
Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology

 ,
,  ,
,
Abstract
1. Introduction
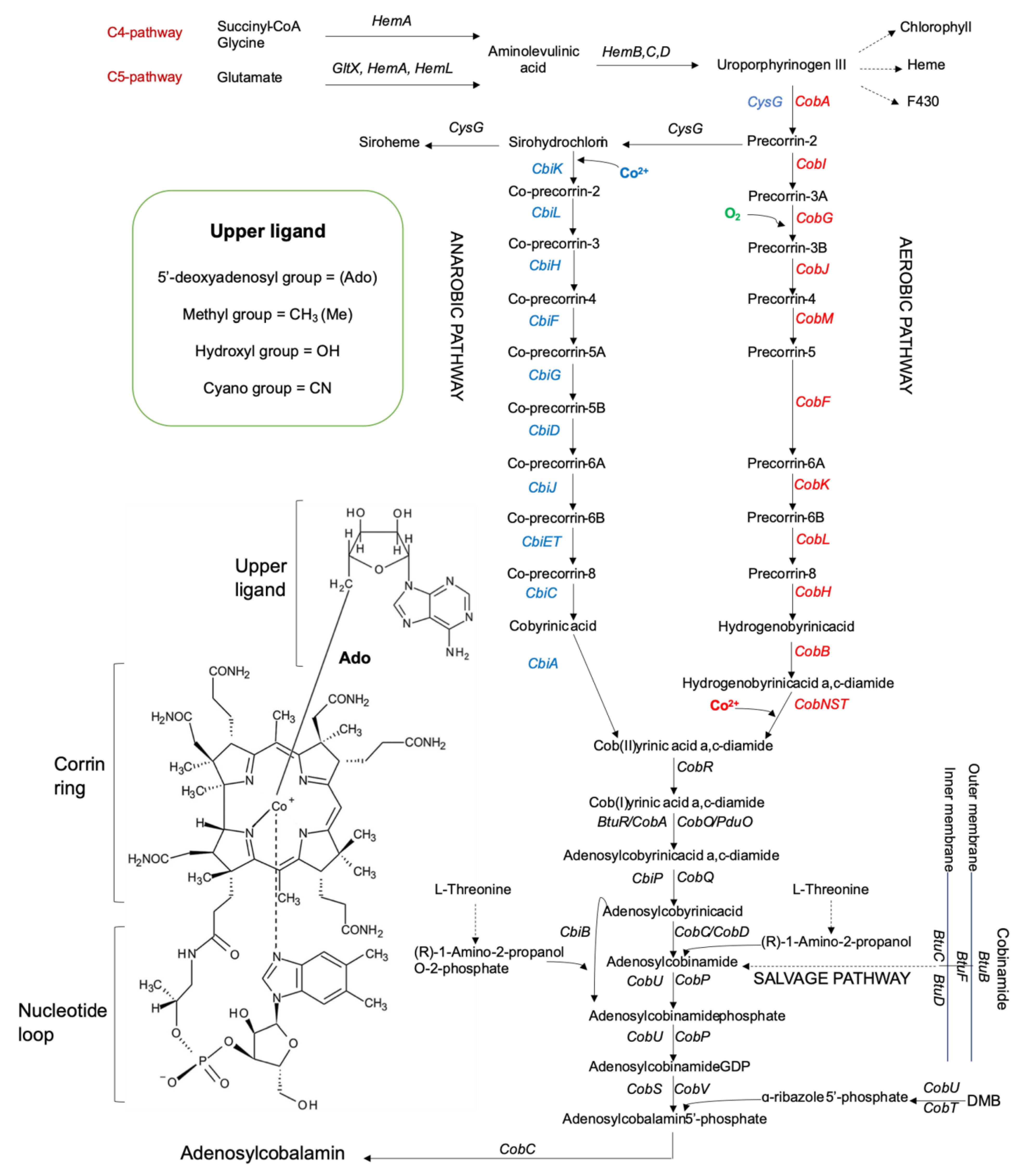
2. An Overview of Cobalamin Biosynthesis: Metabolic Pathways and Catalysts
2.1. Molecular-Genetic Organization of Cobalamin Biosynthetic Pathways
2.1.1. Historical Implication of Cobalamin Biosynthesis Signatures
2.1.2. Aerobic Pathway and Related Enzymes
2.1.3. Anaerobic Pathway and Related Enzymes
2.1.4. Common Pathway and Salvage
2.1.5. Selectivity of Lower Ligand Activation
3. Genetic Diversity of Cobalamin (Cobamide) Biosynthesis Pathways and Transport
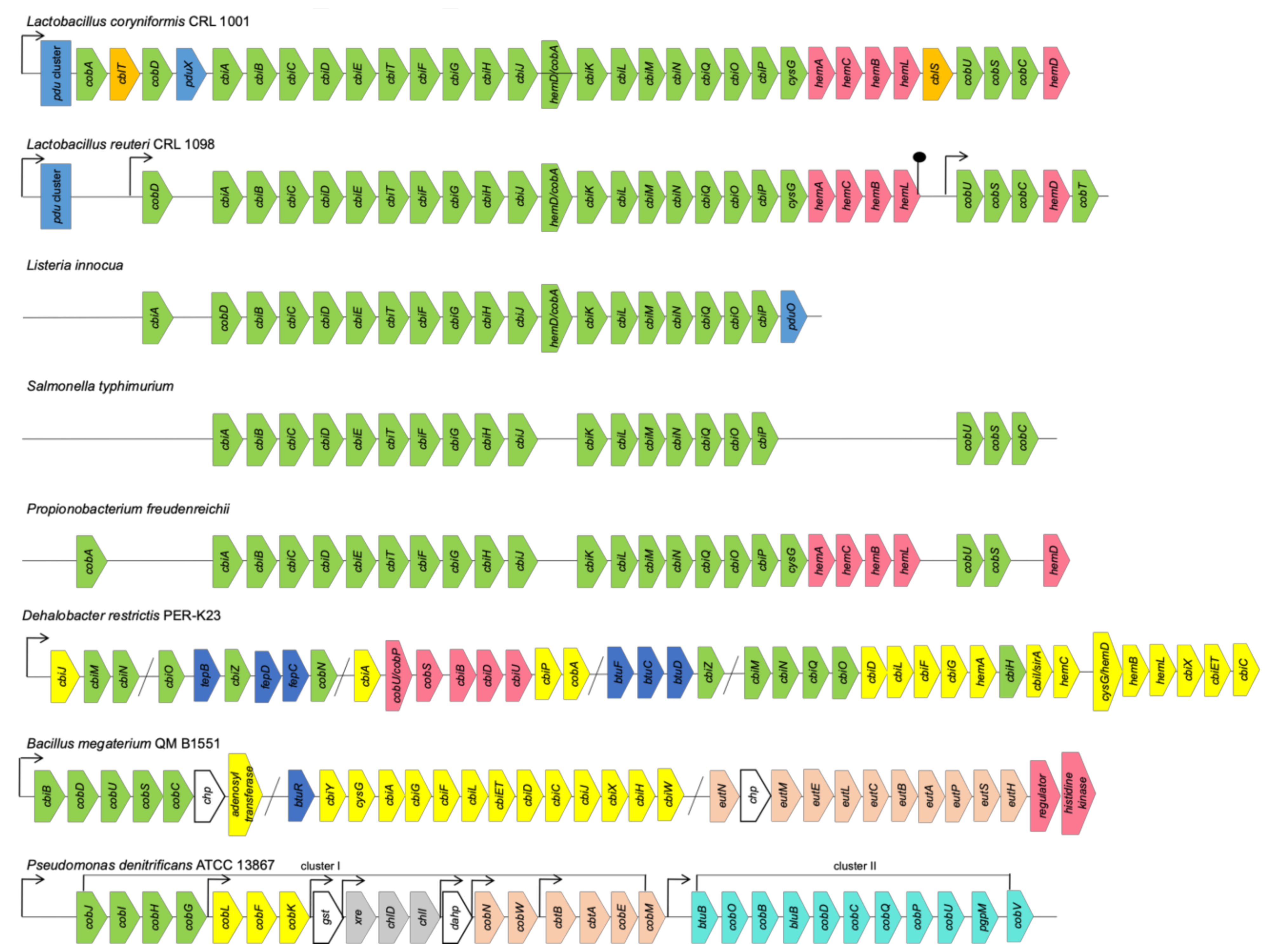
3.1. Chromosomal Organization of Cobalamin Biosynthesis Pathways
3.1.1. Clusters and Operons in Anaerobic Pathway
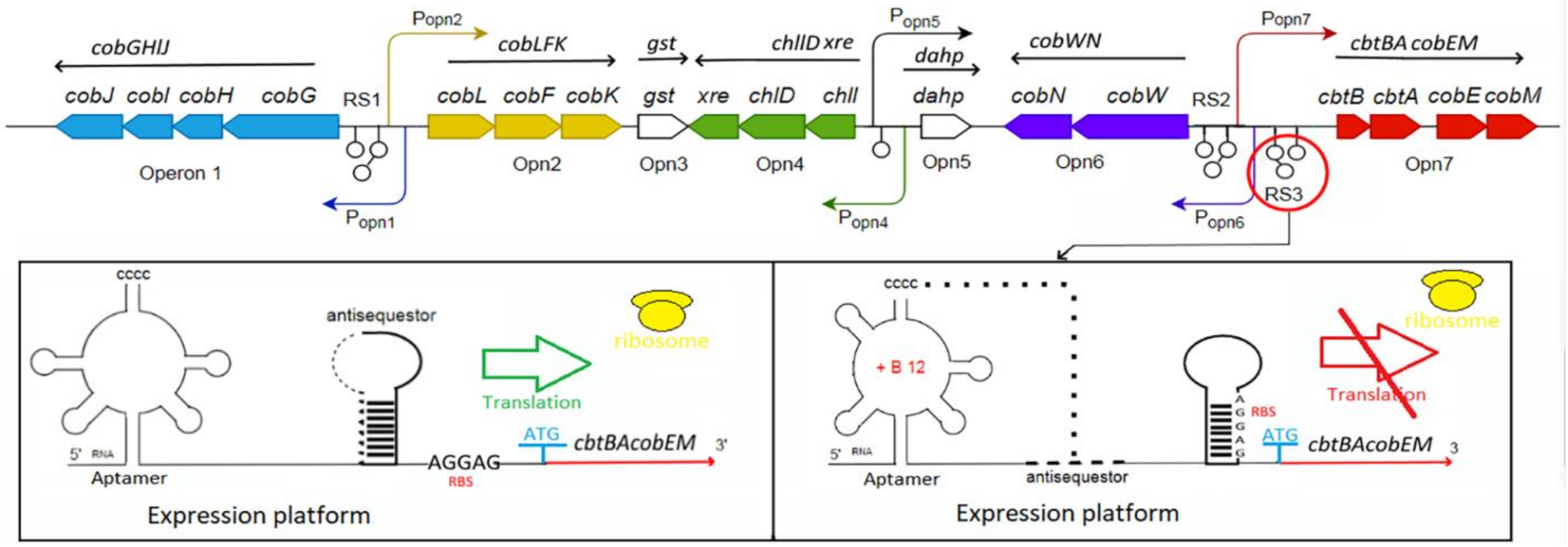
3.1.2. Clusters and Operons in Aerobic Pathway
3.1.3. Transport Systems in Bacteria
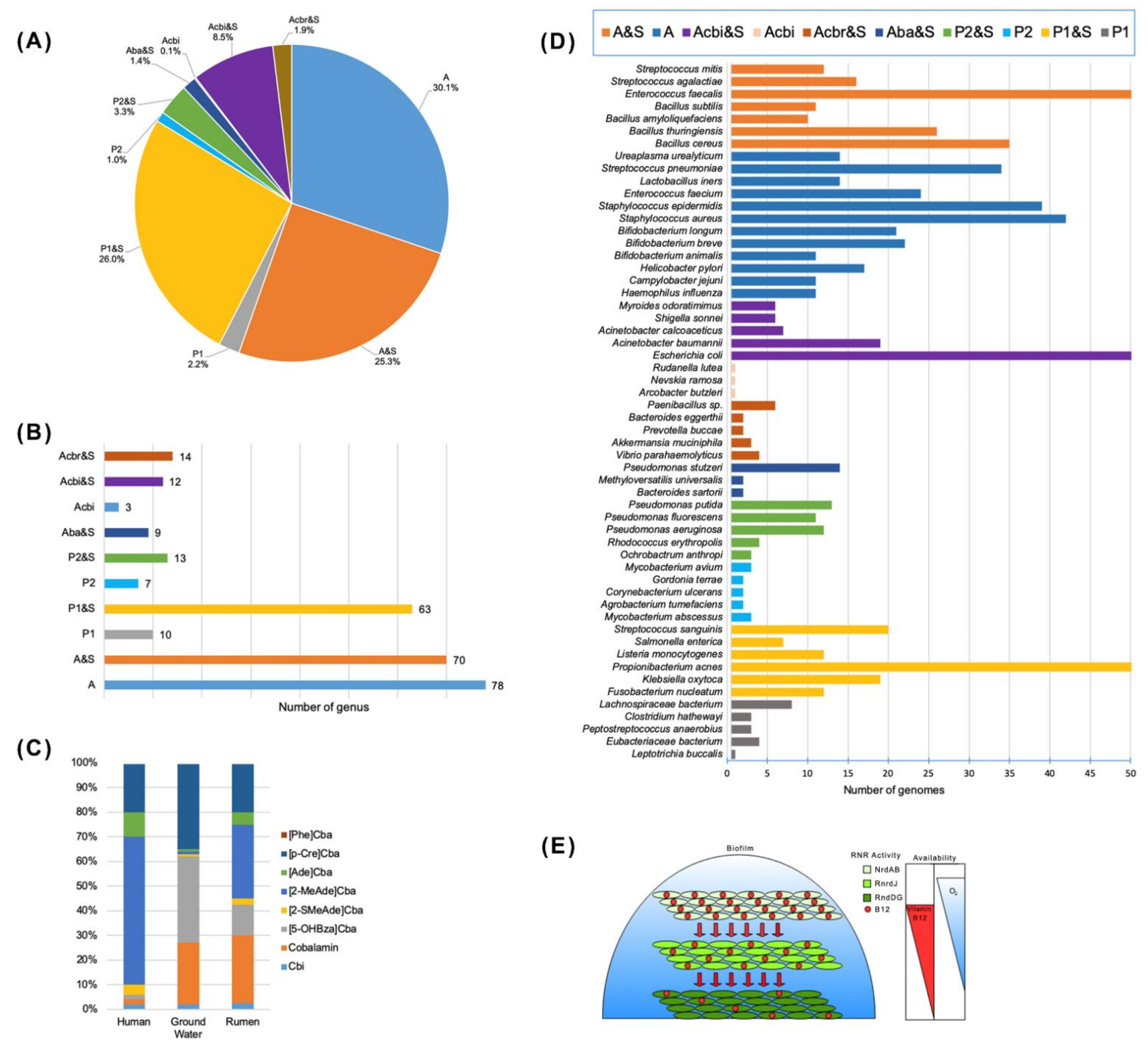
3.2. Cobalamin Biosynthesis Capability Assessment in Bacteria
3.2.1. Genetic Signatures of Cobalamin Pathways and Transport
3.2.2. Metabolic Reconstruction Methods
3.3. Cobamide Uptake in Eukaryotes
4. Cobalamin-Dependent Regulation
4.1. Cobalamin-Dependent Metabolism
4.2. Riboswitch-Mediated Regulation
4.2.1. Riboswitch Structure and Mechanism of Regulation
4.2.2. Riboswitch-Encoding Sequences Location and Function
4.2.3. Ligand Selectivity of Riboswitches
4.3. Light-Dependent Regulation
5. Evolution and Role of B12 Auxotrophy
5.1. Marine Microbial Community Auxotrophy
5.2. Gut Microbial Community Auxothrophy
5.3. Evolutionary Strategy for Cosmopolite Bacteria Auxothrophy
5.4. Mammalian B12 Auxotrophy
6. Cell Factories for Cobalamin Production
6.1. Discovery and Development of Biotechnological B12 Producers
6.1.1. Propionobacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain/Pathway | Strategy/Tactic | Main Precursors | Product Yield | Reference |
|---|---|---|---|---|
| P. denitrificans SC510, aerobic/salvage | Random mutagenesis using radiation (UV light) and chemicals (ethyleneimine and nitrosomethyluretane), overexpression of cobF-cobM gene cluster as well as cobA and cobE genes; optimization of promoters, RBSs, terminators. | sucrose, betaine, DMB | 214 mg/L | [101] |
| S. meliloti MC5-2, aerobic/salvage | Random mutagenesis based on atmospheric and room-temperature plasma (ARTP); overexpression of hemE; deletion of cobI, and usage of a riboswitch based on butB element from Salmonella typhimurium in front of a gfp reporter gene driven by the constitutive promoter PmelA | sucrose, DMB | 156 mg/L | [23] |
| Pseudomonas sp. PCSIR-B-99, aerobic | Optimization of fermentation process | methanol, DMB | 3500 μg/L | [89] |
| S. olivaceus NRRL B-1125, aerobic | Optimization of fermentation process | glucose, DMB | 1–3.3 µg/mL | [90] |
| P. shermanii, anaerobic | Overexpression of biosynthetic genes | glucose, DMB | 206 mg/L | [54] |
| P. freudenreichii CICC 10019 | Optimization of fermentation process | glucose, corn extract, DMB | 58.8 mg/L | [95] |
| B. megaterium, anaerobic | Overexpression of hemACDBL, sirA, cbiXJCDETLFGA, cysGA, cbiY, btuR, glmS, metH, rtpR with xylose-inducible promoter; antisense RNA for hemE, hemZ, sirB. Bypassing of the B12 riboswitch | glucose; ALA; DMB | 0.220 mg/L | [102] |
| B. megaterium, wild strain, anaerobic | Optimization of fermentation process | glucose; ALA; DMB | 204.46 µg/L | [103] |
| E. coli, salvage | The 22 native cob genes located in six operons from P. denitrificans ATCC 13867 were PCR-amplified and cloned in three compatible plasmids under the strong inducible T7 promoter | ALA | 0.65 µg/g | [104] |
| E. coli, aerobic/anaerobic/salvage | Optimization of fermentation process; expression was conducted by assembling six modules comprising 28 genes from R. capsulatus, B. melitensis, S. meliloti, S. typhimurium, and R. palustris | ALA; glycine, succinic acid, betaine | 307 µg/g | [105] |
| P. freudenreichii IFO12426 | Optimization of fermentation process, overexpression of hem genes from R. sphaeroides | glucose; ALA | 1.46 mg/L | [94] |
| Mesophilic methane bacteria from digested sludge | Optimization of fermentation process: enriching trace metal salts by an electrolysis process | H2/CO2 medium (biogas or coal gas) | 185 mg/L | [106] |
