
Long Non-Coding RNAs as Functional Codes for Oral Cancer: Translational Potential, Progress and Promises


Abstract
1. Introduction
2. Mechanism of Oral Cancer-Associated lncRNAs in Tumorigenesis
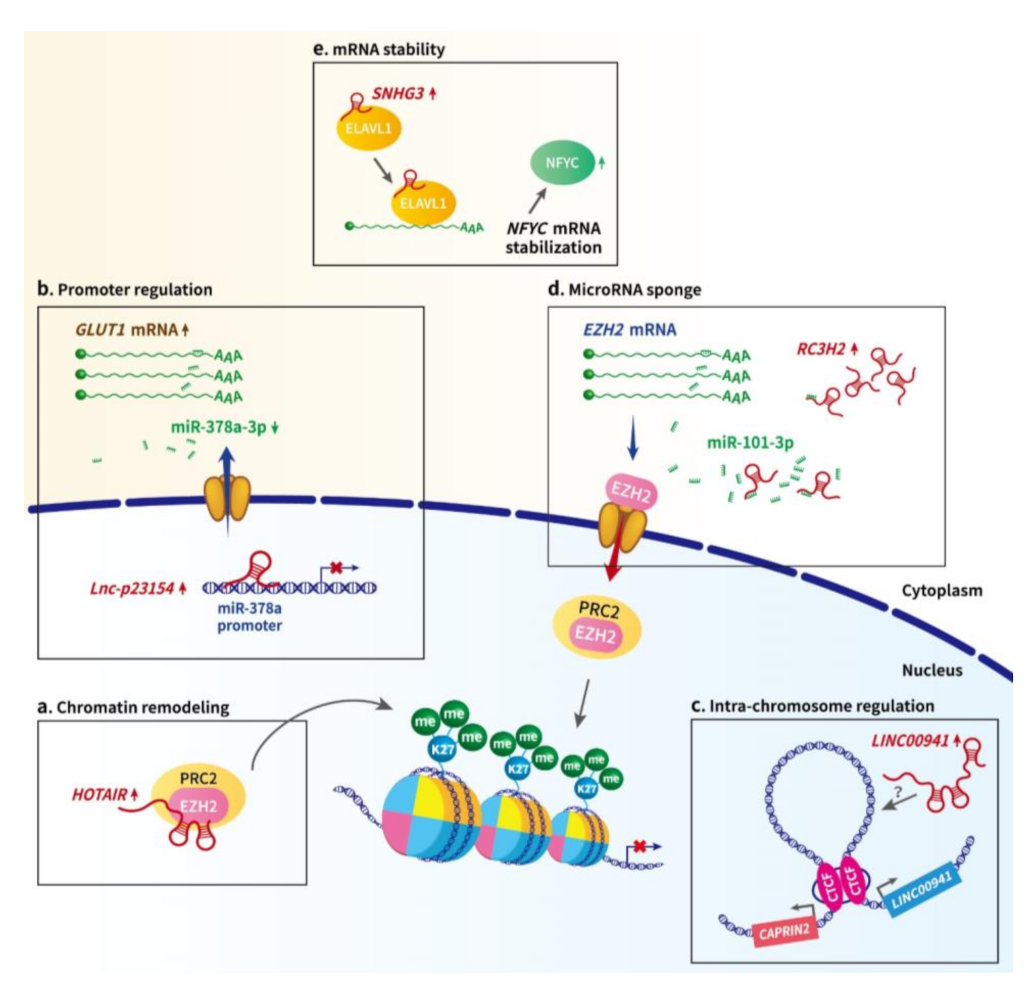
2.1. LncRNA-Mediated Epigenetic and Transcriptional Regulation in Oral Cancer
2.2. LncRNA-Mediated Post-Transcriptional Regulation in Oral Cancer
3. LncRNAs Involved in Crucial Signaling Pathways
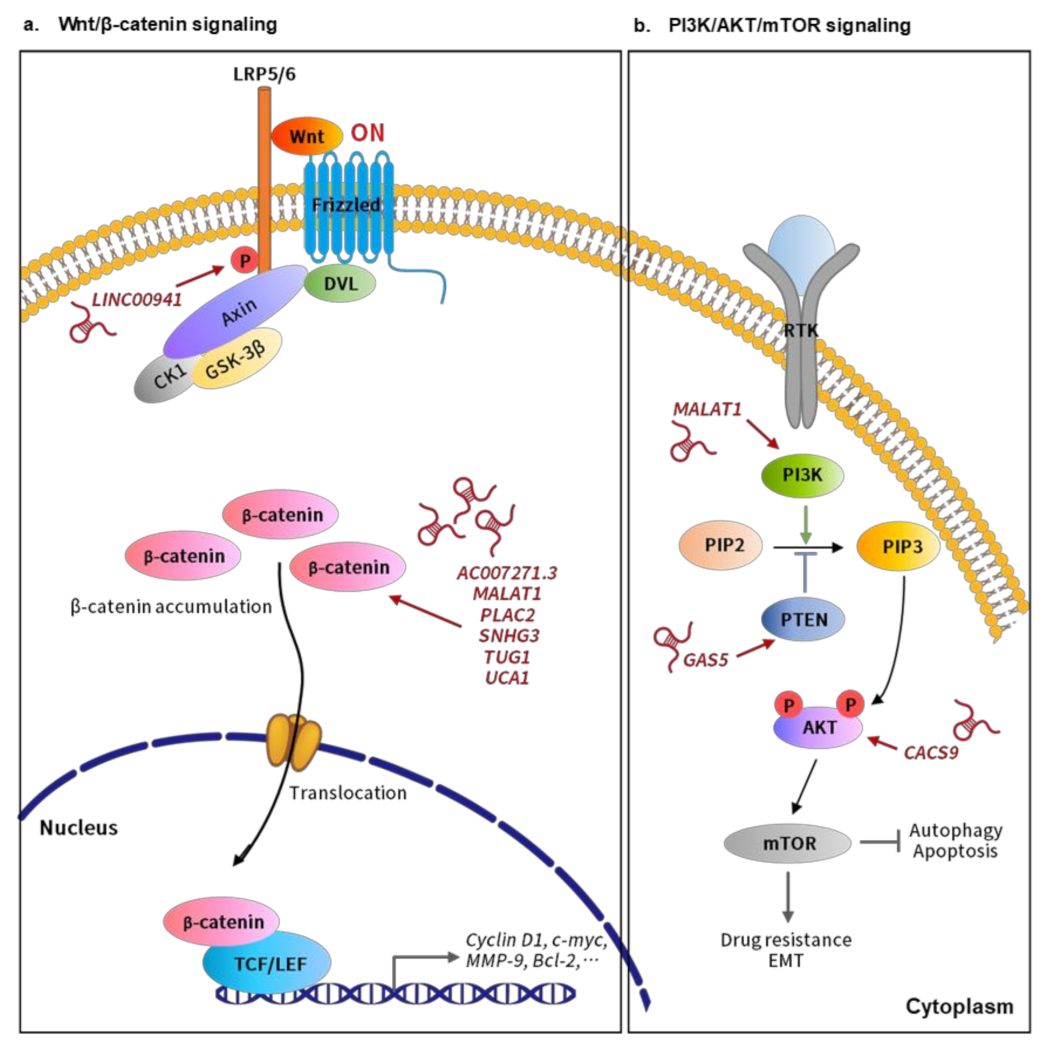
3.1. Wnt/β-Catenin Signaling-Related lncRNAs in Oral Cancer
3.2. PI3K/AKT/mTOR Signaling-Related lncRNAs in Oral Cancer
3.3. Other Oncogenic Signaling Pathways-Related Lncrnas in Oral Cancer
4. LncRNAs as Potential Biomarkers and Therapeutic Targets for Oral Cancer: Challenges and Potential
5. Conclusions and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ASO | Antisense oligonucleotide |
| Bcl-2 | B-cell lymphoma 2 |
| CAPRIN2 | Caprin family member 2 |
| ceRNA | Competing endogenous RNA |
| CTCF | CCCTC-binding factor |
| EMT | Epithelial-mesenchymal transition |
| EZH2 | Enhancer of zeste homolog 2 |
| GLUT1 | Glucose transport 1 |
| H3K27me3 | Histone H3 lysine 27 trimethylation |
| Hes1 | Hairy and enhancer of split 1 |
| HIF-1α | Hypoxia-inducible factor 1-alpha |
| HNSCC | Head and neck squamous cell carcinoma |
| JAK | Janus kinases |
| LC3B | Microtubule-associated protein 1-light chain 3 beta |
| LATS1 | Large tumor suppressor kinase 1 |
| LncRNA | Long non-coding RNA |
| miRNA | microRNA |
| MOB | Monopolar spindle-one-binder protein |
| Notch1 | Neurogenic locus notch homolog protein |
| OSCC | Oral squamous cell carcinoma |
| PRC2 | Polycomb repressive complex 2 |
| RBP | RNA binding protein |
| SOCS | Suppressor of cytokine signaling |
| STAT | Signal transducer and activator of transcription |
| TGF-β | Transforming growth factor-β |
| TSCC | Tongue squamous cell carcinoma |
| UTR | Untranslated region |
| YAP1 | Yes-associated protein 1 |
| ELAVL1 | Embryonic lethal abnormal vision-like protein 1 |
| HuR | Hu antigen R |
| NFYC | Nuclear transcription factor Y subunit gamma |
| APC | Adenomatous polyposis coli |
| CK1 | Casein kinase 1 |
| Dvl | Dishevelled |
| Fzd | Frizzled |
| GSK-3β | Glycogen synthase kinase-3β |
| MMP-9 | Matrix metallopeptidase 9 |
| LEF | Lymphoid-enhancing factor |
| LRP | Lipoprotein receptor-related protein |
| TCF | T-cell factor |
| AKT | Protein kinase B |
| mTOR | Mammalian target of rapamycin |
| PI3K | Phosphoinositide 3-kinase |
| PIP2 | Phosphatidylinositol 4,5-bisphosphate |
| PIP3 | Phosphatidylinositol 3,4,5-triphosphate |
| PTEN | Phosphatase and tensin homolog |
| RTK | Receptor tyrosine kinase |
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar]
- Forastiere, A.; Koch, W.; Trotti, A.; Sidransky, D. Head and Neck Cancer. N. Engl. J. Med. 2001, 345, 1890–1900. [Google Scholar] [CrossRef] [PubMed]
- Zini, A.; Czerninski, R.; Sgan-Cohen, H.D. Oral cancer over four decades: Epidemiology, trends, histology, and survival by anatomical sites. J. Oral Pathol. Med. 2010, 39, 299–305. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.L.; Riaz, M.K.; Qu, X.; Chan, J.; Meehan, K. Profiling of extracellular vesicles in oral cancer, from transcriptomics to proteomics. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Chen, Y.K.; Huang, H.C.; Lin, L.M.; Lin, C.C. Primary oral squamous cell carcinoma: An analysis of 703 cases in southern Taiwan. Oral Oncol. 1999, 35, 173–179. [Google Scholar] [CrossRef]
- Bagan, J.; Sarrion, G.; Jimenez, Y. Oral cancer: Clinical features. Oral Oncol. 2010, 46, 414–417. [Google Scholar] [CrossRef]
- Wang, W.; Han, S.; Yao, Z.; Li, X.; Huang, P.; Zhang, M.; Chen, Y.; He, J. A Study of Epidemiologic and Recurrence Factors of Oral Cancer. J. Oral Maxillofac. Surg. 2012, 70, 2205–2210. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Thavarool, S.B.; Muttath, G.; Nayanar, S.; Duraisamy, K.; Bhat, P.; Shringarpure, K.; Nayak, P.; Tripathy, J.P.; Thaddeus, A.; Philip, S.B.S. Improved survival among oral cancer patients: Findings from a retrospective study at a tertiary care cancer centre in rural Kerala, India. World J. Surg. Oncol. 2019, 17, 15. [Google Scholar] [CrossRef]
- Neville, B.W.; Day, T.A. Oral Cancer and Precancerous Lesions. CA A Cancer J. Clin. 2002, 52, 195–215. [Google Scholar] [CrossRef]
- Alsahafi, E.; Begg, K.; Amelio, I.; Raulf, N.; Lucarelli, P.; Sauter, T.; Tavassoli, M. Clinical update on head and neck cancer: Molecular biology and ongoing challenges. Cell Death Dis. 2019, 10, 540. [Google Scholar] [CrossRef]
- Uchida, S.; Dimmeler, S. Long Noncoding RNAs in Cardiovascular Diseases. Circ. Res. 2015, 116, 737–750. [Google Scholar] [CrossRef]
- Statello, L.; Guo, C.-J.; Chen, L.-L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Rev. Mol. Cell Biol. 2021, 22, 96–118. [Google Scholar] [CrossRef]
- Wu, X.; Tudoran, O.M.; Calin, G.A.; Ivan, M. The Many Faces of Long Noncoding RNAs in Cancer. Antioxid. Redox Signal. 2017, 29, 922–935. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Kuo, T.-C.; Kung, H.-J.; Shih, J.-W. Signaling in and out: Long-noncoding RNAs in tumor hypoxia. J. Biomed. Sci. 2020, 27, 59. [Google Scholar] [CrossRef] [PubMed]
- Nohata, N.; Abba, M.C.; Gutkind, J.S. Unraveling the oral cancer lncRNAome: Identification of novel lncRNAs associated with malignant progression and HPV infection. Oral. Oncol. 2016, 59, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Pentenero, M.; Bowers, L.M.; Jayasinghe, R.; Yap, T.; Cheong, S.C.; Kerr, A.R.; Farah, C.S.; Alevizos, I. World Workshop on Oral Medicine VII: Clinical evidence of differential expression of lncRNAs in oral squamous cell carcinoma: A scoping review. Oral Dis. 2019, 25 (Suppl. 1), 88–101. [Google Scholar] [CrossRef]
- Pentenero, M.; Bowers, L.; Jayasinghe, R.; Cheong, S.C.; Farah, C.S.; Kerr, A.R.; Alevizos, I. World Workshop on Oral Medicine VII: Functional pathways involving differentially expressed lncRNAs in oral squamous cell carcinoma. Oral Dis. 2019, 25 (Suppl. 1), 79–87. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, X.; Zhu, X.W.; Yang, D.C.; Chen, R.; Jiang, Y.; Xu, T. Long non-coding RNAs in Oral squamous cell carcinoma: Biologic function, mechanisms and clinical implications. Mol. Cancer 2019, 18, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Qiu, W.; Wu, B.; Fang, F. Long non‑coding RNAs are novel players in oral inflammatory disorders, potentially premalignant oral epithelial lesions and oral squamous cell carcinoma (Review). Int. J. Mol. Med. 2020, 46, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Jiang, E.; Shao, Z.; Shang, Z. Long Noncoding RNAs in the Metastasis of Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 10, 3014. [Google Scholar] [CrossRef]
- Palazzo, A.F.; Koonin, E.V. Functional Long Non-coding RNAs Evolve from Junk Transcripts. Cell 2020, 183, 1151–1161. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Liu, H.; You, G. Long non-coding RNA C5orf66-AS1 prevents oral squamous cell carcinoma through inhibiting cell growth and metastasis. Int. J. Mol. Med. 2018, 42, 3291–3299. [Google Scholar] [CrossRef]
