
Regulation of Heat Stress in Physcomitrium (Physcomitrella) patens Provides Novel Insight into the Functions of Plant RNase H1s

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Phylogenetic Analysis of RNases H1s
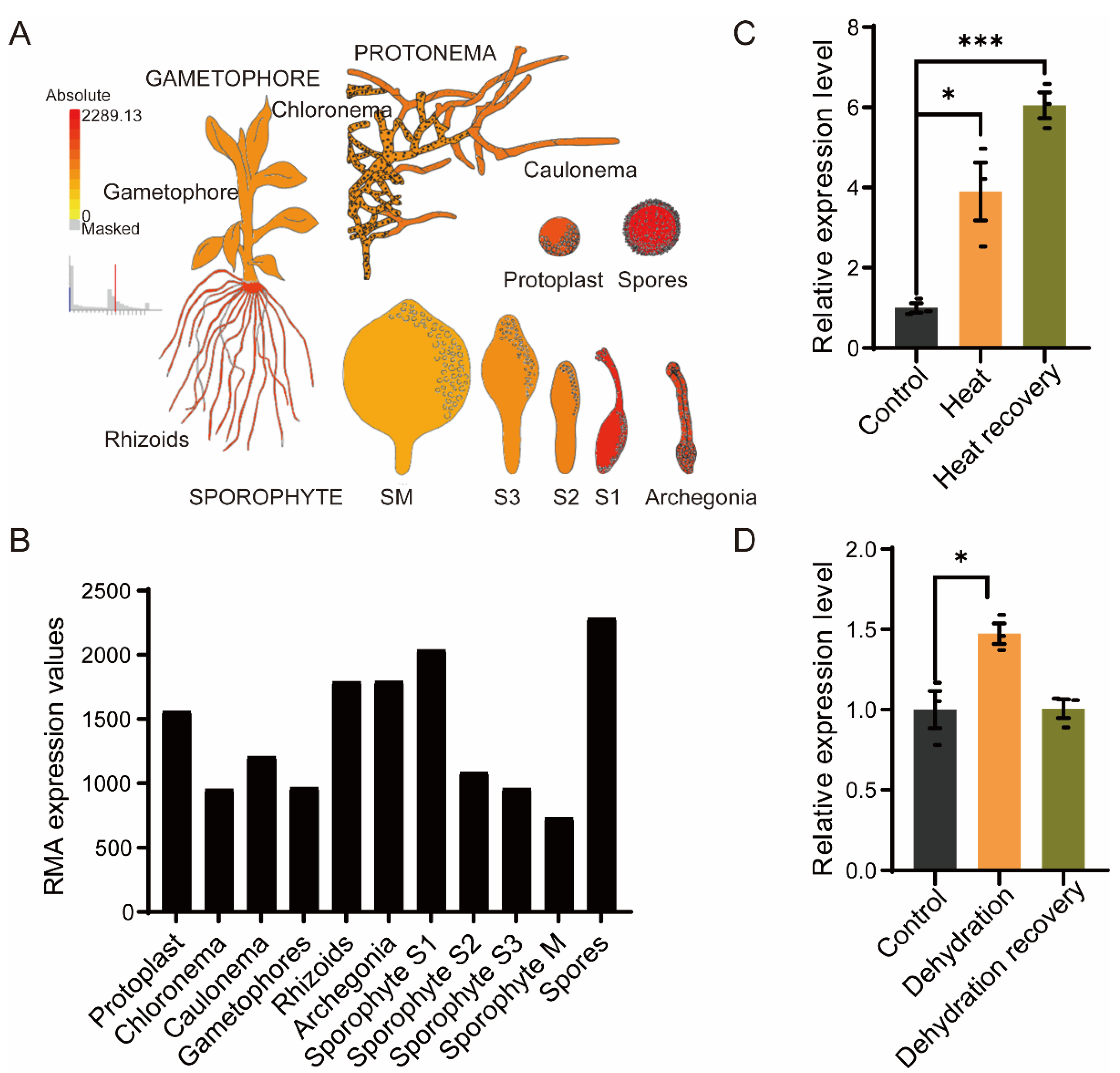
2.2. PpRNH1A Is Involved in Growth Development and Stress Tolerance
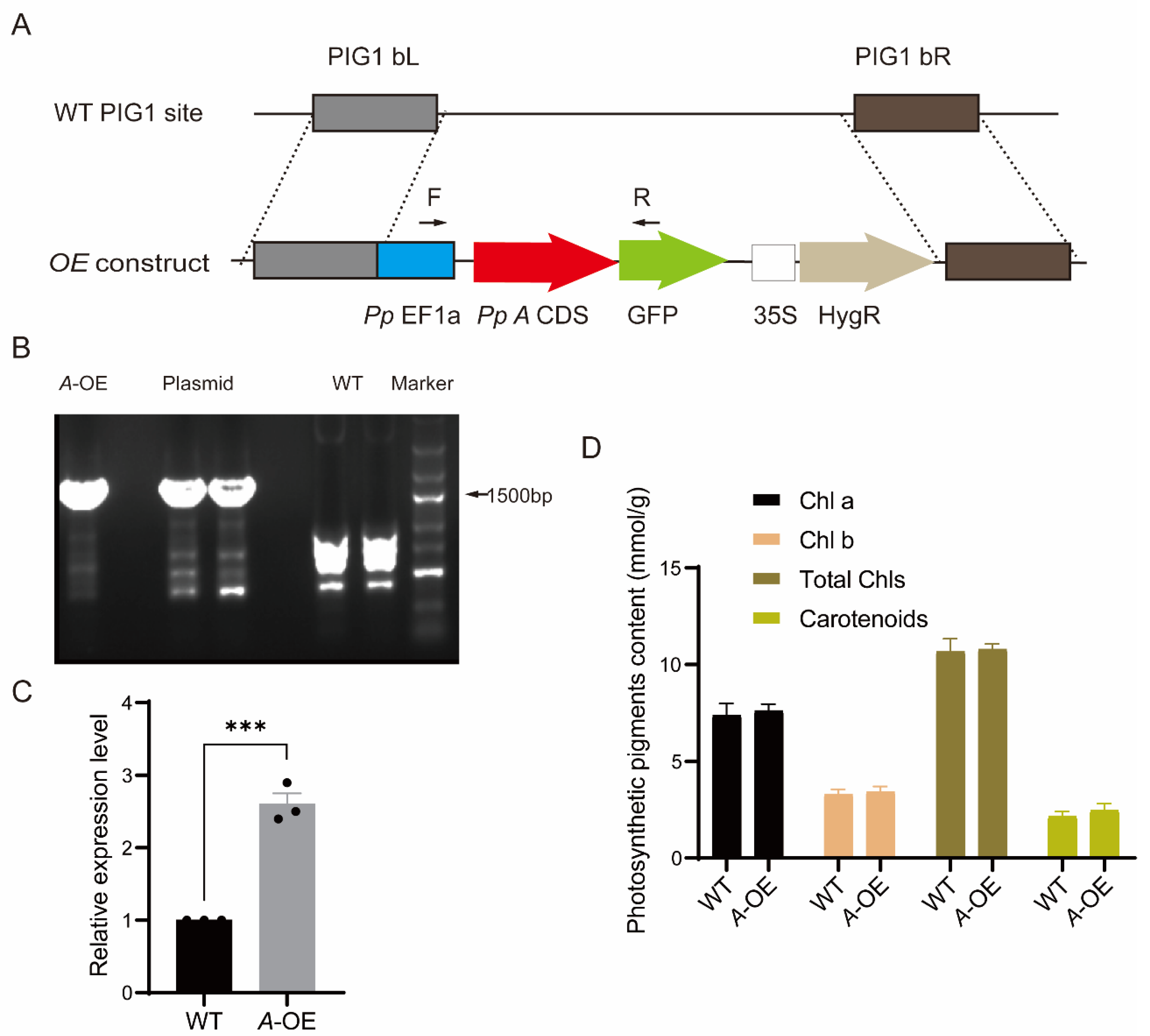
2.3. Creation of Stable Transgenic P. patens Line Overexpressing PpRNH1A
2.4. Overexpression of PpRNH1A Affects Lipid Droplets Metabolism and Mobilization
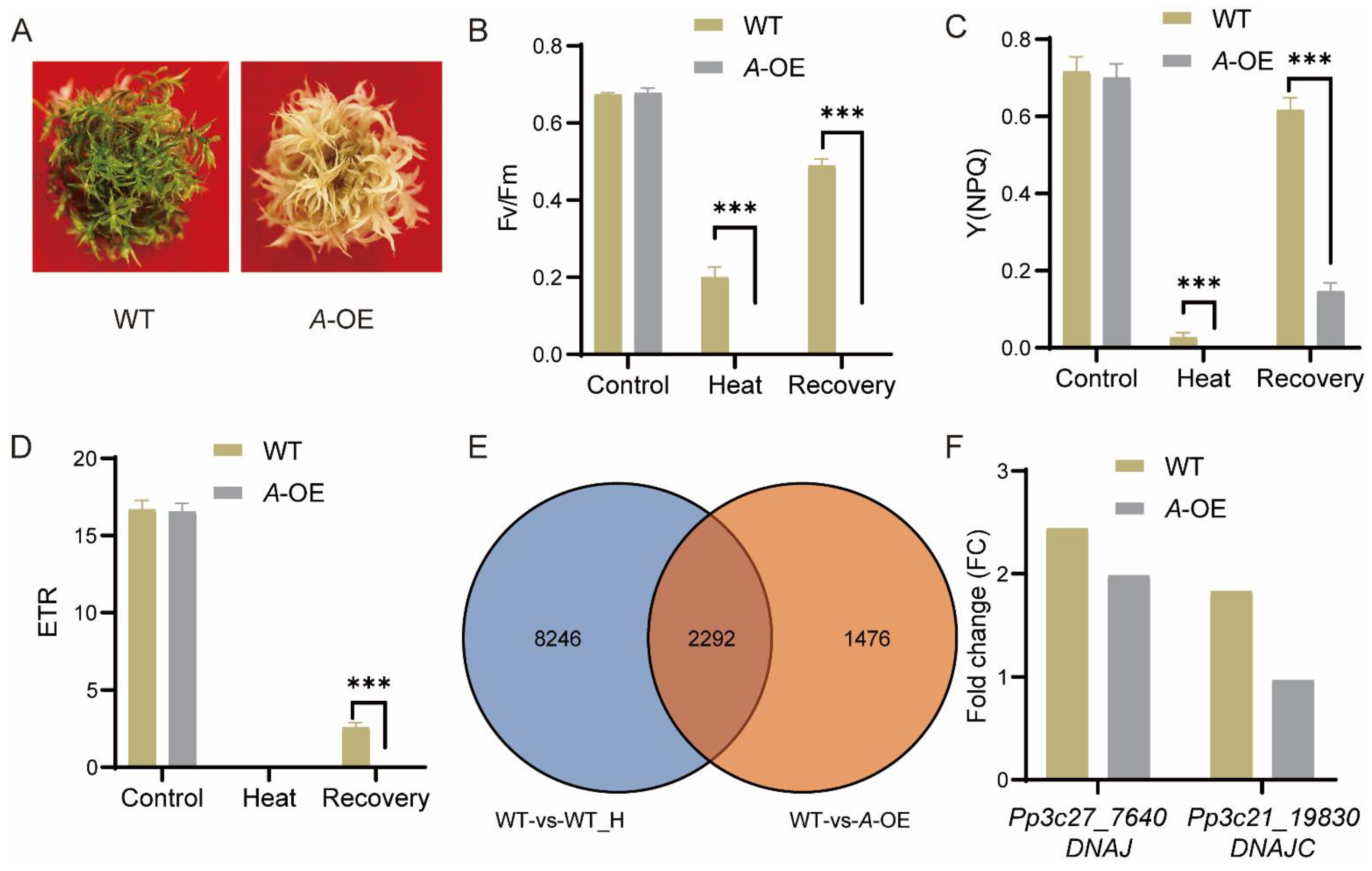
2.5. PpRNH1A-Overexpression Line Is More Sensitive to Heat Stress
2.6. PpRNH1A Regulates the Expression of Heat-Responsive Genes including DNAJ and DNAJC
3. Discussion
4. Materials and Methods
4.1. Phylogenetic Analysis
4.2. Plant Materials and Growth Conditions
4.3. Protoplast Transformation
4.4. PCR and Real-Time qRT-PCR Characterization of Overexpression Plants
4.5. Analysis of Gene Expression Patterns and Analysis of Expression by Stress Treatment
4.6. Observation of Cell Ultrastructure
4.7. Observation of Cell Surface Structure
4.8. Nile Red Staining
4.9. Heat Stress Assay
4.10. Measurement of Chlorophyll Fluorescence (Fv/Fm), Non-Photochemical Energy Dissipation (NPQ), Electron Transport Rate (ETR), and Pigments Content
4.11. Bioinformatics and Data Analysis
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sarwar, M.; Saleem, M.F.; Ullah, N.; Rizwan, M.; Ali, S.; Shahid, M.R.; Alamri, S.A.; Alyemeni, M.N.; Ahmad, P. Exogenously applied growth regulators protect the cotton crop from heat-induced injury by modulating plant defense mechanism. Sci. Rep. 2018, 8, 17086. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Sirhindi, G.; Bhardwaj, R.; Alyemeni, M.N.; Siddique, K.H.M.; Ahmad, P. 28-homobrassinolide regulates antioxidant enzyme activities and gene expression in response to salt- and temperature-induced oxidative stress in Brassica juncea. Sci. Rep. 2018, 8, 8735. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Hayes, S.; Schachtschabel, J.; Mishkind, M.; Munnik, T.; Arisz, S.A. Hot topic: Thermosensing in plants. Plant Cell Environ. 