
Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis stolonifera

 ,
, 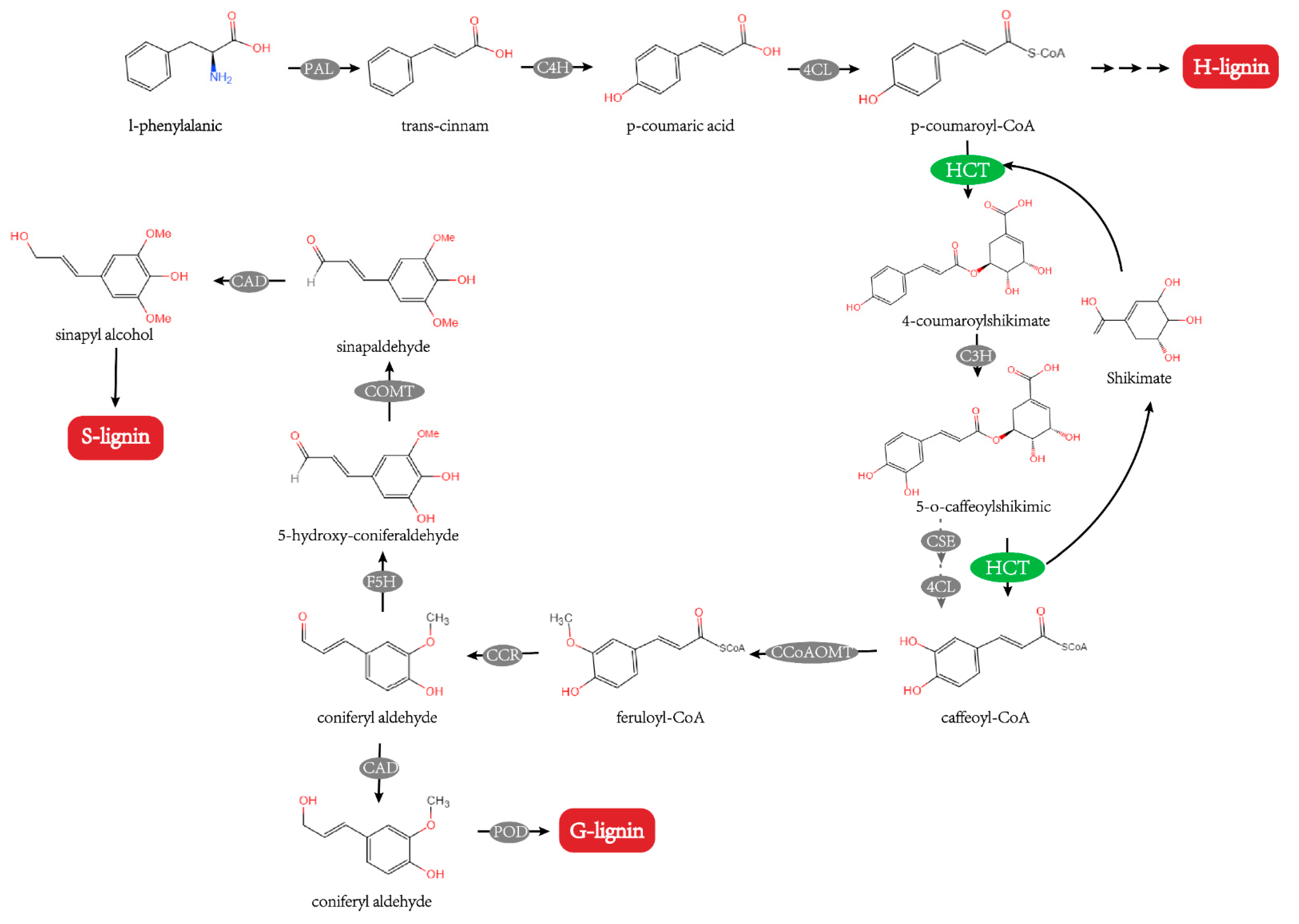{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
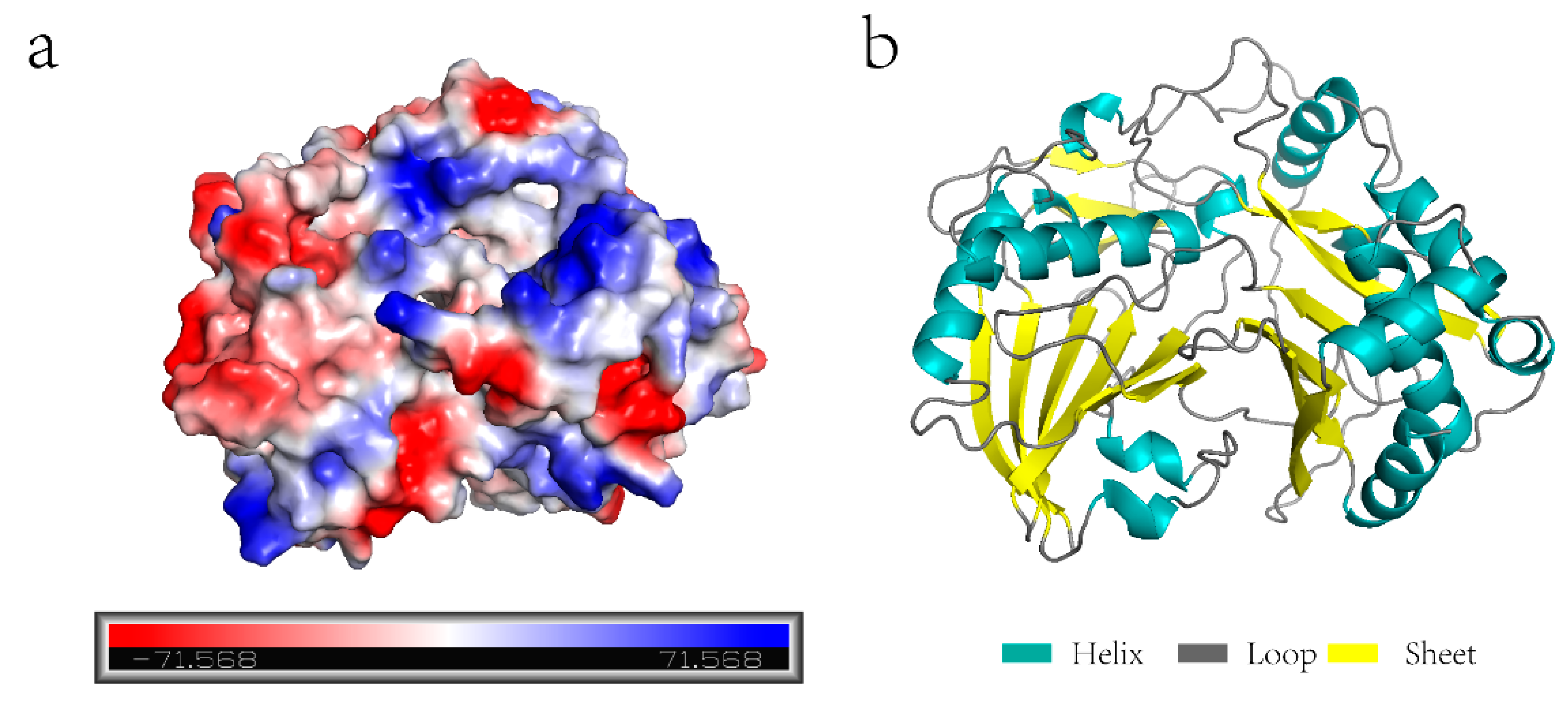
2.1. Identification and Bioinformatic Analysis of the ZjHCT4
2.2. Upstream Sequence Analysis of ZjHCT4
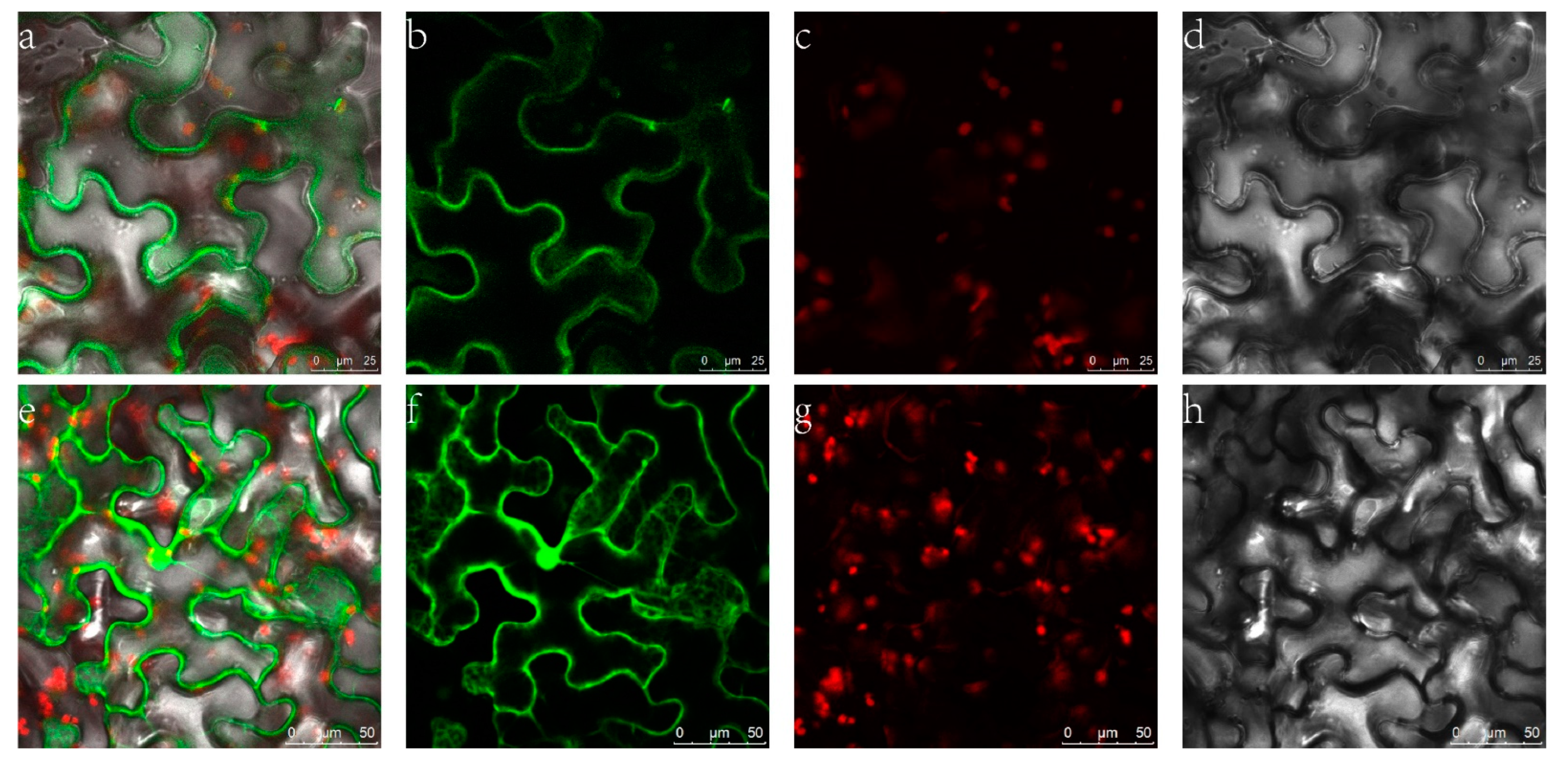
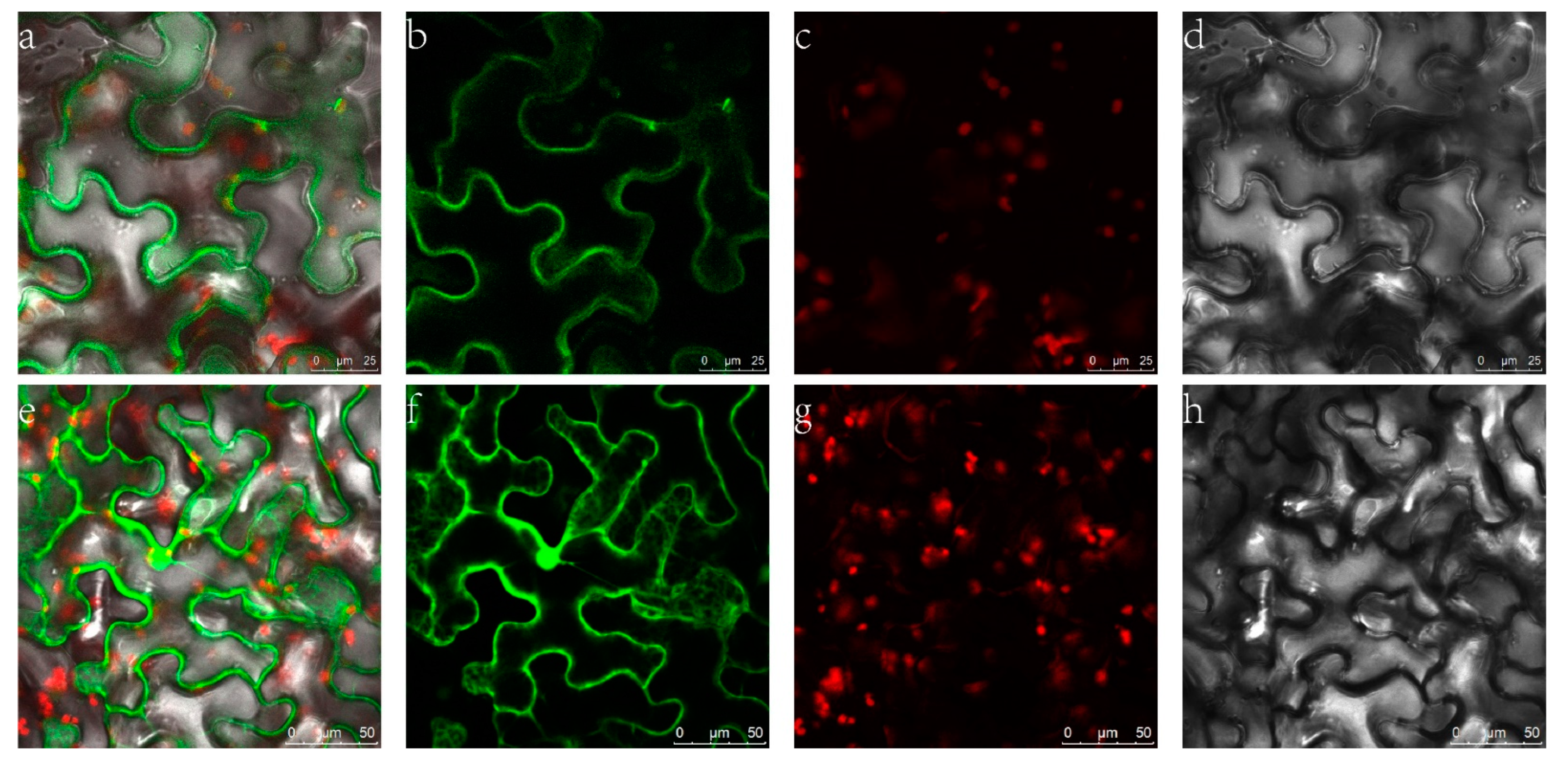
2.3. Subcellular Localization
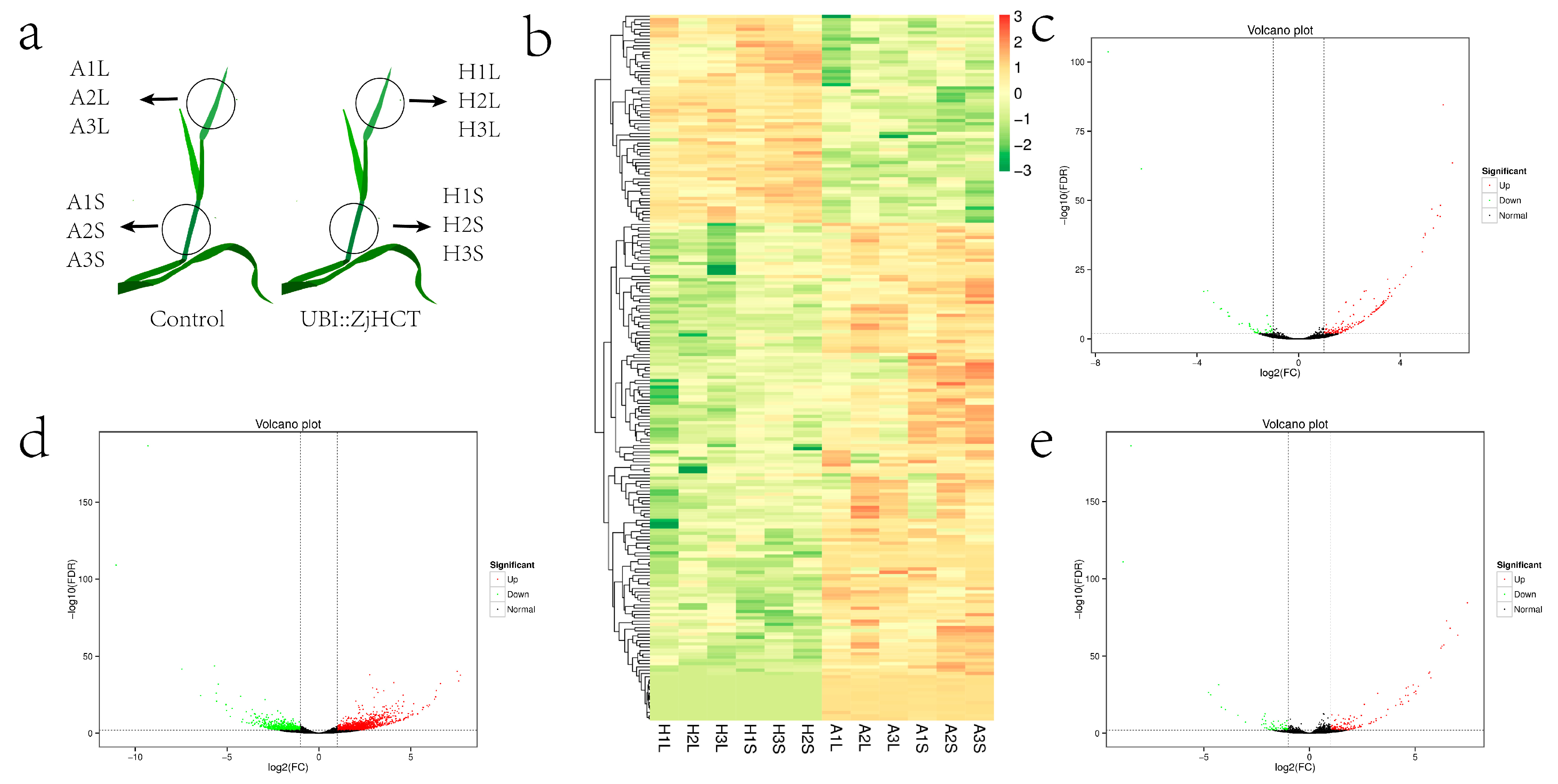
2.4. Ectopic Expression of ZjHCT4 in Creeping Bentgrass
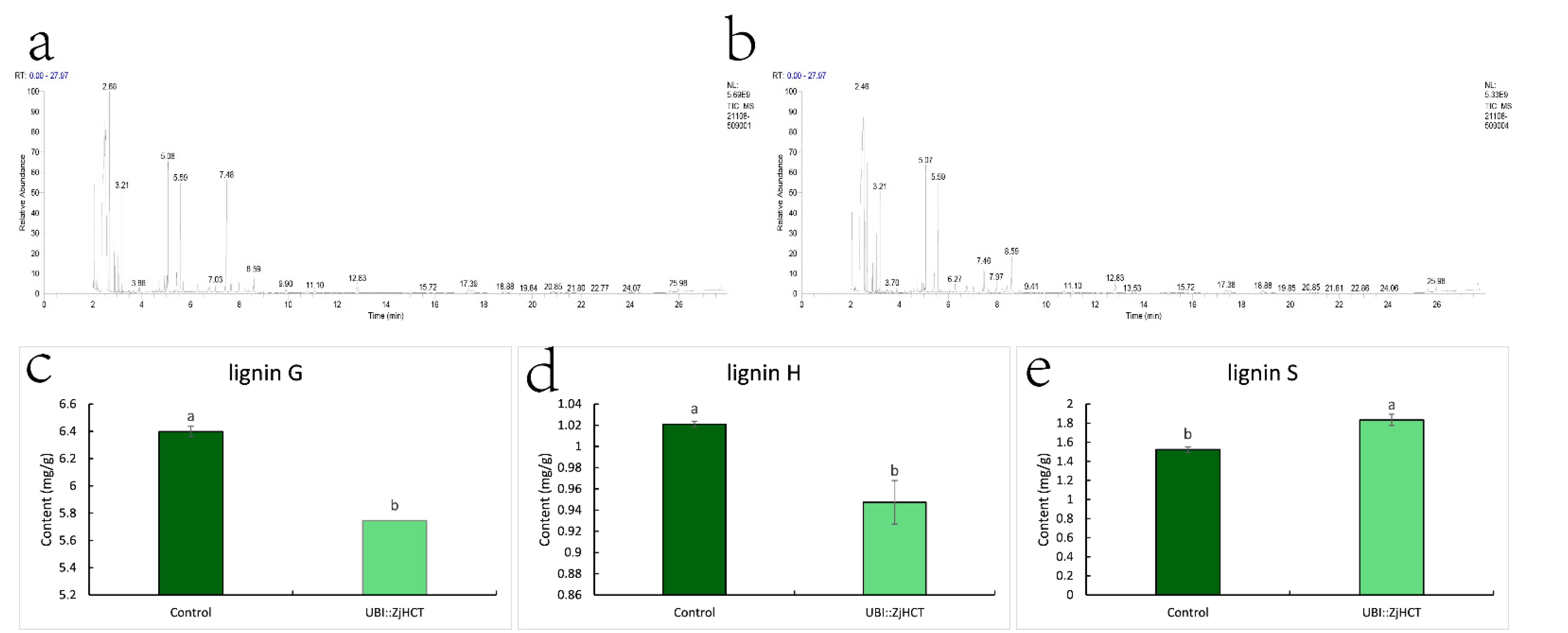
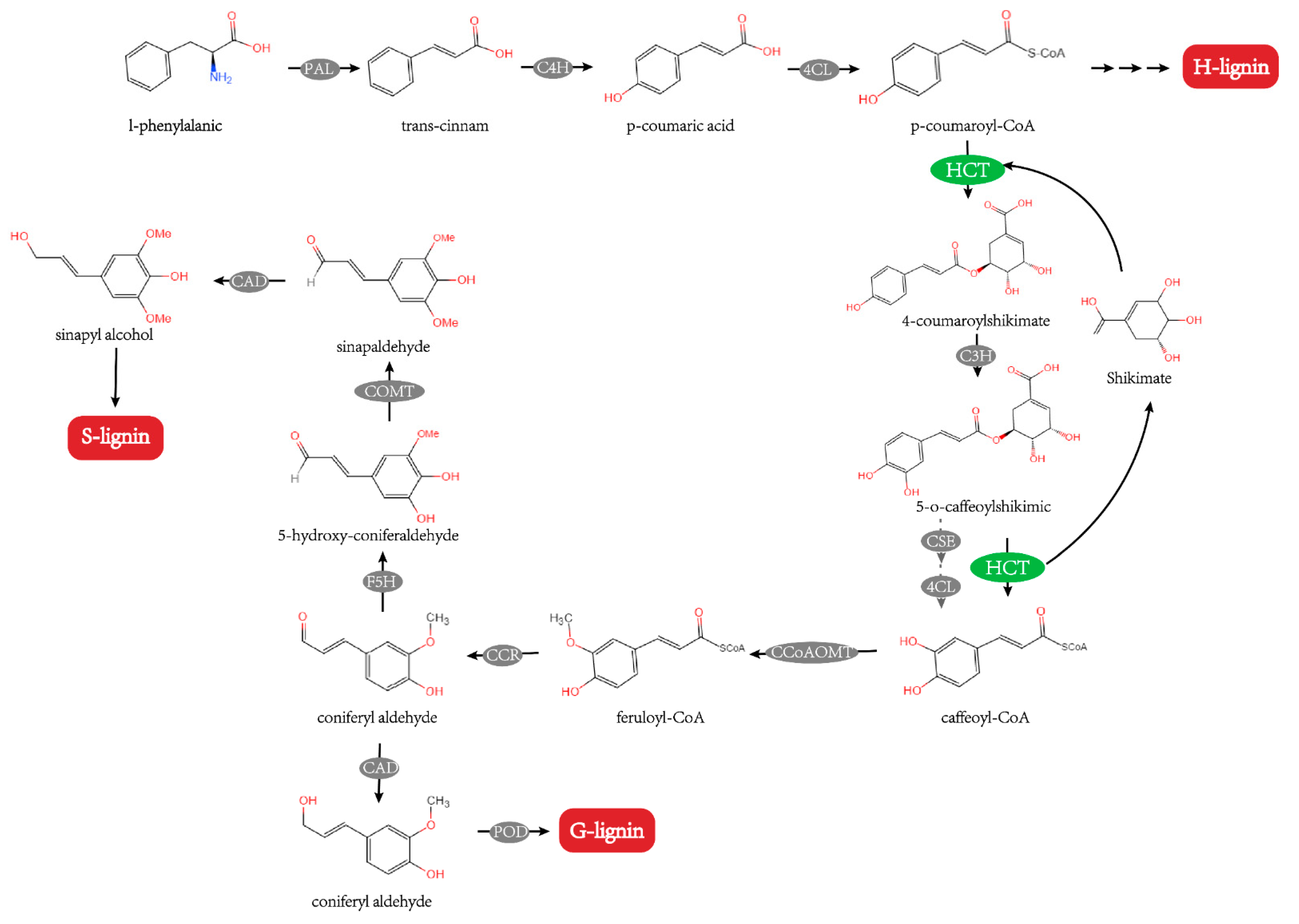
2.5. Determination of Monomer Content of Lignin
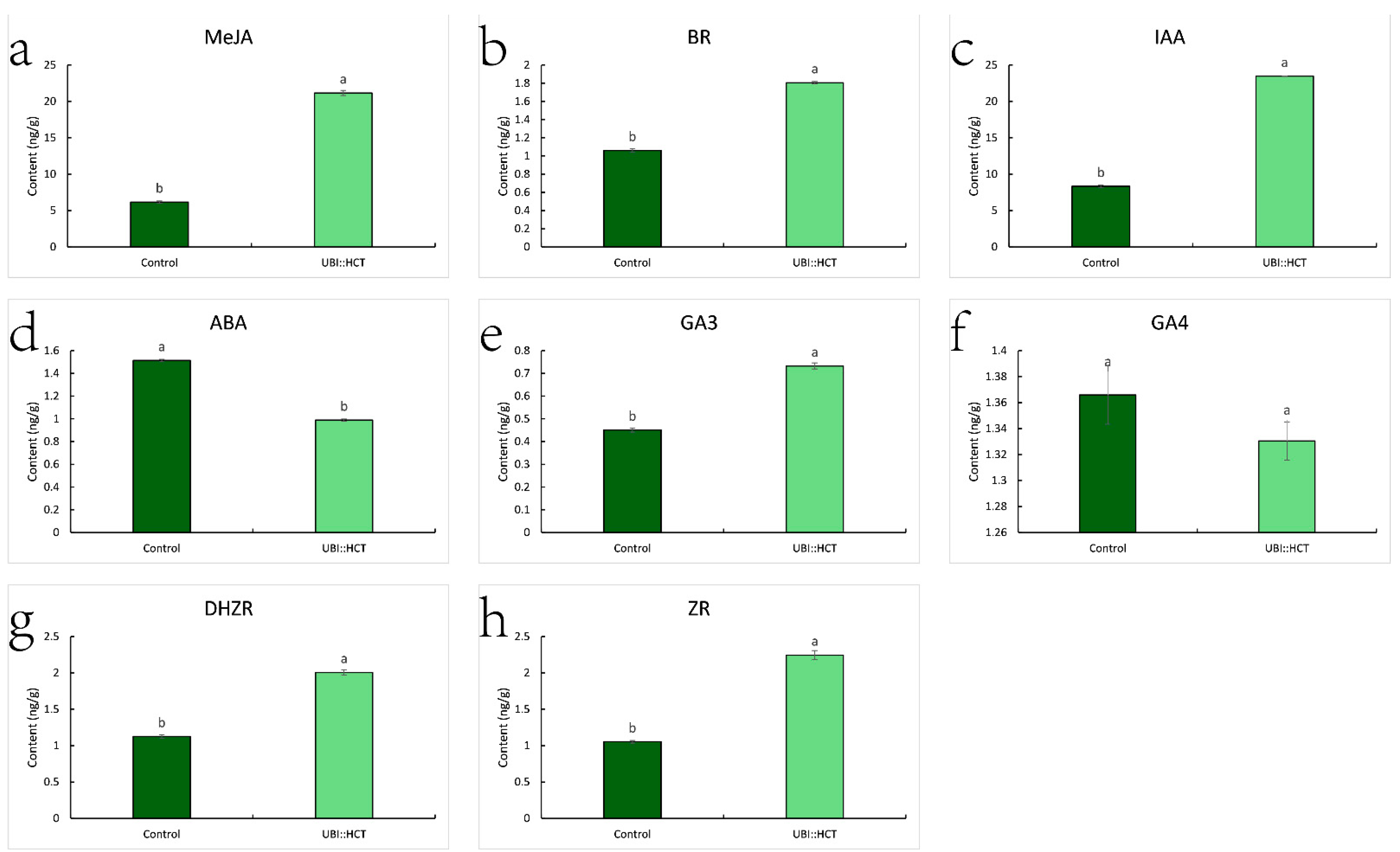
2.6. Ecpotic Expression of ZjHCT4 Causes Hormonal Changes
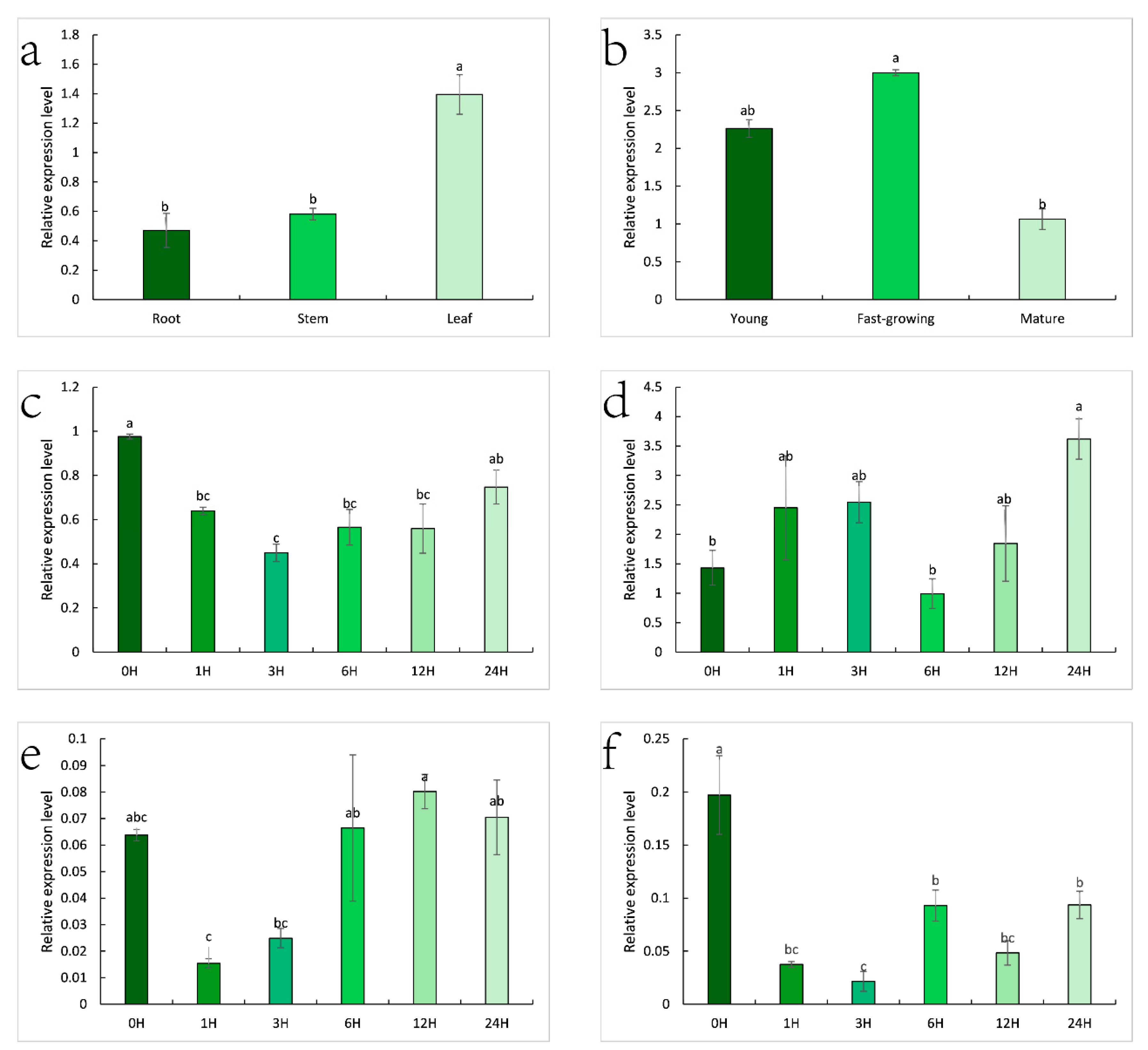
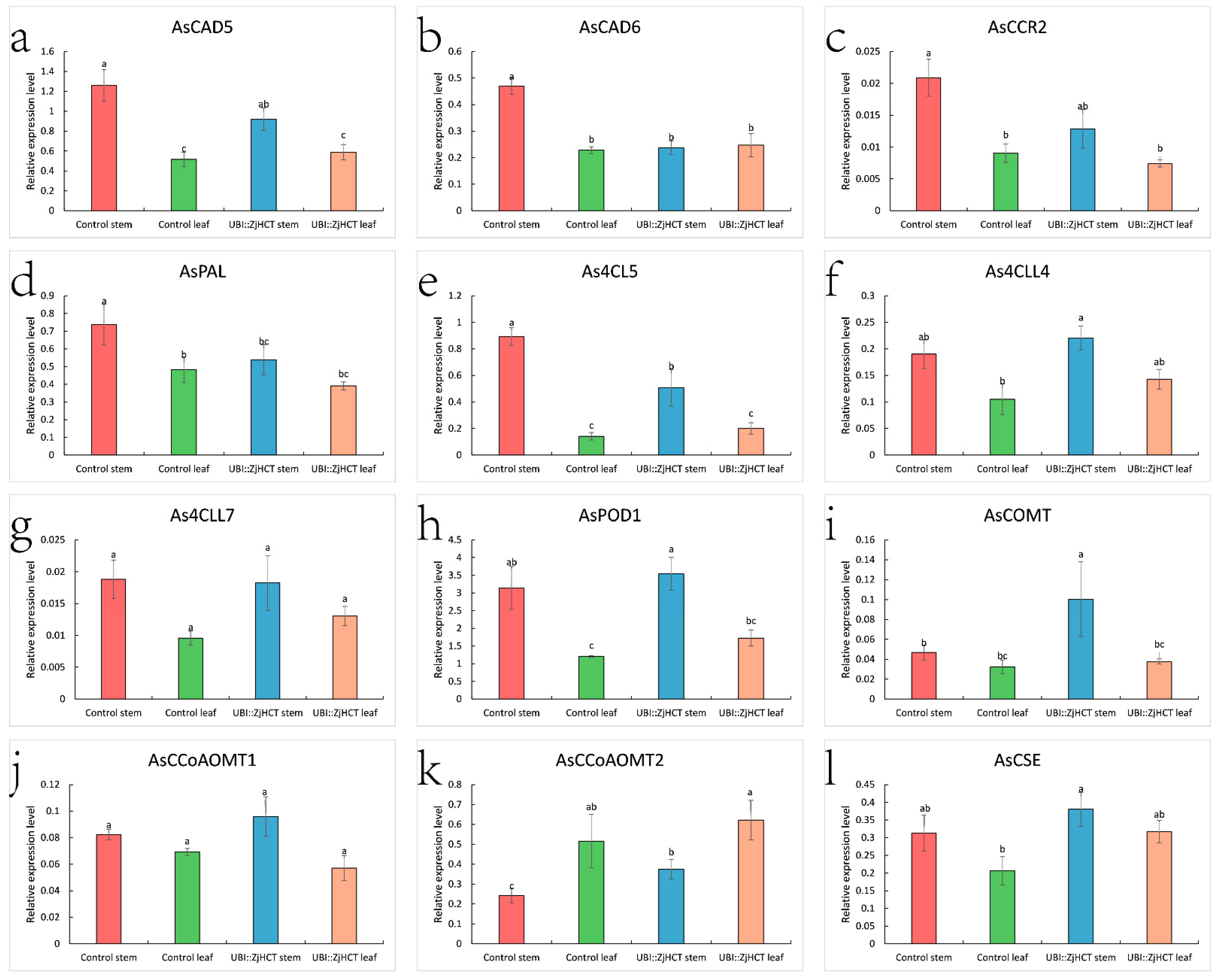
2.7. Expression Pattern of ZjHCT4 and the Lignin-Related Genes
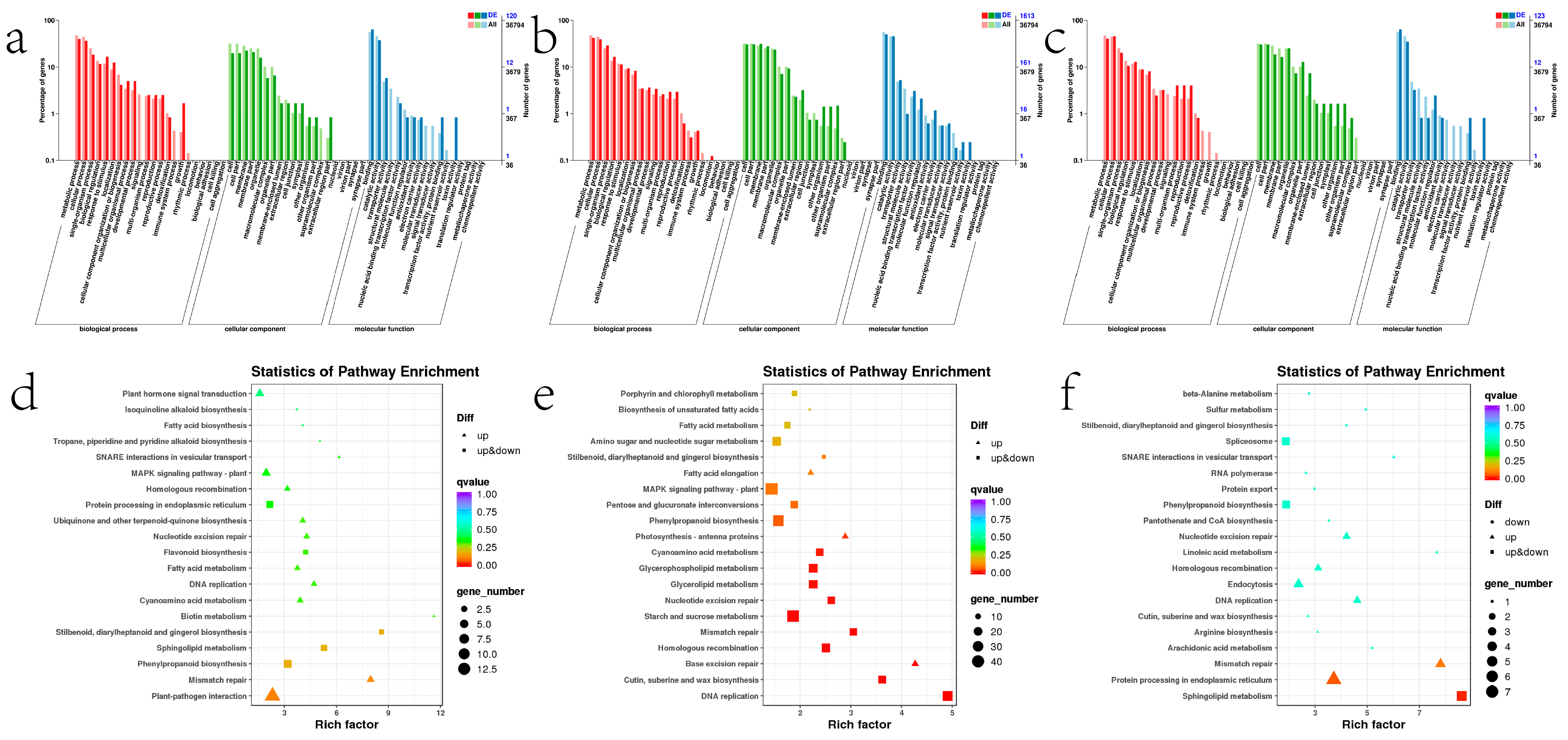
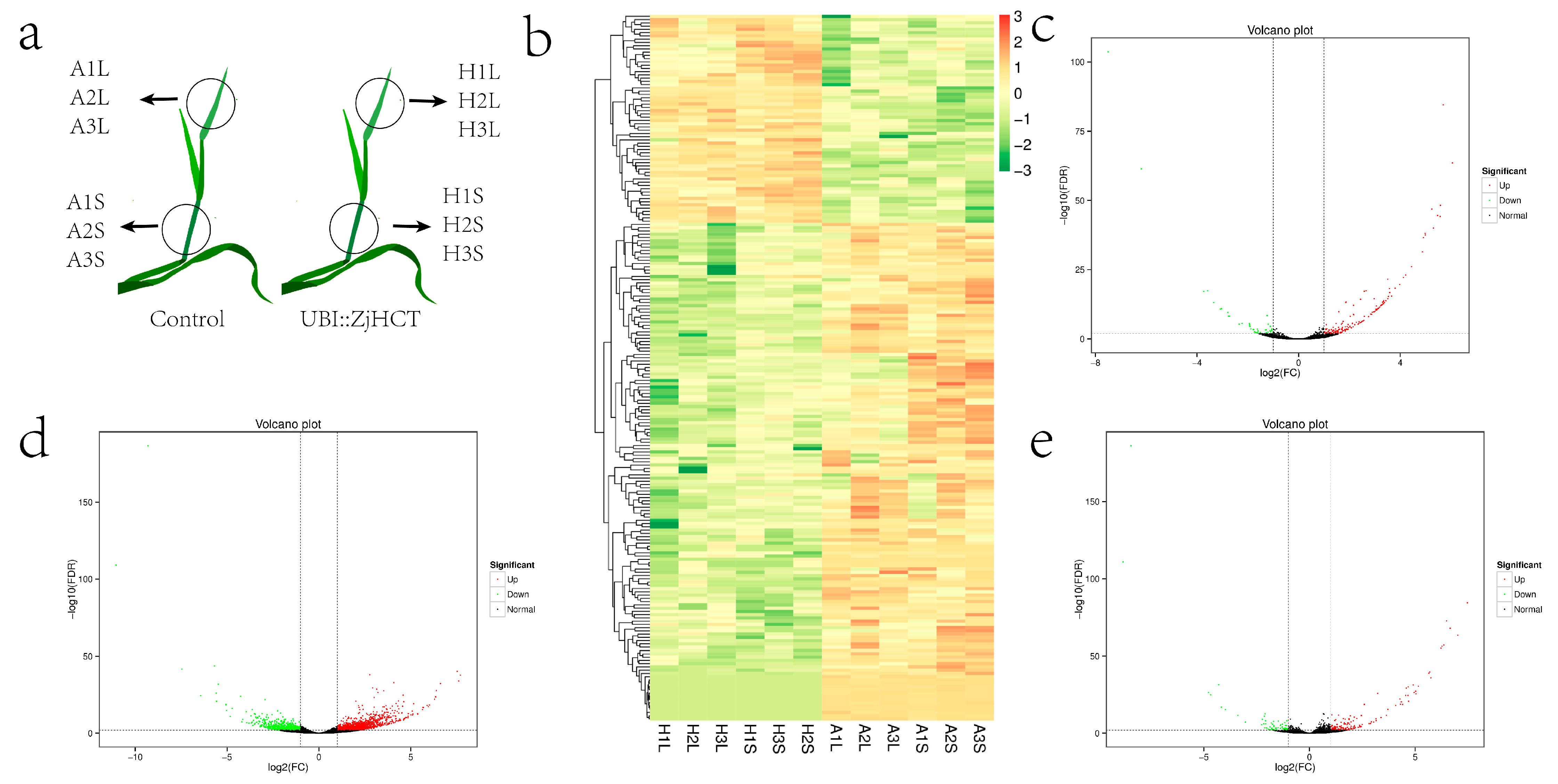
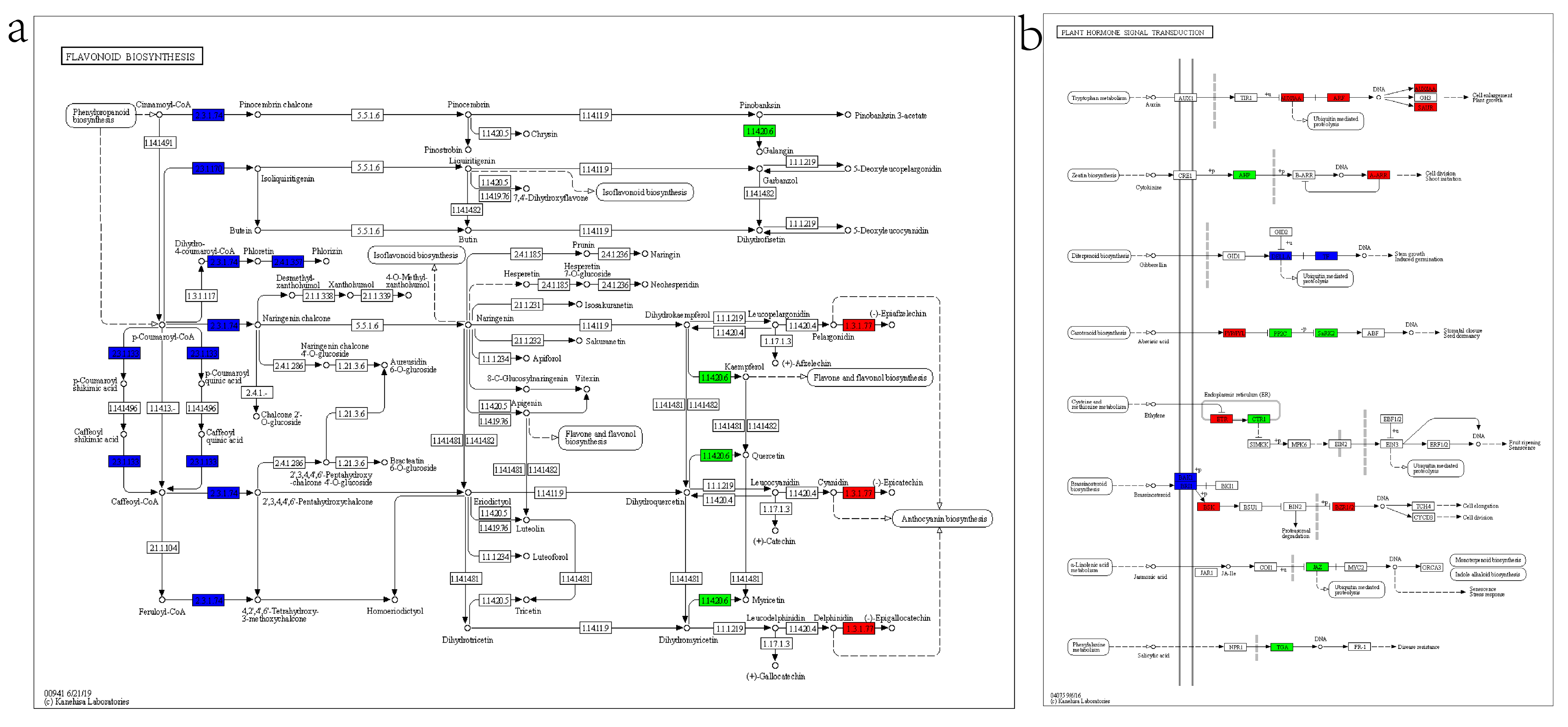
2.8. Analysis of Differentially Expressed Genes
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Cloning of ZjHCT4 and Vector Construction
4.3. Generation of Transgenic Plants
4.4. Subcellular Localization Analysis
4.5. Bioinformatics Analysis
4.6. Expression Levels of ZjHCT4 Gene and Lignin-Related Genes
4.7. Hormone Contents Assay
4.8. Transcriptomic Analysis of DEGs
