
Generation and Identification of the Number of Copies of Exogenous Genes and the T-DNA Insertion Site in SCN-Resistance Transformation Event ZHs1-2



Abstract
:1. Introduction
2. Results
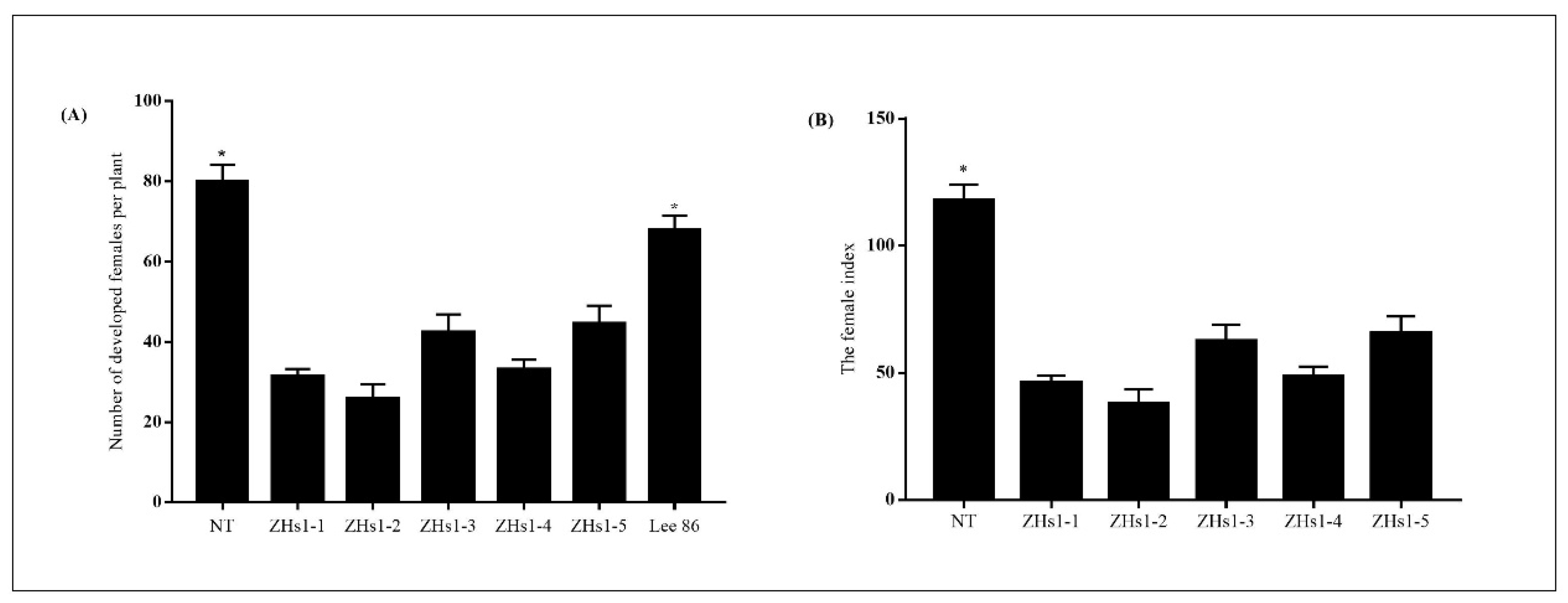
2.1. Generation, Selection, and Inheritances of Transgenic Soybean Lines for Expression of Hs1pro−1 and Bar Genes
