
Macrophage-Specific, Mafb-Deficient Mice Showed Delayed Skin Wound Healing

 and
and 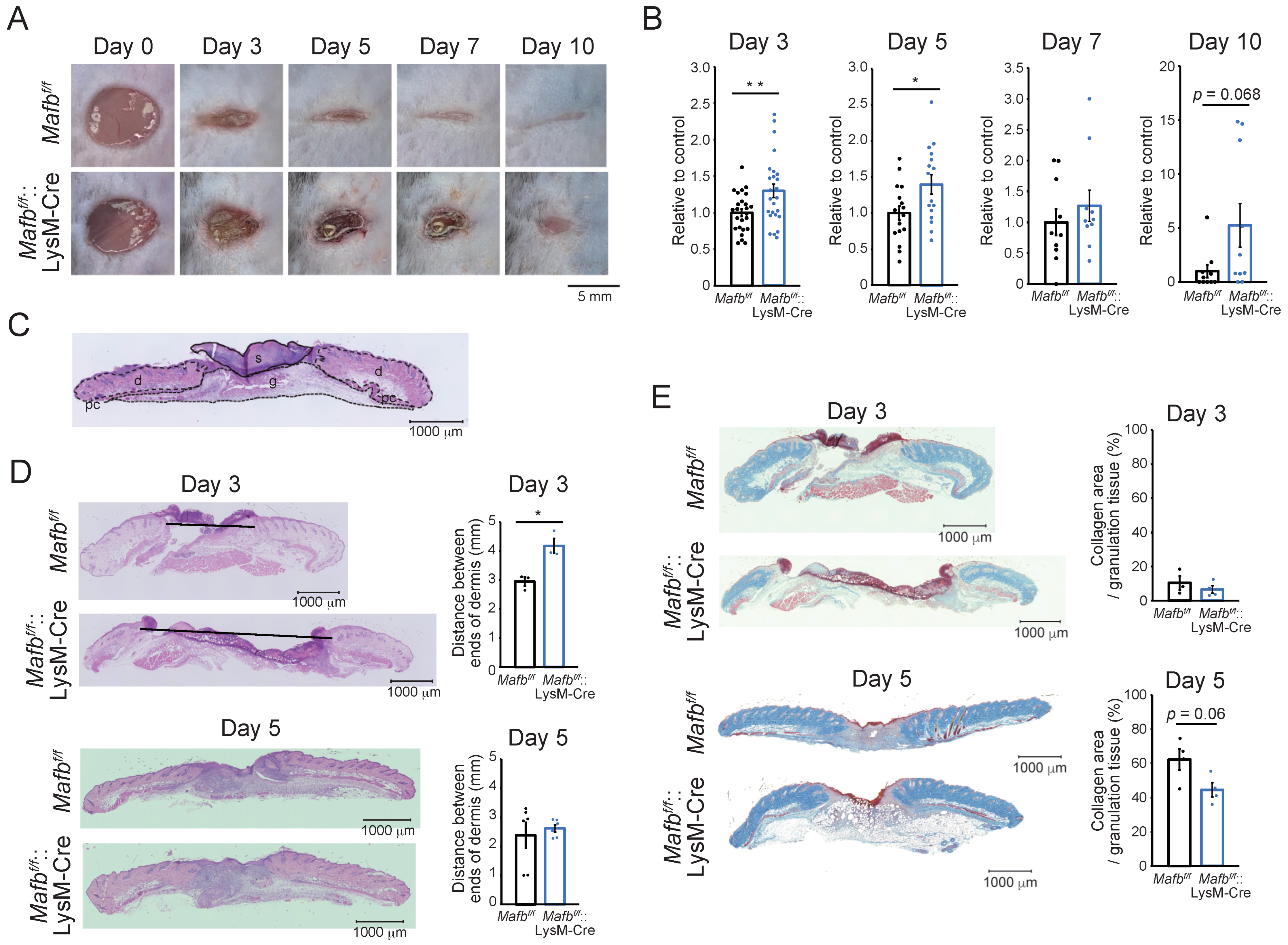{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MAFB Is Required for Proper Wound Healing
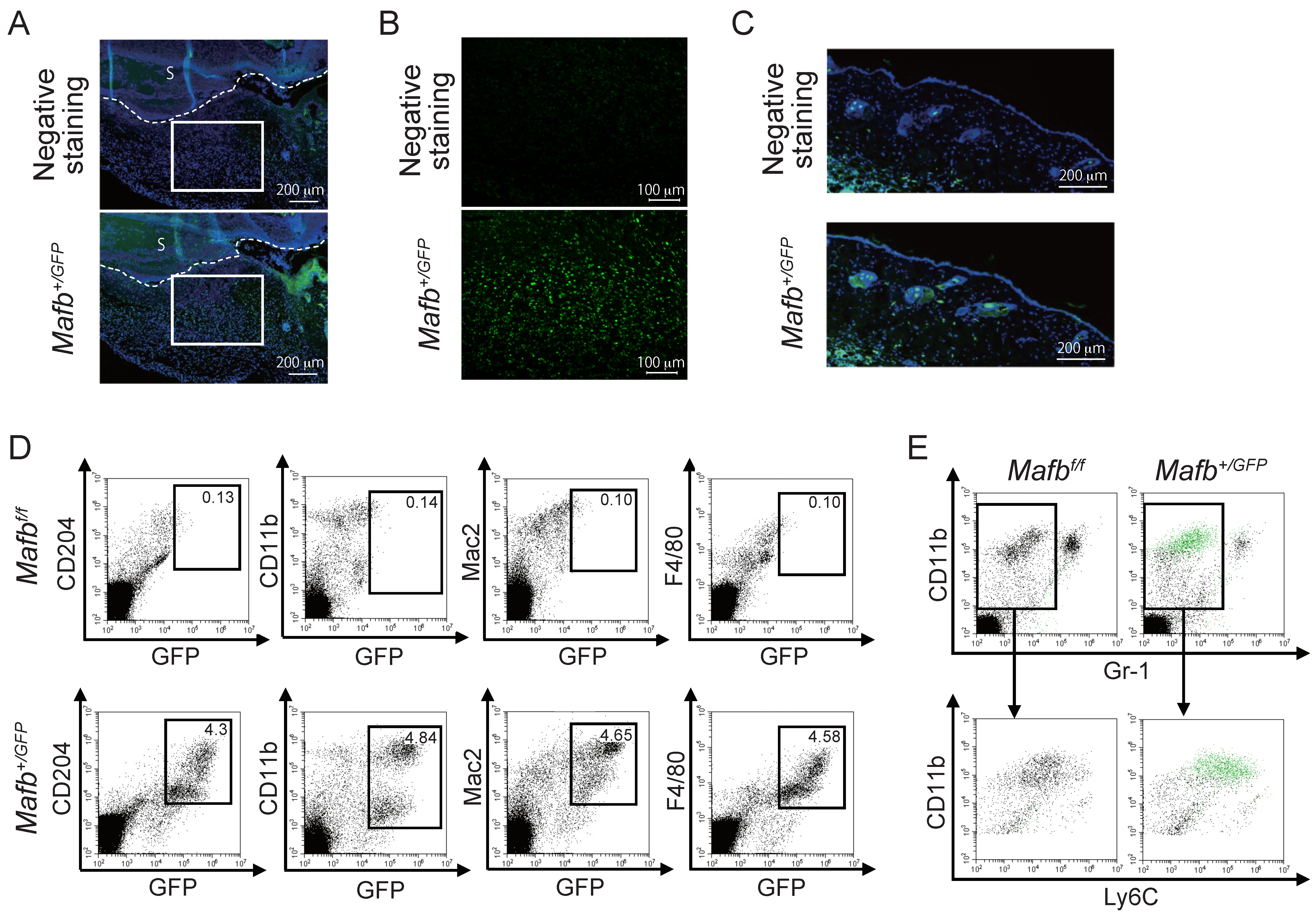
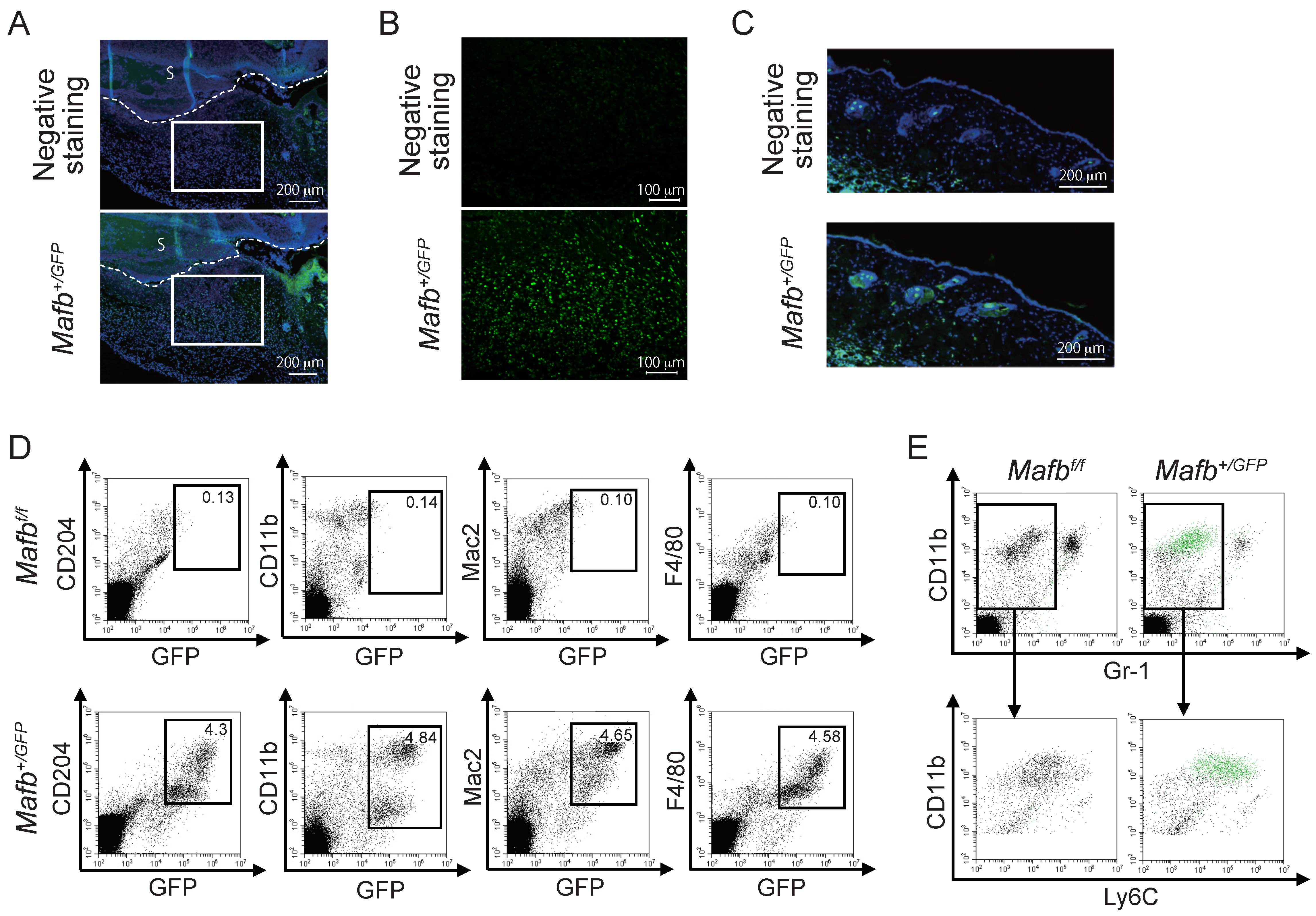
2.2. MAFB Expressed on Ly6C+ Macrophages in Granulation Tissue
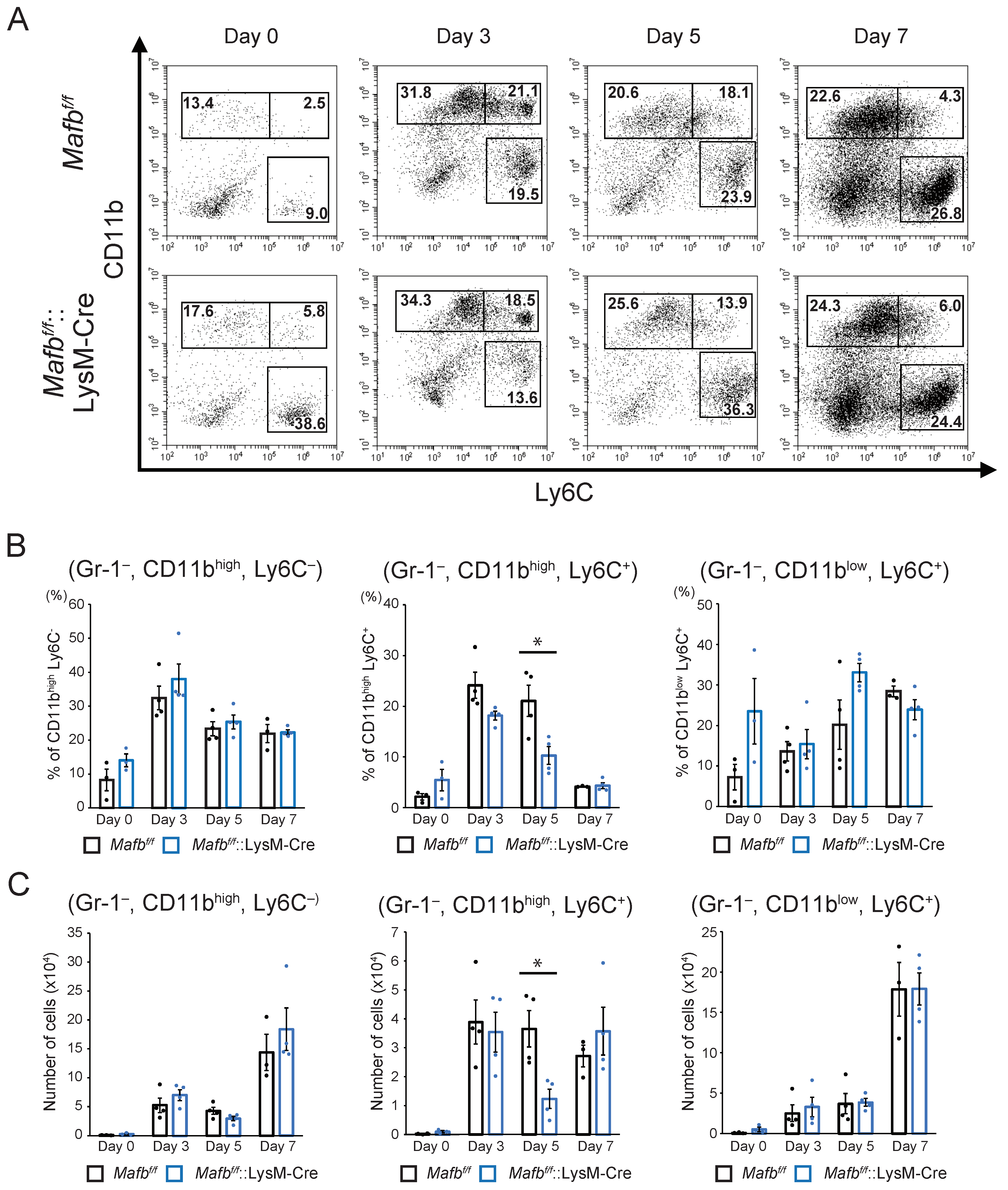
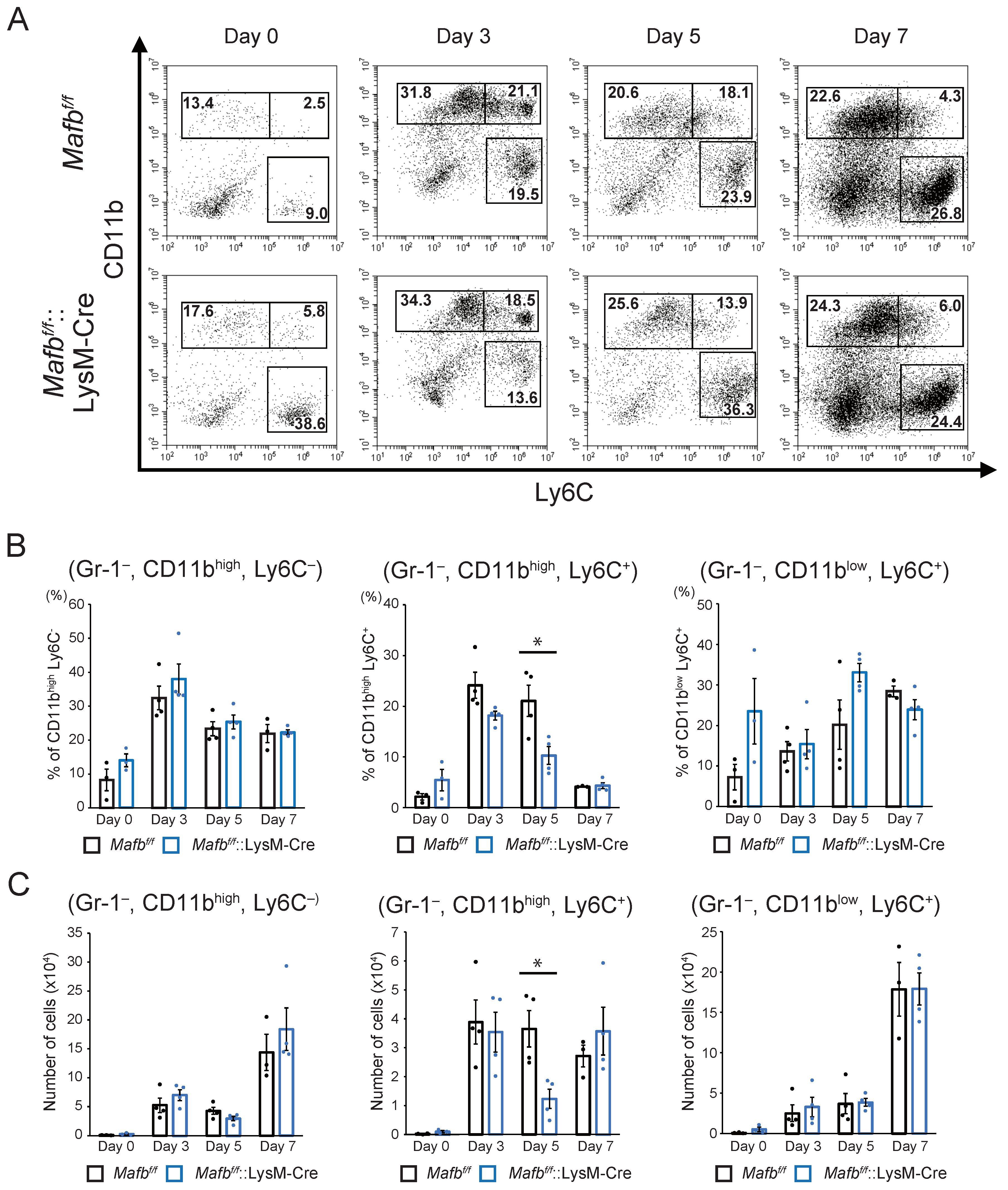
2.3. MAFB Expression in Macrophages Is Related to Their Recruitment in Wounds
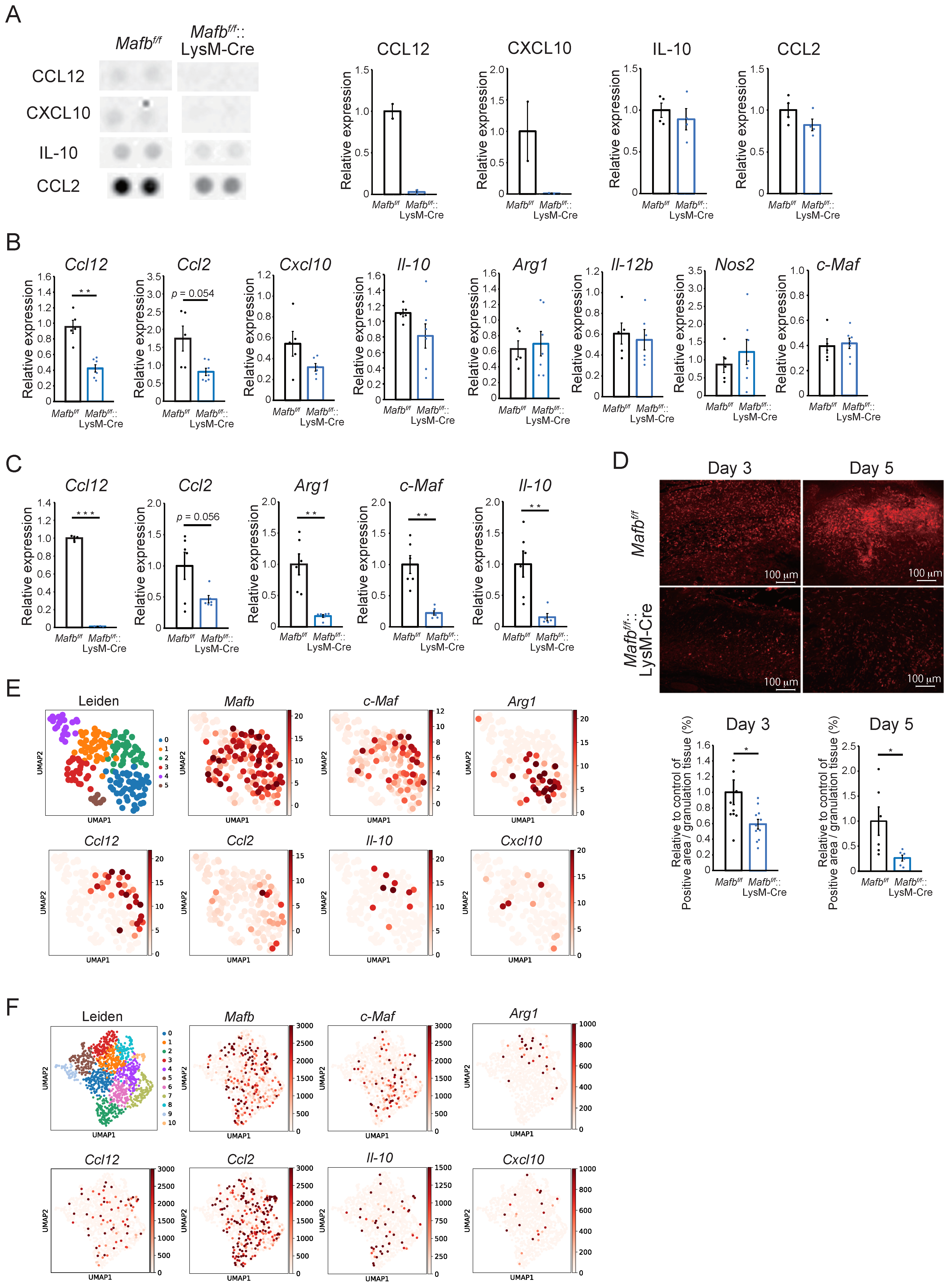
2.4. Mafb Deficiency Is Associated with CCL12, CCL2, and Arg1 Downergulation in Wounds
2.5. Mafb-Positive Macrophages Show Heterogeneous Gene Expression
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Wounding and Wound Collection
4.3. Wound Area Quantification
4.4. Histological Analysis
4.5. Flow Cytometry
4.6. Proteome Analysis
4.7. qRT-PCR
4.8. Bone Marrow Macrophage Culture
4.9. Analysis of scRNA-Seq Data
4.10. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Kataoka, K.; Fujiwara, K.T.; Noda, M.; Nishizawa, M. MafB, a New Maf Family Transcription Activator That Can Associate with Maf and Fos but Not with Jun. Mol. Cell. Biol. 1994, 14, 7581–7591. [Google Scholar] [CrossRef] [PubMed]