4.9. Determination of Lignin Composition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, A.M.; Hayes, R.P.; Youn, B.; Vermerris, W.; Sattler, S.E.; Kang, C. Elucidation of the Structure and Reaction Mechanism of Sorghum Hydroxycinnamoyltransferase and Its Structural Relationship to Other Coenzyme A-Dependent Transferases and Synthases. Plant Physiol. 2013, 162, 640–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin Biosynthesis and Structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, R.A.; Barros, J. Lignin Biosynthesis: Old Roads Revisited and New Roads Explored. Open Biol. 2019, 9, 190215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin Biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519. [Google Scholar] [CrossRef]
- Weng, J.; Chapple, C. The Origin and Evolution of Lignin Biosynthesis. New Phytol. 2010, 187, 273–285. [Google Scholar] [CrossRef]
- Kriegshauser, L.; Knosp, S.; Grienenberger, E.; Tatsumi, K.; Gütle, D.D.; Sørensen, I.; Herrgott, L.; Zumsteg, J.; Rose, J.K.C.; Reski, R.; et al. Function of the HYDROXYCINNAMOYL-CoA:SHIKIMATE HYDROXYCINNAMOYL TRANSFERASE Is Evolutionarily Conserved in Embryophytes. Plant Cell 2021, 33, 1472–1491. [Google Scholar] [CrossRef]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, Cloning, and Properties of an Acyltransferase Controlling Shikimate and Quinate Ester Intermediates in Phenylpropanoid Metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef] [Green Version]