2021, 44, 2018–2033. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Vierling, E. The roles of heat shock proteins in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Rajan, V.B.V.; D’Silva, P. Arabidopsis thaliana J-class heat shock proteins: Cellular stress sensors. Funct. Integr. Genom. 2009, 9, 433. [Google Scholar] [CrossRef]
- Kong, F.; Deng, Y.; Wang, G.; Wang, J.; Liang, X.; Meng, Q. LeCDJ1, a chloroplast DnaJ protein, facilitates heat tolerance in transgenic tomatoes. J. Integr. Plant Biol. 2014, 56, 63–74. [Google Scholar] [CrossRef]
- Li, K.P.; Wong, C.H.; Cheng, C.C.; Cheng, S.S.; Li, M.W.; Mansveld, S.; Bergsma, A.; Huang, T.; van Eijk, M.J.T.; Lam, H.M. GmDNJ1, a type-I heat shock protein 40 (HSP40), is responsible for both Growth and heat tolerance in soybean. Plant Direct 2021, 5, e00298. [Google Scholar] [CrossRef]
- Shao, Q.; Liu, X.; Su, T.; Ma, C.; Wang, P. New insights into the role of seed oil body proteins in metabolism and plant development. Front. Plant Sci. 2019, 10, 1568. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, F.K.; Doner, N.M.; Krawczyk, H.E.; Scholz, P.; Schmitt, K.; Valerius, O.; Braus, G.H.; Mullen, R.T.; Ischebeck, T. Identification of low-abundance lipid droplet proteins in seeds and seedlings. Plant Physiol. 2020, 182, 1326–1345. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.-L.; Huang, M.-D.; Chen, T.-L.L.; Huang, A.H.C. Oleosin of subcellular lipid droplets evolved in green algae. Plant Physiol. 2013, 161, 1862–1874. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Santos, R.; Izquierdo, Y.; López, A.; Muñiz, L.; Martínez, M.; Cascón, T.; Hamberg, M.; Castresana, C. Protein profiles of lipid droplets during the hypersensitive defense response of Arabidopsis against Pseudomonas infection. Plant Cell Physiol. 2020, 61, 1144–1157. [Google Scholar] [CrossRef] [PubMed]