- Hanamura, I.; Iida, S.; Akano, Y.; Hayami, Y.; Kato, M.; Miura, K.; Harada, S.; Banno, S.; Wakita, A.; Kiyoi, H.; et al. Ectopic Expression of MAFB Gene in Human Myeloma Cells Carrying (14;20)(Q32;Q11) Chromosomal Translocations. Jpn. J. Cancer Res. 2001, 92, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjöstedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A Pathology Atlas of the Human Cancer Transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [PubMed]
- Hamada, M.; Tsunakawa, Y.; Jeon, H.; Yadav, M.K.; Takahashi, S. Role of MafB in Macrophages. Exp. Anim. 2020, 69, 1–10. [Google Scholar] [CrossRef]
- Karlsson, M.; Zhang, C.; Méar, L.; Zhong, W.; Digre, A.; Katona, B.; Sjöstedt, E.; Butler, L.; Odeberg, J.; Dusart, P.; et al. A Single–Cell Type Transcriptomics Map of Human Tissues. Sci. Adv. 2021, 7, eabh2169. [Google Scholar] [CrossRef]
- Daassi, D.; Hamada, M.; Jeon, H.; Imamura, Y.; Nhu Tran, M.T.; Takahashi, S. Differential expression patterns of MafB and c-Maf in macrophages in vivo and in vitro. Biochem. Biophys. Res. Commun. 2016, 473, 118–124. [Google Scholar] [CrossRef]
- Tran, M.T.N.; Hamada, M.; Jeon, H.; Shiraishi, R.; Asano, K.; Hattori, M.; Nakamura, M.; Imamura, Y.; Tsunakawa, Y.; Fujii, R.; et al. MafB Is a Critical Regulator of Complement Component C1q. Nat. Commun. 2017, 8, 1700. [Google Scholar] [CrossRef]