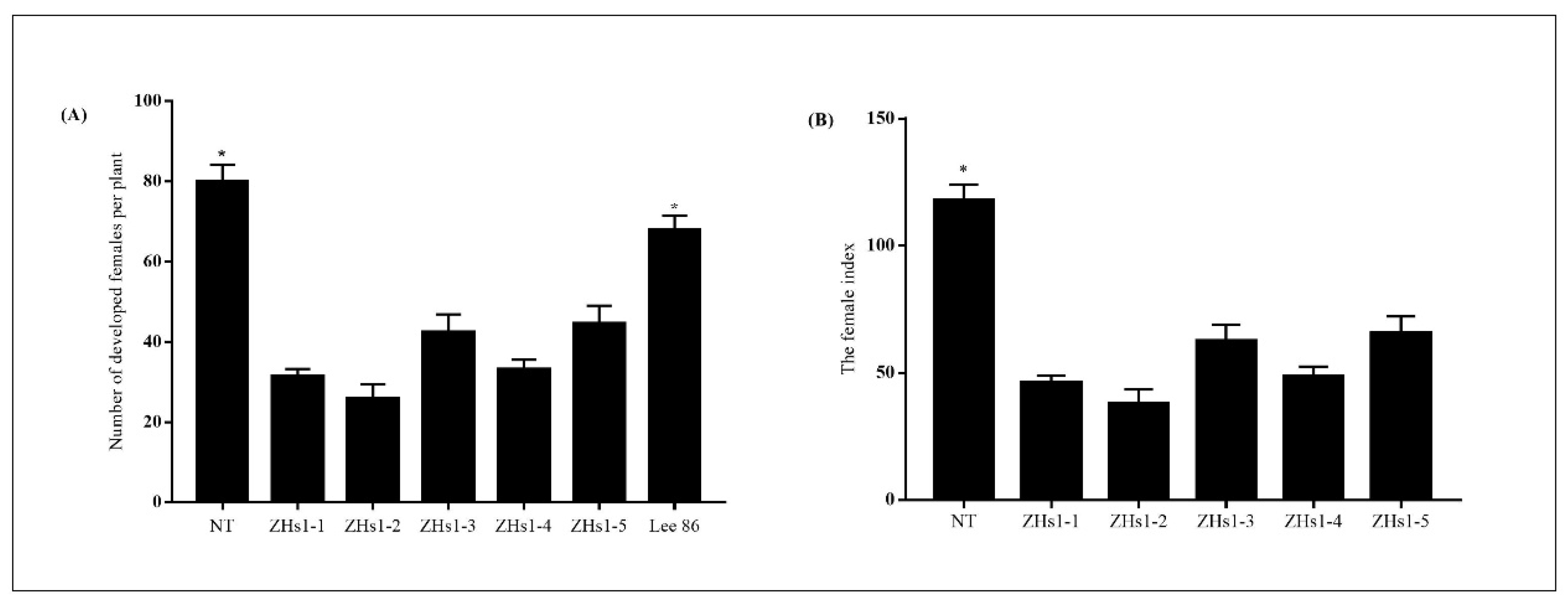
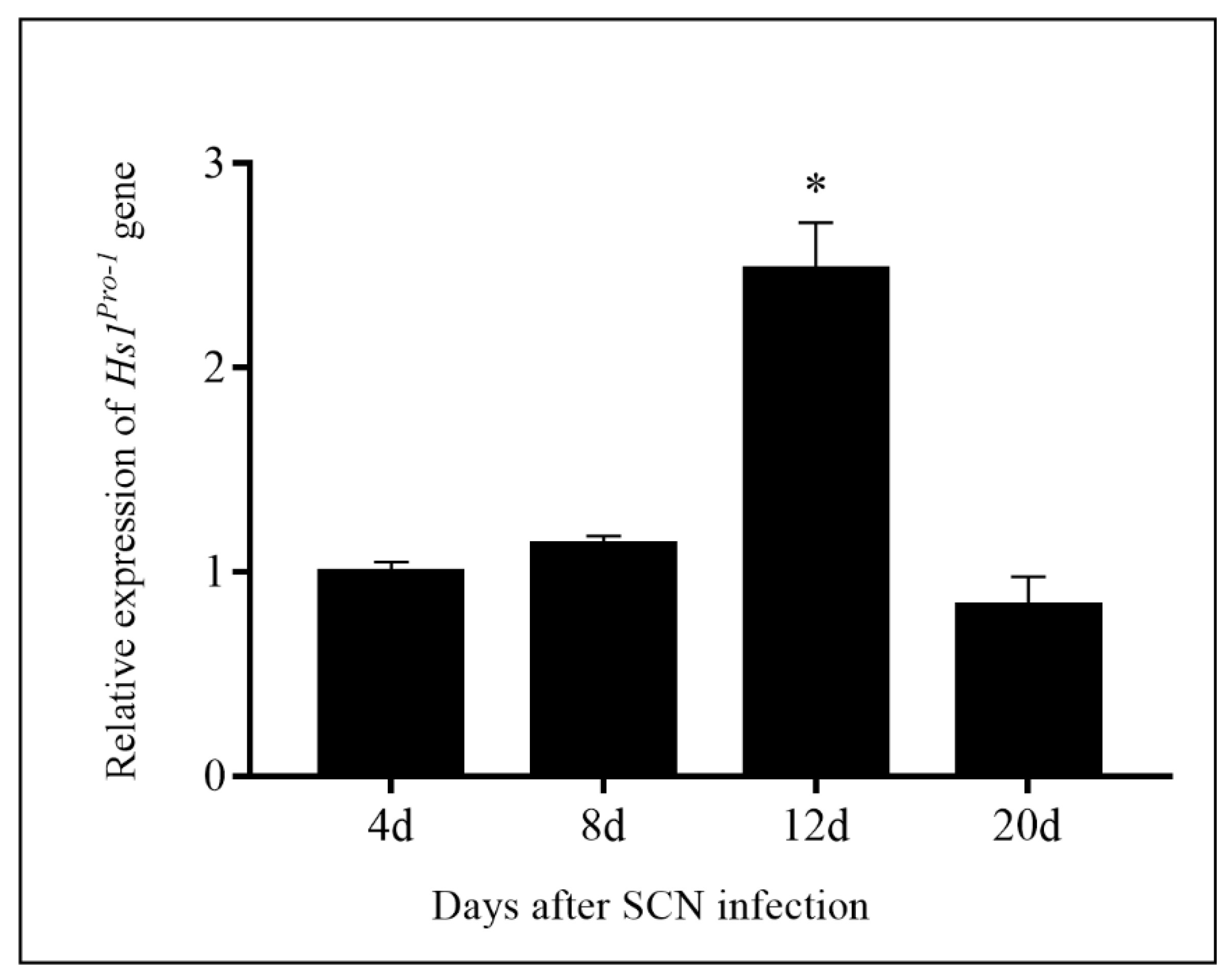
2.2. SCN Resistance in Transformation Events
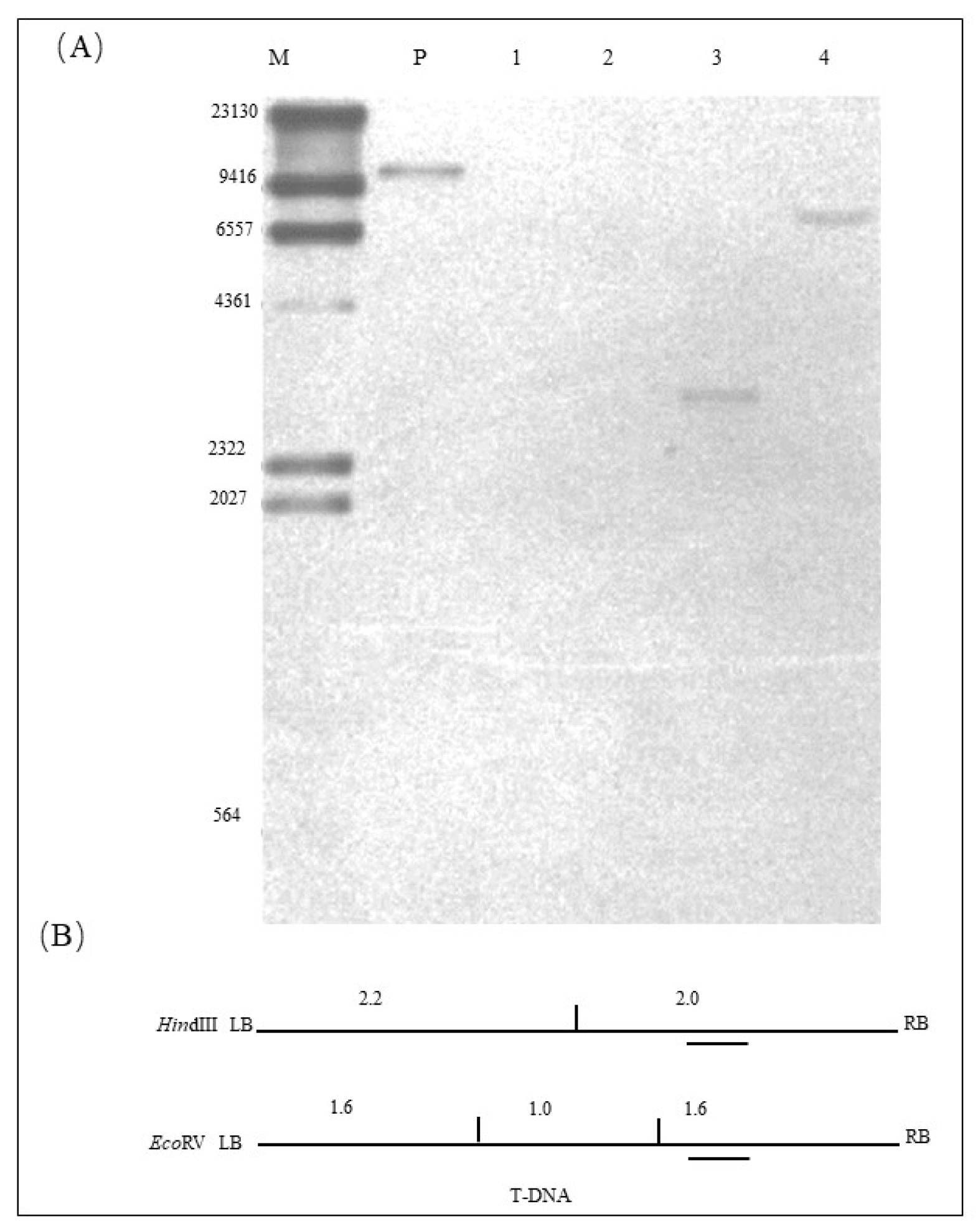
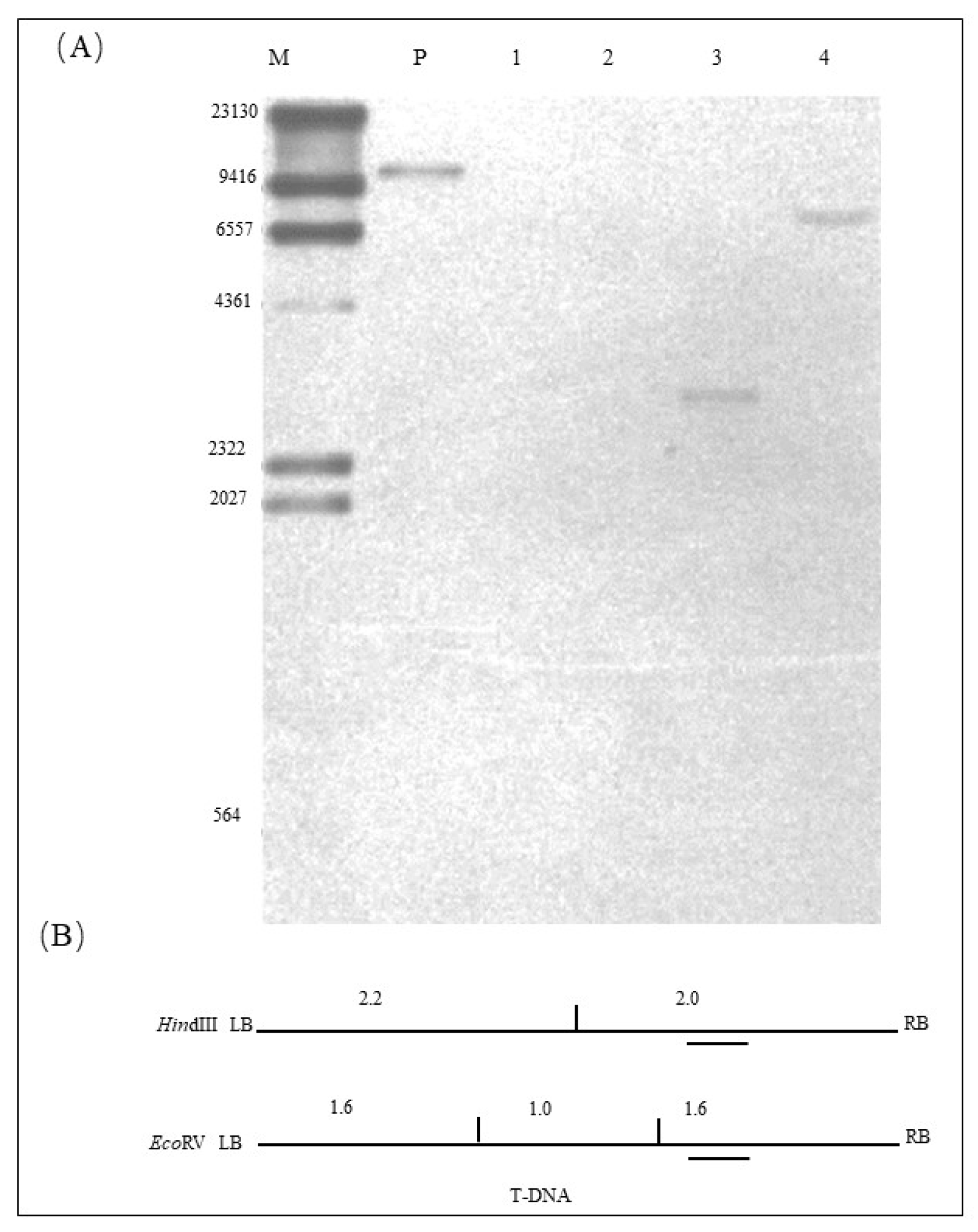
2.3. Copy Number of Exogenous Gene in ZHs1-2 Transformation Event
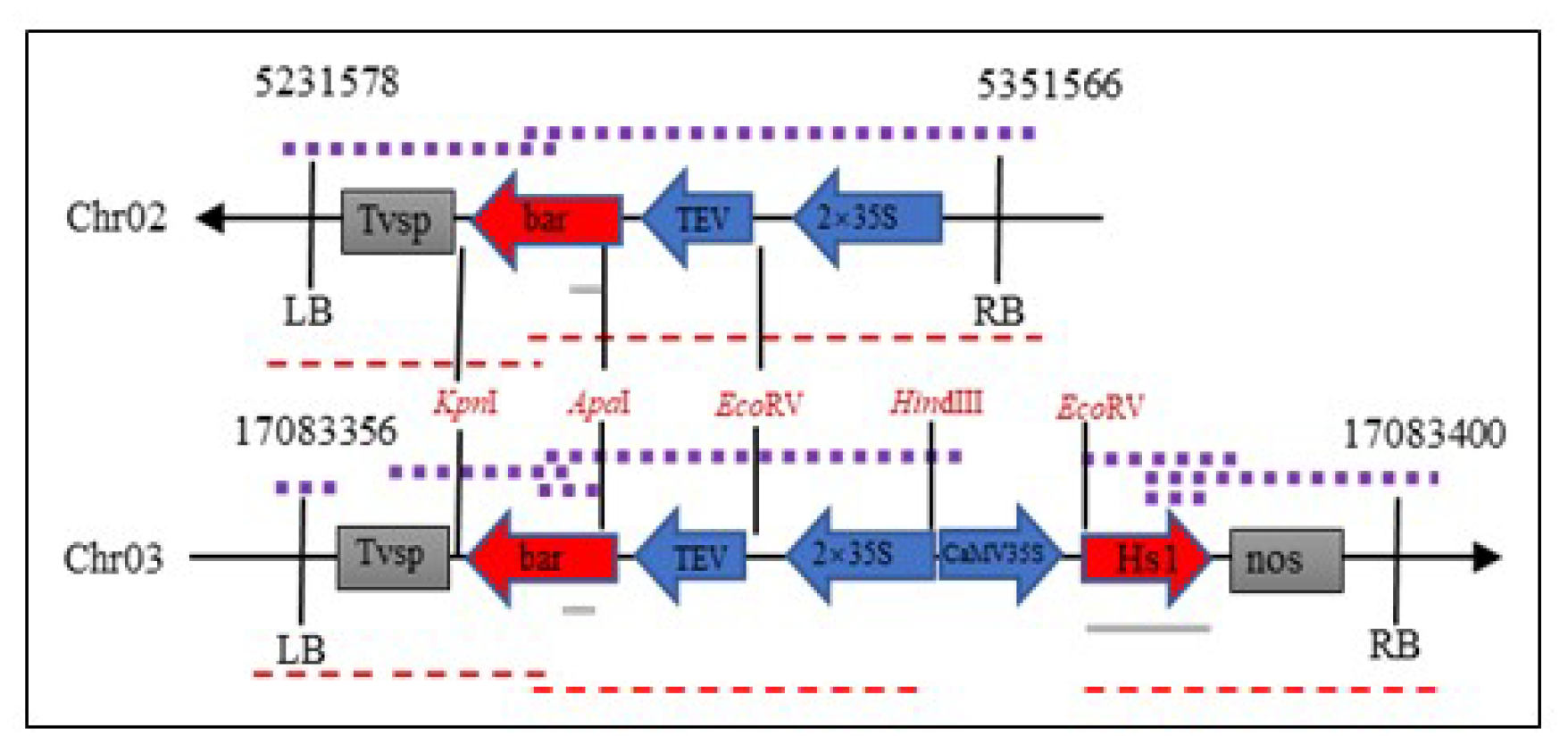