- Feeney, M.; Frigerio, L.; Cui, Y.; Menassa, R. Following vegetative to embryonic cellular changes in leaves of Arabidopsis overexpressing LEAFY COTYLEDON2. Plant Physiol. 2013, 162, 1881–1896. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Xu, Y.; Wang, J.; Singer, S.D.; Chen, G. The role of triacylglycerol in plant stress response. Plants 2020, 9, 472. [Google Scholar] [CrossRef] [PubMed]
- de Vries, J.; Ischebeck, T. Ties between stress and lipid droplets pre-date seeds. Trends Plant Sci. 2020, 25, 1203–1214. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef]
- Légeret, B.; Schulz-Raffelt, M.; Nguyen, H.M.; Auroy, P.; Beisson, F.; Peltier, G.; Blanc, G.; Li-Beisson, Y. Lipidomic and transcriptomic analyses of Chlamydomonas reinhardtii under heat stress unveil a direct route for the conversion of membrane lipids into storage lipids. Plant Cell Environ. 2016, 39, 834–847. [Google Scholar] [CrossRef]
- Aizouq, M.; Peisker, H.; Gutbrod, K.; Melzer, M.; Hölzl, G.; Dörmann, P. Triacylglycerol and phytyl ester synthesis in Synechocystis sp. PCC6803. Proc. Natl. Acad. Sci. USA 2020, 117, 6216–6222. [Google Scholar] [CrossRef]
- Gogna, M.; Bhatla, S. Biochemical mechanisms regulating salt tolerance in sunflower. Plant Signal. Behav. 2019, 14, 1670597. [Google Scholar] [CrossRef] [PubMed]
- Gogna, M.; Bhatla, S. Salt-tolerant and -sensitive seedlings exhibit noteworthy differences in lipolytic events in response to salt stress. Plant Signal. Behav. 2020, 15, 1737451. [Google Scholar] [CrossRef] [PubMed]
- Guan, Y.; Liu, L.; Wang, Q.; Zhao, J.; Li, P.; Hu, J.; Yang, Z.; Running, M.P.; Sun, H.; Huang, J. Gene refashioning through innovative shifting of reading frames in mosses. Nat. Commun. 2018, 9, 1555. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhai, J.; Qi, Z.; Liu, W.; Cui, J.; Zhang, X.; Bai, S.; Li, L.; Shui, G.; Cui, S. The specific glycerolipid composition is responsible for maintaining the membrane stability of Physcomitrella patens under dehydration stress. J. Plant Physiol. 2021, 268, 153590. [Google Scholar] [CrossRef] [PubMed]
- Stein, H.; Hausen, P. Enzyme from calf thymus degrading the RNA moiety of DNA-RNA Hybrids: Effect on DNA-dependent RNA polymerase. Science 1969, 166, 393–395. [Google Scholar] [CrossRef]