- Shichita, T.; Ito, M.; Morita, R.; Komai, K.; Noguchi, Y.; Ooboshi, H.; Koshida, R.; Takahashi, S.; Kodama, T.; Yoshimura, A. MAFB Prevents Excess Inflammation after Ischemic Stroke by Accelerating Clearance of Damage Signals through MSR1. Nat. Med. 2017, 23, 723–732. [Google Scholar] [CrossRef]
- Hamada, M.; Nakamura, M.; Tran, M.T.N.; Moriguchi, T.; Hong, C.; Ohsumi, T.; Dinh, T.T.H.; Kusakabe, M.; Hattori, M.; Katsumata, T.; et al. MafB Promotes Atherosclerosis by Inhibiting Foam-Cell Apoptosis. Nat. Commun. 2014, 5, 3147. [Google Scholar] [CrossRef]
- Aziz, A.; Soucie, E.; Sarrazin, S.; Sieweke, M.H. MafB/c-Maf Deficiency Enables Self-Renewal of Differentiated Functional Macrophages. Science 2009, 326, 867–871. [Google Scholar] [CrossRef]
- Mirza, R.; DiPietro, L.A.; Koh, T.J. Selective and Specific Macrophage Ablation Is Detrimental to Wound Healing in Mice. Am. J. Pathol. 2009, 175, 2454–2462. [Google Scholar] [CrossRef]
- Shook, B.; Xiao, E.; Kumamoto, Y.; Iwasaki, A.; Horsley, V. CD301b+ Macrophages Are Essential for Effective Skin Wound Healing. J. Investig. Dermatol. 2016, 136, 1885–1891. [Google Scholar] [CrossRef]
- Lucas, T.; Waisman, A.; Ranjan, R.; Roes, J.; Krieg, T.; Müller, W.; Roers, A.; Eming, S.A. Differential Roles of Macrophages in Diverse Phases of Skin Repair. J. Immunol. 2010, 184, 3964–3977. [Google Scholar] [CrossRef]
- Whelan, D.S.; Caplice, N.M.; Clover, A.J.P. Mesenchymal Stromal Cell Derived CCL2 Is Required for Accelerated Wound Healing. Sci. Rep. 2020, 10, 2642. [Google Scholar] [CrossRef]