- Xing, H.B.; Qiu, H.M.; Li, Y.; Dong, P.F.; Zhu, X.M. Long noncoding RNA CASC2 alleviates the growth, migration and invasion of oral squamous cell carcinoma via downregulating CDK1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4777–4783. [Google Scholar]
- Dong, Y.; Wu, W. Downregulation of lncRNA CASC2 promotes the postoperative local recurrence of early oral squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2019, 276, 605–610. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.; Chen, H.; Bai, Y.; Wang, Q.; Chen, L. Long non-coding RNA CASC2 serves as a ceRNA of microRNA-21 to promote PDCD4 expression in oral squamous cell carcinoma. Onco. Targets Ther. 2019, 12, 3377–3385. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yan, L.; Wang, J.; Lin, L.; Qiu, Y.; Hu, Z.; Wu, J.; Bao, X.; Lin, L.; Wang, R.; et al. Upregulated long noncoding RNA ENST00000470447.1 inhibits cell migration and invasion and predicts better disease-free survival of oral cancer. Head Neck 2019, 41, 2883–2891. [Google Scholar] [CrossRef]
- Jia, B.; Xie, T.; Qiu, X.; Sun, X.; Chen, J.; Huang, Z.; Zheng, X.; Wang, Z.; Zhao, J. Long noncoding RNA FALEC inhibits proliferation and metastasis of tongue squamous cell carcinoma by epigenetically silencing ECM1 through EZH2. Aging 2019, 11, 4990–5007. [Google Scholar] [CrossRef]
- Chen, F.B.; Wu, P.; Zhou, R.; Yang, Q.X.; Zhang, X.; Wang, R.R.; Qi, S.C.; Yang, X. LINC01315 Impairs microRNA-211-Dependent DLG3 Downregulation to Inhibit the Development of Oral Squamous Cell Carcinoma. Front. Oncol. 2020, 10, 556084. [Google Scholar] [CrossRef]
- Jin, Z.; Jiang, S.; Jian, S.; Shang, Z. Long noncoding RNA MORT overexpression inhibits cancer cell proliferation in oral squamous cell carcinoma by downregulating ROCK1. J. Cell Biochem. 2019, 120, 11702–11707. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, L.; Zhang, L.; Wang, Y.; Li, H.; Ren, X.; Wei, F.; Yu, W.; Liu, T.; Wang, X.; et al. Long non-coding RNA HOTAIR promotes tumor cell invasion and metastasis by recruiting EZH2 and repressing E-cadherin in oral squamous cell carcinoma. Int. J. Oncol. 2015, 46, 2586–2594. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Tsai, M.C.; Manor, O.; Wan, Y.; Mosammaparast, N.; Wang, J.K.; Lan, F.; Shi, Y.; Segal, E.; Chang, H.Y. Long noncoding RNA as modular scaffold of histone modification complexes. Science 2010, 329, 689–693. [Google Scholar] [CrossRef]
- Lu, M.Y.; Liao, Y.W.; Chen, P.Y.; Hsieh, P.L.; Fang, C.Y.; Wu, C.Y.; Yen, M.L.; Peng, B.Y.; Wang, D.P.; Cheng, H.C.; et al. Targeting LncRNA HOTAIR suppresses cancer stemness and metastasis in oral carcinomas stem cells through modulation of EMT. Oncotarget 2017, 8, 98542–98552. [Google Scholar] [CrossRef]
- Wang, X.; Liu, W.; Wang, P.; Li, S. RNA interference of long noncoding RNA HOTAIR suppresses autophagy and promotes apoptosis and sensitivity to cisplatin in oral squamous cell carcinoma. J. Oral Pathol. Med. 2018, 47, 930–937. [Google Scholar] [CrossRef]
- Tao, D.; Zhang, Z.; Liu, X.; Zhang, Z.; Fu, Y.; Zhang, P.; Yuan, H.; Liu, L.; Cheng, J.; Jiang, H. LncRNA HOTAIR promotes the invasion and metastasis of oral squamous cell carcinoma through metastasis-associated gene 2. Mol. Carcinog. 2020, 59, 353–364. [Google Scholar] [CrossRef]
- Pisignano, G.; Pavlaki, I.; Murrell, A. Being in a loop: How long non-coding RNAs organise genome architecture. Essays Biochem. 2019, 63, 177–186. [Google Scholar] [PubMed]
- Ai, Y.; Wu, S.; Zou, C.; Wei, H. LINC00941 promotes oral squamous cell carcinoma progression via activating CAPRIN2 and canonical WNT/β-catenin signaling pathway. J. Cell Mol. Med. 2020, 24, 10512–10524. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, X.; Wang, Z.; Hu, Q.; Wu, J.; Li, Y.; Ren, X.; Wu, T.; Tao, X.; Chen, X.; et al. LncRNA-p23154 promotes the invasion-metastasis potential of oral squamous cell carcinoma by regulating Glut1-mediated glycolysis. Cancer Lett. 2018, 434, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Wang, S.; Chen, J.; Wang, Z.; Liang, X.; Wang, X.; Jiang, J.; Lang, J.; Li, L. Long noncoding RNA HAS2-AS1 mediates hypoxia-induced invasiveness of oral squamous cell carcinoma. Mol. Carcinog. 2017, 56, 2210–2222. [Google Scholar] [CrossRef] [PubMed]
- Vigetti, D.; Deleonibus, S.; Moretto, P.; Bowen, T.; Fischer, J.W.; Grandoch, M.; Oberhuber, A.; Love, D.C.; Hanover, J.A.; Cinquetti, R.; et al. Natural Antisense Transcript for Hyaluronan Synthase 2 (HAS2-AS1) Induces Transcription of HAS2 via Protein O-GlcNAcylation*. J. Biol. Chem. 2014, 289, 28816–28826. [Google Scholar] [CrossRef] [PubMed]
- Shih, J.W.; Chiang, W.F.; Wu, A.T.H.; Wu, M.H.; Wang, L.Y.; Yu, Y.L.; Hung, Y.W.; Wang, W.C.; Chu, C.Y.; Hung, C.L.; et al. Long noncoding RNA LncHIFCAR/MIR31HG is a HIF-1α co-activator driving oral cancer progression. Nat. Commun 2017, 8, 15874. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ma, Z.; Feng, L.; Yang, Y.; Tan, C.; Shi, Q.; Lian, M.; He, S.; Ma, H.; Fang, J. LncRNA MIR31HG targets HIF1A and P21 to facilitate head and neck cancer cell proliferation and tumorigenesis by promoting cell-cycle progression. Mol. Cancer 2018, 17, 162. [Google Scholar] [CrossRef]
- Feng, L.; Wang, R.; Lian, M.; Ma, H.; He, N.; Liu, H.; Wang, H.; Fang, J. Integrated Analysis of Long Noncoding RNA and mRNA Expression Profile in Advanced Laryngeal Squamous Cell Carcinoma. PLOS ONE 2016, 11, e0169232. [Google Scholar] [CrossRef] [PubMed]
- Sigova, A.A.; Mullen, A.C.; Molinie, B.; Gupta, S.; Orlando, D.A.; Guenther, M.G.; Almada, A.E.; Lin, C.; Sharp, P.A.; Giallourakis, C.C.; et al. Divergent transcription of long noncoding RNA/mRNA gene pairs in embryonic stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 2876–2881. [Google Scholar] [CrossRef] [PubMed]
- Romero-Barrios, N.; Legascue, M.F.; Benhamed, M.; Ariel, F.; Crespi, M. Splicing regulation by long noncoding RNAs. Nucleic Acids Res. 2018, 46, 2169–2184. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Ma, Y.; Hu, X.; Song, R.; Zhu, L.; Zhong, M. Long non-coding RNA CEBPA-AS1 correlates with poor prognosis and promotes tumorigenesis via CEBPA/Bcl2 in oral squamous cell carcinoma. Cancer Biol. Ther. 2018, 19, 205–213. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Denzler, R.; Agarwal, V.; Stefano, J.; Bartel, D.P.; Stoffel, M. Assessing the ceRNA hypothesis with quantitative measurements of miRNA and target abundance. Mol. Cell 2014, 54, 766–776. [Google Scholar] [CrossRef]
- Denzler, R.; McGeary, S.E.; Title, A.C.; Agarwal, V.; Bartel, D.P.; Stoffel, M. Impact of MicroRNA Levels, Target-Site Complementarity, and Cooperativity on Competing Endogenous RNA-Regulated Gene Expression. Mol. Cell 2016, 64, 565–579. [Google Scholar] [CrossRef]
- Thomson, D.W.; Dinger, M.E. Endogenous microRNA sponges: Evidence and controversy. Nat. Rev. Genet. 2016, 17, 272–283. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Jiang, Y.; Zhou, W.; Zhang, B.; Li, Y.; Xie, F.; Zhang, J.; Wang, X.; Yan, M.; Xu, Q.; et al. Long Noncoding RNA RC3H2 Facilitates Cell Proliferation and Invasion by Targeting MicroRNA-101-3p/EZH2 Axis in OSCC. Mol. Ther. Nucleic Acids 2020, 20, 97–110. [Google Scholar] [CrossRef]