| S. meliloti CGMCC 9638 aerobic/salvage | Optimization of fermentation process (9–12 days) | sucrose, glycine betaine, corn liquor, DMB | at least 50 mg/L (up to 180 mg/L) | [107] |
| P. freudenreichii (food- grade) | Aqueous cereal-based matrices fermentation | malted barley flour, riboflavin, nicotinamide, cobalt | 712 μg/kg | [96] |
| P. freudenreichii | Feed-back inhibition of propionic acid | glucose, DMB, corn steep liquor, cobalt | 59.5 mg/L (0.59 mg/L/h) | [97] |
| P. freudenreichii DSM 20271 | Co-fermentation with L. brevis ATCC 14869 with pH control (pH 5.0) | wheat bran dough and water (15:85) | 332 ± 44 ng/g dry weigth (3 days) | [100] |
6.1.2. Pseudomonades
6.1.3. Bacillus megaterium
6.1.4. Escherichia coli
6.1.5. Other Microorganisms
7. Industrial Bioprocess of Vitamin B12
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
Appendix A
| Species | Genome ID */ Numbers ** | Target Genes (Basic Pathway Variant) | *** Prototroph(P)/ Auxotroph(A) and Salvager(S) |
|---|---|---|---|
| Eubacteriaceae bacterium Leptotrichia buccalis Leptotrichia goodfellowii Fusobacterium mortiferum Peptostreptococcus anaerobius Clostridium tyrobutyricum | 796939.3/4 523794.5/1 596323.3/1 469616.3/1 1122950.3/3 1121342.3/2 | CbiK/CbiX/CbiX2, CbiL, CbiH, CbiF, CbiG, CbiD, CbiJ, CbiT, CbiE, CbiC, CbiA, Co-transporter, CbiP, CbiB, CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO | P1 628 genomes (28.2%) |
| Clostridium hathewayi Lachnospiraceae bacterium Peptostreptococcus anaerobius Bilophila wadsworthia Bacteroides cellulosilyticus Bacteroides dorei Bacteroides fragilis Bacteroides intestinalis Bacteroides pyogenes Bacteroides stercoris Bacteroides uniformis Bacteroides vulgatus Parabacteroides distasonis Parabacteroides goldsteinii Parabacteroides johnsonii Parabacteroides merdae Porphyromonas asaccharolytica Porphyromonas gingivalis Fusobacterium gonidiaformans Fusobacterium necrophorum Fusobacterium nucleatum Fusobacterium periodonticum Citrobacter amalonaticus Citrobacter freundii Citrobacter koseri Klebsiella oxytoca Klebsiella pneumoniae Salmonella enterica (Agona) Salmonella enterica (Typhum) Salmonella enterica Edwardsiella tarda Hafnia alvei Morganella morganii Yersinia enterocolitica Yersinia frederiksenii Yersinia kristensenii Desulfovibrio desulfuricans Propionibacterium acidipropionici Propionibacterium acnes Propionibacterium granulosum Propionibacterium freudenreichii Propionibacterium subsp. Shermanii Bacillus megaterium Lysinibacillus fusiformis Lysinibacillus sphaericus Listeria innocua Listeria monocytogenes Brevibacillus brevis Lactobacillus reuteri Streptococcus sanguinis Clostridium acetobutylicum Clostridium botulinum Clostridium perfringens Ruminococcus obeum Blautia wexlerae Dorea formicigenerans Eubacterium saburreum | 566550.8/3 658081.3/8 596329.3/3 563192.3/1 246787.4/3 997875.3/4 272559.17/6 329854.7/2 1321819.3/3 449673.7/2 411479.10/6 435590.9/5 435591.13/3 927665.4/3 537006.5/2 411477.4/3 879243.3/2 431947.6/4 469615.3/2 1095747.3/3 525283.3/12 546275.3/3 35703.30/4 1006003.3/7 290338.8/3 1191061.3/19 1037908.3/9 1265617.3/2 99287.12/6 550537.5/7 500638.3/6 1002364.3/4 582.34/4 630.33/7 349966.8/2 28152.3/2 525146.4/3 1748.13/3 1091045.3/83 1160719.7/2 66712.6/2 754252.3/1 1348623.7/6 1231627.3/2 1421.39/6 1002366.3/2 882095.3/12 1129895.3/2 557436.4/7 997356.4/20 272562.8/2 441770.6/7 195103.9/4 411459.7/2 1121115.4/2 411461.4/2 887325.3/2 | CbiK/CbiX/CbiX2, CbiL, CbiH, CbiF, CbiG, CbiD, CbiJ, CbiT, CbiE, CbiC, CbiA, Co-transporter, CbiP, CbiB, CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO; +transporters: CbrUVT/BtuBCDF | P1&S |
| Lachnobacterium bovis Clostridium bolteae Clostridium clostridioforme Clostridium symbiosum Roseburia intestinalis Eubacterium rectale Desulfitobacterium hafniense Clostridium difficile Clostridium sordellii Clostridium bifermentans Faecalibacterium prausnitzii Selenomonas ruminantium Veillonella atypica Veillonella parvula | 140626.5/3 997893.5/6 999403.4/9 411472.5/3 536231.5/3 515619.6/3 272564.6/4 499175.4/17 1292036.3/2 1233170.3/3 411483.3/4 1280706.4/2 866776.4/3 686660.3/3 | ChlID, CobN, CbiL, CobG, CbiH, CbiF, CobF, CbiJ, CbiT, CbiE, CbiC, CbiA, Co-transporter, CbiP, CbiB, CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO | P2 97 genomes (4,4%) |