- del Ramirez-Ahumada, M.C.; Timmermann, B.N.; Gang, D.R. Biosynthesis of Curcuminoids and Gingerols in Turmeric (Curcuma Longa) and Ginger (Zingiber Officinale): Identification of Curcuminoid Synthase and Hydroxycinnamoyl-CoA Thioesterases. Phytochemistry 2006, 67, 2017–2029. [Google Scholar] [CrossRef]
- Shadle, G.; Chen, F.; Reddy, M.; Jackson, L.; Nakashima, J.; Dixon, R.A. Down-Regulation of Hydroxycinnamoyl CoA: Shikimate Hydroxycinnamoyl Transferase in Transgenic Alfalfa Affects Lignification, Development and Forage Quality. Phytochemistry 2007, 68, 1521–1529. [Google Scholar] [CrossRef]
- Gallego-Giraldo, L.; Jikumaru, Y.; Kamiya, Y.; Tang, Y.; Dixon, R.A. Selective Lignin Downregulation Leads to Constitutive Defense Response Expression in Alfalfa (Medicago Sativa L.). New Phytol. 2011, 190, 627–639. [Google Scholar] [CrossRef]
- Serrani-Yarce, J.C.; Escamilla-Trevino, L.; Barros, J.; Gallego-Giraldo, L.; Pu, Y.; Ragauskas, A.; Dixon, R.A. Targeting Hydroxycinnamoyl CoA: Shikimate Hydroxycinnamoyl Transferase for Lignin Modification in Brachypodium Distachyon. Biotechnol. Biofuels 2021, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A.; Ralph, J.; Akiyama, T.; Flint, H.; Phillips, L.; Torr, K.; Nanayakkara, B.; Te Kiri, L. Exploring Lignification in Conifers by Silencing Hydroxycinnamoyl-CoA:Shikimate Hydroxycinnamoyltransferase in Pinus Radiata. Proc. Natl. Acad. Sci. USA 2007, 104, 11856–11861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escamilla-Treviño, L.L.; Shen, H.; Hernandez, T.; Yin, Y.; Xu, Y.; Dixon, R.A. Early lignin pathway