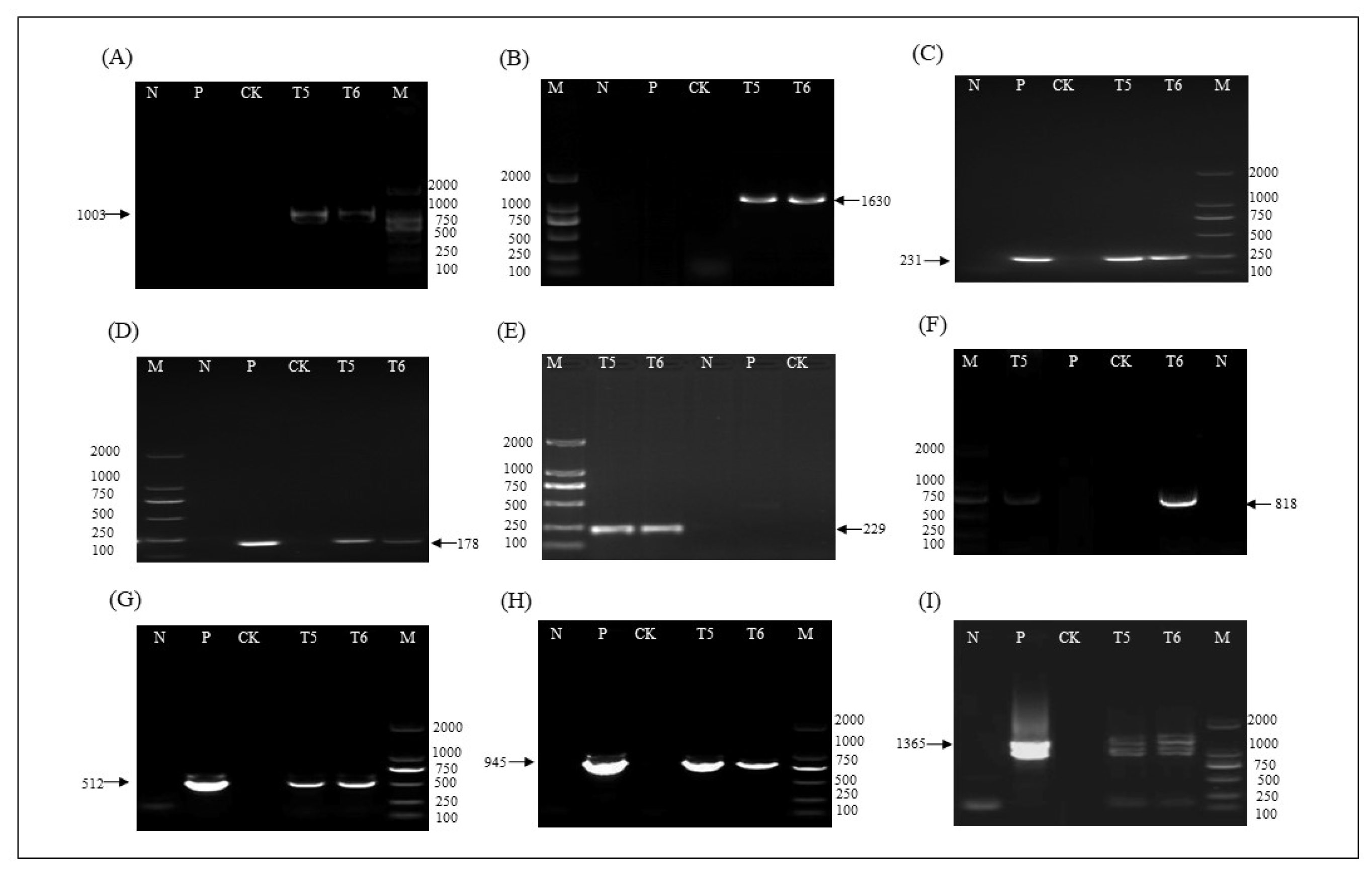
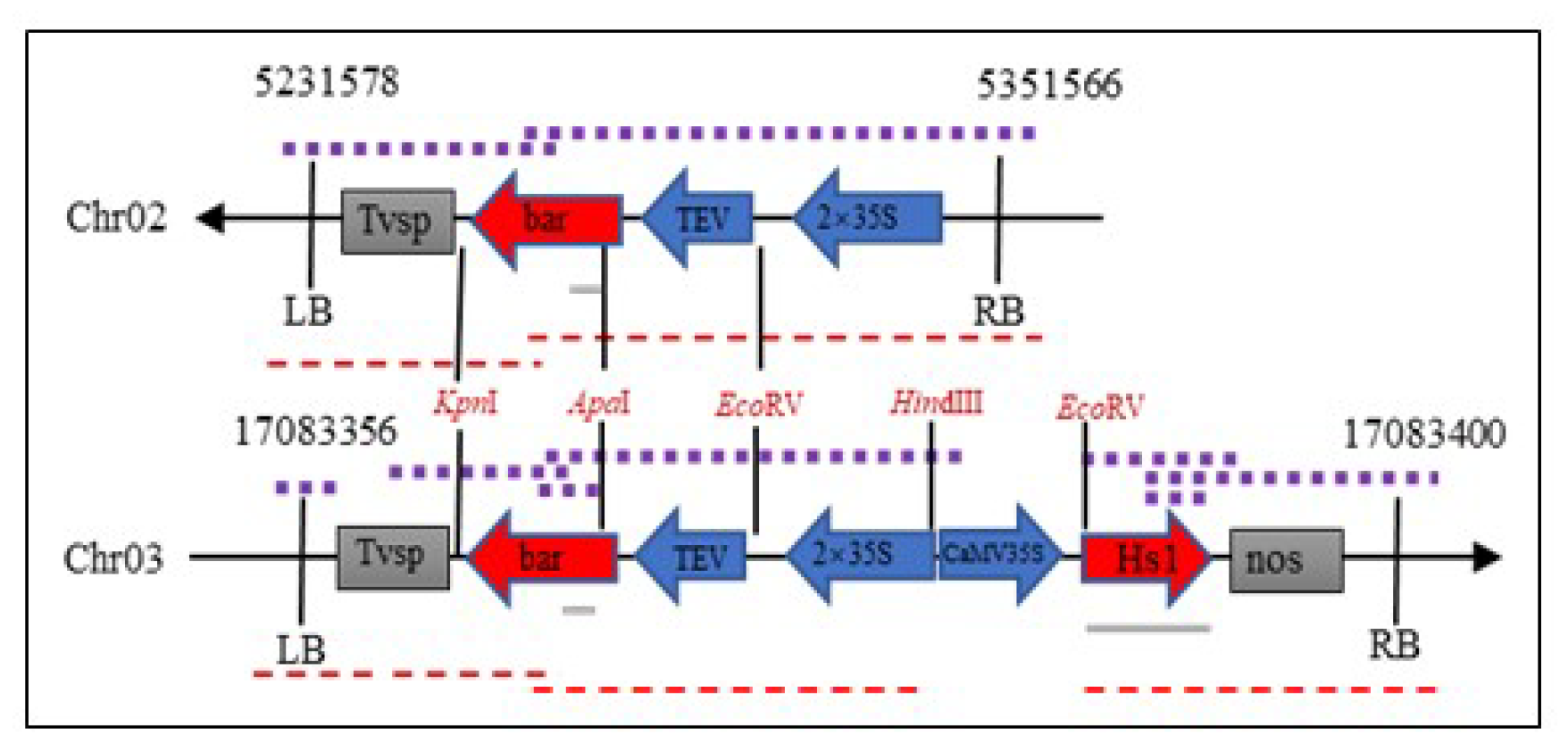
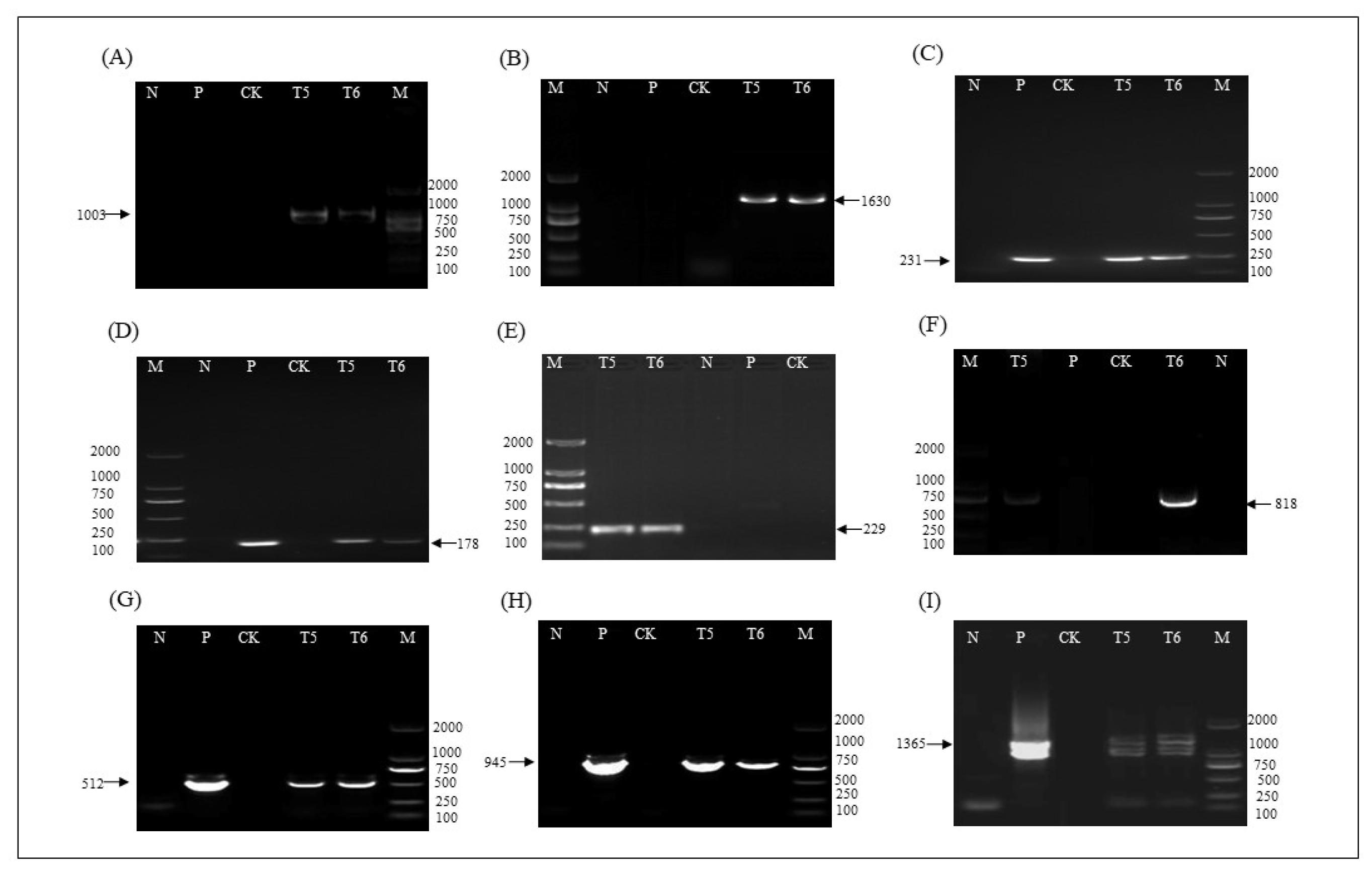
2.4. T-DNA Insertion Sites of ZHs1-2 Transformation Event
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. The ZHs1-2 Transformation Events Generation
4.3. Resistance to SCN Race 4 Assay
4.4. RNA Extraction and qRT-PCR Analysis
4.5. Exogenous Gene Copy Number Verification
4.6. T-DNA Insertion Site Analysis and Verification
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, L.L.; Li, J.F.; Shu, Y.; Chen, X.Y.; Tang, G.X. Current situation of soybean production and consumption in China and strategies to improve self-sufficiency rate. Chin. J. Oil Crop Sci. 2022, 44, 242–248. [Google Scholar] [CrossRef]
- Chen, S.; Kurle, J.; Malvick, D.; Potter, B.; Orf, J. Soybean Cyst Nematode Management Guide. 2018. Available online: https://extension.umn.edu/soybean-pest-management (accessed on 18 April 2022).