| Mesorhizobium loti Agrobacterium tumefaciens Corynebacterium glucuronolyticum Corynebacterium kroppenstedtii Corynebacterium ulcerans Gordonia terrae Mycobacterium avium Mycobacterium abscessus Bradyrhizobium japonicum Ochrobactrum anthropi Burkholderia cepacia Comamonas testosteroni Delftia acidovorans Pseudomonas aeruginosa Pseudomonas fluorescens Pseudomonas putida Dietzia cinnamea Rhodococcus equi Rhodococcus erythropolis Rhodococcus rhodochrous | 266835.9/2 176299.3/2 548478.3/2 645127.4/2 945711.3/2 1316928.3/2 243243.7/3 36809.5/3 224911.1/3 439375.7/3 292.26/3 1009852.3/3 883100.3/3 208964.1/12 294.123/11 303.200/13 1223524.3/2 525370.3/2 1833.80/4 1330040.3/2 | +transporters: CbrUVT/BtuBCDF | P2&S |
| Bacteroides sartorii Methyloversatilis universalis Pseudomonas stutzeri | 1236538.3/2 999628.3/2 32042.3/14 | CbiA, CbiP, CbiB, CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO; +transporters: CbrUVT/BtuBCDF | Aba&S, 32 genomes (1.4%) |
| Bacteroides eggerthii Prevotella buccae Akkermansia muciniphila Vibrio furnissii Vibrio parahaemolyticus Vibrio mimicus Paenibacillus sp. Paenisporosarcina sp. Turicibacter sp. | 665953.3/2 873513.3/2 349741.6/3 675811.4/2 670.436/4 674.46/3 1033743.3/6 1078085.3/2 702450.3/2 | CbiP, CbiB, CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO; +transporters: CbrUVT/BtuBCDF | Acbr&S, 43 genomes (1.9%) |
| Rudanella lutea Nevskia ramosa Arcobacter butzleri Bacteroides ovatus Prevotella oralis Myroides odoratimimus Escherichia albertii Escherichia coli Enterobacter aerogenes Shigella dysenteriae Shigella flexneri | 1089547.3/1 1122603.3/1 888827.3/1 411476.11/6 873533.3/2 76832.7/6 208962.36/3 481805.6/88 548.155/5 622.8/3 623.158/5 | CobU, CobS, CobC/CblZ, CobT, CobD, BtuR/PduO | Acbi 193 genomes (8.7%) |
| Shigella sonnei Acinetobacter baumannii Acinetobacter calcoaceticus Acinetobacter pittii Acinetobacter haemolyticus Acinetobacter johnsonii Acinetobacter junii Acinetobacter radioresistens | 624.634/6 400667.7/19 981331.5/7 48296.83/4 707232.3/3 1217662.4/2 1217665.3/2 981334.6/4 | +transporters: CbrUVT/BtuBCDF | Acbi&S, mostly E. coli and Acinetobacter spp. |
| Lactobacillus brevis Lactobacillus crispatus Capnocytophaga ochracea Sutterella wadsworthensis Burkholderiales bacterium Neisseria flavescens Neisseria mucosa Actinobacillus pleuropneumoniae Haemophilus influenza Moraxella catarrhalis Campylobacter coli Campylobacter jejuni Campylobacter ureolyticus Helicobacter pylori Actinomyces odontolyticus Mobiluncus curtisii Mobiluncus mulieris Varibaculum cambriense Bifidobacterium adolescentis Bifidobacterium angulatum Bifidobacterium animalis Bifidobacterium bifidum Bifidobacterium breve Bifidobacterium dentium Bifidobacterium longum Micrococcus luteus Rothia mucilaginosa Cellulosimicrobium cellulans Gemella haemolysans Staphylococcus aureus Staphylococcus epidermidis Staphylococcus intermedius Staphylococcus lugdunensis Aerococcus viridans Enterococcus faecium Lactobacillus fermentum Lactobacillus iners Lactobacillus johnsonii Lactobacillus plantarum Lactobacillus ruminis Lactobacillus salivarius Pediococcus acidilactici Leuconostoc mesenteroides Weissella koreensis Lactococcus lactis Streptococcus intermedius Streptococcus parasanguinis Streptococcus pneumoniae Erysipelotrichaceae bacterium | 525310.3/2 679188.3/7 521097.5/3 742823.3/3 469610.4/2 546264.5/2 546266.6/2 1224142.3/5 727.532/11 480.212/2 195.462/6 683083.3/11 883165.3/3 85962.8/17 411466.7/3 887326.3/3 585199.3/4 184870.3/3 367928.6/3 518635.5/2 442563.4/11 500634.6/5 518634.7/22 871562.3/5 216816.111/21 1270.32/8 553201.3/4 1710.9/2 546270.5/2 1229492.3/42 176280.1/39 1141106.7/1 1034809.4/5 655812.3/2 1138892.3/24 525325.3/5 888801.3/14 525330.3/5 525338.3/7 525362.3/3 1041521.3/5 862514.3/4 203120.7/2 1045854.4/2 746361.3/6 1316583.3/6 1114965.3/4 561276.4/34 658659.3/6 | BtuR/PduO | A 1235 genomes (55.4%) |