- Amon, J.D.; Koshland, D. RNase H enables efficient repair of R-loop induced DNA damage. eLife 2016, 5, e20533. [Google Scholar] [CrossRef]
- Ohtani, N.; Haruki, M.; Morikawa, M.; Crouch, R.J.; Itaya, M.; Kanaya, S. Identification of the genes encoding Mn2+-dependent RNase HII and Mg2+-dependent RNase HIII from Bacillus subtilis: Classification of RNases H into three families. Biochemistry 1999, 38, 605–618. [Google Scholar] [CrossRef]
- Chon, H.; Matsumura, H.; Koga, Y.; Takano, K.; Kanaya, S. Crystal Structure and Structure-based Mutational Analyses of RNase HIII from Bacillus stearothermophilus: A New Type 2 RNase H with TBP-like Substrate-binding Domain at the N Terminus. J. Mol. Biol. 2006, 356, 165–178. [Google Scholar] [CrossRef]
- Jongruja, N.; You, D.J.; Angkawidjaja, C.; Kanaya, E.; Koga, Y.; Kanaya, S. Structure and characterization of RNase H3 from Aquifex aeolicus. FEBS J. 2012, 279, 2737–2753. [Google Scholar] [CrossRef]
- Kalhorzadeh, P.; Hu, Z.; Cools, T.; Amiard, S.; Willing, E.-M.; De Winne, N.; Gevaert, K.; De Jaeger, G.; Schneeberger, K.; White, C.I. Arabidopsis thaliana RNase H2 deficiency counteracts the needs for the WEE1 checkpoint kinase but triggers genome instability. Plant Cell 2014, 26, 3680–3692. [Google Scholar] [CrossRef]
- Eekhout, T.; Kalhorzadeh, P.; De Veylder, L. Lack of RNase H2 activity rescues HU-sensitivity of WEE1 deficient plants. Plant Signal. Behav. 2015, 10, e1001226. [Google Scholar] [PubMed]
- Hyjek, M.; Figiel, M.; Nowotny, M. RNases H: Structure and mechanism. DNA Repair 2019, 84, 102672. [Google Scholar] [CrossRef] [PubMed]
- Holmes, J.B.; Akman, G.; Wood, S.R.; Sakhuja, K.; Cerritelli, S.M.; Moss, C.; Bowmaker, M.R.; Jacobs, H.T.; Crouch, R.J.; Holt, I.J. Primer retention owing to the absence of RNase H1 is catastrophic for mitochondrial DNA replication. Proc. Natl. Acad. Sci. USA 2015, 112, 9334–9339. [Google Scholar] [CrossRef]
- Akman, G.; Desai, R.; Bailey, L.J.; Yasukawa, T.; Dalla Rosa, I.; Durigon, R.; Holmes, J.B.; Moss, C.F.; Mennuni, M.; Houlden, H.; et al. Pathological ribonuclease H1 causes R-loop depletion and aberrant DNA segregation in mitochondria. Proc. Natl. Acad. Sci. USA 2016, 113, E4276–E4285. [Google Scholar] [CrossRef] [PubMed]