enzymes and routes to chlorogenic acid in switchgrass (Panicum virgatumL.). Plant Mol. Biol. 2014, 84, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Pollet, B.; Legrand, M. Silencing of hydroxycinnamoyl-coenzyme A shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [Green Version]
- Andrea, M.; Cinzia, C.; Sergio, L.; Ric, D.V.; Pieter, D.W.; Teris, A.V.B.; Luca, G.; Saverio, F.R.; Jules, B. Production of novel antioxidative phenolic amides through heterologous expression of the plant’s chlorogenic acid biosynthesis genes in yeast. Metab. Eng. 2010, 12, 223–232. [Google Scholar] [CrossRef]
- Sander, M.; Petersen, M. Distinct substrate specificities and unusual substrate flexibilities of two hydroxycinnamoyltransferases, rosmarinic acid synthase and hydroxycinnamoyl-CoA: Shikimate hydroxycinnamoyl-transferase, from Coleus blumei Benth. Planta 2011, 233, 1157–1171. [Google Scholar] [CrossRef]
- Kim, I.A.; Kim, B.G.; Kim, M.; Ahn, J.H. Characterization of hydroxycinnamoyltransferase from rice and its application for biological synthesis of hydroxycinnamoyl glycerols. Phytochemistry 2012, 76, 25–31. [Google Scholar] [CrossRef]
- Chen, F.; Li, Y.; Li, X.; Li, W.; Xu, J.; Cao, H.; Wang, Z.; Li, Y.; Soppe, W.J.; Liu, Y. Ectopic expression of the Arabidopsis florigen gene FLOWERING LOCUS T in seeds enhances seed dormancy via the GA and DOG1 pathways. Plant J. 2021, 107, 909–924. [Google Scholar] [CrossRef]