| Anaerococcus prevotii Finegoldia magna Mycoplasma hominis Mycoplasma pneumoniae Ureaplasma parvum Ureaplasma urealyticum Lactobacillus reuteri Bacteroides caccae Bacteroides salyersiae Bacteroides thetaiotaomicron Bacteroides xylanisolvens Prevotella bivia Prevotella corporis Prevotella intermedia Prevotella melaninogenica Prevotella nigrescens Achromobacter xylosoxidans Alcaligenes faecalis Aeromonas caviae Aeromonas hydrophila Aeromonas media Aeromonas veronii Cronobacter sakazakii Enterobacter asburiae Enterobacter cloacae Enterobacter hormaechei Escherichia fergusonii Pantoea agglomerans Proteus mirabilis Proteus vulgaris Providencia alcalifaciens Providencia rettgeri Providencia stuartii Serratia marcescens Yersinia pseudotuberculosis Plesiomonas shigelloides Aggregatibacter aphrophilus Haemophilus parainfluenzae Stenotrophomonas maltophilia Brachyspira pilosicoli Tropheryma whipplei Eggerthella lenta Bacillus cereus Bacillus mycoides Bacillus thuringiensis Bacillus amyloliquefaciens Bacillus licheniformis Bacillus subtilis Enterococcus faecalis Enterococcus casseliflavus Lactobacillus acidophilus Lactobacillus amylovorus Lactobacillus casei Lactobacillus delbrueckii Lactobacillus gasseri Lactobacillus helveticus Lactobacillus rhamnosus Lactococcus garvieae Streptococcus agalactiae Streptococcus infantis Streptococcus mitis Streptococcus mutans Streptococcus oralis Streptococcus parauberis Streptococcus pyogenes Streptococcus thermophilus Eubacterium siraeum Ruminococcus albus | 879305.3/2 334413.6/5 1267000.3/2 272634.6/4 515608.4/5 565575.4/14 349123.6/3 411901.7/2 997887.3/2 226186.1/4 657309.4/2 868129.3/2 28128.5/2 28131.5/2 553174.6/2 702439.3/2 762376.5/2 511.9/2 648.77/3 644.34/4 651.7/3 654.45/5 290339.8/5 61645.42/3 716541.4/6 1259823.3/3 550694.3/3 549.25/7 525369.4/4 585.10/2 520999.6/2 521000.6/2 471874.6/4 615.109/5 349747.9/4 703.7/2 985008.3/4 888828.3/5 40324.126/4 1133568.3/2 203267.6/2 479437.5/2 1396.420/35 526997.3/4 527031.3/26 692420.6/10 279010.13/4 1423.136/11 525271.4/62 565654.4/4 1314884.3/4 695560.3/2 321967.11/6 321956.7/4 324831.13/7 585520.4/4 1088720.3/6 420889.6/3 1309806.3/16 889204.3/4 864567.3/12 1257041.3/9 1161421.3/9 936154.3/5 864568.3/9 1051074.3/6 428128.7/3 246199.4/1 | BtuR/PduO; +transporters: CbrUVT/BtuBCDF | A&S |
| Species | Biosynthesis Pathway | Classification |
|---|---|---|
| Streptomyces coelicolor | aerobic | Actinobacteria |
| Streptomyces griseus | ||
| Agrobacterium tumefaciens Methylobacterium extorquens M. dichloromethanicum Methylosinus trichosporium Sinorhizobium meliloti | aerobic | α-Proteobacteria Rhizobiales |
| Dinoroseobacter shibae Rhodobacter capsulatus Rhodobacter sphaeroides Ruegeria pomeryoi | aerobic | α-Proteobacteria Rhodobacterales |
| Rhodospirillum rubrum | aerobic | α-Proteobacteria Rhodospirillales |
| Methylobacter luteus Methylococcus capsulatus | aerobic | γ-Proteobacteria Methylococcales |
| Pseudomonas denitrificans Pseudomonas putida | aerobic | γ-Proteobacteria Pseudomonadales |
| Propionibacterium acidipropionici Propionibacterium freudenreichii Propionibacterium shermanii | anaerobic | Actinobacteria |
| Prochlorococcus sp. MIT9313 Synechococcus elongates Synechocystis sp. PCC6803 | anaerobic | Cyanobacteria |
| Bacillus megaterium Listeria monocytogenes | anaerobic | Firmicutes Bacilli Bacillales |
| Lactobacillus coryniformis Lactobacillus reuteri Lactobacillus rossiae | anaerobic | Firmicutes Bacilli Lactobacillales |
| Clostridium cochlearium Clostridium kluyveri | anaerobic | Firmicutes Clostridia Clostridiales Clostridiaceae |
| Acetobacterium woodii Eubacterium barkeri Eubacterium hallii Eubacterium limosum | anaerobic | Firmicutes Clostridia Clostridiales Eubacteriaceae |
| Blautia hydrogenotrophica Clostridium phytofermentans | anaerobic | Firmicutes Clostridia Clostridiales Lachnospiraceae |
| Dehalobacter restrictus Desulfitobacterium hafniense Desulfitobacterium sp. PCE1 Desulfotomaculum reducens | anaerobic | Firmicutes Clostridia Clostridiales Peptococcaceae |
| Moorella thermoacetica | anaerobic | Firmicutes Clostridia Thermoanaerobacterale |
| Pelosinus fermentans Sporomusa ovata Veillonella parvula | anaerobic | Firmicutes Negativicutes |
| Desulfobacterium autotrophicum Desulfobulbus propionicus | anaerobic | δ-Proteobacteria Desulfobacterales |
| Desulfovibrio desulfuricans Desulfovibrio vulgaris | anaerobic | δ-Proteobacteria Desulfovibrionales |
| Geobacter lovleyi Geobacter sulfurreducens Pelobacter propionicus | anaerobic | δ-Proteobacteria Desulfuromonadales |
| Sulfurospirillum multivorans | anaerobic | ε-Proteobacteria |
| Salmonella typhimurium Yersinia enterocolitica | anaerobic | γ-Proteobacteria |
| Thermosipho africanus H1760334 Thermosipho africanus TCF52B | anaerobic | Thermotogae |
| Aphanizomenon flos-aquae Crocosphaera watsonii | unknown | Cyanobacteria |
