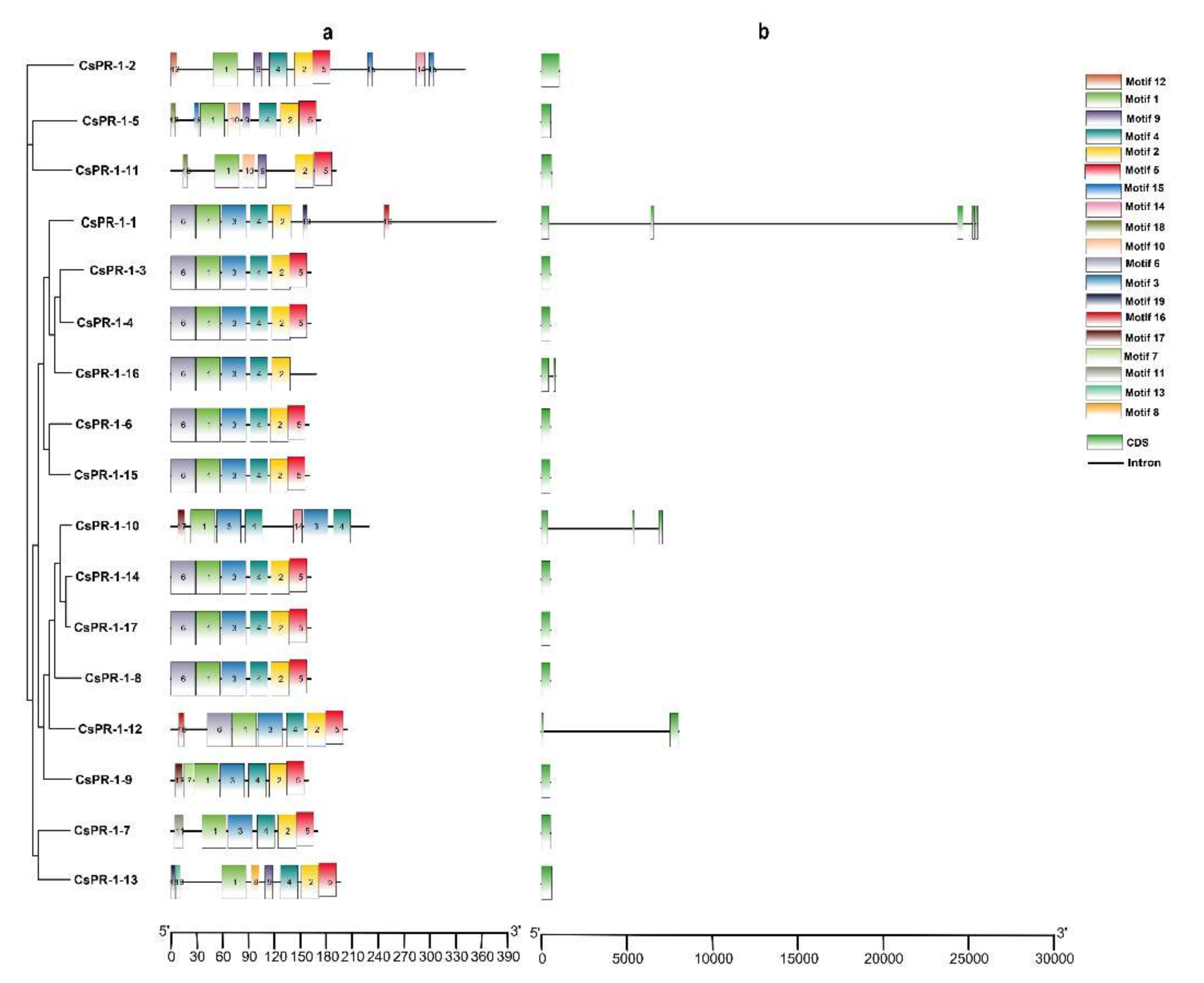
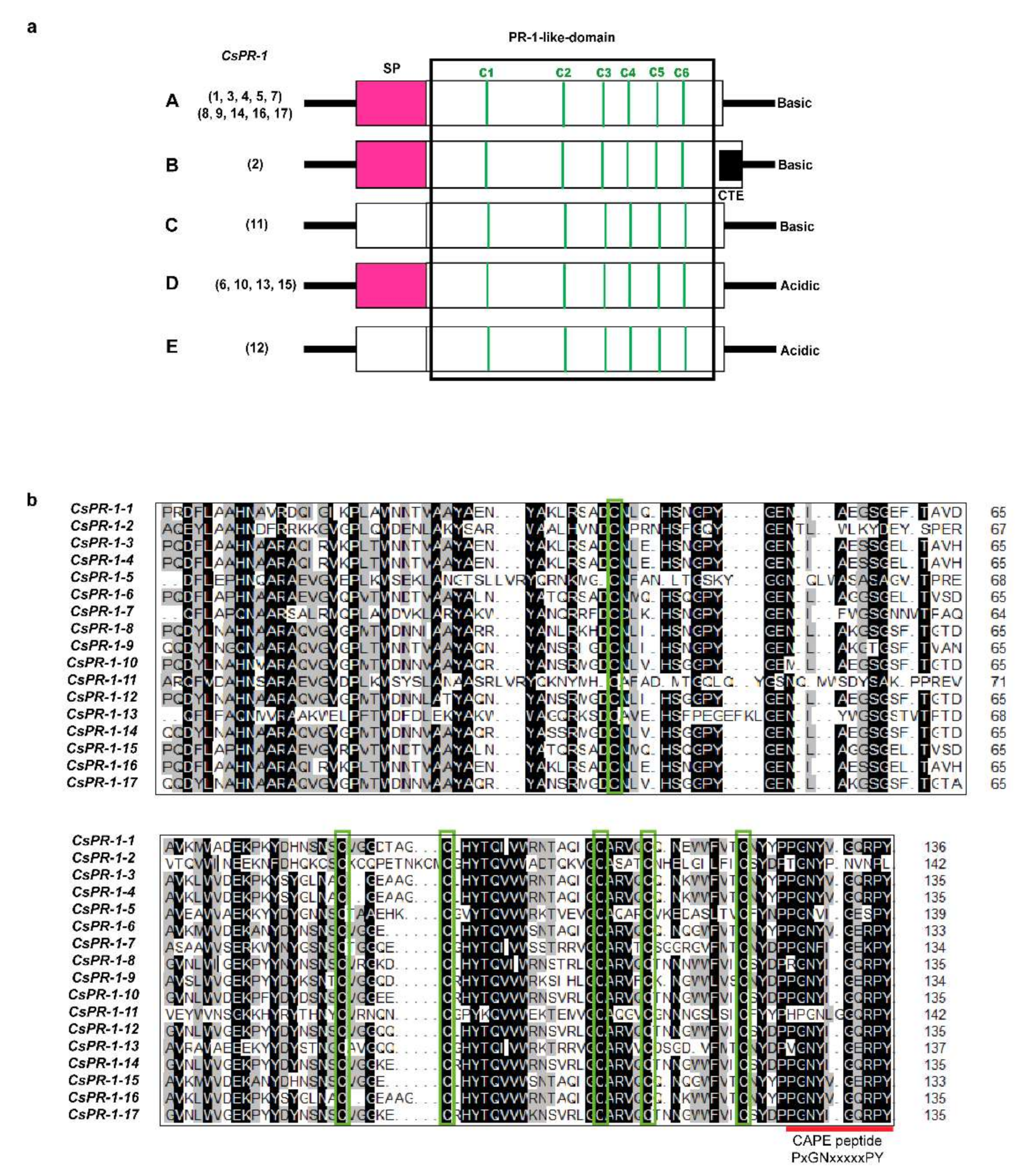
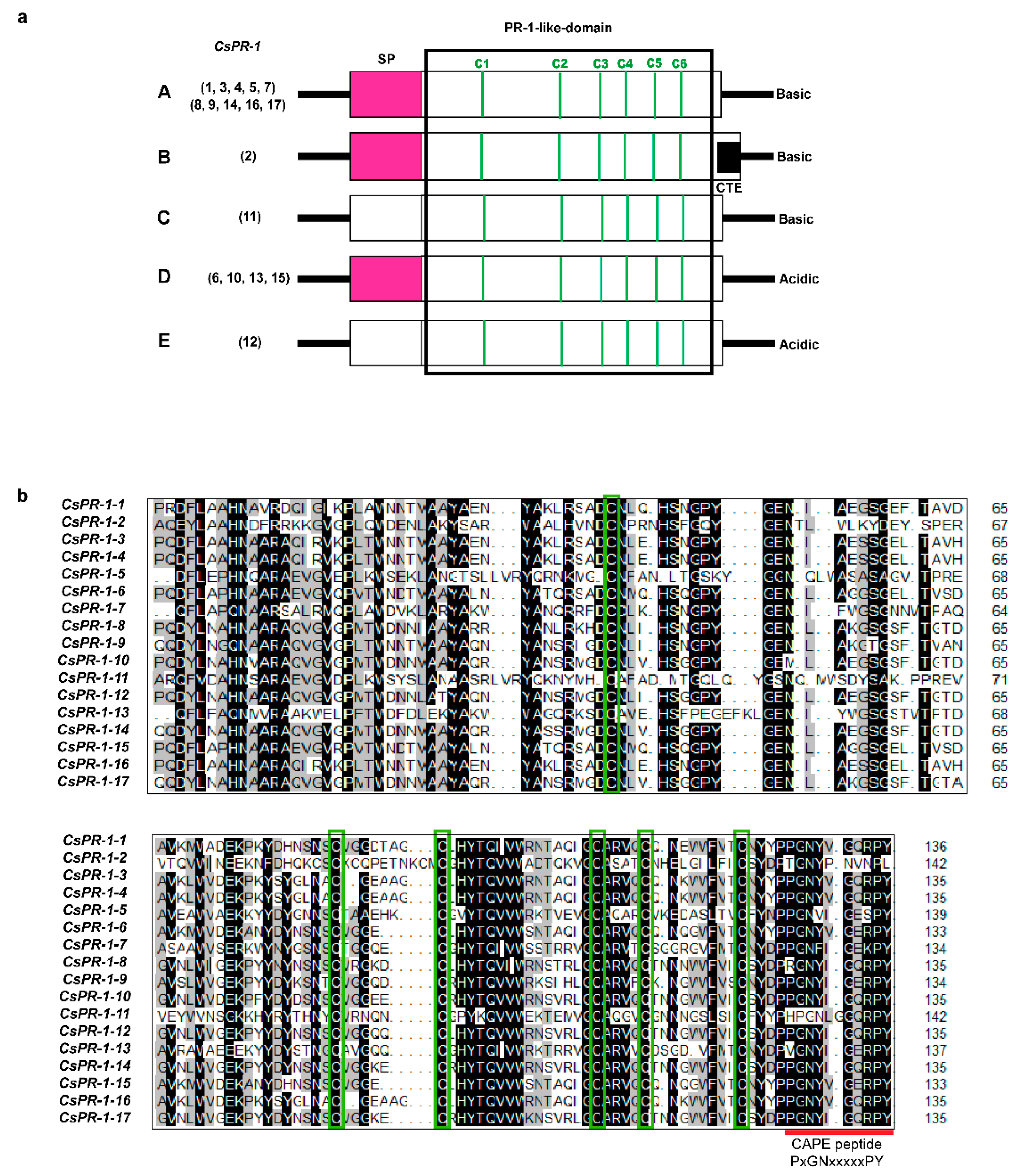
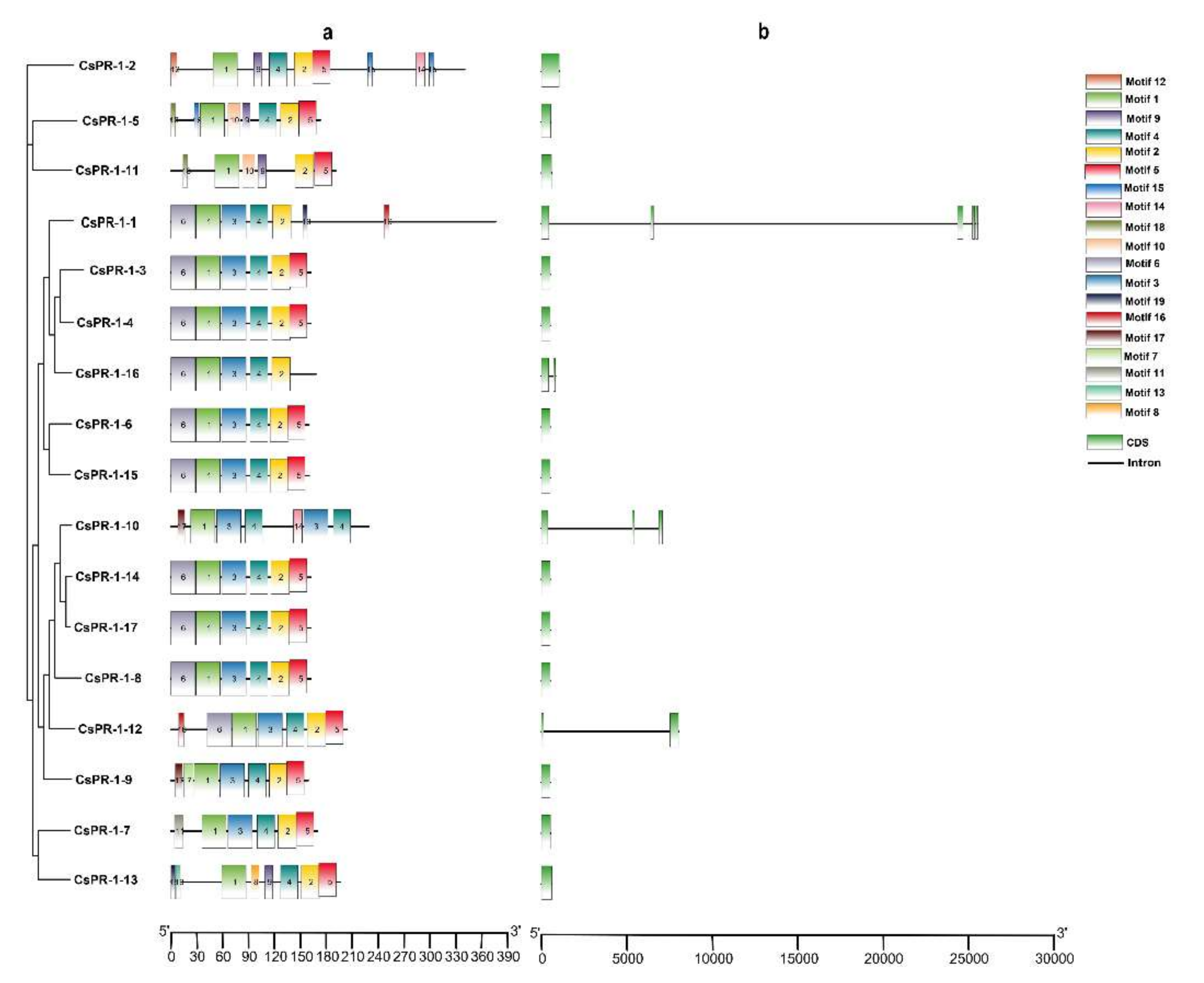
2.2. Analysis of the Genomic and Domain Structures of CsPR-1 Genes
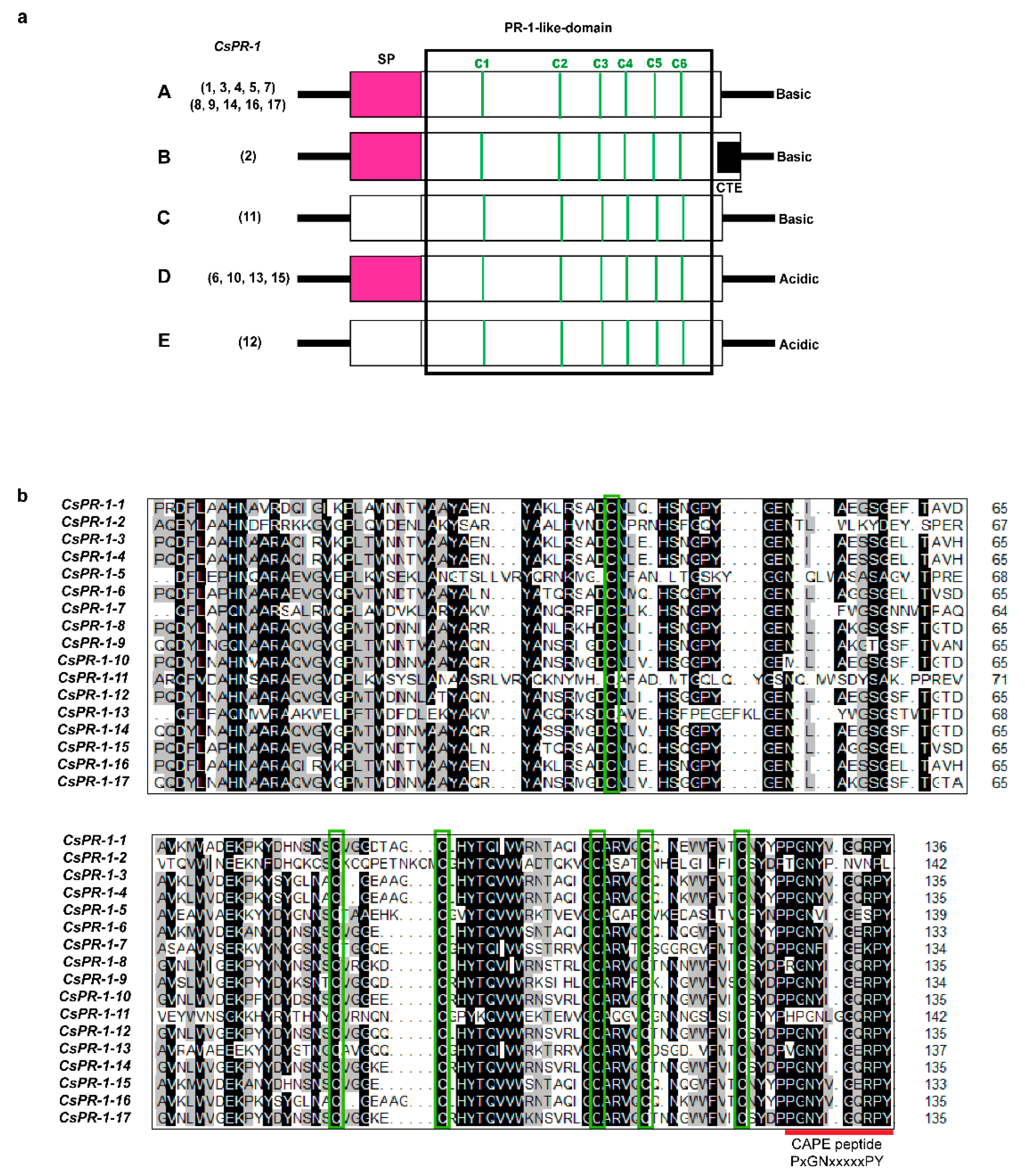
2.3. Conserved Motifs of the CsPR-1 Protein Structure
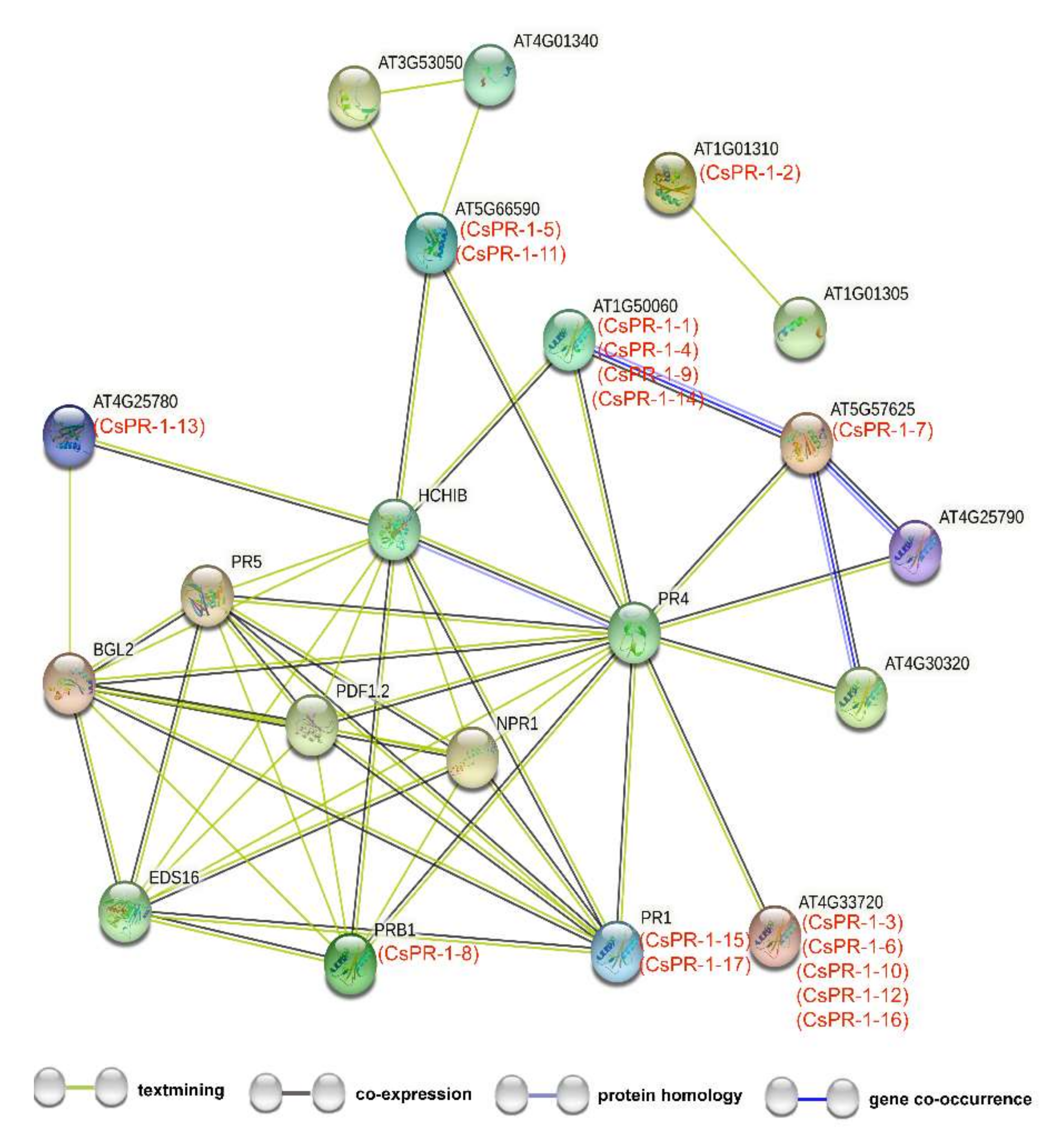
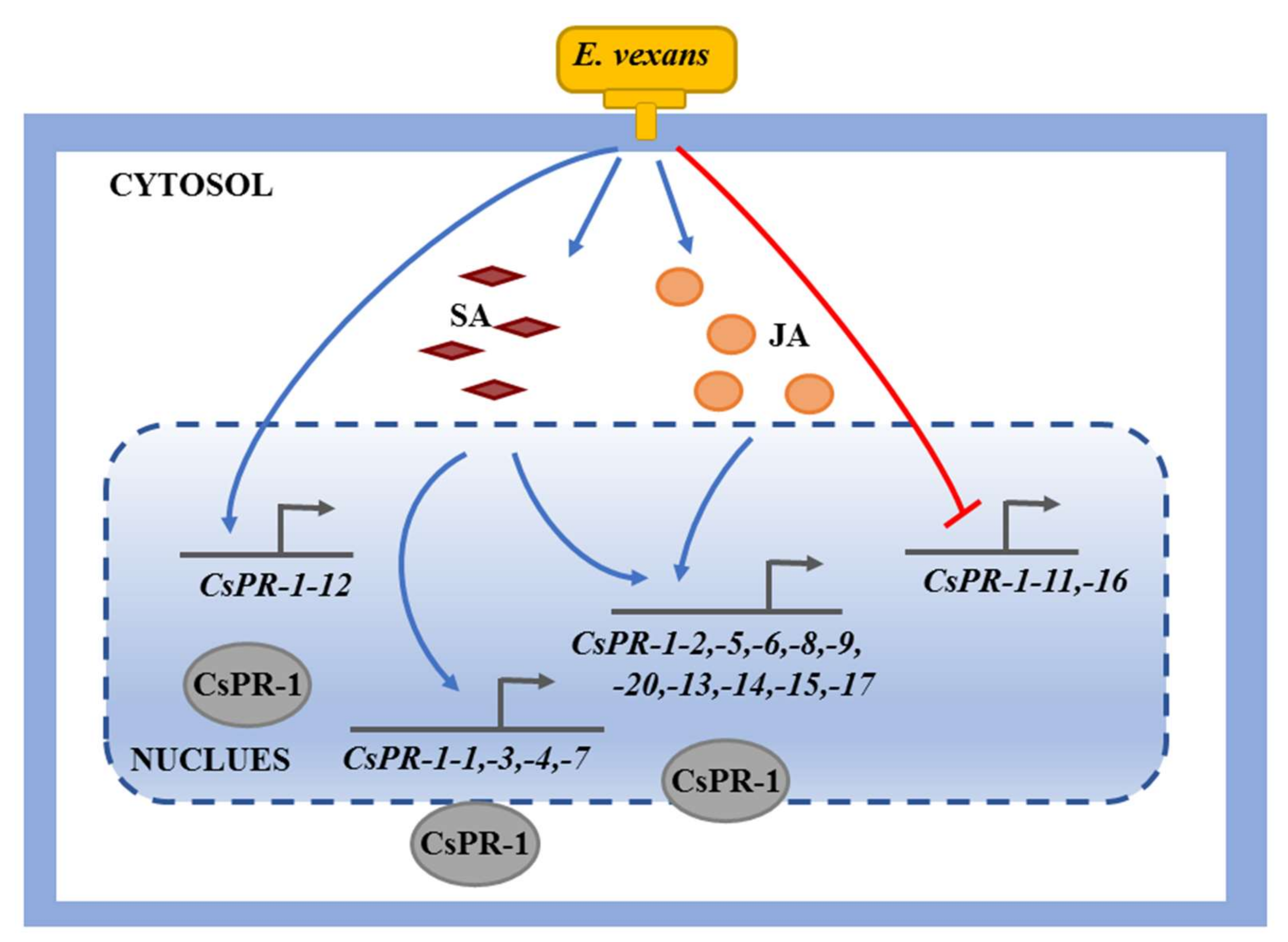
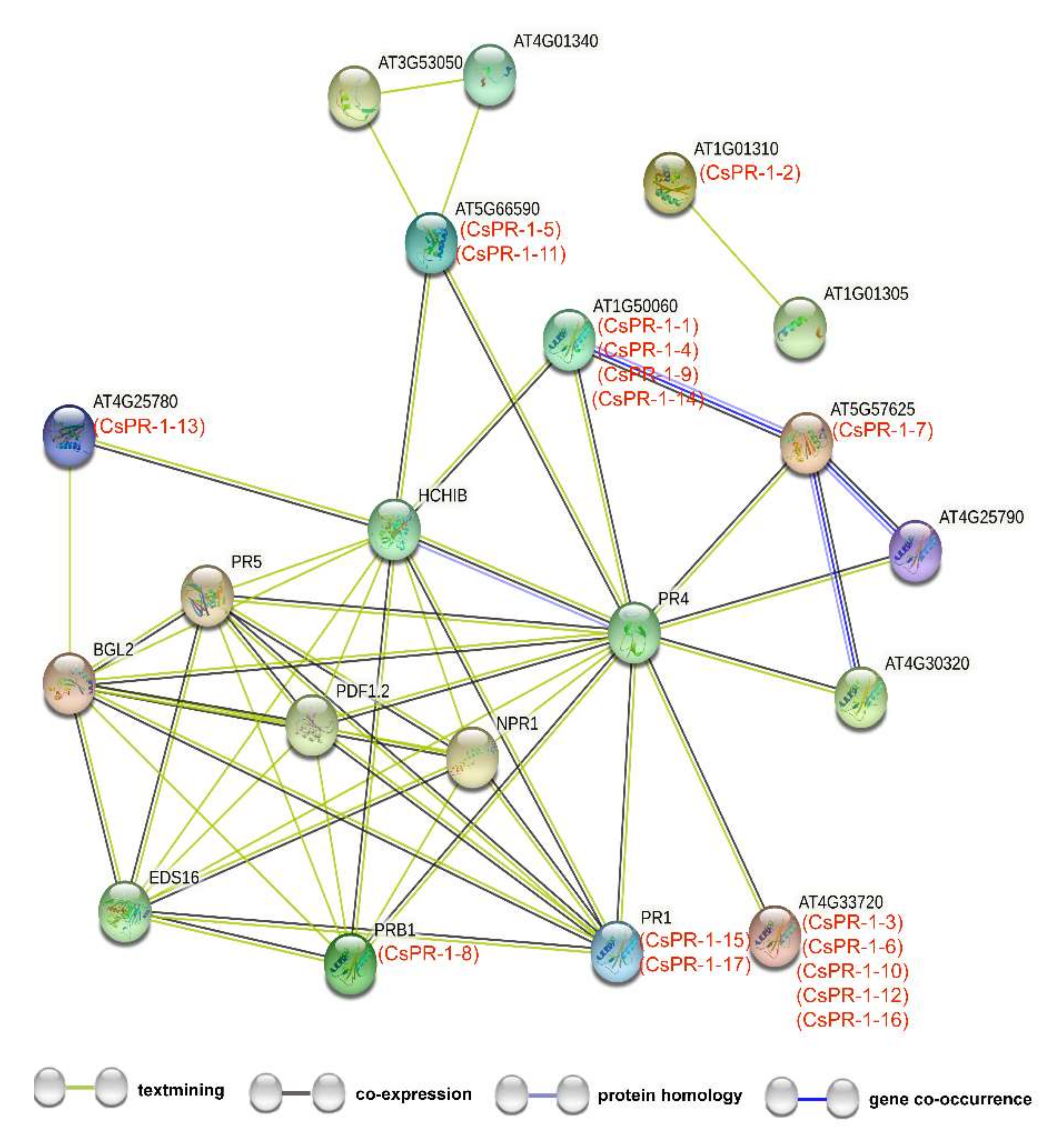
2.4. Functional Interaction Networks of CsPR-1 Proteins
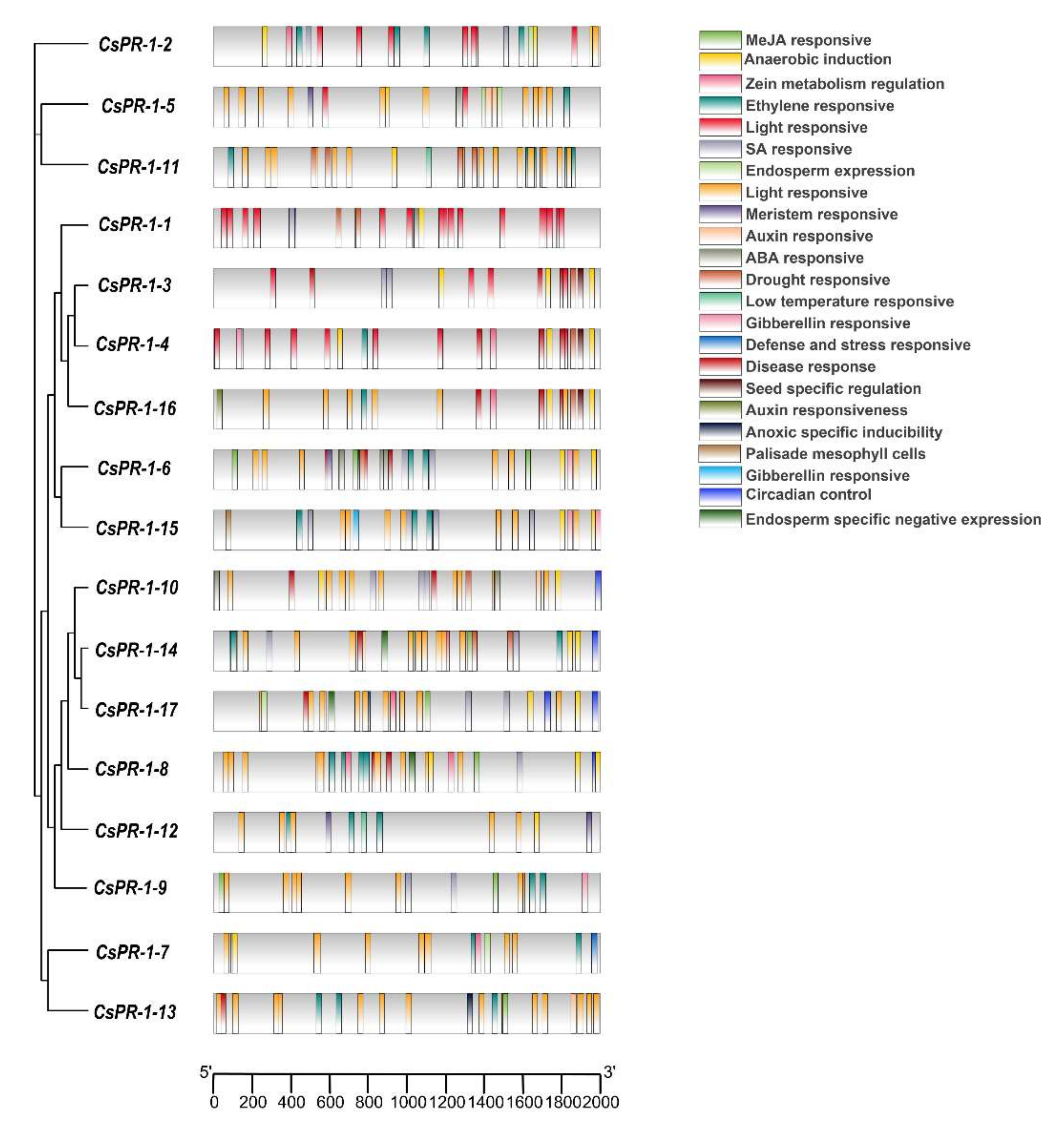
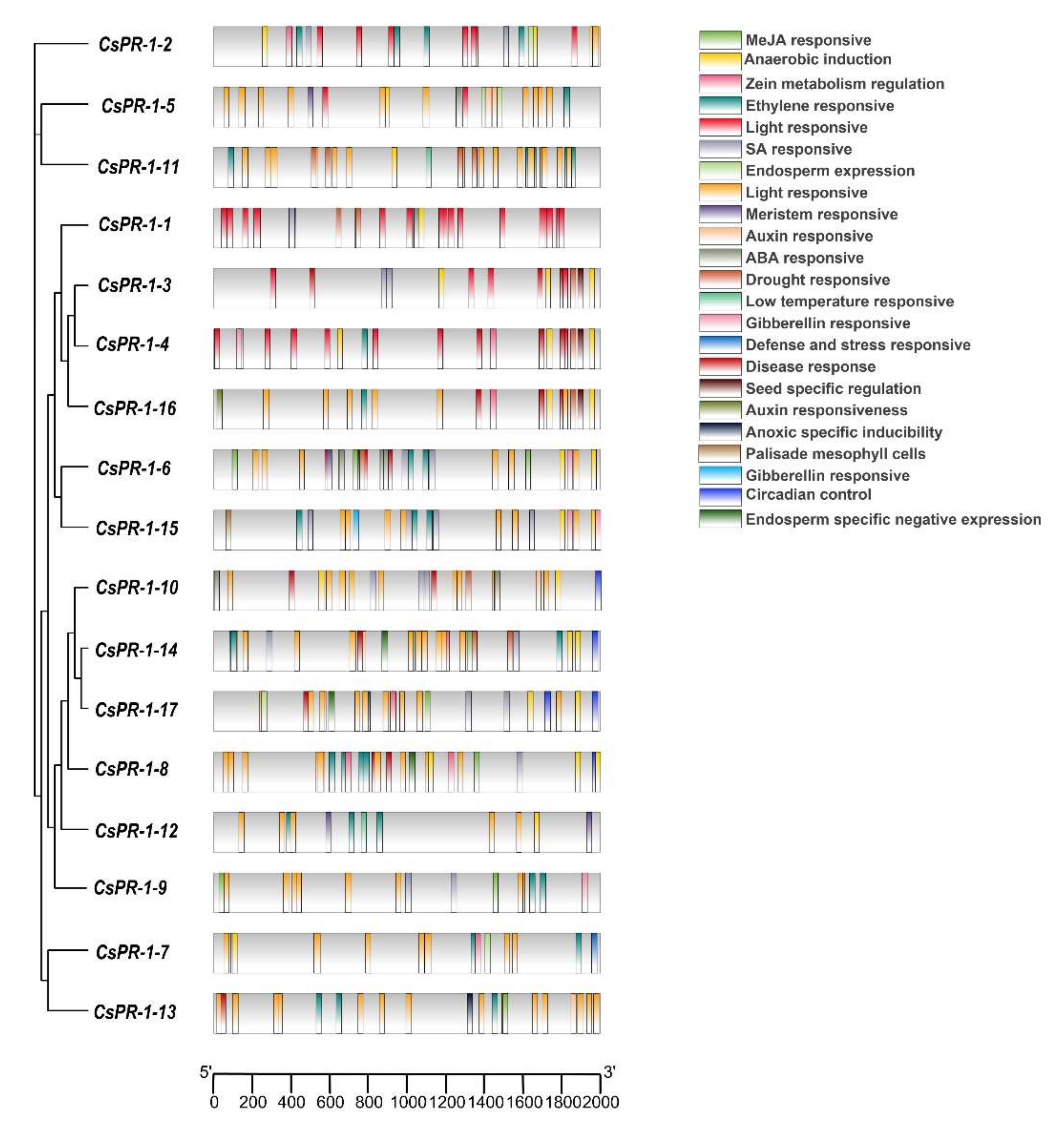
2.5. Analysis of Cis Elements of the CsPR-1 Genes
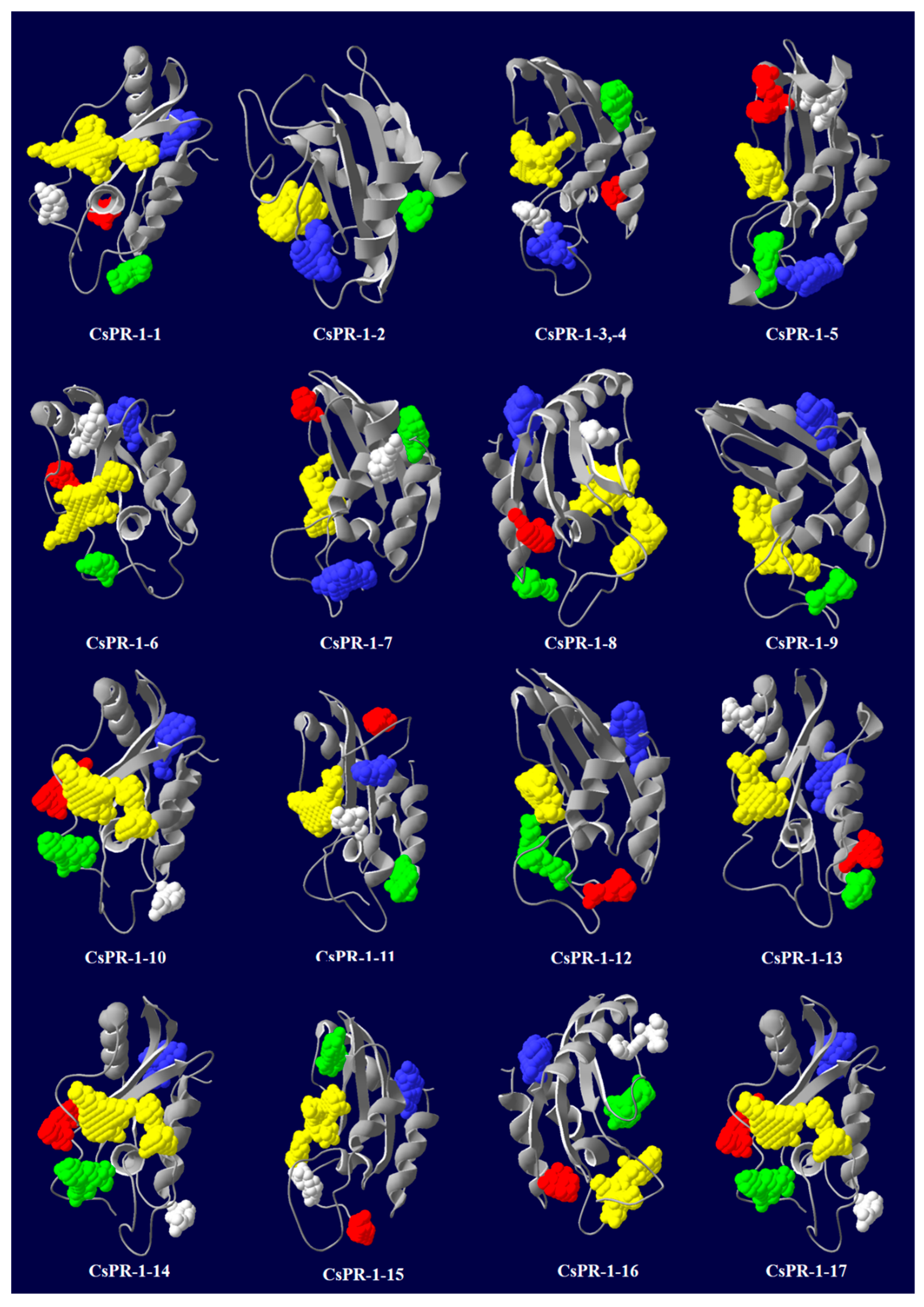
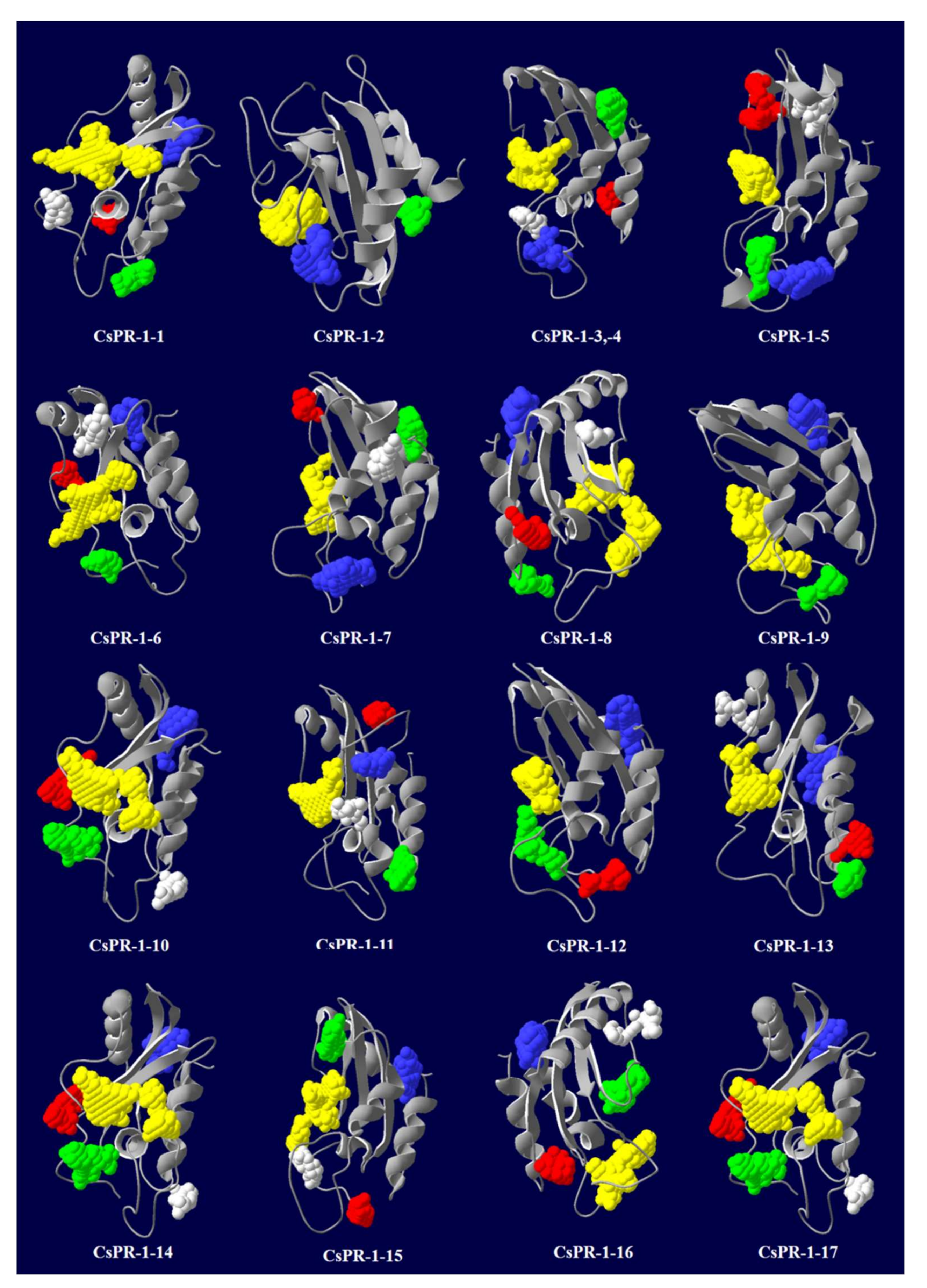
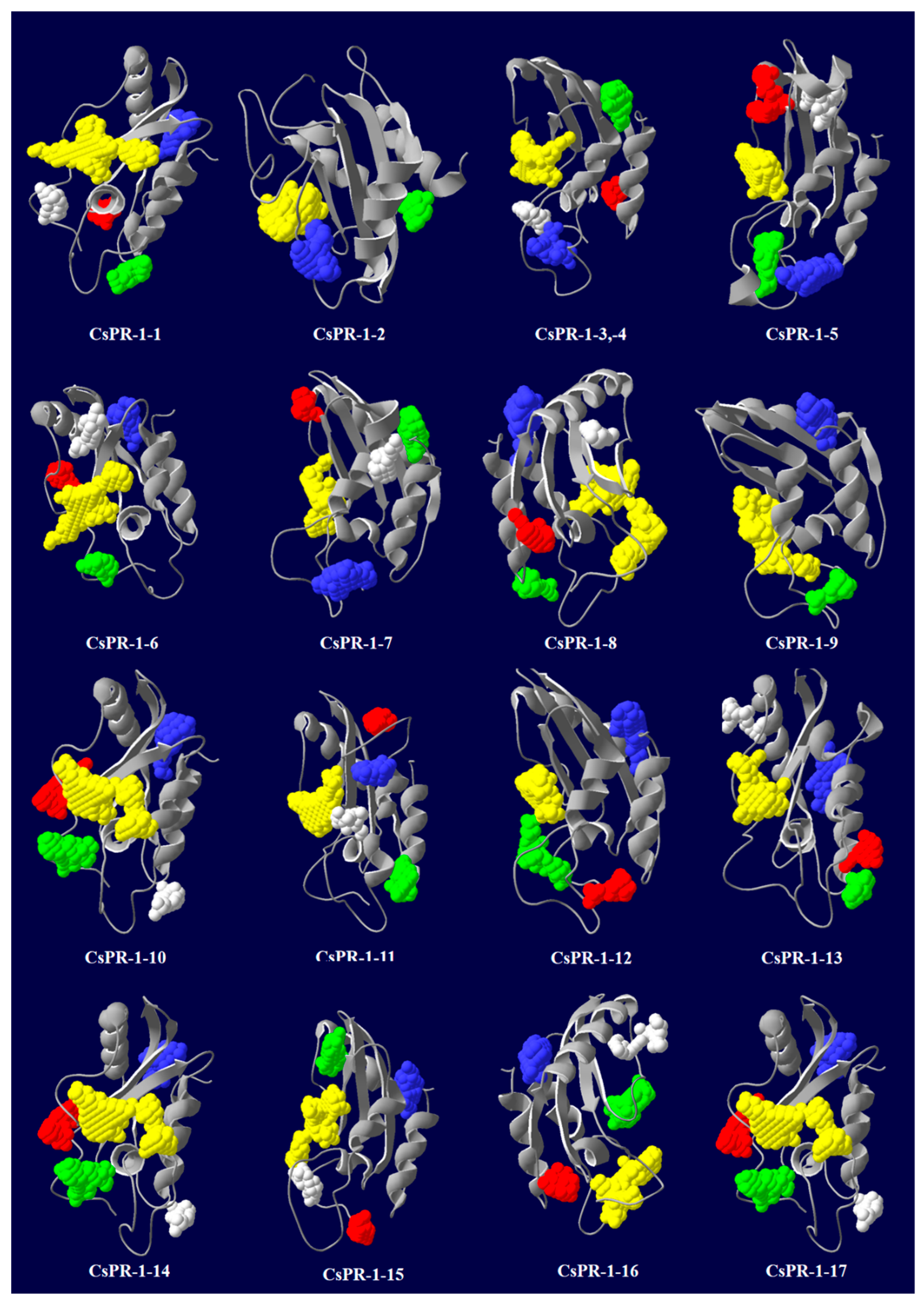
2.6. Predicted Secondary and 3D Structures of CsPR-1 Proteins
2.7. Histomorphological Observation of Tea Leaves
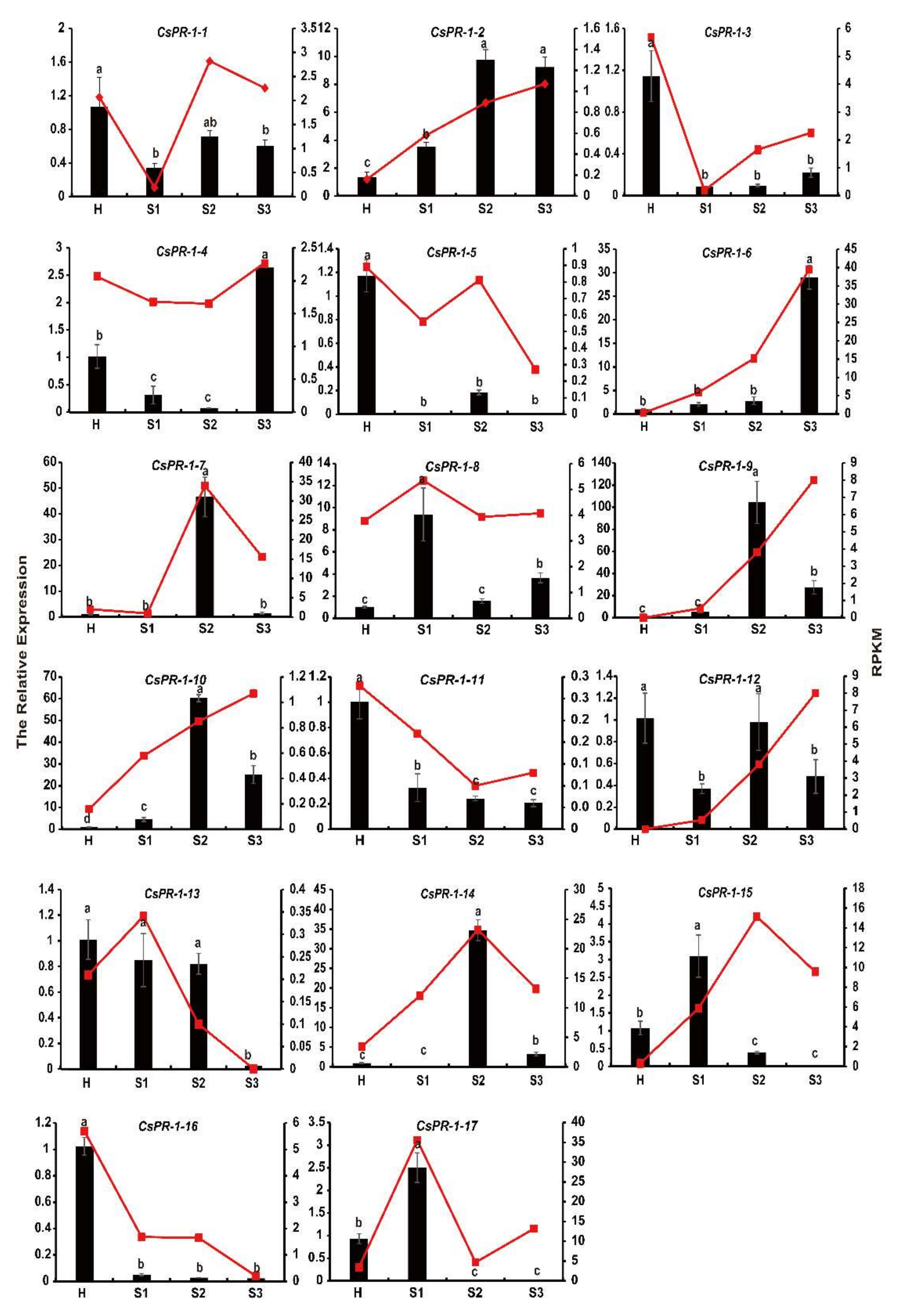
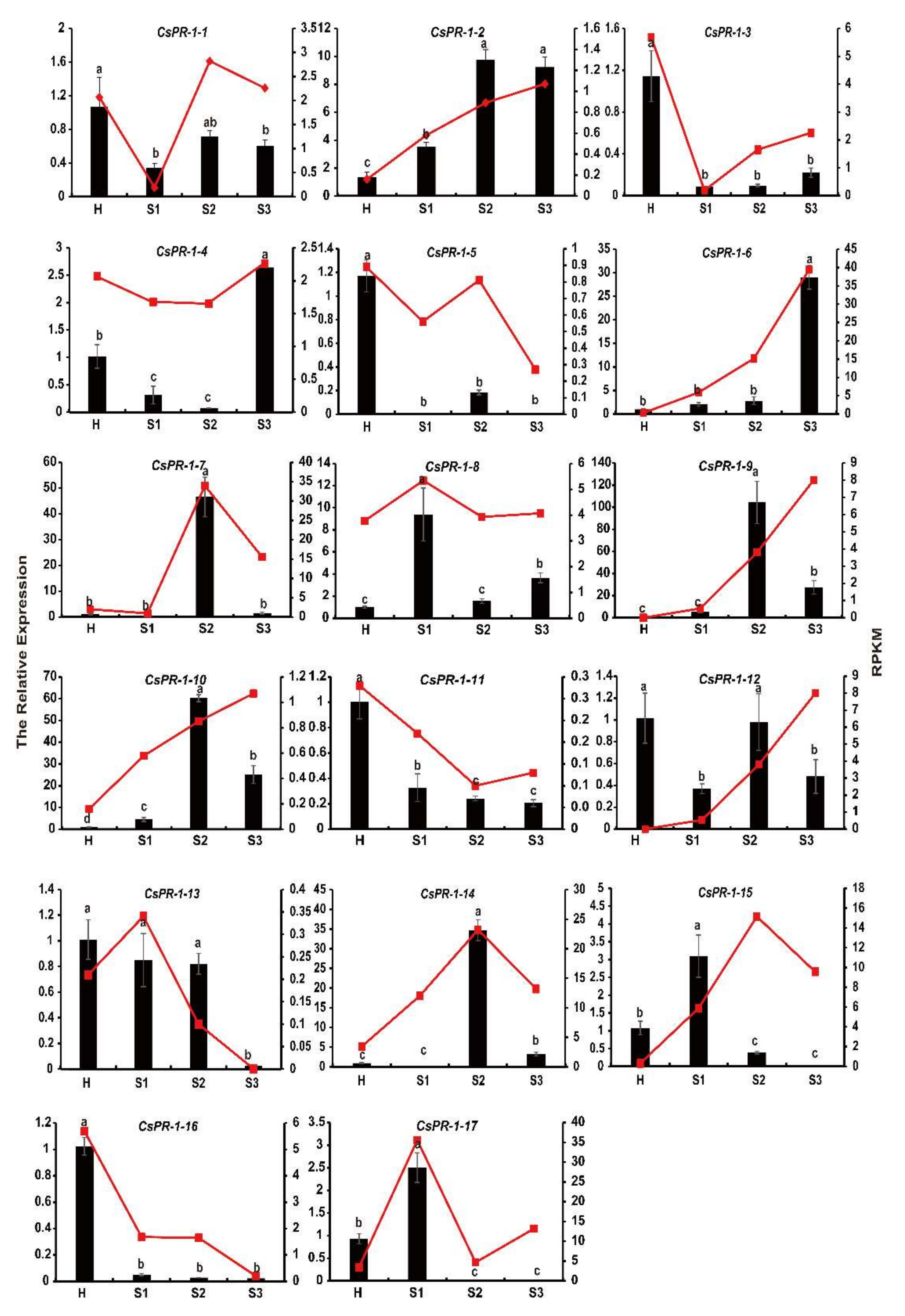
2.8. Expression Patterns of CsPR-1 Genes
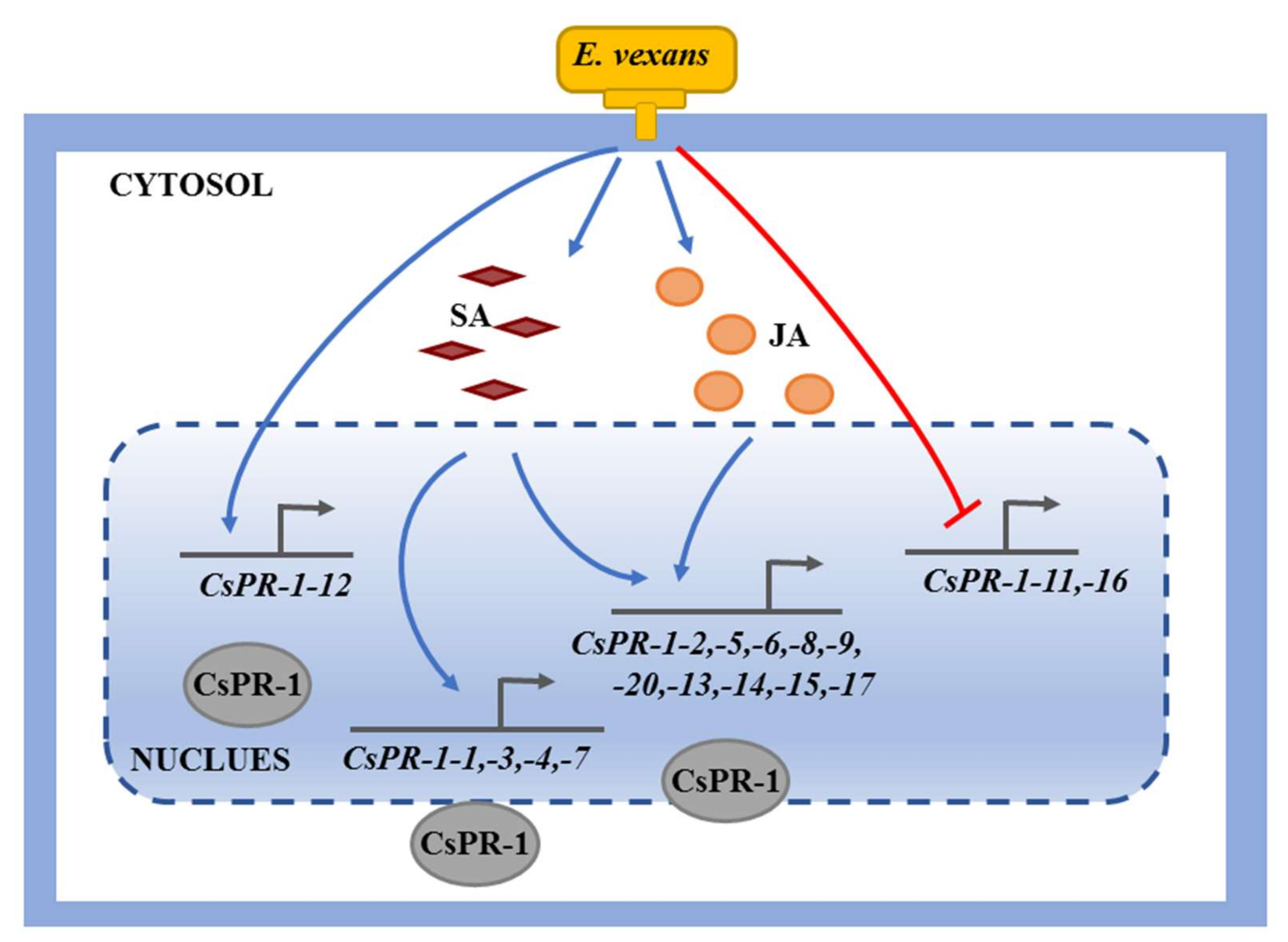
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Database Mining and Identification of CsPR-1 Genes
4.3. Analysis of the Conserved Motifs, Gene Structures, and Protein Functional Networks
4.4. Analysis of the Cis Elements of the CsPR-1 Gene Promoters
4.5. Structural Analysis of CsPR-1 Proteins
4.6. PAS Staining
4.7. RNA Extraction and Quantitative RT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Takken, F.; Tameling, W. To Nibble at Plant Resistance Proteins. Science 2009, 324, 744–746. [Google Scholar] [CrossRef] [PubMed]
- Bigeard, J.; Colcombet, J.; Hirt, H. Signaling Mechanisms in Pattern-Triggered Immunity (PTI). Mol. Plant 2015, 19, 521–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tena, G. PTI and ETI are One. Nat. Plants 2021, 7, 1527. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.S.; Arraes, F.B.M.; Campos, D.A.; Grossi-de-Sa, M.; Fernandez, D.; Cândido, E.S.; Cardoso, M.H.; Franco, O.L.; Grossi-de-Sa, M.F. Review: Potential Biotechnological Assets Related to Plant Immunity Modulation Applicable in Engineering Disease-Resistant Crops. Plant Sci. 2018, 270, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Hamamouch, N.; Chunying, L.I.; Seo, P.J.; Park, C.M. Expression of Arabidopsis Pathogenesis-Related Genes During Nematode Infection. Mol. Plant Pathol. 