- Jang, S.; Hur, J.; Kim, S.J.; Han, M.J.; Kim, S.R.; An, G. Ectopic expression of OsYAB1 causes extra stamens and carpels in rice. Plant Mol. Biol. 2004, 56, 133–143. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, X.; Zheng, Y.; Liu, X. Ectopic expression of an antisense BpCCoAOMT gene from Betula platyphylla Suk. affects growth and development of tobacco due to lignin content reductio. J. Plant Biochem. Biotechnol. 2019, 29, 266–275. [Google Scholar] [CrossRef]
- Mao, P.S.; Wang, X.G.; Wang, Y.H.; Han, J.G. Effect of Storage Temperature and Duration on the Vigor of Zoysiagrass (Zoysia Japonica Steud.) Seed Harvested at Different Maturity Stages. Grassl. Sci. 2009, 55, 1–5. [Google Scholar] [CrossRef]
- Xu, Y.; Huang, B. Comparative Transcriptomic Analysis Reveals Common Molecular Factors Responsive to Heat and Drought Stress in Agrostis Stolonifera. Sci. Rep. 2018, 8, 15181. [Google Scholar] [CrossRef] [PubMed]
- Chin, S.W.; Ow, L.F. Performance of Zoysia Spp. and Axonopus Compressus Turf on Turf-Paver Complex under Simulated Traffic. Weed Turfgrass Sci. 2016, 5, 88–94. [Google Scholar] [CrossRef]
- D’Auria, J.C.; Pichersky, E.; Schaub, A.; Hansel, A.; Gershenzon, J. Characterization of a BAHD Acyltransferase Responsible for Producing the Green Leaf Volatile (Z)-3-hexen-1-yl Acetate in Arabidopsis Thaliana. Plant J. 2007, 49, 194–207. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.-H.; Yang, C.-Y.; Tzen, J.T.C. Molecular Identification and Characterization of Hydroxycinnamoyl Transferase in Tea Plants (Camellia sinensis L.). Int. J. Mol. Sci. 2018, 19, 3938. [Google Scholar] [CrossRef] [Green Version]