| Clostridium tetanomorphum | unknown | Firmicutes |
References
- Froese, D.S.; Fowler, B.; Baumgartner, M.R. Vitamin B12, folate, and the methionine remethylation cycle—biochemistry, pathways, and regulation. J. Inherit. Metab. Dis. 2019, 42, 673–685. [Google Scholar] [CrossRef] [PubMed]
- Elías-Arnanz, M. Anaerobic bacteria need their vitamin B12 to digest estrogen. Proc. Natl. Acad. Sci. USA 2020, 117, 1833–1835. [Google Scholar] [CrossRef]
- Jugder, B.-E.; Ertan, H.; Bohl, S.; Lee, M.; Marquis, C.P.; Manefield, M. Organohalide respiring bacteria and reductive dehalogenases: Key tools in organohalide bioremediation. Front. Microbiol. 2016, 7, 249. [Google Scholar] [CrossRef]
- Guo, M.; Chen, Y. Coenzyme cobalamin: Biosynthesis, overproduction and its application in dehalogenation-a review. Rev. Environ. Sci. Biotechnol. 2018, 17, 259–284. [Google Scholar] [CrossRef]
- Acevedo-Rocha, C.G.; Gronenberg, L.S.; Mack, M.; Commichau, F.M.; Genee, H.J. Microbial cell factories for the sustainable manufacturing of B vitamins. Curr. Opin. Biotechnol. 2019, 56, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Danchin, A.; Braham, S. Coenzyme B12 synthesis as a baseline to study metabolite contribution of animal microbiota. Microb. Biotechnol. 2017, 10, 688–701. [Google Scholar] [CrossRef] [PubMed]
- Shelton, A.N.; Seth, E.C.; Mok, K.C.; Han, A.W.; Jackson, S.N.; Haft, D.R.; Tag, M.E. Uneven distribution of cobamide biosynthesis and dependence in bacteria predicted by comparative genomics. ISME J. 2018, 13, 789–804. [Google Scholar] [CrossRef] [PubMed]
- Rempel, S.; Colucci, E.; de Gier, J.W.; Guskov, A.; Slotboom, D.J. Cysteine-mediated decyanation of vitamin B12 by the predicted membrane transporter BtuM. Nat. Commun. 2018, 9, 3038. [Google Scholar] [CrossRef]
- Martens, J.H.; Barg, H.; Warren, M.J.; Jahn, D. Microbial production of vitamin B12. Appl. Microbiol. Biotechnol. 2002, 58, 275–285. [Google Scholar] [CrossRef]
- Biedendieck, R.; Malten, M.; Barg, H.; Bunk, B.; Martens, J.-H.; Deery, E.; Leech, H.; Warren, M.J.; Jahn, D. Metabolic engineering of cobalamin (vitamin B12) production in Bacillus megaterium. Microbial. Biotechnol. 2010, 3, 24–37. [Google Scholar] [CrossRef]
- Xu, Y.; Xiang, S.; Ye, K.; Zheng, Y.; Feng, X.; Zhu, X.; Chen, J.; Chen, Y. Cobalamin (Vitamin B12) Induced a shift in microbial composition and metabolic activity in an in vitro colon simulation. Front. Microbiol. 2018, 9, 2780. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Arzamasov, A.A.; Khoroshkin, M.S.; Iablokov, S.N.; Leyn, S.A.; Peterson, S.N.; Novichkov, P.S.; Osterman, A.L. Micronutrient Requirements and Sharing Capabilities of the Human Gut Microbiome. Front. Microbiol. 2019, 10, 1316. [Google Scholar] [CrossRef]
- Magnúsdóttir, S.; Ravcheev, D.; de Crécy-Lagard, V.; Thiele, I. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Front. Genet. 2015, 6, 148. [Google Scholar] [CrossRef]
- Heal, K.R.; Qin, W.; Ribalet, F.; Bertagnolli, A.D.; Coyote-Maestas, W.; Hmelo, L.R.; Moffett, J.W.; Devol, A.H.; Armbrust, E.V.; Stahl, D.A.; et al. Two distinct pools of B12 analogs reveal community interdependencies in the ocean. Proc. Natl. Acad. Sci. USA 2017, 114, 364–369. [Google Scholar] [CrossRef]
- Romine, M.F.; Rodionov, D.A.; Maezato, Y.; Anderson, L.N.; Nandhikonda, P.; Rodionova, I.A.; Carre, A.; Li, X.; Xu, C.; Clauss, T.R.W. Roles for vitamin B12 in regulation. Proc. Natl. Acad. Sci. USA 2017, 114, E1205–E1214. [Google Scholar] [CrossRef]
- Frischkorn, K.R.; Haley, S.T.; Dyhrman, S.T. Coordinated gene expression between Trichodesmium and its microbiome over day-night cycles in the North Pacific Subtropical Gyre. ISME J. 2018, 12, 997–1007. [Google Scholar] [CrossRef]
- Walworth, N.G.; Lee, M.D.; Suffridge, C.; Qu, P.; Fu, F.; Saito, M.A.; Webb, E.A.; Sanu-do-Wilhelmy, E.A.; Hutchins, D.A. Functional genomics and phylogenetic evidence suggest genus-wide cobalamin production by the globally distributed marine nitrogen fixer Trichodesmium. Front. Microbiol. 2018, 9, 189. [Google Scholar] [CrossRef]
- Joglar, V.; Álvarez-Salgado, X.; Gago-Martinez, A.; Leao, J.M.; Pérez-Martínez, C.; Pontiller, B.; Lundin, D.; Pinhassi, J.; Fernández, E.; Teira, E. Cobalamin and microbial plankton dynamics along a coastal to offshore transect in the Eastern North Atlantic Ocean. Environ. Microbiol. 2020, 23, 1559–1583. [Google Scholar] [CrossRef]