2011, 12, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Sung, Y.C.; Outram, M.A.; Breen, S.; Wang, C.; Dagvadorj, B.; Winterberg, B.; Kobe, B.; Williams, S.J.; Solomon, P.S. PR1-Mediated Defence via C-Terminal Peptide Release is Targeted by a Fungal Pathogen Effector. New Phytol. 2021, 229, 3467–3480. [Google Scholar] [CrossRef] [PubMed]
- Datta, K.; Tu, J.; Oliva, N.; Ona, I.; Velazhahan, R.; Mew, T.W.; Muthukrishnan, S.; Datta, S.K. Enhanced Resistance to Sheath Blight by Constitutive Expression of Infection-Related Rice Chitinase in Transgenic Elite Indica Rice Cultivars. Plant Sci. 2001, 160, 405–414. [Google Scholar] [CrossRef]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M.J. Significance of Inducible Defense-Related Proteins in Infected Plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, G.M.; Kim, R.; O’Bryan, M.K. The CAP Superfamily: Cysteine-Rich Secretory Proteins, Antigen 5, and Pathogenesis-Related 1 Proteins-Roles in Reproduction, Cancer, and Immune Defense. Endocr. Rev. 2008, 7, 865–897. [Google Scholar] [CrossRef]
- Sheng, J.; Olrichs, N.K.; Gadella, B.M.; Kaloyanova, D.V.; Helms, J.B. Regulation of Functional Protein Aggregation by Multiple Factors: Implications for the Amyloidogenic Behavior of the CAP Superfamily Proteins. Int. J. Mol. Sci. 2020, 21, 6530. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.Y.; Cheng, K.T.; Chang, W.H.; Huang, R.N.; Nam, H.G.; Chen, Y.R. Quantitative Peptidomics Study Reveals that a Wound-Induced Peptide from PR-1 Regulates Immune Signaling in Tomato. Plant Cell 2014, 26, 4135–4148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiraga, J.; Mackiewicz, P.; Mackiewicz, D.; Kowalczuk, M.; Biecek, P.; Polak, N.; Smolarczyk, K.; Dudek, M.R.; Cebrat, S. The Relationships Between theIsoelectric Point and: Length of Proteins, Taxonomy and Ecology of Organisms. BMC Genomics 2007, 8, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kattupalli, D.; Sreenivasan, A.; Soniya, E.V. A Genome-Wide Analysis of Pathogenesis-Related Protein-1 (PR-1) Genes from Piper nigrum Reveals Its Critical Role during Phytophthora capsici Infection. Genes 2021, 12, 1007. [Google Scholar] [CrossRef]