- Wu, J.; Zhao, W.; Wang, Z.; Xiang, X.; Zhang, S.; Liu, L. Long non-coding RNA SNHG20 promotes the tumorigenesis of oral squamous cell carcinoma via targeting miR-197/LIN28 axis. J. Cell Mol. Med. 2019, 23, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Sun, H.; Wang, X.; Yu, X.; Zhang, J.; Guo, B.; Hexige, S. Metabolic changes during malignant transformation in primary cells of oral lichen planus: Succinate accumulation and tumour suppression. J. Cell Mol. Med. 2020, 24, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Chai, L.; Yuan, Y.; Chen, C.; Zhou, J.; Wu, Y. The role of long non-coding RNA ANRIL in the carcinogenesis of oral cancer by targeting miR-125a. Biomed. Pharmacother. 2018, 103, 38–45. [Google Scholar] [CrossRef]
- Ge, C.; Dong, J.; Chu, Y.; Cao, S.; Zhang, J.; Wei, J. LncRNA FGD5-AS1 promotes tumor growth by regulating MCL1 via sponging miR-153-3p in oral cancer. Aging 2020, 12, 14355–14364. [Google Scholar] [CrossRef]
- Hong, Y.; He, H.; Sui, W.; Zhang, J.; Zhang, S.; Yang, D. Long non-coding RNA H19 promotes cell proliferation and invasion by acting as a ceRNA of miR-138 and releasing EZH2 in oral squamous cell carcinoma. Int. J. Oncol 2018, 52, 901–912. [Google Scholar] [CrossRef]
- Chang, S.M.; Hu, W.W. Long non-coding RNA MALAT1 promotes oral squamous cell carcinoma development via microRNA-125b/STAT3 axis. J. Cell Physiol. 2018, 233, 3384–3396. [Google Scholar] [CrossRef]
- Sun, C.C.; Zhang, L.; Li, G.; Li, S.J.; Chen, Z.L.; Fu, Y.F.; Gong, F.Y.; Bai, T.; Zhang, D.Y.; Wu, Q.M.; et al. The lncRNA PDIA3P Interacts with miR-185-5p to Modulate Oral Squamous Cell Carcinoma Progression by Targeting Cyclin D2. Mol. Ther. Nucleic Acids 2017, 9, 100–110. [Google Scholar] [CrossRef]
- Li, C.; Ye, J.; Zhang, Z.; Gong, Z.; Lin, Z.; Ding, M. Long non-coding RNA RBM5-AS1 promotes the aggressive behaviors of oral squamous cell carcinoma by regulation of miR-1285-3p/YAP1 axis. Biomed. Pharmacother. 2020, 123, 109723. [Google Scholar] [CrossRef]
- Liu, S.; Liu, L.H.; Hu, W.W.; Wang, M. Long noncoding RNA TUG1 regulates the development of oral squamous cell carcinoma through sponging miR-524-5p to mediate DLX1 expression as a competitive endogenous RNA. J. Cell Physiol. 2019, 234, 20206–20216. [Google Scholar] [CrossRef]
- Fu, S.W.; Zhang, Y.; Li, S.; Shi, Z.Y.; Zhao, J.; He, Q.L. LncRNA TTN-AS1 promotes the progression of oral squamous cell carcinoma via miR-411-3p/NFAT5 axis. Cancer Cell Int. 2020, 20, 415. [Google Scholar] [CrossRef]
- He, R.Z.; Luo, D.X.; Mo, Y.Y. Emerging roles of lncRNAs in the post-transcriptional regulation in cancer. Genes Dis. 2019, 6, 6–15. [Google Scholar] [CrossRef]
- Wu, X.; Gardashova, G.; Lan, L.; Han, S.; Zhong, C.; Marquez, R.T.; Wei, L.; Wood, S.; Roy, S.; Gowthaman, R.; et al. Targeting the interaction between RNA-binding protein HuR and FOXQ1 suppresses breast cancer invasion and metastasis. Commun. Biol. 2020, 3, 193. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tao, H. Small nucleolar RNA host gene 3 facilitates cell proliferation and migration in oral squamous cell carcinoma via targeting nuclear transcription factor Y subunit gamma. J. Cell Biochem. 2020, 121, 2150–2158. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Mo, Y.; Peng, M.; Tang, T.; Zhong, Y.; Deng, X.; Xiong, F.; Guo, C.; Wu, X.; Li, Y.; et al. Emerging role of tumor-related functional peptides encoded by lncRNA and circRNA. Mol. Cancer 2020, 19, 22. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Orera, J.; Villanueva-Cañas, J.L.; Albà, M.M. Evolution of new proteins from translated sORFs in long non-coding RNAs. Exp. Cell Res. 2020, 391, 111940. [Google Scholar] [CrossRef]
- Shao, T.; Huang, J.; Zheng, Z.; Wu, Q.; Liu, T.; Lv, X. SCCA, TSGF, and the Long Non-Coding RNA AC007271.3 are Effective Biomarkers for Diagnosing Oral Squamous Cell Carcinoma. Cell Physiol. Biochem. 2018, 47, 26–38. [Google Scholar] [CrossRef]
- Shao, T.R.; Zheng, Z.N.; Chen, Y.C.; Wu, Q.Q.; Huang, G.Z.; Li, F.; Zeng, W.S.; Lv, X.Z. LncRNA AC007271.3 promotes cell proliferation, invasion, migration and inhibits cell apoptosis of OSCC via the Wnt/β-catenin signaling pathway. Life Sci. 2019, 239, 117087. [Google Scholar] [CrossRef]
- Liu, L.; Ning, S.B.; Fu, S.; Mao, Y.; Xiao, M.; Guo, B. Effects of lncRNA ANRIL on proliferation and apoptosis of oral squamous cell carcinoma cells by regulating TGF-β/Smad pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 6194–6201. [Google Scholar]
- Zhang, D.; Ding, L.; Li, Y.; Ren, J.; Shi, G.; Wang, Y.; Zhao, S.; Ni, Y.; Hou, Y. Midkine derived from cancer-associated fibroblasts promotes cisplatin-resistance via up-regulation of the expression of lncRNA ANRIL in tumour cells. Sci Rep. 2017, 7, 16231. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Zhou, R.; Mao, L.; Deng, C.; Dang, X. Esophageal cancer related gene-4 inhibits the migration and proliferation of oral squamous cell carcinoma through BC200 lncRNA/MMP-9 and -13 signaling pathway. Cell Signal. 2019, 62, 109327. [Google Scholar] [CrossRef] [PubMed]
- Dai, D.; Feng, X.D.; Zhu, W.Q.; Bao, Y.N. LncRNA BLACAT1 regulates the viability, migration and invasion of oral squamous cell carcinoma cells by targeting miR-142-5p. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10313–10323. [Google Scholar] [PubMed]
- Yang, Y.; Chen, D.; Liu, H.; Yang, K. Increased expression of lncRNA CASC9 promotes tumor progression by suppressing autophagy-mediated cell apoptosis via the AKT/mTOR pathway in oral squamous cell carcinoma. Cell Death Dis. 2019, 10, 41. [Google Scholar] [CrossRef]
- Zhang, X.; Guo, B.; Zhu, Y.; Xu, W.; Ning, S.; Liu, L. Up-regulation of plasma lncRNA CACS15 distinguished early-stage oral squamous cell carcinoma patient. Oral Dis. 2020, 26, 1619–1624. [Google Scholar] [CrossRef]
- Arunkumar, G.; Murugan, A.K.; Prasanna Srinivasa Rao, H.; Subbiah, S.; Rajaraman, R.; Munirajan, A.K. Long non-coding RNA CCAT1 is overexpressed in oral squamous cell carcinomas and predicts poor prognosis. Biomed. Rep. 2017, 6, 455–462. [Google Scholar] [CrossRef]
- Li, G.H.; Ma, Z.H.; Wang, X. Long non-coding RNA CCAT1 is a prognostic biomarker for the progression of oral squamous cell carcinoma via miR-181a-mediated Wnt/β-catenin signaling pathway. Cell Cycle 2019, 18, 2902–2913. [Google Scholar] [CrossRef]
- Wang, Y.; Tong, J.; Lin, H.; Ma, L.; Xu, Y. CCHE1 accelerated the initiation of oral squamous cell carcinoma through enhancing PAK2 expression by sponging miR-922. J. Oral Pathol. Med. 2020, 49, 636–644. [Google Scholar] [CrossRef]
- Dai, J.; Mu, J.W.; Mu, H. Long non-coding RNA CRNDE regulates cell proliferation, migration, invasion, epithelial-mesenchymal transition and apoptosis in oral squamous cell carcinoma. Oncol. Lett. 2019, 17, 3330–3340. [Google Scholar] [CrossRef]
- Qu, X.H.; Shi, Y.L.; Ma, Y.; Bao, W.W.; Yang, L.; Li, J.C.; Zhang, F. LncRNA DANCR regulates the growth and metastasis of oral squamous cell carcinoma cells via altering miR-216a-5p expression. Hum. Cell 2020, 33, 1281–1293. [Google Scholar] [CrossRef]