- Lu, X.; Katherine, R.H.; Ingalls, A.E.; Doxey, A.C.; Nefeld, J.D. Metagenomic and chemical characterization of soil cobalamin production. ISME J. 2020, 14, 53–66. [Google Scholar] [CrossRef]
- Bryant, D.A.; Hunter, C.N.; Warren, M.J. Biosynthesis of the modified tetrapyrroles—The pigments of life. J. Biol. Chem. 2020, 295, 6888–6925. [Google Scholar] [CrossRef]
- Koyyalamudi, S.R.; Jeong, S.-C.; Cho, K.Y.; Pang, G. Vitamin B-12 Is the Active Corrinoid Produced in Cultivated White Button Mushrooms (Agaricus bisporus). J. Agric. Food Chem. 2009, 57, 14–6327. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A.D.; Nemoto-Smith, E.; Deery, E.; Baker, J.A.; Schroeder, S.; Brown, D.G.; Tullet, J.M.A.; Howard, M.J.; Brown, I.R.; Smith, A.G.; et al. Construction of Fluorescent Analogs to Follow the Uptake and Distribution of Cobalamin (Vitamin B12) in Bacteria, Worms, and Plants. Cell Chem. Biol. 2018, 25, 941–951. [Google Scholar] [CrossRef]
- Cai, Y.; Xia, M.; Dong, H.; Qian, Y.; Zhang, T.; Zhu, B.; Wu, J.; Zhang, D. Engineering a vitamin B12 high throughput screening system by riboswitch sensor in Sinorhizobium meliloti. BMC Biotechnol. 2018, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Kang, J.; Zhang, D. Microbial production of vitamin B12: A review and future perspectives. Microb. Cell Factories 2017, 16, 15. [Google Scholar] [CrossRef] [PubMed]
- Piwowarek, K.; Lipińska, E.; Hać-Szymańczuk, E.; Kieliszek, M.; Ścibisz, I. Propionibacterium spp.-source of propionic acid, vita-min B12, and other metabolites important for the industry. Appl. Microbiol. Biotechnol. 2018, 102, 515–538. [Google Scholar] [PubMed]
- Nguyen-Vo, T.P.; Ainala, S.K.; Kim, J.R.; Park, S. Analysis and characterization of coenzyme B12 biosynthetic gene clusters and improvement of B12 biosynthesis in Pseudomonas denitrificans ATCC 13867. FEMS Microbiol. Lett. 2018, 365, fny211. [Google Scholar] [CrossRef] [PubMed]
- Deptula, P.; Kylli, P.; Chamlagain, B.; Holm, L.; Kostiainen, R.; Piironen, V.; Savijoki, K.; Varmanen, P. BluB/CobT2 fusion enzyme activity reveals mechanisms responsible for production of active form of vitamin B12 by Propionibacterium freudenreichii. Microb. Cell Fact. 2015, 14, 186. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.J.; Warren, M.J. The anaerobic biosynthesis of vitamin B12. Biochem. Soc. Trans. 2012, 40, 581–586. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Midford, P.E.; Ong, W.K.; Paley, S.; Subhraveti, P.; Karp, P.D. The MetaCyc database of metabolic pathways and enzymes—A 2019 update. Nucleic Acids Res. 2020, 48, D445–D453. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2020, 30, gkaa970. [Google Scholar] [CrossRef]
- Rodionov, D.A.; Vitreschak, A.G.; Mironov, A.A.; Gelfand, M.S. Comparative genomics of the vitamin B12 metabolism and regulation in prokaryotes. J. Biol. Chem. 2003, 278, 41148–41159. [Google Scholar] [CrossRef]
- Torres, A.C.; Vannini, V.; Bonacina, J.; Font, G.; Saavedra, L.; Taranto, M.P. Cobalamin production by Lactobacillus coryniformis: Biochemical identification of the synthetized corrinoid and genomic analysis of the biosynthetic cluster. BMC Microbiol. 2016, 16, 240. [Google Scholar] [CrossRef]
- Perruchon, C.; Vasileiadis, S.; Papadopoulou, E.S.; Karpouzas, D.G. Genome-Based Metabolic Reconstruction Unravels the Key Role of B12 in Methionine Auxotrophy of an Ortho-Phenylphenol-Degrading Sphingomonas haloaromaticamans. Front. Microbiol. 2020, 10, 3009. [Google Scholar] [CrossRef]
- Rupakula, A.; Lu, Y.; Kruse, T.; Boeren, S.; Holliger, C.; Smidt, H.; Maillard, J. Functional genomics of corrinoid starvation in the organohalide-respiring bacterium Dehalobacter restrictus strain PER-K23. Front. Microbiol. 2015, 5, 751. [Google Scholar] [CrossRef]
- Torres, A.C.; Vannini, V.; Font, G.; Saavedra, L.; Taranto, M.P. Novel pathway for corrinoid compounds production in Lactobacillus. Front. Microbiol. 2018, 9, 2256. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Zayas, C.L.; Escalante-Semerena, J.C. Reassessment of the late steps of coenzyme B12 synthesis in Salmonella enterica: Evidence that dephosphorylation of adenosylcobalamin-5′-phosphate by the CobC phosphatase is the last step of the pathway. J. Bacteriol. 2007, 189, 2210–2218. [Google Scholar] [CrossRef]