- Cerritelli, S.M.; Frolova, E.G.; Feng, C.; Grinberg, A.; Love, P.E.; Crouch, R.J. Failure to Produce Mitochondrial DNA Results in Embryonic Lethality in Rnaseh1 Null Mice. Mol. Cell 2003, 11, 807–815. [Google Scholar] [CrossRef]
- Kramara, J.; Osia, B.; Malkova, A. Break-Induced Replication: The Where, The Why, and The How. Trends Genet. 2018, 34, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.L.; Lei, Y.N.; Liu, X.Q.; Xue, W.; Zhang, Y.; Nan, F.; Gao, X.; Zhang, J.; Wei, J.; et al. Linking circular intronic RNA degradation and function in transcription by RNase H1. Sci. China Life Sci. 2021, 64, 1795–1809. [Google Scholar] [CrossRef]
- Yang, Z.; Hou, Q.; Cheng, L.; Xu, W.; Hong, Y.; Li, S.; Sun, Q. RNase H1 cooperates with DNA gyrases to restrict R-loops and maintain genome integrity in Arabidopsis chloroplasts. Plant Cell 2017, 29, 2478–2497. [Google Scholar] [CrossRef]
- Kuciński, J.; Chamera, S.; Kmera, A.; Rowley, M.J.; Fujii, S.; Khurana, P.; Nowotny, M.; Wierzbicki, A.T. Evolutionary history and activity of RNase H1-like proteins in Arabidopsis thaliana. Plant Cell Physiol. 2020, 61, 1107–1119. [Google Scholar] [CrossRef]
- Wang, W.; Li, K.; Yang, Z.; Hou, Q.; Zhao, W.W.; Sun, Q. RNase H1C collaborates with ssDNA binding proteins WHY1/3 and recombinase RecA1 to fulfill the DNA damage repair in Arabidopsis chloroplasts. Nucleic Acids Res. 2021, 49, 6771–6787. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, W.; Yao, Y.; Sun, Q. Mitochondrial RNase H1 activity regulates R-loop homeostasis to maintain genome integrity and enable early embryogenesis in Arabidopsis. PLoS Biol. 2021, 19, e3001357. [Google Scholar] [CrossRef] [PubMed]