- Fang, X.; Tang, Z.; Zhang, H.; Quan, H. Long non-coding RNA DNM3OS/miR-204-5p/HIP1 axis modulates oral cancer cell viability and migration. J. Oral Pathol. Med. 2020, 49, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Li, Z.; Gan, Z.; Yang, Z.; Wu, Z.; Rong, M. LncRNA ELF3-AS1 is involved in the regulation of oral squamous cell carcinoma cell proliferation by reprogramming glucose metabolism. Onco Targets Ther. 2019, 12, 6857–6863. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Jiao, Y. LncRNA FAL1 promotes the development of oral squamous cell carcinoma through regulating the microRNA-761/CRKL pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5779–5786. [Google Scholar]
- Liu, L.; Zhan, Y.; Huang, Y.; Huang, L. LncRNA FGD5-AS1 can be predicted as therapeutic target in oral cancer. J. Oral Pathol. Med. 2020, 49, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xiong, X.; Chen, L.; Yang, L.; Li, X. Identification and validation long non-coding RNAs of oral squamous cell carcinoma by bioinformatics method. Oncotarget 2017, 8, 107469–107476. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Li, Y.; Jiang, F.; Wei, C.; Chen, G.; Zhang, W.; Zhao, W.; Yu, D. LncRNA GAS5 suppresses proliferation, migration, invasion, and epithelial-mesenchymal transition in oral squamous cell carcinoma by regulating the miR-21/PTEN axis. Exp. Cell Res. 2019, 374, 365–373. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, X.; Feng, C.; Shang, X.; Xi, Y. Long Non-Coding RNA HCP5 Facilitates Cell Invasion And Epithelial-Mesenchymal Transition In Oral Squamous Cell Carcinoma By miR-140-5p/SOX4 Axis. Cancer Manag. Res. 2019, 11, 10455–10462. [Google Scholar] [CrossRef]
- Liu, Z.; Li, H.; Fan, S.; Lin, H.; Lian, W. STAT3-induced upregulation of long noncoding RNA HNF1A-AS1 promotes the progression of oral squamous cell carcinoma via activating Notch signaling pathway. Cancer Biol Ther 2019, 20, 444–453. [Google Scholar] [CrossRef]
- Xiong, L.; Tang, Y.; Tang, J.; Liu, Z.; Wang, X. Downregulation of lncRNA HOTTIP Suppresses the Proliferation, Migration, and Invasion of Oral Tongue Squamous Cell Carcinoma by Regulation of HMGA2-Mediated Wnt/β-Catenin Pathway. Cancer Biother. Radiopharm. 2020, 35, 720–730. [Google Scholar] [CrossRef]
- Mu, M.; Li, Y.; Zhan, Y.; Li, X.; Zhang, B. Knockdown of HOXA transcript at the distal tip suppresses the growth and invasion and induces apoptosis of oral tongue squamous carcinoma cells. Onco Targets Ther. 2018, 11, 8033–8044. [Google Scholar] [CrossRef]
- Li, B.; Wang, W.; Miao, S.; Li, G.; Lv, Y.; Xiang, C.; Pei, R. HOXA11-AS promotes the progression of oral squamous cell carcinoma by targeting the miR-518a-3p/PDK1 axis. Cancer Cell Int. 2019, 19, 140. [Google Scholar] [CrossRef]
- Wang, X.; Li, H.; Shi, J. LncRNA HOXA11-AS Promotes Proliferation and Cisplatin Resistance of Oral Squamous Cell Carcinoma by Suppression of miR-214-3p Expression. Biomed. Res. Int 2019, 2019, 8645153. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.; Yang, B.; Liu, F.; Fang, Q. LncRNA HOXA11-AS promotes OSCC progression by sponging miR-98-5p to upregulate YBX2 expression. Biomed. Pharmacother. 2020, 121, 109623. [Google Scholar] [CrossRef]
- Li, W.; Zhu, Q.; Zhang, S.; Liu, L.; Zhang, H.; Zhu, D. HOXC13-AS accelerates cell proliferation and migration in oral squamous cell carcinoma via miR-378g/HOXC13 axis. Oral Oncol. 2020, 111, 104946. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Tang, J.; Wang, Y.; Sun, S.; Shen, Y.; Yang, H. Long non-coding RNA highly up-regulated in liver cancer promotes epithelial-to-mesenchymal transition process in oral squamous cell carcinoma. J. Cell Mol. Med. 2019, 23, 2645–2655. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Chen, S.; Lu, N.; Yin, Y.; Liu, Z. LncRNA JPX overexpressed in oral squamous cell carcinoma drives malignancy via miR-944/CDH2 axis. Oral Dis. 2021, 27, 924–933. [Google Scholar] [CrossRef]
- Zhang, C.; Bao, C.; Zhang, X.; Lin, X.; Pan, D.; Chen, Y. Knockdown of lncRNA LEF1-AS1 inhibited the progression of oral squamous cell carcinoma (OSCC) via Hippo signaling pathway. Cancer Biol. Ther. 2019, 20, 1213–1222. [Google Scholar] [CrossRef]
- Li, M.; Ning, J.; Li, Z.; Wang, J.; Zhao, C.; Wang, L. LINC00152 promotes the growth and invasion of oral squamous cell carcinoma by regulating miR-139-5p. Onco Targets Ther. 2018, 11, 6295–6304. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, J.; Li, S.; Jia, B.; Huang, Z.; Shen, J.; Luo, H.; Zhao, J. CCL18-induced LINC00319 promotes proliferation and metastasis in oral squamous cell carcinoma via the miR-199a-5p/FZD4 axis. Cell Death Dis. 2020, 11, 777. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, X.; Dong, P.; Cai, J. Long noncoding RNA LINC00958 promotes the oral squamous cell carcinoma by sponging miR-185-5p/YWHAZ. Life Sci. 2020, 242, 116782. [Google Scholar] [CrossRef]
- Lee, S.P.; Hsieh, P.L.; Fang, C.Y.; Chu, P.M.; Liao, Y.W.; Yu, C.H.; Yu, C.C.; Tsai, L.L. LINC00963 Promotes Cancer Stemness, Metastasis, and Drug Resistance in Head and Neck Carcinomas via ABCB5 Regulation. Cancers 2020, 12, 1073. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Guo, Y.; Huang, Z.; Zhang, Q.; Huang, Z.; Zhang, Y.; Huang, Z. Vitamin D inhibits the proliferation of Oral Squamous Cell Carcinoma by suppressing lncRNA LUCAT1 through the MAPK pathway. J. Cancer 2020, 11, 5971–5981. [Google Scholar] [CrossRef]
- Zhu, M.; Zhang, C.; Chen, D.; Chen, S.; Zheng, H. lncRNA MALAT1 potentiates the progression of tongue squamous cell carcinoma through regulating miR-140-5p-PAK1 pathway. Onco. Targets Ther. 2019, 12, 1365–1377. [Google Scholar] [CrossRef]
- Xiao, L.; Wang, W.; Zhao, J.; Xu, H.; Li, S.; Yang, X. lncRNA MALAT1 promotes cell proliferation and invasion by regulating the miR-101/EZH2 axis in oral squamous cell carcinoma. Oncol. Lett. 2020, 20, 164. [Google Scholar] [CrossRef]
- Yu, L.; Shao, X.; Huo, L.; Zhang, T. Long Non-Coding RNA (lncRNA) Metastasis-Associated Lung Adenocarcinoma Transcript 1 (MALAT1) Promotes Cell Proliferation and Migration by Regulating miR-143-3p and MAGE Family Member A9 (MAGEA9) in Oral Squamous Cell Carcinoma. Med. Sci. Monit. 2020, 26, e924187. [Google Scholar] [CrossRef]
- Liang, J.; Liang, L.; Ouyang, K.; Li, Z.; Yi, X. MALAT1 induces tongue cancer cells’ EMT and inhibits apoptosis through Wnt/β-catenin signaling pathway. J. Oral Pathol. Med. 2017, 46, 98–105. [Google Scholar] [CrossRef]
- Wang, R.; Lu, X.; Yu, R. lncRNA MALAT1 Promotes EMT Process and Cisplatin Resistance of Oral Squamous Cell Carcinoma via PI3K/AKT/m-TOR Signal Pathway. Onco. Targets Ther. 2020, 13, 4049–4061. [Google Scholar] [CrossRef]
- Hou, C.; Wang, X.; Du, B. lncRNA MCM3AP-AS1 promotes the development of oral squamous cell carcinoma by inhibiting miR-363-5p. Exp. Ther. Med. 2020, 20, 978–984. [Google Scholar] [CrossRef]
- Li, H.; Jiang, J. LncRNA MCM3AP-AS1 promotes proliferation, migration and invasion of oral squamous cell carcinoma cells via regulating miR-204-5p/FOXC1. J. Investig. Med. 2020, 68, 1282–1288. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Hu, D.; Zou, L.H. Low expression of lncRNA MEG3 promotes the progression of oral squamous cell carcinoma by targeting miR-21. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 8315–8323. [Google Scholar] [PubMed]