- Moore, B.B.; Murray, L.; Das, A.; Wilke, C.A.; Herrygers, A.B.; Toews, G.B. The Role of CCL12 in the Recruitment of Fibrocytes and Lung Fibrosis. Am. J. Respir. Cell Mol. Biol. 2006, 35, 175–181. [Google Scholar] [CrossRef]
- Short, W.D.; Rae, M.; Lu, T.; Padon, B.; Prajapati, T.J.; Faruk, F.; Olutoye, O.O.; Yu, L.; Bollyky, P.; Keswani, S.G.; et al. Endogenous IL-10 Contributes to Wound Healing and Regulates Tissue Repair. bioRxiv 2022. [Google Scholar] [CrossRef]
- Campbell, L.; Saville, C.R.; Murray, P.J.; Cruickshank, S.M.; Hardman, M.J. Local Arginase 1 Activity Is Required for Cutaneous Wound Healing. J. Investig. Dermatol. 2013, 133, 2461–2470. [Google Scholar] [CrossRef]
- Suga, H.; Sugaya, M.; Fujita, H.; Asano, Y.; Tada, Y.; Kadono, T.; Sato, S. TLR4, Rather than TLR2, Regulates Wound Healing through TGF-β and CCL5 Expression. J. Dermatol. Sci. 2014, 73, 117–124. [Google Scholar] [CrossRef]
- Minutti, C.M.; Knipper, J.A.; Allen, J.E.; Zaiss, D.M.W. Tissue-Specific Contribution of Macrophages to Wound Healing. Semin. Cell Dev. Biol. 2017, 61, 3–11. [Google Scholar] [CrossRef]
- Shook, B.A.; Wasko, R.R.; Mano, O.; Rutenberg-Schoenberg, M.; Rudolph, M.C.; Zirak, B.; Rivera-Gonzalez, G.C.; López-Giráldez, F.; Zarini, S.; Rezza, A.; et al. Dermal Adipocyte Lipolysis and Myofibroblast Conversion Are Required for Efficient Skin Repair. Cell Stem Cell 2020, 26, 880–895.e6. [Google Scholar] [CrossRef]