- Cove, D. The moss Physcomitrella patens. Annu. Rev. Genet. 2005, 39, 339–358. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Rensing, S.A.; Goffinet, B.; Meyberg, R.; Wu, S.-Z.; Bezanilla, M. The moss Physcomitrium (Physcomitrella) patens: A model organism for non-seed plants. Plant Cell 2020, 32, 1361–1376. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Dong, X.; Yang, Z.; Hou, X.; Liu, L. Regulation of the Development in Physcomitrium (Physcomitrella) patens implicates the functional differentiation of plant RNase H1s. Plant Sci. 2021, 313, 111070. [Google Scholar] [CrossRef]
- Dong, X.M.; Pu, X.J.; Zhou, S.Z.; Li, P.; Luo, T.; Chen, Z.X.; Chen, S.L.; Liu, L. Orphan gene PpARDT positively involved in drought tolerance potentially by enhancing ABA response in Physcomitrium (Physcomitrella) patens. Plant Sci. 2022, 319, 11122. [Google Scholar] [CrossRef]
- Ul Haq, S.; Khan, A.; Ali, M.; Khattak, A.M.; Gai, W.X.; Zhang, H.X.; Wei, A.M.; Gong, Z.H. Heat shock proteins: Dynamic biomolecules to counter plant biotic and abiotic stresses. Int. J. Mol. Sci. 2019, 20, 5321. [Google Scholar] [CrossRef]
- Lee, H.G.; Park, M.-E.; Park, B.Y.; Kim, H.U.; Seo, P.J. The Arabidopsis MYB96 transcription factor mediates ABA-dependent triacylglycerol accumulation in vegetative tissues under drought stress conditions. Plants 2019, 8, 296. [Google Scholar] [CrossRef]
- Angkawijaya, A.E.; Nguyen, V.C.; Nakamura, Y. LYSOPHOSPHATIDIC ACID ACYLTRANSFERASES 4 and 5 are involved in glycerolipid metabolism and nitrogen starvation response in Arabidopsis. New Phytol. 2019, 224, 336–351. [Google Scholar] [CrossRef]
- Shimada, T.L.; Shimada, T.; Takahashi, H.; Fukao, Y.; Hara-Nishimura, I. A novel role for oleosins in freezing tolerance of oilseeds in Arabidopsis thaliana. Plant J. 2008, 55, 798–809. [Google Scholar] [CrossRef]
- Miquel, M.; Ghassen, T.; d’Andréa, S.; Kelemen, Z.; Baud, S.; Berger, A.; Deruyffelaere, C.; Trubuil, A.; Lepiniec, L.; Dubreucq, B. Specialization of oleosins in oil body dynamics during seed development in Arabidopsis Seeds. Plant Physiol. 2014, 164, 1866–1878. [Google Scholar] [CrossRef] [PubMed]
- Lundquist, P.K.; Shivaiah, K.-K.; Espinoza-Corral, R. Lipid droplets throughout the evolutionary tree. Prog. Lipid Res. 2020, 78, 101029. [Google Scholar] [CrossRef] [PubMed]