- Tan, J.; Xiang, L.; Xu, G. LncRNA MEG3 suppresses migration and promotes apoptosis by sponging miR-548d-3p to modulate JAK-STAT pathway in oral squamous cell carcinoma. IUBMB Life 2019, 71, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Wu, C.; Xie, N.; Wang, P. Long non-coding RNA MEG3 inhibits the proliferation and metastasis of oral squamous cell carcinoma by regulating the WNT/β-catenin signaling pathway. Oncol. Lett. 2017, 14, 4053–4058. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, C.; Liu, J.; Geng, F.; Shi, X.; Li, Q.; Lu, Z.; Pan, Y. Fusobacterium nucleatum promotes epithelial-mesenchymal transiton through regulation of the lncRNA MIR4435-2HG/miR-296-5p/Akt2/SNAI1 signaling pathway. FEBS J. 2020, 287, 4032–4047. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.G.; Li, H.; Xiao, Y.; Yang, Q.C.; Yang, L.L.; Chen, L.; Bu, L.L.; Zhang, W.F.; Zhang, J.L.; Sun, Z.J. Long noncoding RNA MYOSLID promotes invasion and metastasis by modulating the partial epithelial-mesenchymal transition program in head and neck squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 278. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; He, X.; Wei, X.L. lncRNA NEAT1 promotes cell proliferation and invasion by regulating miR-365/RGS20 in oral squamous cell carcinoma. Oncol. Rep. 2018, 39, 1948–1956. [Google Scholar] [CrossRef]
- Arunkumar, G.; Anand, S.; Raksha, P.; Dhamodharan, S.; Prasanna Srinivasa Rao, H.; Subbiah, S.; Murugan, A.K.; Munirajan, A.K. LncRNA OIP5-AS1 is overexpressed in undifferentiated oral tumors and integrated analysis identifies as a downstream effector of stemness-associated transcription factors. Sci. Rep. 2018, 8, 7018. [Google Scholar] [CrossRef]
- Li, M.; Ning, J.; Li, Z.; Fei, Q.; Zhao, C.; Ge, Y.; Wang, L. Long noncoding RNA OIP5-AS1 promotes the progression of oral squamous cell carcinoma via regulating miR-338-3p/NRP1 axis. Biomed. Pharmacother. 2019, 118, 109259. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, Y.; Li, C.; Zhang, L.; Liu, Q.; Jiang, T. LncRNA PAPAS promotes oral squamous cell carcinoma by upregulating transforming growth factor-β1. J. Cell Biochem. 2019, 120, 16120–16127. [Google Scholar] [CrossRef]
- Chen, F.; Qi, S.; Zhang, X.; Wu, J.; Yang, X.; Wang, R. lncRNA PLAC2 activated by H3K27 acetylation promotes cell proliferation and invasion via the activation of Wnt/β-catenin pathway in oral squamous cell carcinoma. Int. J. Oncol. 2019, 54, 1183–1194. [Google Scholar] [CrossRef]
- Li, X.; Ren, H. Long noncoding RNA PVT1 promotes tumor cell proliferation, invasion, migration and inhibits apoptosis in oral squamous cell carcinoma by regulating miR-150-5p/GLUT-1. Oncol Rep. 2020, 44, 1524–1538. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ji, X.; Wang, J.; Ma, X.; Yang, Y.; Zuo, J.; Cui, J. LncRNA PVT1 Enhances Proliferation and Cisplatin Resistance via Regulating miR-194-5p/HIF1a Axis in Oral Squamous Cell Carcinoma. Onco Targets Ther. 2020, 13, 243–252. [Google Scholar] [CrossRef]
- Liu, G.M.; Lu, T.C.; Sun, M.L.; Ji, X.; Zhao, Y.A.; Jia, W.Y.; Luo, Y.G. RP11-874J12.4 promotes oral squamous cell carcinoma tumorigenesis via the miR-19a-5p/EBF1 axis. J. Oral Pathol. Med. 2020, 49, 645–654. [Google Scholar] [CrossRef]
- Yin, Y.; Tan, Y.; Yao, Y.; Lu, N.; Zhang, F. SNHG12/miR-326/E2F1 feedback loop facilitates the progression of oral squamous cell carcinoma. Oral Dis. 2020, 26, 1631–1639. [Google Scholar] [CrossRef] [PubMed]
- Tong, F.; Guo, J.; Miao, Z.; Li, Z. LncRNA SNHG17 promotes the progression of oral squamous cell carcinoma by modulating miR-375/PAX6 axis. Cancer Biomark. 2021, 30, 1–12. [Google Scholar] [CrossRef]
- Gao, P.; Fan, R.; Ge, T. SNHG20 serves as a predictor for prognosis and promotes cell growth in oral squamous cell carcinoma. Oncol. Lett. 2019, 17, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.F.; Wang, Y.; Sun, L.L.; Ding, S.Y.; Jinag, H. LncRNA SNHG20 enhances the progression of oral squamous cell carcinoma by regulating the miR-29a/DIXDC1/Wnt regulatory axis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 5436–5445. [Google Scholar] [PubMed]
- Lu, N.; Yin, Y.; Yao, Y.; Zhang, P. SNHG3/miR-2682-5p/HOXB8 promotes cell proliferation and migration in oral squamous cell carcinoma. Oral Dis. 2020. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-M.; Wang, T.-H.; Chen, H.-C.; Li, S.-C.; Lee, M.-C.; Liou, H.-H.; Liu, P.-F.; Tseng, Y.-K.; Shiue, Y.-L.; Ger, L.-P.; et al. Aberrant DNA hypermethylation-silenced SOX21-AS1 gene expression and its clinical importance in oral cancer. Clin. Epigenetics 2016, 8, 129. [Google Scholar] [CrossRef]
- Jin, N.; Jin, N.; Bu, W.; Li, X.; Liu, L.; Wang, Z.; Tong, J.; Li, D. Long non-coding RNA TIRY promotes tumor metastasis by enhancing epithelial-to-mesenchymal transition in oral cancer. Exp. Biol Med. 2020, 245, 585–596. [Google Scholar] [CrossRef]
- Liang, S.; Zhang, S.; Wang, P.; Yang, C.; Shang, C.; Yang, J.; Wang, J. LncRNA, TUG1 regulates the oral squamous cell carcinoma progression possibly via interacting with Wnt/β-catenin signaling. Gene 2017, 608, 49–57. [Google Scholar] [CrossRef]
- Fang, Z.; Zhao, J.; Xie, W.; Sun, Q.; Wang, H.; Qiao, B. LncRNA UCA1 promotes proliferation and cisplatin resistance of oral squamous cell carcinoma by sunppressing miR-184 expression. Cancer Med. 2017, 6, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Xu, M.; Wu, M.; Zhang, X.; Gan, M.; Jiang, H. Long noncoding RNA UCA1 promotes cell growth, migration, and invasion by targeting miR-143-3p in oral squamous cell carcinoma. Cancer Med. 2020, 9, 3115–3129. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, L.; Wu, K.; Ge, W.; Zhang, Z.; Gong, L.; Yuan, D. Knockdown of long noncoding RNA urothelial cancer-associated 1 enhances cisplatin chemosensitivity in tongue squamous cell carcinoma cells. Pharmazie 2016, 71, 598–602. [Google Scholar] [PubMed]
- Yang, Y.T.; Wang, Y.F.; Lai, J.Y.; Shen, S.Y.; Wang, F.; Kong, J.; Zhang, W.; Yang, H.Y. Long non-coding RNA UCA1 contributes to the progression of oral squamous cell carcinoma by regulating the WNT/β-catenin signaling pathway. Cancer Sci. 2016, 107, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Noguti, J.; De Moura, C.F.G.; Hossaka, T.A.; Franco, M.; Oshima, C.T.F.; Dedivitis, R.A.; Ribeiro, D.A. The Role of Canonical WNT Signaling Pathway in Oral Carcinogenesis: A Comprehensive Review. Anticancer Res. 2012, 32, 873–878. [Google Scholar] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef]
- Lyu, X.; Li, J.; Yun, X.; Huang, R.; Deng, X.; Wang, Y.; Chen, Y.; Xiao, G. miR-181a-5p, an inducer of Wnt-signaling, facilitates cell proliferation in acute lymphoblastic leukemia. Oncol Rep. 2017, 37, 1469–1476. [Google Scholar] [CrossRef]
- Custódio, M.; Biddle, A.; Tavassoli, M. Portrait of a CAF: The story of cancer-associated fibroblasts in head and neck cancer. Oral Oncol. 2020, 110, 104972. [Google Scholar] [CrossRef]
- Ding, Y.; Xi, Y.; Chen, T.; Wang, J.-y.; Tao, D.-l.; Wu, Z.-L.; Li, Y.-p.; Li, C.; Zeng, R.; Li, L. Caprin-2 enhances canonical Wnt signaling through regulating LRP5/6 phosphorylation. J. Cell Biol. 2008, 182, 865–872. [Google Scholar] [CrossRef]