- Willenborg, S.; Sanin, D.E.; Jais, A.; Ding, X.; Ulas, T.; Nüchel, J.; Popović, M.; MacVicar, T.; Langer, T.; Schultze, J.L.; et al. Mitochondrial Metabolism Coordinates Stage-Specific Repair Processes in Macrophages during Wound Healing. Cell Metab. 2021, 33, 2398–2414.e9. [Google Scholar] [CrossRef]
- Sarafi, M.N.; Garcia-Zepeda, E.A.; MacLean, J.A.; Charo, I.F.; Luster, A.D. Murine Monocyte Chemoattractant Protein (MCP)-5: A Novel CC Chemokine That Is a Structural and Functional Homologue of Human MCP-1. J. Exp. Med. 1997, 185, 99–109. [Google Scholar] [CrossRef]
- Yang, J.; Agarwal, M.; Ling, S.; Teitz-Tennenbaum, S.; Zemans, R.L.; Osterholzer, J.J.; Sisson, T.H.; Kim, K.K. Diverse Injury Pathways Induce Alveolar Epithelial Cell CCL2/12, Which Promotes Lung Fibrosis. Am. J. Respir. Cell Mol. Biol. 2020, 62, 622–632. [Google Scholar] [CrossRef]
- Moore, B.B.; Kolodsick, J.E.; Thannickal, V.J.; Cooke, K.; Moore, T.A.; Hogaboam, C.; Wilke, C.A.; Toews, G.B. CCR2-Mediated Recruitment of Fibrocytes to the Alveolar Space after Fibrotic Injury. Am. J. Pathol. 2005, 166, 675–684. [Google Scholar] [CrossRef]