- Bandara, A.Y.; Weerasooriya, D.K.; Bradley, C.A.; Allen, T.W.; Esker, P.D. Dissecting the economic impact of soybean diseases in the United States over two decades. PLoS ONE 2020, 15, e0231141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niblack, T.L.; Lambert, K.N.; Tylka, G.L. A model plant pathogen from the kingdom Animalia: Heterodera glycines, the soybean cyst nematode. Annu. Rev. Phytopathol. 2006, 44, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Roth, M.G.; Webster, R.; Mueller, D.S.; Chilvers, M.I.; Faske, T.R.; Mathew, F.M.; Bradley, C.A.; Damicone, J.P.; Kabbage, M.K.; Smith, D.L. Integrated management of important soybean pathogens of the United States in changing climate. J. Integr. Pest Manag. 2020, 11, 28. [Google Scholar] [CrossRef]
- Mitchum, M.G. Soybean resistance to the soybean cyst nematode Heterodera glycines: An update. Phytopathology 2016, 106, 1444–1450. [Google Scholar] [CrossRef] [Green Version]
- Kahn, T.W.; Duck, N.B.; Mccarville, M.T.; Schouten, L.C.; Daum, J. A Bacillus thuringiensis Cry protein controls soybean cyst nematode in transgenic soybean plants. Nat. Commun. 2021, 12, 13. [Google Scholar] [CrossRef]
- Piya, S.; Hawk, T.; Patel, B.; Baldwin, L.; Rice, J.H.; Stewart, N.C.; Hewezi, T., Jr. Kinase-dead mutation: A novel strategy for improving soybean resistance to soybean cyst nematode Heterodera glycines. Mol. Plant Pathol. 2022, 23, 417–430. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, Q.; Zhang, J.; Niu, L.; Yang, J.; Liu, X.; Xing, G.; Zhong, X.; Yang, X. Enhanced resistance to soybean cyst nematode in transgenic soybean via host-induced silencing of vital Heterodera glycines genes. Transgenic Res. 2022, 31, 239–248. [Google Scholar] [CrossRef]
- Brucker, E.; Carlson, S.; Wright, E.; Niblack, T.; Diers, B. Rhg1 alleles from soybean PI 437654 and PI 88788 respond differentially to isolates of Heterodera glycines in the greenhouse. Theor. Appl. Genet. 2005, 11, 44–49. [Google Scholar] [CrossRef]
- Liu, S.M.; Kandoth, P.K.; Warren, S.D.; Yeckel, G.; Heinz, R.; Alden, J.; Yang, C.L.; Jamai, A.; El-Mellouki, T.; Juvale, P.S.; et al. A soybean cyst nematode resistance gene points to a new mechanism of plant resistance to pathogens. Nature 2012, 492, 256–260. [Google Scholar] [CrossRef]
- Cai, D.; Kleine, M.; Kifle, S.; Harloff, H.J.; Sandal, N.N.; Marcker, K.A.; Klein-Lankhorst, R.M.; Salentijn, E.M.J.; Lange, W.; Stiekema, W.J.; et al. Positional cloning of a gene for nematode resistance in sugar beet. Science 1997, 275, 832–834. [Google Scholar] [CrossRef]
- Thurau, T.; Kifle, S.; Jung, C.; Cai, D. The promoter of the nematode resistance gene Hs1pro-1 activates a nematode-responsive and feeding site-specific gene expression in sugar beet (Beta vulgaris L.) and Arabidopsis thaliana. Plant Mol. Biol. 2003, 52, 643–660. [Google Scholar] [CrossRef]
- McLean, M.D.; Hoover, G.J.; Bancroft, B. Identification of the full-length Hs1pro-1 coding sequence and preliminary evaluation of soybean cyst nematode resistance in soybean transformed with Hs1pro-1 cDNA. Can. J. Bot. 2007, 85, 437–441. [Google Scholar] [CrossRef]
- Zhong, X.B.; Zhou, Q.Z.; Cui, N.; Cai, D.G.; Tang, G.X. BvcZR3 and BvHs1pro-1 genes pyramiding enhanced beet cyst nematode (Heterodera schachtii Schm.) resistance in oilseed rape (Brassica napus L.). Int. J. Mol. Sci. 2019, 20, 1740. [Google Scholar] [CrossRef] [Green Version]
- Yuan, C.P.; Zhou, G.A.; Li, Y.H.; Wang, K.J.; Wang, Z.; Li, X.H.; Chang, R.Z.; Qiu, L.J. Cloning and sequence diversity analysis of GmHs1pro-1 in Chinese domesticated and wild soybeans. Mol. Breed. 2008, 22, 593–602. [Google Scholar] [CrossRef]
- Wei, L.; Li, Y.H.; Lu, W.G.; Chang, R.Z.; Qiu, L.J. Inducing expression analysis of Gmhs1pro-1 by inoculating soybean cyst nematode 4 race. J. Agric. Biotechnol. 2011, 19, 77–84. [Google Scholar] [CrossRef]
- ISAAA. Brief 55: Executive Summary Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier. Available online: default (accessed on 14 May 2022).
- Dany, M.; David, D.; Sandrine, H.; Jana, Z.; Kristina, G.N. Target amplification strategy allowing quantitative on-chip detection of GMOs. Nucletic Acids Res. 2008, 36, 11. [Google Scholar] [CrossRef]
- Kanwar, P.; Ghosh, S.; Sanyal, S.K.; Pandey, G.K. Identification of gene copy number in the transgenic plants by quantitative polymerase chain reaction (qPCR). Methods Mol. Biol. 2022, 2392, 161–171. [Google Scholar] [CrossRef]
- Ahmad, A.O.; Marty, G.H.D.; James, H.G.; Jude, W.G. Estimation transgene copy number in transformed citrus plants by quantitative multiplex real-time PCR. Biotechnol. Prog. 2008, 24, 1241–1248. [Google Scholar] [CrossRef]
- Wang, P.; Ge, X.Y.; Li, F.G. Analysis on the insertion sites of exogenous genes of two transgenic insect-resistant cotton L280 and L282. China Cotton 2020, 47, 10–15. [Google Scholar] [CrossRef]