- Duckett, J.G.; Ligrone, R. The formation of catenate foliar gemmae and the origin of oil bodies in the liverwort Odontoschisma denudatum (Mart.) Dum. (Jungermanniales): A light and electron microscope study. Ann. Bot. 1995, 76, 405–419. [Google Scholar] [CrossRef]
- Scholz, P.; Chapman, K.D.; Mullen, R.T.; Ischebeck, T. Finding new friends and revisiting old ones—How plant lipid droplets connect with other subcellular structures. New Phytol. 2022. [Google Scholar] [CrossRef]
- Romani, F.; Flores, J.; Tolopka, J.; Suarez, G.; He, X.; Moreno, J. Liverwort oil bodies: Diversity, biochemistry, and molecular cell biology of the earliest secretory structure of land plants. J. Exp. Bot. 2022, 73, 4427–4439. [Google Scholar] [CrossRef]
- Schuster, R.M. The oil-bodies of the Hepaticae. I. Introduction. J. Hattori Bot. Lab. 1992, 72, 151–162. [Google Scholar]
- Schuster, R.M.; Konstantinova, N. Studies on Treubiales, I. On Apotreubia Hatt. et al. and A. hortonae Schust. & Konstantinova, sp. n. J. Hattori Bot. Lab. 1995, 78, 41–61. [Google Scholar]
- Wang, G.; Cai, G.; Xu, N.; Zhang, L.; Sun, X.; Guan, J.; Meng, Q. Novel DnaJ protein facilitates thermotolerance of transgenic tomatoes. Int. J. Mol. Sci. 2019, 20, 367. [Google Scholar] [CrossRef]
- Lee, K.-W.; Rahman, M.A.; Kim, K.-Y.; Choi, G.J.; Cha, J.-Y.; Cheong, M.S.; Shohael, A.M.; Jones, C.; Lee, S.-H. Overexpression of the alfalfa DnaJ-like protein (MsDJLP) gene enhances tolerance to chilling and heat stresses in transgenic tobacco plants. Turk. J. Biol. 2018, 42, 12–22. [Google Scholar] [CrossRef]
- Hori, K.; Maruyama, F.; Fujisawa, T.; Togashi, T.; Yamamoto, N.; Seo, M.; Sato, S.; Yamada, T.; Mori, H.; Tajima, N. Klebsormidium flaccidum genome reveals primary factors for plant terrestrial adaptation. Nat. Commun. 2014, 5, 3978. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.T.; Taylor, W.R.; Thornton, J.M. The rapid generation of mutation data matrices from protein sequences. Comput. Appl. Biosci. 1992, 8, 275–282. [Google Scholar] [CrossRef]