- Harsha, C.; Banik, K.; Ang, H.L.; Girisa, S.; Vikkurthi, R.; Parama, D.; Rana, V.; Shabnam, B.; Khatoon, E.; Kumar, A.P.; et al. Targeting AKT/mTOR in Oral Cancer: Mechanisms and Advances in Clinical Trials. Int. J. Mol. Sci 2020, 21, 3285. [Google Scholar] [CrossRef]
- Peng, W.X.; Koirala, P.; Mo, Y.Y. LncRNA-mediated regulation of cell signaling in cancer. Oncogene 2017, 36, 5661–5667. [Google Scholar] [CrossRef] [PubMed]
- Alzahrani, A. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Seminars Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Invest. 2015, 125, 25–32. [Google Scholar] [CrossRef]
- Siegel, P.M.; Massagué, J. Cytostatic and apoptotic actions of TGF-beta in homeostasis and cancer. Nat. Rev. Cancer 2003, 3, 807–821. [Google Scholar] [CrossRef]
- Papoutsoglou, P.; Moustakas, A. Long non-coding RNAs and TGF-β signaling in cancer. Cancer Sci. 2020, 111, 2672–2681. [Google Scholar] [CrossRef] [PubMed]
- FDA-NIH Biomarker Working Group. BEST (Biomarkers, EndpointS, and other Tools) Resource; Food and Drug Administration (US), National Institutes of Health (US): Bethesda, MD, USA, 2016. [Google Scholar]
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366. [Google Scholar]
- Mazumder, S.; Datta, S.; Ray, J.G.; Chaudhuri, K.; Chatterjee, R. Liquid biopsy: miRNA as a potential biomarker in oral cancer. Cancer Epidemiol. 2019, 58, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Simonetti, M.; Rossell, C.; Mignardi, M.; Mirzazadeh, R.; Annaratone, L.; Marchiò, C.; Sapino, A.; Bienko, M.; Crosetto, N.; et al. RollFISH achieves robust quantification of single-molecule RNA biomarkers in paraffin-embedded tumor tissue samples. Commun. Biol. 2018, 1, 209. [Google Scholar] [CrossRef]
- Crosetto, N.; Bienko, M.; van Oudenaarden, A. Spatially resolved transcriptomics and beyond. Nat. Rev. Genet. 2015, 16, 57–66. [Google Scholar] [CrossRef]
- Mathai, R.; Vidya, R.; Reddy, B.; Thomas, L.; Udupa, K.; Kolesar, J.; Rao, M. Potential Utility of Liquid Biopsy as a Diagnostic and Prognostic Tool for the Assessment of Solid Tumors: Implications in the Precision Oncology. J. Clin. Med. 2019, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- De Rubis, G.; Rajeev Krishnan, S.; Bebawy, M. Liquid Biopsies in Cancer Diagnosis, Monitoring, and Prognosis. Trends Pharmacol. Sci. 2019, 40, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Arantes, L.; De Carvalho, A.C.; Melendez, M.E.; Lopes Carvalho, A. Serum, plasma and saliva biomarkers for head and neck cancer. Expert Rev. Mol. Diagn. 2018, 18, 85–112. [Google Scholar] [CrossRef] [PubMed]
- Lousada-Fernandez, F.; Rapado-Gonzalez, O.; Lopez-Cedrun, J.L.; Lopez-Lopez, R.; Muinelo-Romay, L.; Suarez-Cunqueiro, M.M. Liquid Biopsy in Oral Cancer. Int. J. Mol. Sci. 2018, 19, 1704. [Google Scholar] [CrossRef]
- Wu, J.; Hu, S.; Zhang, L.; Xin, J.; Sun, C.; Wang, L.; Ding, K.; Wang, B. Tumor circulome in the liquid biopsies for cancer diagnosis and prognosis. Theranostics 2020, 10, 4544–4556. [Google Scholar] [CrossRef]
- De Rubis, G.; Krishnan, S.R.; Bebawy, M. Circulating tumor DNA - Current state of play and future perspectives. Pharmacol. Res. 2018, 136, 35–44. [Google Scholar] [CrossRef]
- Fan, C.; Wang, J.; Tang, Y.; Zhang, S.; Xiong, F.; Guo, C.; Zhou, Y.; Li, Z.; Li, X.; Li, Y.; et al. Upregulation of long non-coding RNA LOC284454 may serve as a new serum diagnostic biomarker for head and neck cancers. BMC Cancer 2020, 20, 917. [Google Scholar] [CrossRef] [PubMed]
- Cristaldi, M.; Mauceri, R.; Di Fede, O.; Giuliana, G.; Campisi, G.; Panzarella, V. Salivary Biomarkers for Oral Squamous Cell Carcinoma Diagnosis and Follow-Up: Current Status and Perspectives. Front. Physiol. 2019, 10, 1476. [Google Scholar] [CrossRef]
- Tang, H.; Wu, Z.; Zhang, J.; Su, B. Salivary lncRNA as a potential marker for oral squamous cell carcinoma diagnosis. Mol. Med. Rep. 2013, 7, 761–766. [Google Scholar] [CrossRef]
- Blondal, T.; Jensby Nielsen, S.; Baker, A.; Andreasen, D.; Mouritzen, P.; Wrang Teilum, M.; Dahlsveen, I.K. Assessing sample and miRNA profile quality in serum and plasma or other biofluids. Methods 2013, 59, S1–S6. [Google Scholar] [CrossRef]
- Livak, K.J.; Wills, Q.F.; Tipping, A.J.; Datta, K.; Mittal, R.; Goldson, A.J.; Sexton, D.W.; Holmes, C.C. Methods for qPCR gene expression profiling applied to 1440 lymphoblastoid single cells. Methods 2013, 59, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, S.; Herold, A.; Bergauer, T.; Belousov, A.; Betzl, G.; Demario, M.; Dietrich, M.; Luistro, L.; Poignee-Heger, M.; Schostack, K.; et al. Gene expression analysis in biomarker research and early drug development using function tested reverse transcription quantitative real-time PCR assays. Methods 2013, 59, 10–19. [Google Scholar] [CrossRef]
- Supper, J.; Gugenmus, C.; Wollnik, J.; Drueke, T.; Scherf, M.; Hahn, A.; Grote, K.; Bretschneider, N.; Klocke, B.; Zinser, C.; et al. Detecting and visualizing gene fusions. Methods 2013, 59, S24–S28. [Google Scholar] [CrossRef]
- Hiraga, C.; Yamamoto, S.; Hashimoto, S.; Kasahara, M.; Minamisawa, T.; Matsumura, S.; Katakura, A.; Yajima, Y.; Nomura, T.; Shiba, K. Pentapartite fractionation of particles in oral fluids by differential centrifugation. Sci. Rep. 2021, 11, 3326. [Google Scholar] [CrossRef]
- Minotti, L.; Agnoletto, C.; Baldassari, F.; Corrà, F.; Volinia, S. SNPs and Somatic Mutation on Long Non-Coding RNA: New Frontier in the Cancer Studies? High Throughput 2018, 7, 34. [Google Scholar] [CrossRef]
- Yuan, Z.; Yu, Y.; Zhang, B.; Miao, L.; Wang, L.; Zhao, K.; Ji, Y.; Wang, R.; Ma, H.; Chen, N.; et al. Genetic variants in lncRNA H19 are associated with the risk of oral squamous cell carcinoma in a Chinese population. Oncotarget 2018, 9, 23915–23922. [Google Scholar] [CrossRef]
- Yu, A.-M.; Jian, C.; Yu, A.H.; Tu, M.-J. RNA therapy: Are we using the right molecules? Pharmacol. Ther. 2019, 196, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Li, D.; Leng, S.; Zhu, X. RNA-based pharmacotherapy for tumors: From bench to clinic and back. Biomed. Pharmacother. 2020, 125, 109997. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.G. Management of familial hypercholesterolemia: A review of the recommendations from the National Lipid Association Expert Panel on Familial Hypercholesterolemia. J. Manag. Care Pharm. 2013, 19, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Wood, H. FDA approves patisiran to treat hereditary transthyretin amyloidosis. Nat. Rev. Neurol. 2018, 14, 570. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| lncRNA | Expression Levels in Cancer | Clinical Association | Functional Regulation | Interactor | Target/Effect | Mechanistic Classification | Refs |
|---|---|---|---|---|---|---|---|
| AC007271.3* | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | N.D. | β-catenin and its downstream target molecules CyclinD1, c-myc and Bcl-2 | Unclear mechanism | [69,70] |
| ANRIL* | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis ↓ Cisplatin cytotoxicity | miR-125a | Estrogen Related Receptor α (ESRRA) | Sequestration of miRNAs (ANRIL acts as a miR-125a sponge, thereby enhancing ESRRA expression) | [56] |
| N.D. | TGF-β/Smad signaling pathway | Unclear mechanism | [71] | ||||