- Iyer, L.M.; Kumpatia, S.P.; Chandrasekharan, M.B.; Hall, T.C. Transgene silencing in monocots. Plant Mol. Biol. 2000, 43, 322–346. [Google Scholar] [CrossRef] [PubMed]
- James, V.A.; Avart, C.; Worland, B.; Snape, J.W.; Vain, P. The relationship between homozygous and hemizygous transgene expression levels over generation in population of transgenic rice plants. Theor. Appl. Genet. 2002, 104, 553–561. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Newton, R.J.; Weidner, D.A. Genetic transformation and gene silencing mediated by multiple copies of a transgene in eastern white pine. J. Exp. Bot. 2007, 58, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Southern, E.M. Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 1975, 98, 503–517. [Google Scholar] [CrossRef]
- Wei, Y.D.; Mao, X.H.; He, W.; Wang, J.L.; Lian, L.; Xu, X.B.; Lin, Y.L.; Xie, H.A.; Zhang, J.F. Determination of the copy number of exogenous gene in transgenic rice (Oryza sativa) by multiplex qPCR. J. Agric. Biotechnol. 2017, 25, 2072–2078. [Google Scholar] [CrossRef]
- Zhong, X.F.; Yang, J.; He, H.L.; Niu, L.; Xing, G.J.; Yang, X.D. Analysis of the T-DNA flanking sequences and event-specific PCR detection of high-content oleic acid transgenic soybean (Glycine max) based on genome re-sequencing. J. Agric. Biotechnol. 2018, 26, 2017–2026. [Google Scholar] [CrossRef]
- Xu, J.M.; Hu, H.; Mao, W.X.; Mao, C.Z. Identifying T-DNA insertion site(s) of transgenic plants by whole-genome resequencing. Heredita 2018, 40, 676–682. [Google Scholar] [CrossRef]
- Lu, W.G.; Gai, J.Y. Advances in resistance to soybean cyst nematode (Heterodera glycines Ichinohe) and resistant molecular markers in soybean (Glycines max Merr.). Soybean Sci. 2004, 23, 59–65, (in Chinese English abstract). [Google Scholar] [CrossRef]
- Tang, G.X.; Qin, Y.B.; Yang, X.F.; Knecht, K.; Cai, D. Effect of transgenic resistant gene oilseed rape (Brassica napus L.) on the cyst nematode resistance. J. Plant Prot. 2008, 35, 571–572. [Google Scholar] [CrossRef]
- Lechtenberg, B.; Schubert, D.; Forsbach, A.; Gils, M.; Schmidt, R. Neither inverted repeat T-DNA configurations nor arrangements of tandemly repeated transgenes are sufficient to trigger transgene silencing. Plant J. 2003, 34, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Qian, Q.; Wang, H.H.; Huang, D.N. Hereditary behavior of bar gene cassette is complex in rice mediated by particle bombardment. J. Genet. Genom. 2007, 34, 824–835. [Google Scholar] [CrossRef]
- Guo, B.F.; Guo, Y.; Hong, H.L.; Qiu, L.J. Identification of genomic insertion and flanking sequence of G2 EPSPS and GAT transgenes in soybean using whole genome sequencing method. Front. Plant Sci. 2016, 7, 9. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Q.; Li, H.Y.; Hu, X.N.; Shan, Z.H.; Tang, G.X. Agrobacterium tumefaciens mediated transformation of RNAi CP gene into soybean (Glycine max L.). Acta Agron. Sin. 2013, 39, 1594–1601. [Google Scholar] [CrossRef]
- Matthews, B.F.; Youssef, R.M. Soybean Cyst Nematode, Heterodera glycines, Infection assay using soybean roots. Bio-Protocol 2016, 6, e1707. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Batches | Number of Explants | Number of Rooting Plants | Identification of T0 Putative Transgenic Rooting Plants | Transformation Efficiency % | |||
|---|---|---|---|---|---|---|---|
| Herbicide Tolerance Assay | Quick Bar Protein Assay | PCR for Hs1 Gene | PCR for Bar Gene | ||||
| X1 | 156 | 0 | 0 | 0 | 0 | 0 | 0.00 |
| X2 | 112 | 6 | 4 | 4 | 4 | 4 | 3.57 |
| X3 | 180 | 0 | 0 | 0 | 0 | 0 | 0.00 |
| X4 | 120 | 8 | 4 | 4 | 4 | 4 | 3.33 |
| X5 | 270 | 8 | 4 | 4 | 4 | 4 | 1.48 |
| L1 | 150 | 13 | 5 | 5 | 5 | 5 | 3.33 |
| L2 | 200 | 20 | 10 | 10 | 10 | 10 | 5.00 |
| L3 | 200 | 14 | 12 | 12 | 12 | 12 | 6.00 |
| Total | 1388 | 69 | 39 | 39 | 39 | 39 | 2.81 |
| Transgenic Plant Code | Number of T1 Seeds | Number of Positive T1 Seeds | T1 Segregation Ratio (Positive:Negative) | χ2 Value | Significant χ2 Value |
|---|---|---|---|---|---|
| 150 | 35 | 3 | 1:10 | 78.87 | No |
| 152 | 40 | 32 | 4:1 | 0.3 | Yes |
| 154 | 4 | 0 | 0 | ||
| 156 | 10 | 4 | 2:3 | 4.8 | No |
| 159 | 11 | 0 | 0 | ||
| 163 | 33 | 18 | 6:5 | 6.31 | No |
| 164 | 11 | 0 | 0 | ||
| 171 | 2 | 0 | 0 | ||
| 173 | 6 | 0 | 0 | ||
| 174 | 16 | 0 | 0 | ||
| 175 | 5 | 2 | 2:3 | 1.67 | Yes |
| 178 | 25 | 0 | 0 | ||
| 186 | 5 | 0 | 0 | ||
| 187 | 4 | 3 | 3:1 | 0.33 | Yes |
| 188 | 3 | 1 | 1:2 | 1 | Yes |
| 189 | 4 | 0 | 0 | ||
| 190 | 3 | 0 | 0 | ||
| 191 | 9 | 0 | 0 | ||
| 192 | 13 | 6 | 1:1.67 | 5.77 | No |
| 193 | 3 | 0 | 0 | ||
| 194 | 25 | 0 | 0 | ||
| 195 | 5 | 0 | 0 | ||
| 196 | 12 | 10 | 5:1 | 0.11 | Yes |
| Name of Primers | Primer Sequence (5′-3′) | Location in Vector (bp) | Amplified Size (bp) | Usage |
|---|---|---|---|---|