- Boniakowski, A.E.; Kimball, A.S.; Joshi, A.; Schaller, M.; Davis, F.M.; denDekker, A.; Obi, A.T.; Moore, B.B.; Kunkel, S.L.; Gallagher, K.A. Murine Macrophage Chemokine Receptor CCR2 Plays a Crucial Role in Macrophage Recruitment and Regulated Inflammation in Wound Healing. Eur. J. Immunol. 2018, 48, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Metabolism, A.; Healing, W. Basic Science Review: Arginine Metabolism and Wound Healing. Wound Health S. Afr. 2008, 1, 48–50. [Google Scholar]
- Crompton, R.A.; Williams, H.; Campbell, L.; Hui Kheng, L.; Saville, C.; Ansell, D.M.; Reid, A.; Wong, J.; Vardy, L.A.; Hardman, M.J.; et al. An Epidermal-Specific Role for Arginase1 during Cutaneous Wound Repair. J. Investig. Dermatol. 2022, 142, 1206–1216.e8. [Google Scholar] [CrossRef] [PubMed]
- Imbratta, C.; Hussein, H.; Andris, F.; Verdeil, G. C-MAF, a Swiss Army Knife for Tolerance in Lymphocytes. Front. Immunol. 2020, 11, 206. [Google Scholar] [CrossRef]
- Lopez-Pajares, V.; Qu, K.; Zhang, J.; Webster, D.E.; Barajas, B.C.; Siprashvili, Z.; Zarnegar, B.J.; Boxer, L.D.; Rios, E.J.; Tao, S.; et al. A LncRNA-MAF:MAFB Transcription Factor Network Regulates Epidermal Differentiation. Dev. Cell 2015, 32, 693–706. [Google Scholar] [CrossRef]
- King, A.; Balaji, S.; Le, L.D.; Crombleholme, T.M.; Keswani, S.G. Regenerative Wound Healing: The Role of Interleukin-10. Adv. Wound Care 2014, 3, 315–323. [Google Scholar] [CrossRef]
- Cao, S.; Liu, J.; Song, L.; Ma, X. The Protooncogene C-Maf Is an Essential Transcription Factor for IL-10 Gene Expression in Macrophages. J. Immunol. 2005, 174, 3484–3492. [Google Scholar] [CrossRef]