- Li, Z.T.; Dhekney, S.A.; Gray, D.J. PR-1 Gene Family of Grapevine: A Uniquely Duplicated PR-1 Gene from a Vitis Interspecific Hybrid Confers High Level Resistance to Bacterial Disease in Transgenic Tobacco. Plant Cell Rep. 2011, 30, 1–11. [Google Scholar] [CrossRef]
- Kiba, A.; Nishihara, M.; Nakatsuka, T.; Yamamure, S. Pathogenesis-Related Protein 1 Homologue is an Antifungal Protein in Wasabia japonica Leaves and Confers Resistance to Botrytis cinerea in Transgenic Tobacco. Plant Biotechnol. 2007, 24, 247–253. [Google Scholar] [CrossRef]
- Breen, S.; Williams, S.J.; Outram, M.; Kobe, B.; Solomon, P.S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends Plant Sci. 2017, 22, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Baruah, P.M.; Krishnatreya, D.B.; Bordoloi, K.S.; Gill, S.S.; Agarwalaa, N. Genome Wide Identification and Characterization of Abiotic Stress Responsive lncRnas in Capsicum annuum. Plant Physiol. Bioch. 2021, 162, 221–236. [Google Scholar] [CrossRef]
- Jiang, M.; Chen, H.; Liu, J.; Du, Q.; Lu, S.; Liu, C. Genome-Wide Identification and Functional Characterization of Natural Antisense Transcripts in Salvia miltiorrhiza. Sci. Rep. 2021, 11, 4769. [Google Scholar] [CrossRef]
- Kesawat, M.S.; Kherawat, B.S.; Singh, A.; Dey, P.; Kabi, M.; Debnath, D.; Saha, D.; Khandual, A.; Rout, S.; Manorama; et al. Genome-Wide Identification and Characterization of the Brassinazole-Resistant (BZR) Gene Family and Its Expression in the Various Developmental Stage and Stress Conditions in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2021, 22, 8743. [Google Scholar] [CrossRef]
- Kumar, M.; Kherawat, B.S.; Dey, P.; Saha, D.; Singh, A.; Bhatia, S.K.; Ghodake, G.S.; Kadam, A.A.; Kim, H.-U.; Manorama; et al. Genome-Wide Identification and Characterization of PIN-FORMED (PIN) Gene Family Reveals Role in Developmental and Various Stress Conditions in Triticum aestivum L. Int. J. Mol. Sci. 2021, 22, 7396. [Google Scholar] [CrossRef]
- Tong, T.; Fang, Y.; Zhang, Z.; Zheng, J.; Zhang, X.; Li, J.; Niu, C.; Xue, D.; Zhang, X. Genome-Wide Identification and Expression Pattern Analysis of the KCS Gene Family in Barley. Plant Growth Regul. 2021, 93, 89–103. [Google Scholar] [CrossRef]