- Shoji, T.; Ogawa, T.; Hashimoto, T. Jasmonate-Induced Nicotine Formation in Tobacco Is Mediated by Tobacco COI1 and JAZ Genes. Plant Cell Physiol. 2008, 49, 1003–1012. [Google Scholar] [CrossRef]
- Gallego-Giraldo, L.; Shadle, G.; Shen, H.; Barros-Rios, J.; Corrales, S.F.; Wang, H.; Dixon, R.A. Combining enhanced biomass density with reduced lignin level for improved forage quality. Plant Biotechnol. J. 2016, 14, 895–904. [Google Scholar] [CrossRef]
- Anterola, A.M.; Lewis, N.G. Trends in lignin modification: A comprehensive analysis of the effects of genetic manipulations/mutations on lignification and vascular integrity. Phytochemistry 2002, 61, 221–294. [Google Scholar] [CrossRef]
- Bonawitz, N.D.; Chapple, C. The genetics of lignin biosynthesis: Connecting genotype to phenotype. Ann. Rev. Genet. 2010, 44, 337–363. [Google Scholar] [CrossRef]
- Besseau, S.; Hoffmann, L.; Geoffroy, P.; Lapierre, C.; Pollet, B.; Legrand, M. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef] [Green Version]
- Chan, M.H.; Fine, D.; Bhatia, A.; Rao, X.; Dixon, R.A. Ectopic defense gene expression is associated with growth defects in medicago truncatula lignin pathway mutants. Plant Physiol. 2019, 181, 63–84. [Google Scholar]