- Hayuningtyas, R.A.; Han, M.; Choi, S.; Kwak, M.S.; Park, I.H.; Lee, J.-H.; Choi, J.E.; Kim, D.K.; Son, M.; Shin, J.-S. The Collagen Structure of C1q Induces Wound Healing by Engaging Discoidin Domain Receptor 2. Mol. Med. 2021, 27, 125. [Google Scholar] [CrossRef]
- Bossi, F.; Tripodo, C.; Rizzi, L.; Bulla, R.; Agostinis, C.; Guarnotta, C.; Munaut, C.; Baldassarre, G.; Papa, G.; Zorzet, S.; et al. C1q as a Unique Player in Angiogenesis with Therapeutic Implication in Wound Healing. Proc. Natl. Acad. Sci. USA 2014, 111, 4209–4214. [Google Scholar] [CrossRef]
- Usui, T.; Morito, N.; Shawki, H.H.; Sato, Y.; Tsukaguchi, H.; Hamada, M.; Jeon, H.; Yadav, M.K.; Kuno, A.; Tsunakawa, Y.; et al. Transcription Factor MafB in Podocytes Protects against the Development of Focal Segmental Glomerulosclerosis. Kidney Int. 2020, 98, 391–403. [Google Scholar] [CrossRef]
- Miyai, M.; Hamada, M.; Moriguchi, T.; Hiruma, J.; Kamitani-Kawamoto, A.; Watanabe, H.; Hara-Chikuma, M.; Takahashi, K.; Takahashi, S.; Kataoka, K. Transcription Factor MafB Coordinates Epidermal Keratinocyte Differentiation. J. Investig. Dermatol. 2016, 136, 1848–1857. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Iijima, S.; Hasegawa, Y.; Suzuki, Y.; Daitoku, Y.; Mizuno, S.; Ishige, T.; Kudo, T.; Takahashi, S.; Kunita, S.; et al. Embryonic Stem Cells Derived from C57BL/6J and C57BL/6N Mice. Comp. Med. 2008, 58, 347–352. [Google Scholar]