- Jeske, L.; Placzek, S.; Schomburg, I.; Chang, A.; Schomburg, D. BRENDA in 2019: A European ELIXIR core data resource. Nucleic Acids Res. 2018, 47, D542–D549. [Google Scholar] [CrossRef]
- Mera, P.E.; Maurice, M.S.; Rayment, I.; Escalante-Semerena, J.C. Structural and functional analyses of the human-type corrinoid adenosyltransferase (PduO) from Lactobacillus reuteri. Biochemistry 2007, 46, 13829–13836. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.-Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, D351–D360. [Google Scholar] [CrossRef]
- Heal, K.R.; Kellogg, N.A.; Carlson, L.T.; Lionheart, R.M.; Ingalls, A.E. Metabolic consequences of cobalamin scarcity in the diatom Thalassiosira pseudonana as revealed through metabolomics. Protist 2019, 170, 328–348. [Google Scholar] [CrossRef] [PubMed]
- Tavares, N.K.; VanDrisse, C.M.; Escalante-Semerena, J.C. Rhodobacterales use a unique L-threonine kinase for the assembly of the nucleotide loop of coenzyme B12. Mol. Microbiol. 2018, 110, 239–261. [Google Scholar] [CrossRef] [PubMed]
- Gray, M.J.; Tavares, N.K.; Escalante-Semerena, J.C. The genome of Rhodobacter sphaeroides strain 2.4.1 encodes functional cobinamide salvaging systems of archaeal and bacterial origins. Mol. Microbiol. 2008, 70, 824–836. [Google Scholar]
- Swithers, K.S.; Petrus, A.K.; Secinaro, M.A.; Nesbø, C.L.; Gogarten, J.P.; Noll, K.M.; Butzin, N.C. Vitamin B-12 synthesis and salvage pathways were acquired by horizontal gene transfer to the thermotogales. Genome Biol. Evol. 2012, 4, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Bi, M.; Bourdon, A.K.; Farmer, A.T.; Wang, P.-H.; Molenda, O.; Quaile, A.T.; Jiang, N.; Yang, Y.; Yin, Y.; et al. Purinyl-cobamide is a native prosthetic group of reductive dehalogenases. Nat. Chem. Biol. 2018, 14, 8–14. [Google Scholar] [CrossRef]
- Sokolovskaya, O.M.; Mok, K.C.; Park, J.D.; Tran, J.L.A.; Quanstrom, K.A.; Taga, M.E. Cofactor selectivity in methylmalonyl coenzyme A mutase, a model cobamide-dependent enzyme. mBio 2019, 10, e01303-19. [Google Scholar] [CrossRef] [PubMed]
- Terence, S.C.; Seth, E.C.; Hazra, A.B.; Taga, M.E. Cobamide structure depends on both lower ligand availability and CobT substrate specificity. Chem. Biol. 2013, 20, 1265–1274. [Google Scholar]
- Mok, K.C.; Taga, M.E. Growth inhibition of Sporomusa ovata by incorporation of benzimidazole bases into cobamides. J. Bacteriol. 2013, 195, 1902–1911. [Google Scholar] [CrossRef]
- Chan, C.H.; Escalante-Semerena, J.C. ArsAB, a novel enzyme from Sporomusa ovata activates phenolic bases for adenosylcobamide biosynthesis. Mol. Microbiol. 2011, 81, 952–967. [Google Scholar] [CrossRef]
- Reimer, L.C.; Vetcininova, A.; Carbasse, J.S.; Söhngen, C.; Gleim, D.; Ebeling, C.; Overmann, J. BacDive in 2019: Bacterial phenotypic data for High-throughput biodiversity analysis. Nucleic Acids Res. 2019, 47, D631–D636. [Google Scholar] [CrossRef]
- Mathur, Y.; Sreyas, S.; Datar, P.M.; Sathian, M.B.; Hazra, A.B. CobT and BzaC catalyze the regiospecific activation and methylation of the 5-hydroxybenzimidazole lower ligand in anaerobic cobamide biosynthesis. J. Biol. Chem. 2020, 295, 10522–10534. [Google Scholar] [CrossRef]
- Eppinger, M.; Bunk, B.; Johns, M.A.; Edirisinghe, J.N.; Kutumbaka, K.K.; Koenig, S.S.K.; Creasy, H.H.; Rosovitz, M.J.; Riley, D.R.; Daugherty, S.; et al. Genome sequences of the biotechnologically important Bacillus megaterium strains QM B1551 and DSM319. J. Bacteriol. 2011, 193, 4199–4213. [Google Scholar] [CrossRef]
- Santos, F.; Vera, J.L.; van der Heijden, R.; Valdez, G.; de Vos, W.M.; Sesma, F.; Hugenholtz, J. The complete coenzyme B12 biosynthesis gene cluster of Lactobacillus reuteri CRL1098. Microbiology 2008, 154, 81–93. [Google Scholar] [CrossRef]
- Sych, M.J.; Lacroix, C.; Stevens, M.J.A. Vitamin B12-physiology, production and application. In Industrial Biotechnology of Vitamins, Biopigments, and Antioxidants; Vandamme, E.J., Revuelta, J.L., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 129–159. [Google Scholar]
- Parizzi, L.P.; Grassi, M.C.B.; Llerena, L.A.; Carazzolle, M.F.; Queiroz, V.L.; Lunardi, I.; Zeidler, A.F.; Teixeira, P.J.P.L.; Mieczkowski, P.; Rincones, J.; et al. The genome sequence of Propionibacterium acidipropionici provides insights into its biotechnological and industrial potential. BMC Genom. 2012, 13, 562. [Google Scholar] [CrossRef]