- Wang, M.; Chen, B.; Zhou, W.; Xie, L.; Wang, L.; Zhang, Y.; Zhang, Q. Genome-Wide Identification and Expression Analysis of the AT-Hook Motif Nuclear Localized Gene Family in Soybean. BMC Genomics 2021, 22, 361. [Google Scholar] [CrossRef]
- Akbudak, M.A.; Yildiz, S.; Filiz, E. Pathogenesis Related Protein-1 (PR-1) Genes in Tomato (Solanum lycopersicum L.): Bioinformatics Analyses and Expression Profiles in Response to Drought Stress. Genomics 2020, 112, 4089–4099. [Google Scholar] [CrossRef]
- Lu, S.; Friesen, T.L.; Faris, J.D. Molecular Characterization and Genomic Mapping of the Pathogenesis-Related protein 1 (PR-1) Gene Family in Hexaploid Wheat (Triticum aestivum L.). Mol. Gen. Genet. 2011, 285, 485–503. [Google Scholar] [CrossRef] [PubMed]
- Almeida, S.F.; Venancio, T.M. Pathogenesis-Related Protein 1 (PR-1) Genes in Soybean: Genome-Wide Identification, Structural Analysis and Expression Profiling under Multiple Biotic and Abiotic Stresses. Gene. 2021, 809, 146013. [Google Scholar] [CrossRef] [PubMed]
- Higdon, J.V.; Frei, B. Tea Catechins and Polyphenols: Health Effects, Metabolism, and Antioxidant Functions. Crit. Rev. Food Sci. 2003, 43, 89–143. [Google Scholar] [CrossRef] [PubMed]
- Nagao, H. Effect of Aqueous Vitamin B on the Growth of Blister Blight Pathogen, Exobasidium vexans. Songklanakarin J. Sci. Technol. 2012, 34, 601–606. [Google Scholar]
- Punyasiri, P.A.; Abeysinghe, S.B.; Kumar, V. Preformed and Induced Chemical Resistance of Tea Leaf against Exobasidium vexans Infection. J. Chem. Ecol. 2005, 31, 1315–1324. [Google Scholar] [CrossRef] [Green Version]
- Ponmurugan, P.; Gnanamangai, B.M.; Manjukarunambika, K. Architectural Effect of Different Tea Clones on the Development of Blister Blight Disease. J. Appl. Bot. Food Qual. 2019, 92, 7–14. [Google Scholar]
- Jayaswall, K.; Mahajan, P.; Singh, G.; Parmar, R.; Seth, R.; Raina, A.; Swarnkar, M.K.; Singh, A.K.; Shankar, R.; Sharma, R.K. Transcriptome Analysis Reveals Candidate Genes Involved in Blister Blight Defense in Tea (Camellia sinensis (L) Kuntze). Sci. Rep. 2016, 6, 30412. [Google Scholar] [CrossRef] [Green Version]