- Clausen, B.E.; Burkhardt, C.; Reith, W.; Renkawitz, R.; Förster, I. Conditional Gene Targeting in Macrophages and Granulocytes Using LysMcre Mice. Transgenic Res. 1999, 8, 265–277. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Inoue, Y.; Liao, C.-W.; Tsunakawa, Y.; Tsai, I.-L.; Takahashi, S.; Hamada, M. Macrophage-Specific, Mafb-Deficient Mice Showed Delayed Skin Wound Healing. Int. J. Mol. Sci. 2022, 23, 9346. https://doi.org/10.3390/ijms23169346
Inoue Y, Liao C-W, Tsunakawa Y, Tsai I-L, Takahashi S, Hamada M. Macrophage-Specific, Mafb-Deficient Mice Showed Delayed Skin Wound Healing. International Journal of Molecular Sciences. 2022; 23(16):9346. https://doi.org/10.3390/ijms23169346
Chicago/Turabian StyleInoue, Yuri, Ching-Wei Liao, Yuki Tsunakawa, I-Lin Tsai, Satoru Takahashi, and Michito Hamada. 2022. "Macrophage-Specific, Mafb-Deficient Mice Showed Delayed Skin Wound Healing" International Journal of Molecular Sciences 23, no. 16: 9346. https://doi.org/10.3390/ijms23169346
APA StyleInoue, Y., Liao, C.-W., Tsunakawa, Y., Tsai, I.-L., Takahashi, S., & Hamada, M. (2022). Macrophage-Specific, Mafb-Deficient Mice Showed Delayed Skin Wound Healing. International Journal of Molecular Sciences, 23(16), 9346. https://doi.org/10.3390/ijms23169346