| N.D. | Drug transporters (MRP1 and ABCC2) | Unclear mechanism | [72] | ||||
| BC200 | Upregulated | N.D. | ↑ Proliferation ↑ Migration | N.D. | MMP-9 and MMP-13 expression | Unclear mechanism | [73] |
| BLACAT1 | Upregulated | N.D. | ↑ Viability ↑ Migration ↑ Invasion | miR-142-5p | N.D. | Sequestration of miRNAs (BLACAT1 acts as a miR-142-5p sponge) | [74] |
| C5orf66-AS1 | Downregulated | N.D. | ↓ Cell growth ↓ Metastasis | N.D. | CYC1 expression | Unclear mechanism | [24] |
| CASC2 | Downregulated |
| ↓ Migration ↓ Invasion ↓ Proliferation ↑ Apoptosis | N.D. | Downregulation of CDK1 | Unclear mechanism | [25] |
| miRNA-21 | PDCD4 expression | Sequestration of miRNAs (CASC2 acts as a miRNA-21 sponge, thereby enhancing PDCD4 expression) | [27] | ||||
| CASC9* | Upregulated |
| ↑ Proliferation ↓ Autophagy-mediated cell apoptosis | N.D. | AKT/mTOR pathway | Unclear mechanism | [75] |
| CASC15 | Upregulated |
| ↑ Proliferation | N.D. | MEG3 | Unclear mechanism | [76] |
| CCAT1* | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-181a | Wnt/β-catenin signaling | Sequestration of miRNAs (CCAT1 acts as a miRNA sponge) | [77,78] |
| miR155-5p and let7b-5p | N.D. | ||||||
| CCHE1 | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-922 | PAK2 expression | Sequestration of miRNAs (CCHE1 acts as a miR-922sponge, thereby enhancing PAK2 expression) | [79] |
| CEBPA-AS1* | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | CEBPA | Bcl-2 expression | Post-transcriptional regulation (CEBPA-AS1 might form a “lncRNA-mRNA” pair with CEBPA and regulate CEBPA expression in a cis manner) | [48] |
| CRNDE | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis | N.D. | Wnt/β-catenin signaling | Unclear mechanism | [80] |
| DANCR | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-216a-5p | Bcl-2 expression KLF12 expression | Sequestration of miRNAs (DANCR acts as a miR-216a-5p sponge, thereby enhancing Bcl-2 and KLF12 expression) | [81] |
| DNM3OS | Upregulated | N.D. | ↑ Viability ↑ Migration | miR-204-5p | HIP1 expression | Sequestration of miRNAs (DNM3OS acts as a miR-204-5p sponge, thereby enhancing HIP1 expression) | [82] |
| ELF3-AS1 | Upregulated | N.D. | ↑ Proliferation | N.D. | GLUT1 expression | Unclear mechanism | [83] |
| ENST00000470447.1 | Downregulated |
| ↓ Proliferation ↓ Migration ↓ Invasion ↑ Apoptosis | N.D. | N.D. | Unclear mechanism | [28] |
| FAL1 | Upregulated | N.D. | ↑ Proliferation | miR-761 | CRKL expression | Sequestration of miRNAs (FAL1 acts as a miR-761 sponge, thereby enhancing CRKL expression) | [84] |
| FALEC * | Downregulated | N.D. | ↓ Proliferation ↓ Migration | EZH2 | ECM1 expression | Epigenetic and transcriptional regulation (FALEC inhibited transcription through recruiting EZH2 to the promoter of ECM1) | [29] |
| FGD5-AS1 * | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-153-3p | MCL1 expression | Sequestration of miRNAs (FGD5-AS1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [57,85] |
| miR-520b | USP21 expression | ||||||
| GAS5 * | Downregulated | N.D. | ↓ Proliferation ↓ Migration ↓ Invasion ↓ EMT | miR-21 | Regulation of PI3K/Akt pathwayPTEN expression | Sequestration of miRNAs (GAS5 acts as a miR-21 sponge, thereby enhancing PTEN expression) | [86,87] |
| H19 * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis | miR-138 | EZH2 expression | Sequestration of miRNAs (H19 acts as a miR-138 sponge, thereby enhancing EZH2 expression) | [58] |
| HAS2-AS1 * | Upregulated |
| ↑ Invasion ↑ EMT | HAS2 gene | Transcription of HAS2 | Transcriptional regulation (HAS2-AS1 is necessary for the transcription of its sense counterpart HAS2 upon hypoxia treatment) | [41] |
| HCP5 | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT | miR-140-5p | SOX4 expression | Sequestration of miRNAs (HCP5 acts as a miR-140-5p sponge, thereby enhancing SOX4 expression) | [88] |
| HIFCAR/ MIR31HG * | Upregulated |
| ↑ Tumor progression ↑ Metastatic potential | HIF-1α | HIF-1α signaling | Transcriptional regulation (LncHIFCAR acts as HIF-1α coactivator) | [43] |
| p21 | Unclear mechanism | [44] | |||||
| HNF1A-AS1 * | Upregulated |
| ↑ Migration ↑ Invasion ↑ EMT | N.D. | Notch1 and Hes1 expression | Unclear mechanism | [89] |
| HOTAIR * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis ↑ Autophagy | EZH2 and H3K27me3 | Regulation of E-cadherin | Epigenetic and transcriptional regulation (HOTAIR regulated E-cadherin expression through partly associating with EZH2 and mediating H3K27me3 at the promoter of E-cadherin) | [32] |
| miR-326 | MTA2 expression | Sequestration of miRNAs (HOTAIR acts as a miR-326 sponge, thereby enhancing MTA2 expression) | [37] | ||||
| N.D. | Regulation of mTOR and the autophagy-related factors | Unclear mechanism | [36] | ||||
| HOTTIP | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis | miR-124-3p | HMGA2 expressionWnt/β-catenin pathway | Sequestration of miRNAs (HOTTIP acts as a miR-124-3p sponge, thereby enhancing HMGA2 expression) | [90] |
| N.D. | Cell cycle arrest at G1 phase | Unclear mechanism | [91] | ||||
| HOXA11-AS | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis ↓ CDDP cytotoxicity | miR-98-5p | YBX2 expression | Sequestration of miRNAs (HOXA11-AS acts as a miRNA sponge, thereby enhancing the expression of their targets) | [92,93,94] |
| miR-518a-3p | PDK1 expression | ||||||
| miR-214-3p | PIM1 expression | ||||||
| HOXC13-AS | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ EMT | miR-378g | HOXC13 expression | Sequestration of miRNAs (HOXC13-AS acts as a miR-378g sponge, thereby enhancing HOXC13 expression) | [95] |
| HULC | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis ↑ EMT ↑ CDDP tolerance | N.D. | N.D. | Unclear mechanism | [96] |
| JPX | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion | miR-944 | CDH2 expression | Sequestration of miRNAs (JPX acts as a miR-944sponge, thereby enhancing CDH2 expression) | [97] |
| LEF1-AS1* | Upregulated |
| ↑ Survival and proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | LATS1 | Regulation of Hippo signaling | Interaction Decoy (LEF1-AS1 can interact with LATS1 and therefore regulates Hippo signaling) | [98] |
| LINC00152 | Upregulated |
| ↑ Proliferation ↑ Colony formation ↑ Migration ↑ Invasion ↑ EMT | miR-139-5p | N.C. | Sequestration of miRNAs (LINC00152 acts as a miR-139-5p sponge) | [99] |
| LINC00319 | Upregulated | N.D. | ↑ Proliferation ↑ Metastasis ↑ EMT ↑ Angiogenesis | miR-199a-5p | FZD4 expression | Sequestration of miRNAs (LINC00319 acts as a miR-199a-5p sponge, thereby enhancing FZD4 expression) | [100] |
| LINC00941 * | Upregulated | N.D. | ↑ Proliferation ↑ Colony formation | CAPRIN2 | CAPRIN2 expression Canonical WNT/β-catenin signaling | Transcriptional regulation (LINC00941 acts as transcriptional activator by looping to CAPRIN2 promoter) | [39] |
| LINC00958 | Upregulated |
| ↑ Proliferation ↑ Invasion ↓ Apoptosis | miR-185-5p | YWHAZ expression | Sequestration of miRNAs (LINC00958 acts as a miR-185-5p sponge, thereby enhancing YWHAZ expression) | [101] |
| LINC00963 | Upregulated | N.D. | ↑ CSC hallmarks ↑ Chemoresistance | N.D. | Stemness marker ALDH1Multidrug-resistance transporter ABCB5 | Unclear mechanism | [102] |