- Chandra, S.; Chakraborty, N.; Panda, K.; Acharya, K. Chitosan-Induced Immunity in Camellia sinensis (L.) O. Kuntze against Blister Blight Disease is Mediated by Nitric-Oxide. Plant Physiol. Biochem. 2017, 115, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Niderman, T. Pathogenesis-Related PR-1 Proteins are Antifungal-Isolation and Characterization of 3 14-Kilodalton Proteins of Tomato and of a Basic PR-1 of Tobacco with Inhibitory Activity against Phytophthora -Infestans. Plant Physiol. 1995, 108, 17–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarowar, S.; Kim, Y.J.; Kim, E.N.; Kim, K.D.; Hwang, B.K.; Islam, R.; Shin, J.S. Overexpression of a Pepper Basic Pathogenesis-Related Protein 1 Gene in Tobacco Plants Enhances Resistance to Heavy Metal and Pathogen Stresses. Plant Cell Rep. 2005, 24, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Wang, Y.; Wei, W.; Lin, Y.; Yin, W.; Luo, C. Development of Novel Methods for Functional Evaluation of the Signal Peptide of Secreted Protein. Physiol. Mol. Plant P 2019, 106, 182–186. [Google Scholar] [CrossRef]
- Lu, S.; Faris, J.D.; Sherwood, R.; Friesen, T.L.; Edwards, M.C. A Dimeric PR-1-Type Pathogenesis-Related Protein Interacts with ToxA and Potentially Mediates ToxA-Induced Necrosis in Sensitive Wheat. Mol. Plant Pathol. 2014, 15, 650–663. [Google Scholar] [CrossRef]
- Pfitzner, U.M.; Goodman, H.M. Isolation and Characterization of cDNA Clones Encoding Pathogenesis-Related Proteins from Tobacco Mosaic Virus Infected Tobacco Plants. Nucleic Acids Res. 1987, 11, 4449–4465. [Google Scholar]
- Tornero, P.; Conejero, V.; Vera, P. A Gene Encoding a Novel Isoform of the PR-1 Protein Family from Tomato is Induced upon Viroid Infection. Mol. Gen. Genet. 1994, 243, 47–53. [Google Scholar] [CrossRef]
- Pazos, F.; Pietrosemoli, N.; García-Martín, J.A.; Solano, R. Protein Intrinsic Disorder in Plants. Front. Plant Sci. 2013, 4, 00363. [Google Scholar] [CrossRef] [Green Version]
- He, X.; Zhang, J. Rapid Subfunctionalization Accompanied by Prolonged and Substantial Neofunctionalization in Duplicate Gene Evolution. Genetics 2005, 169, 1157–1164. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Tong, G.; Wan, S.; Zhang, Y.; Yang, J.; Yu, Y.; Wang, W. Genome-Wide Identification, Classification and Expression Analysis of the HSP Gene Superfamily in Tea Plant (Camellia sinensis). Int. J. Mol. Sci. 2018, 19, 2633. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Xue, Q. Computational Identification of Novel PR-1-Type Genes in Oryza sativa. J. Genet. 2006, 85, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Mir, Z.A.; Tyagi, A.; Bhat, J.A.; Chandrashekar, N.; Papolu, P.K.; Rawat, S.; Grover, A. Identification and Comparative Analysis of Brassica juncea Pathogenesis-Related Genes in Response to Hormonal, Biotic and Abiotic Stresses. Acta Physiol. Plant 2017, 39, 268. [Google Scholar] [CrossRef]
- Gao, L.; Wang, S.; Li, X.Y. Expression and Functional Analysis of a Pathogenesis-Related Protein 1 Gene, TcLr19-PR1, Involved in Wheat Resistance against Leaf Rust Fungus. Plant Mol. Biol. Rep. 2015, 33, 797–805. [Google Scholar] [CrossRef]
- Seo, P.J.; Lee, A.K.; Xiang, F.; Park, C.M. Molecular and Functional Profiling of Arabidopsis Pathogenesis-Related Genes: Insights into Their Roles in Salt Response of Seed Germination. Plant Cell Physiol. 2008, 49, 334–344. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Wu, J.; Zhang, Y.; Yin, L.; Lu, J. Isolation of a WRKY30 Gene from Muscadinia rotundifolia (Michx) and Validation of Its Function Under Biotic and Abiotic Stresses. Protoplasma 2015, 252, 1361–1374. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, Z.; Gao, J.; Zhou, X.; Zhu, S.; Wang, X.Y.; Wang, X.L.; Ren, G.D.; Kuai, B.K. The NPR1-WRKY46-WRKY6 Signaling Cascade Mediates Probenazole/Salicylic Acid-Elicited Leaf Senescence in Arabidopsis thaliana. J. Integr. Plant Biol. 2021, 63, 924–936. [Google Scholar] [CrossRef]
- Xia, E.; Li, F.; Tong, W.; Li, P.; Wu, Q.; Zhao, H.; Ge, H.R.; Li, R.; Li, Y.; Zhang, Z. Tea Plant Information Archive: A Comprehensive Genomics and Bioinformatics Platform for Tea Plant. Plant Biotechnol. J. 2019, 17, 1938–1953. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and Analyzing DNA and Protein Sequence Motifs. Nucl. Acids Res. 2006, 34, 369–373. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C-T Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID | Gene | Chromosome | Protein | |||||
|---|---|---|---|---|---|---|---|---|
| Length (aa) | MW (kD) | pI | Gravy | SP | Subcellular Location | |||