- Dong, D.; Zhao, Y.; Teng, K.; Tan, P.; Liu, Z.; Yang, Z.; Han, L.; Chao, Y. Expression of ZjPSY, a Phytoene Synthase Gene from Zoysia Japonica Affects Plant Height and Photosynthetic Pigment Contents. Plants 2022, 11, 395. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral Dip: A Simplified Method for Agrobacterium-Mediated Transformation of Arabidopsis Thaliana. Plant J. 2010, 16, 735–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Hu, Q.; Nelson, K.; Longo, C.; Kausch, A.P.; Chandlee, J.M.; Wipff, J.K.; Fricker, C.R. Agrobacterium Tumefaciens-Mediated Creeping Bentgrass (Agrostis Stolonifera L.) Transformation Using Phosphinothricin Selection Results in a High Frequency of Single-Copy Transgene Integration. Plant Cell Rep. 2004, 22, 645–652. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a Database of Plant Cis-Acting Regulatory Elements and a Portal to Tools for in Silico Analysis of Promoter Sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, J.; Burgess, P.; Rossi, S.; Huang, B. Interactive Effects of Melatonin and Cytokinin on Alleviating Drought-Induced Leaf Senescence in Creeping Bentgrass (Agrostis Stolonifera). Environ. Exp. Bot. 2018, 145, 1–11. [Google Scholar] [CrossRef]
- Schmittgen, T.D. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Zhao, J.; Li, G.; Yi, G.-X.; Wang, B.M.; Deng, A.X.; Nan, T.G.; Li, Z.H.; Li, Q.X. Comparison between Conventional Indirect Competitive Enzyme-Linked Immunosorbent Assay (IcELISA) and Simplified IcELISA for Small Molecules. Anal. Chim. Acta 2006, 571, 79–85. [Google Scholar] [CrossRef]