- Liu, L.; McNeilage, R.T.; Shi, L.-x.; Theg, S.M. ATP requirement for chloroplast protein import is set by the Km for ATP hydrolysis of stromal Hsp70 in Physcomitrella patens. Plant Cell 2014, 26, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.F.; Liu, Z.; Rochfort, S.; Mouradov, A. Lipid production in aquatic plant Azolla at vegetative and reproductive stages and in response to abiotic stress. Plant Physiol. Biochem. 2018, 124, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, H.; Liu, G.; Ma, W.; Li, C.; Huo, H.; He, J.; Liu, L. PpSARK regulates moss senescence and salt tolerance through ABA related pathway. Int. J. Mol. Sci. 2018, 19, 2609. [Google Scholar] [CrossRef]
- Pu, X.; Yang, L.; Liu, L.; Dong, X.; Chen, S.; Chen, Z.; Liu, G.; Jia, Y.; Yuan, W.; Liu, L. Genome-wide analysis of the MYB transcription factor superfamily in Physcomitrella patens. Int. J. Mol. Sci. 2020, 21, 975. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Z.; Duan, L.; Li, H.; Tang, T.; Chen, L.; Hu, K.; Yang, H.; Liu, L. Regulation of Heat Stress in Physcomitrium (Physcomitrella) patens Provides Novel Insight into the Functions of Plant RNase H1s. Int. J. Mol. Sci. 2022, 23, 9270. https://doi.org/10.3390/ijms23169270
Yang Z, Duan L, Li H, Tang T, Chen L, Hu K, Yang H, Liu L. Regulation of Heat Stress in Physcomitrium (Physcomitrella) patens Provides Novel Insight into the Functions of Plant RNase H1s. International Journal of Molecular Sciences. 2022; 23(16):9270. https://doi.org/10.3390/ijms23169270
Chicago/Turabian StyleYang, Zhuo, Liu Duan, Hongyu Li, Ting Tang, Liuzhu Chen, Keming Hu, Hong Yang, and Li Liu. 2022. "Regulation of Heat Stress in Physcomitrium (Physcomitrella) patens Provides Novel Insight into the Functions of Plant RNase H1s" International Journal of Molecular Sciences 23, no. 16: 9270. https://doi.org/10.3390/ijms23169270
APA StyleYang, Z., Duan, L., Li, H., Tang, T., Chen, L., Hu, K., Yang, H., & Liu, L. (2022). Regulation of Heat Stress in Physcomitrium (Physcomitrella) patens Provides Novel Insight into the Functions of Plant RNase H1s. International Journal of Molecular Sciences, 23(16), 9270. https://doi.org/10.3390/ijms23169270