| TEA022692.1 | CsPR1-1 | Chr04 | 163 | 18.07 | 7.08 | −0.353 | 24 | extr: 7, chlo: 6, cyto: 1 |
| TEA002521.1 | CsPR1-2 | Chr06 | 340 | 38.28 | 10.07 | −1.089 | 27 | vacu: 5, nucl: 3, cyto: 2, plas: 2, chlo: 1, E.R.: 1 |
| TEA017943.1 | CsPR1-3 | Chr06 | 162 | 17.93 | 8.74 | −0.213 | 24 | extr: 6, vacu: 5, chlo: 2, golg: 1 |
| TEA021774.1 | CsPR1-4 | Chr13 | 162 | 17.93 | 8.74 | −0.213 | 24 | extr: 6, vacu: 5, chlo: 2, golg: 1 |
| TEA022146.1 | CsPR1-5 | Chr01 | 173 | 18.61 | 6.81 | −0.227 | 21 | extr: 4, chlo: 3, vacu: 3, cyto: 2, plas: 2 |
| TEA004542.1 | CsPR1-6 | Chr06 | 160 | 17.59 | 4.96 | −0.235 | 24 | extr: 12, vacu: 2 |
| TEA022585.1 | CsPR1-7 | Chr08 | 170 | 19.25 | 9.22 | −0.375 | 24 | chlo: 12, extr: 1, vacu: 1 |
| TEA028234.1 | CsPR1-8 | Chr01 | 162 | 18.26 | 9.16 | −0.399 | 24 | chlo: 4, vacu: 3, extr: 2, E.R.: 2, nucl: 1, mito: 1, plas: 1 |
| TEA004551.1 | CsPR1-9 | Chr06 | 159 | 17.52 | 8.47 | −0.27 | 22 | chlo: 14 |
| TEA028218.1 | CsPR1-10 | Chr01 | 156 | 17.37 | 4.84 | −0.406 | 28 | chlo: 11, cyto: 2, nucl: 1 |
| TEA022150.1 | CsPR1-11 | Chr01 | 191 | 21.53 | 9.16 | −0.34 | \ | vacu: 6, chlo: 4, extr: 2, nucl: 1, plas: 1 |
| TEA030748.1 | CsPR1-12 | unkown | 204 | 22.60 | 5.66 | −0.257 | \ | E.R.: 5.5, cyto: 5, E.R._plas: 3.5, nucl: 1, mito: 1, vacu: 1 |
| TEA025681.1 | CsPR1-13 | Chr03 | 196 | 22.61 | 6.09 | −0.479 | 23 | extr: 7, chlo: 4, E.R.: 2, vacu: 1 |
| TEA022240.1 | CsPR1-14 | unkown | 162 | 17.88 | 6.86 | −0.297 | 24 | chlo: 13, vacu: 1 |
| TEA021361.1 | CsPR1-15 | Chr03 | 160 | 17.62 | 5.38 | −0.269 | 24 | extr: 11, vacu: 2, cyto: 1 |
| TEA011597.1 | CsPR1-16 | unkown | 168 | 17.94 | 8.57 | −0.213 | 24 | chlo: 6, vacu: 5, extr: 2, golg: 1 |
| TEA004541.1 | CsPR1-17 | Chr06 | 162 | 17.87 | 8.62 | −0.345 | 24 | extr: 7, vacu: 4, chlo: 2, mito: 1 |
| Proteins | α-Helices (%) | Extended Strand (%) | Beta Turn (%) | Random Coil (%) | Ramachandran Plot (%) | Number of Channels | Disordered Regions (%) |
|---|---|---|---|---|---|---|---|
| CsPR-1-1 | 38.65 | 13.50 | 4.29 | 43.56 | 98.3 | 2 | 23.93 |
| CsPR-1-2 | 23.82 | 8.24 | 4.12 | 63.82 | 99.2 | 2 | 61.76 |
| CsPR-1-3 | 37.04 | 14.20 | 6.17 | 42.59 | 98.3 | 4 | 24.07 |
| CsPR-1-4 | 37.04 | 14.20 | 6.17 | 42.59 | 98.3 | 4 | 24.07 |