- Guo, D.; Chen, F.; Inoue, K.; Blount, J.W.; Dixon, R.A. Downregulation of Caffeic Acid 3-O-Methyltransferase and Caffeoyl CoA 3-O-Methyltransferase in Transgenic Alfalfa: Impacts on Lignin Structure and Implications for the Biosynthesis of G and S Lignin. Plant Cell 2001, 13, 73–88. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, D.; Yang, Z.; Ma, Y.; Li, S.; Wang, M.; Li, Y.; Liu, Z.; Jia, C.; Han, L.; Chao, Y. Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis stolonifera. Int. J. Mol. Sci. 2022, 23, 9500. https://doi.org/10.3390/ijms23169500
Dong D, Yang Z, Ma Y, Li S, Wang M, Li Y, Liu Z, Jia C, Han L, Chao Y. Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis stolonifera. International Journal of Molecular Sciences. 2022; 23(16):9500. https://doi.org/10.3390/ijms23169500
Chicago/Turabian StyleDong, Di, Zhuoxiong Yang, Yuan Ma, Shuwen Li, Mengdi Wang, Yinruizhi Li, Zhuocheng Liu, Chenyan Jia, Liebao Han, and Yuehui Chao. 2022. "Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis stolonifera" International Journal of Molecular Sciences 23, no. 16: 9500. https://doi.org/10.3390/ijms23169500
APA StyleDong, D., Yang, Z., Ma, Y., Li, S., Wang, M., Li, Y., Liu, Z., Jia, C., Han, L., & Chao, Y. (2022). Expression of a Hydroxycinnamoyl-CoA Shikimate/Quinate Hydroxycinnamoyl Transferase 4 Gene from Zoysia japonica (ZjHCT4) Causes Excessive Elongation and Lignin Composition Changes in Agrostis stolonifera. International Journal of Molecular Sciences, 23(16), 9500. https://doi.org/10.3390/ijms23169500