| LC0266 | CATTTCACCCTAGTATAACCC | genomic | 1003 | Detection of Chr02 left boundary sequence, Tvsp, and Bar gene |
| LC0264 | CTGGCATGACGTGGGTTT | 7314–7331 | ||
| LC0084 | CCAGAAACCCACGTCATGCCA | 7310–7330 | 1630 | Detection of Chr02 right boundary sequence, 2 × 35S, and Bar gene |
| LC0270 | ATTGGAGTGGCAAAGGGA | genomic | ||
| LC0236 | CAGGTGGGTGTAGAGCGTG | 7472–7490 | 231 | Detection of Bar gene |
| LC0237 | GTCAACTTCCGTACCGAGCC | 7683–7702 | ||
| LC0222 | TTGCTGTGATTGGTGGTTCTAC | 9816–9837 | 178 | Detection of Hs1pro−1 gene |
| LC0223 | TTCGCAGTCCGATTCTTCC | 9975–9993 | ||
| LC0276 | TTGGGGAAGGAAAAGAAT | genomic | 229 | Detection of Chr03 left boundary sequence |
| LC0277 | TTGTCTAAGCGTCAATT | 6453–6469 | ||
| LC0222 | TTGCTGTGATTGGTGGTTCTAC | 9816–9837 | 818 | Detection of Chr03 left boundary sequence, nos, and Hs1pro−1 gene |
| LC0267 | CTGCGAGTTGTGAGTTGTGGT | genomic | ||
| LC0238 | TGGAACAAGGGCAGAAGA | 7060–7077 | 512 | Detection of Tvsp and Bar gene |
| LC0083 | GAAGGCACGCAACGCCTACGA | 7551–7571 | ||
| LC0220 | ATGAGAAGGTGTGGGTATAGTTTG | 9152–9175 | 945 | Detection of nos and Hs1pro−1 gene |
| LC0244 | GCAAGACCGGCAACAGGA | 10,079–10,096 | ||
| LC0236 | CAGGTGGGTGTAGAGCGTG | 7472–7490 | 1365 | Detection of 2 × 35S and Bar gene |
| LC0242 | AGGAGGTTTCCGGATATTACC | 8816–8836 |
| Traits | T6 ZHs1-2 | NT |
|---|---|---|
| Plant height(cm) | 78.6 ± 0.9 | 78.6 ± 1.9 |
| Height of bottom pod (cm) | 8.6 ± 0.7 | 7.6 ± 1.2 |
| Number of main stems | 19.2 ± 0.7 | 19.6 ± 0.4 |
| Number of branches | 4.2 ± 0.7 | 4.6 ± 0.8 |
| Number of pods per plant | 61.8 ± 5.2 | 47.0 ± 6.1 |
| Number of seeds per plant | 155.0 ± 13.2 | 114.8 ± 17.4 |
| Grain yield per plant | 26.0 ± 2.2 | 19.1 ± 2.9 |
| 100-seed weight (g) | 16.8 ± 0.1 | 16.7 ± 0.1 |
| Grain yield (kg/hm2) | 1733.5 ± 83.5 | 1495.4 ± 51.6 |
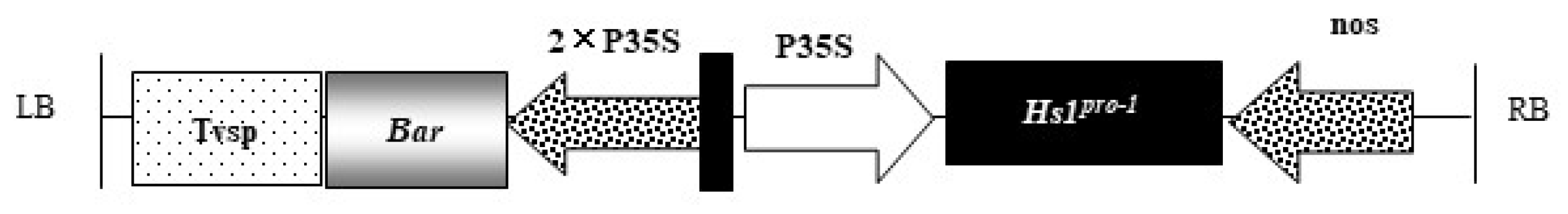
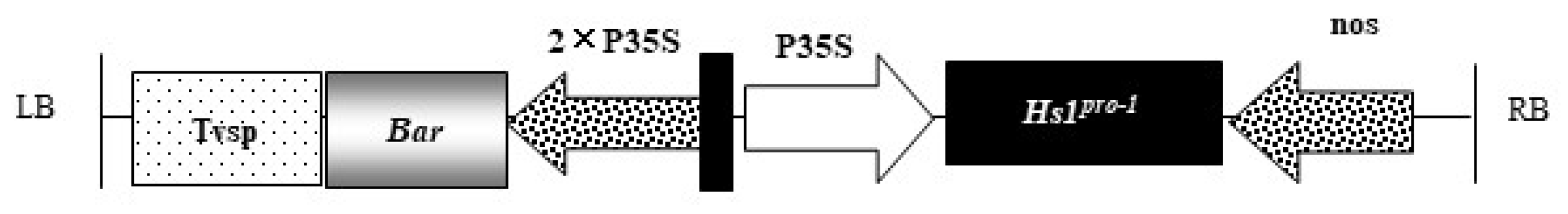
| T-DNA Vector Elements | Location in Vector | Size (bp) | Function |
|---|---|---|---|
| LB | 6426–6451 | 26 | T-DNA left border sequence of Agrobacterium C58, required for T-DNA transfer |
| Tvsp | 6741–7292 | 552 | Soybean storage protein terminator |
| Bar | 7294–7869 | 564 | Code PAT protein, relieves toxicity of glufosinate |
| TEV enhancer | 7870–8014 | 145 | 5’ leader sequence of tobacco etch virus to enhance transcriptional level |
| 2 × 35S | 8015–8731 | 717 | The tandem 35S promoter of cauliflower mosaic virus (CaMV) |
| CaMV 35S | 8732–9145 | 414 | The 35S promoter of cauliflower mosaic virus |
| Hs1 | 9152–10,000 | 849 | Encodes sugar beet Hs1pro−1 protein to improve the soybean cyst nematode resistance |
| nos | 10,067–10,296 | 230 | The terminator of the nopaline synthase gene to promote transcription termination |
| RB | 10,693–10,717 | 26 | T-DNA right border sequence of Agrobacterium C58, required for T-DNA transfer. |
| Target Gene Primers | Primer Sequence | Length of Amplified Fragments (bp) |
|---|---|---|
| Bar-F | CAGCTGCCAGAAACCCACGT | 436 |
| Bar-R | CTGCACCATCGTCAACCACT | |
| Hs1-1-F | GCTCTAGAATGAGAAGGTGTGGGTATAG | 849 |
| Hs1-1-R | GCTCTAGATCATTGTTTCGCAGTCCG | |
| Hs1-2-F | GGCACCATCCAAACTCGG | 543 |
| Hs1-2-R | CGAATAAGTGAGAGGATC |
| Name of Primer | Primer Sequence (5′-3′) | Location in Vector (bp) | Amplified Size (bp) | Usage |
|---|---|---|---|---|
| LC0220 | ATGAGAAGGTGTGGGTATAGTTTG | 9152–9175 | 849 | Check Hs1pro−1 gene |
| LC0221 | TCATTGTTTCGCAGTCCGATT | 9980–10,000 | ||
| LC0236 | CAGGTGGGTGTAGAGCGTG | 7472–7490 | 231 | Check Bar gene |
| LC0237 | GTCAACTTCCGTACCGAGCC | 7683–7702 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, G.; Zhong, X.; Hong, W.; Li, J.; Shu, Y.; Liu, L. Generation and Identification of the Number of Copies of Exogenous Genes and the T-DNA Insertion Site in SCN-Resistance Transformation Event ZHs1-2. Int. J. Mol. Sci. 2022, 23, 6849. https://doi.org/10.3390/ijms23126849
Tang G, Zhong X, Hong W, Li J, Shu Y, Liu L. Generation and Identification of the Number of Copies of Exogenous Genes and the T-DNA Insertion Site in SCN-Resistance Transformation Event ZHs1-2. International Journal of Molecular Sciences. 2022; 23(12):6849. https://doi.org/10.3390/ijms23126849
Chicago/Turabian StyleTang, Guixiang, Xuanbo Zhong, Wei Hong, Jianfei Li, Yue Shu, and Lulu Liu. 2022. "Generation and Identification of the Number of Copies of Exogenous Genes and the T-DNA Insertion Site in SCN-Resistance Transformation Event ZHs1-2" International Journal of Molecular Sciences 23, no. 12: 6849. https://doi.org/10.3390/ijms23126849
APA StyleTang, G., Zhong, X., Hong, W., Li, J., Shu, Y., & Liu, L. (2022). Generation and Identification of the Number of Copies of Exogenous Genes and the T-DNA Insertion Site in SCN-Resistance Transformation Event ZHs1-2. International Journal of Molecular Sciences, 23(12), 6849. https://doi.org/10.3390/ijms23126849