| CsPR-1-5 | 30.64 | 17.34 | 1.73 | 50.29 | 96.0 | 3 | 20.23 |
| CsPR-1-6 | 29.38 | 20.62 | 4.38 | 45.62 | 98.3 | 3 | 23.75 |
| CsPR-1-7 | 29.41 | 17.65 | 4.71 | 48.24 | 97.4 | 3 | 6.47 |
| CsPR-1-8 | 32.72 | 16.05 | 2.47 | 48.77 | 99.1 | 2 | 17.28 |
| CsPR-1-9 | 33.33 | 15.09 | 3.77 | 47.80 | 99.1 | 3 | 18.24 |
| CsPR-1-10 | 36.54 | 16.67 | 7.05 | 39.74 | 99.1 | 3 | 16.03 |
| CsPR-1-11 | 39.27 | 15.18 | 4.71 | 40.84 | 96.7 | 5 | 9.42 |
| CsPR-1-12 | 32.21 | 17.45 | 6.04 | 44.30 | 99.1 | 3 | 22.15 |
| CsPR-1-13 | 28.57 | 22.45 | 3.06 | 45.92 | 96.7 | 9 | 16.33 |
| CsPR-1-14 | 33.95 | 15.43 | 4.32 | 46.30 | 99.1 | 3 | 17.28 |
| CsPR-1-15 | 38.12 | 16.25 | 4.38 | 41.25 | 98.3 | 3 | 27.50 |
| CsPR-1-16 | 39.88 | 11.31 | 4.76 | 44.05 | 97.5 | 6 | 32.74 |
| CsPR-1-17 | 37.04 | 16.05 | 4.32 | 42.59 | 99.1 | 3 | 18.52 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Q.; Guo, N.; Zhang, Y.; Yu, Y.; Liu, S. Genome-Wide Characterization and Expression Analysis of Pathogenesis-Related 1 (PR-1) Gene Family in Tea Plant (Camellia sinensis (L.) O. Kuntze) in Response to Blister-Blight Disease Stress. Int. J. Mol. Sci. 2022, 23, 1292. https://doi.org/10.3390/ijms23031292
Zhang Q, Guo N, Zhang Y, Yu Y, Liu S. Genome-Wide Characterization and Expression Analysis of Pathogenesis-Related 1 (PR-1) Gene Family in Tea Plant (Camellia sinensis (L.) O. Kuntze) in Response to Blister-Blight Disease Stress. International Journal of Molecular Sciences. 2022; 23(3):1292. https://doi.org/10.3390/ijms23031292
Chicago/Turabian StyleZhang, Qiqi, Nini Guo, Yongheng Zhang, Youben Yu, and Shuyuan Liu. 2022. "Genome-Wide Characterization and Expression Analysis of Pathogenesis-Related 1 (PR-1) Gene Family in Tea Plant (Camellia sinensis (L.) O. Kuntze) in Response to Blister-Blight Disease Stress" International Journal of Molecular Sciences 23, no. 3: 1292. https://doi.org/10.3390/ijms23031292
APA StyleZhang, Q., Guo, N., Zhang, Y., Yu, Y., & Liu, S. (2022). Genome-Wide Characterization and Expression Analysis of Pathogenesis-Related 1 (PR-1) Gene Family in Tea Plant (Camellia sinensis (L.) O. Kuntze) in Response to Blister-Blight Disease Stress. International Journal of Molecular Sciences, 23(3), 1292. https://doi.org/10.3390/ijms23031292