| LINC01315 * | Downregulated | N.D. | ↓ Proliferation ↓ Migration ↓ Invasion ↑ Apoptosis | miR-211 | DLG3 expression Regulation of Hippo signaling pathway | Sequestration of miRNAs (LINC01315 acts as a miR-211 sponge, thereby enhancing DLG3 expression) | [30] |
| LncRNA-p23154 * | Upregulated | N.D. | ↑ Metastasis ↑ Glycolysis | Promoter region of miR-378a-3p | GLUT1 expression | Transcriptional regulation (LncRNA-p23154 binds to the promoter region of miR-378a-3p) | [40] |
| LUCAT1 | Upregulated |
| ↑ Cell growth ↑ Proliferation | N.D. | MAPK signaling | Unclear mechanism | [103] |
| MALAT1 * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT ↓ Apoptosis ↓ DDP-resistance | miR-143-3p | MAGEA9 expression | Sequestration of miRNAs (MALAT1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [59,104,105,106] |
| miR-140-5p | PAK1 expression | ||||||
| miR-125b | STAT3 expression | ||||||
| miR-101 | EZH2 expression | ||||||
| N.D. | P-glycoprotein expression PI3K/AKT/m-TOR signaling | Unclear mechanism | [107,108] | ||||
| N.D. | Wnt/β-catenin signaling | ||||||
| MCM3AP-AS1 | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion | miR-204-5p | FOXC1 expression | Sequestration of miRNAs (MCM3AP-AS1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [109,110] |
| miR-363-5p | N.D. | ||||||
| MEG3 * | Downregulated | N.D. | ↓ Proliferation ↓ Migration ↓ Invasion ↑ Apoptosis | miR-548d-3p | SOCS5 and SOCS6 expression Regulation of JAK-STAT signaling | Sequestration of miRNAs (MEG3 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [55,111,112] |
| miR-21 | N.D. | ||||||
| miR-361-5p | Regulation of succinate dehydrogenase (SDH) | ||||||
| N.D. | Wnt/β-catenin signaling | Unclear mechanism | [113] | ||||
| MIR4435-2HG | Upregulated | N.D. | ↑ EMT | miR-296-5p | Expression of EMT markers | Sequestration of miRNAs (MIR4435-2HG acts as a miR-296-5p sponge) | [114] |
| MORT | Downregulated |
| ↓ Proliferation | N.D. | ROCK1 expression | Unclear mechanism | [31] |
| MYOSLID | Upregulated |
| ↑ Migration ↑ Invasion | N.D. | Expression of EMT-related markers | Unclear mechanism | [115] |
| NEAT1 | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-365 | RGS20 expression | Sequestration of miRNAs (NEAT1 acts as a miR-365 sponge, thereby enhancing RGS20 expression) | [116] |
| OIP5-AS1 | Upregulated |
| ↑ Proliferation ↑ Migration↑ Invasion | miR-338-3p | NRP1 expression | Sequestration of miRNAs (OIP5-AS1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [117,118] |
| miR-137 miR148a-3p miR-30a-5p miR-30b-5p miR-338-3p miR-22-3p | N.D. | ||||||
| PAPAS * | Upregulated |
| ↑ Migration ↑ Invasion | N.D. | TGF-β signaling | Unclear mechanism | [119] |
| PLAC2 * | Upregulated | N.D. | ↑ Proliferation ↑ Invasion | N.D. | Wnt/β-catenin signaling | Unclear mechanism | [120] |
| PDIA3P * | Upregulated |
| ↑ Proliferation | miR-185-5p | CCND2 expression | Sequestration of miRNAs (PDIA3P acts as a miR-185-5p sponge, thereby enhancing CCND2 expression) | [60] |
| PVT1 | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ Cisplatin-resistance ↓ Apoptosis | miR-150-5p | GLUT-1 expression | Sequestration of miRNAs (PVT1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [121,122] |
| miR-194-5p | HIF-1α expression | ||||||
| RBM5-AS1 * | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion | miR-1285-3p | YAP1 expression | Sequestration of miRNAs (RBM5-AS1 acts as a miR-1285-3p sponge, thereby enhancing YAP1 expression) | [61] |
| RC3H2 * | Upregulated | N.D. | ↑ Cell growth ↑ Colony formation ↑ Migration ↑ Invasion | miR-101-3p | EZH2 expression | Sequestration of miRNAs (RC3H2 acts as a miR-101-3p sponge, thereby enhancing EZH2 expression) | [53] |
| RP11-874J12.4 | Upregulated | N.D. | ↑ Proliferation ↑ Migration | miR-19a-5p | EBF1 expression | Sequestration of miRNAs (RP11-874J12.4 acts as a miR-19a-5p sponge, thereby enhancing EBF1 expression) | [123] |
| SNHG12 | Upregulated | N.D. | ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-326 | E2F1 expression | Sequestration of miRNAs (SNHG12 acts as a miR-326 sponge, thereby enhancing E2F1 expression) | [124] |
| SNHG17 | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↓ Apoptosis | miR-375 | PAX6 expression | Sequestration of miRNAs (SNHG17 acts as a miR-375 sponge, thereby enhancing PAX6 expression) | [125] |
| SNHG20 * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ CSC properties | miR-29a | DIXDC1 expression Regulation of Wnt signaling | Sequestration of miRNAs (SNHG20 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [54,126,127] |
| miR-197 | LIN28 expression | ||||||
| SNHG3 * | Upregulated | N.D. | ↑ Proliferation ↑ Migration | ELAVL1 | NFYC expression Wnt/β-catenin signaling | Protein scaffold (SNHG3 can bind to ELAVL1 and therefore stabilize and upregulate NFYC expression) | [66] |
| miR-2682-5p | HOXB8 expression | Sequestration of miRNAs (SNHG3 acts as a miR-2682-5p sponge, thereby enhancing HOXB8 expression) | [128] | ||||
| SOX21-AS1 | Downregulated |
| ↓ Cell growth ↓ Invasion | N.D. | N.D. | Unclear mechanism | [129] |
| TIRY * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ EMT | miR-14 | Expression of EMT markers Wnt/β-catenin signaling | Unclear mechanism | [130] |
| TTN-AS1 * | Upregulated | ↑ Proliferation ↑ Migration ↓ Apoptosis | miR-411-3p | NFAT5 expression | Sequestration of miRNAs (TTN-AS1 acts as a miR-411-3p sponge, thereby enhancing NFAT5 expression) | [63] | |
| TUG1 * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion | miR-524-5p | DLX1 expression | Sequestration of miRNAs (TUG1 acts as a miR-524-5p sponge, thereby enhancing DLX1 expression) | [62] |
| N.D. | Wnt/β-catenin signaling | Unclear mechanism | [131] | ||||
| UCA1 * | Upregulated |
| ↑ Proliferation ↑ Migration ↑ Invasion ↑ Chemoresistance↓ Apoptosis | miR-143-3p | MYO6 expression | Sequestration of miRNAs (UCA1 acts as a miRNA sponge, thereby enhancing the expression of their targets) | [132,133] |
| miR-184 | SF1 expression | ||||||
| N.D. | Regulation of cisplatin-activated PI3K/Akt signaling | Unclear mechanism | [134,135] | ||||
| N.D. | Wnt/β-catenin signaling |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, C.-S.; Kung, H.-J.; Shih, J.-W. Long Non-Coding RNAs as Functional Codes for Oral Cancer: Translational Potential, Progress and Promises. Int. J. Mol. Sci. 2021, 22, 4903. https://doi.org/10.3390/ijms22094903
Lei C-S, Kung H-J, Shih J-W. Long Non-Coding RNAs as Functional Codes for Oral Cancer: Translational Potential, Progress and Promises. International Journal of Molecular Sciences. 2021; 22(9):4903. https://doi.org/10.3390/ijms22094903
Chicago/Turabian StyleLei, Cing-Syuan, Hsing-Jien Kung, and Jing-Wen Shih. 2021. "Long Non-Coding RNAs as Functional Codes for Oral Cancer: Translational Potential, Progress and Promises" International Journal of Molecular Sciences 22, no. 9: 4903. https://doi.org/10.3390/ijms22094903
APA StyleLei, C.-S., Kung, H.-J., & Shih, J.-W. (2021). Long Non-Coding RNAs as Functional Codes for Oral Cancer: Translational Potential, Progress and Promises. International Journal of Molecular Sciences, 22(9), 4903. https://doi.org/10.3390/ijms22094903