- Yang, M.; Simpson, D.M.; Wenner, N.; Brownridge, P.; Harman, V.M.; Hinton, J.; Beynon, R.J.; Liu, L.N. Decoding the stoichiometric composition and organisation of bacterial metabolosomes. Nat. Commun. 2020, 11, 1976. [Google Scholar] [CrossRef]
- Cheng, S. Recycling of Vitamin B12 and NAD+ within the Pdu Microcompartment of Salmonella enterica. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2010. [Google Scholar]
- Crespo, A.; Blanco-Cabra, N.; Torrents, E. Aerobic Vitamin B12 Biosynthesis Is Essential for Pseudomonas aeruginosa Class II Ribonucleotide Reductase Activity During Planktonic and Biofilm Growth. Front. Microbiol. 2018, 9, 986. [Google Scholar] [CrossRef]
- Ma, A.T.; Tyrell, B.; Beld, J. Specificity of cobamide remodeling, uptake and utilization in Vibrio cholerae. Mol. Microbiol. 2020, 113, 89–102. [Google Scholar] [CrossRef]
- Degnan, P.H.; Barry, N.A.; Mok, K.C.; Taga, M.E. Goodman AL. Human gut microbes use multiple transporters to distinguish vitamin B₁₂ analogs and compete in the gut. Cell Host Microbe 2014, 15, 47–57. [Google Scholar] [CrossRef]
- Gude, S.; Taga, M.E. Multi-faceted approaches to discovering and predicting microbial nutritional interactions. Curr. Opin. Biotechnol. 2020, 62, 58–64. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Lawrence, A.D.; Holzer, A.; Kudahl, U.J.; Sasso, S.; Krautler, B.; Scanlan, D.J.; Warren, M.J.; Smith, A.G. Cyanobacteria and eukaryotic algae use different chemical variants of vitamin B12. Curr. Biol. 2016, 26, 999–1008. [Google Scholar] [CrossRef]
- Zengler, K.; Zaramela, L.S. The social network of microorganisms - how auxotrophies shape complex communities. Nat. Rev. Microbiol. 2018, 16, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Noh, M.H.; Lim, H.G.; Moon, D.; Park, S.; Jung, G.Y. Auxotrophic Selection Strategy for Improved Production of Coenzyme B12 in Escherichia coli. Science 2020, 23, 100890. [Google Scholar] [CrossRef] [PubMed]
- Kazamia, E.; Czesnick, H.; Nguyen, T.T.; Croft, M.T.; Sherwood, E.; Sasso, S.; Hodson, S.J.; Warren, M.J.; Smit, A.G. Mutualistic interactions between vitamin B12-dependent algae and heterotrophic bacteria exhibit regulation. Environ. Microbiol. 2012, 14, 1466–1476. [Google Scholar] [CrossRef] [PubMed]
- Degnan, P.H.; Taga, M.E.; Goodman, A.L. Vitamin B12 as a modulator of gut microbial ecology. Cell Metab. 2014, 20, 769–778. [Google Scholar] [CrossRef]
- Putnam, E.E.; Goodman, A.L. B vitamin acquisition by gut commensal bacteria. PLoS Pathog. 2020, 16, e1008208. [Google Scholar] [CrossRef]
- Klug, G. Beyond catalysis: Vitamin B12 as a cofactor in gene regulation. Mol. Microbiol. 2014, 91, 635–640. [Google Scholar] [CrossRef]
- Chen, G.E.; Canniffe, D.P.; Hunter, C.N. O2-dependent cyclases of chlorophyll biosynthesis. Proc. Natl. Acad. Sci. USA 2017, 114, 6280–6285. [Google Scholar] [CrossRef]
- Parent, A.; Guillot, A.; Benjdia, A.; Chartier, G.; Leprince, J.; Berteau, O. The B12-radical SAM enzyme PoyC catalyzes valine C-beta-methylation during polytheonamide biosynthesis. J. Am. Chem. Soc. 2016, 138, 15515–15518. [Google Scholar] [CrossRef]
- Bridwell-Rabb, J.; Drennan, C.L. Vitamin B12 in the spotlight again. Curr. Opin. Chem. Biol. 2017, 37, 63–70. [Google Scholar] [CrossRef]
- Padmanabhan, S.; Jost, M.; Drennan, C.L.; Elías-Arnanz, M. A new facet of vitamin B 12: Gene regulation by cobalamin-based photoreceptors. Annu. Rev. Biochem. 2017, 86, 485–514. [Google Scholar] [CrossRef]
- Nef, C.; Jung, S.; Mairet, F.; Kaas, R.; Garnier, M. How haptophytes microalgae mitigate vitamin B12 limitation. Sci. Rep. 2019, 9, 8417. [Google Scholar] [CrossRef]
- Polaski, J.T.; Kletzien, O.A.; Drogalis, L.K.; Batey, R.T. A functional genetic screen reveals sequence preferences within a key tertiary interaction in cobalamin riboswitches required for ligand selectivity. Nucleic Acids Res. 2018, 46, 9094–9105. [Google Scholar] [CrossRef]
- Sinumvayo, J.P.; Zhao, C.; Tuyishime, P. Recent advances and future trends of riboswitches: Attractive regulatory tools. World J. Microbiol. Biotechnol. 2018, 34, 171. [Google Scholar] [CrossRef]
- Zhu, X.; Wang, X.; Zhang, C.; Wang, X.; Gu, Q. A riboswitch sensor to determine vitamin B12 in fermented foods. Food Chem. 2015, 175, 523–528. [Google Scholar] [CrossRef]
- Singh, S.K.; Srivastava, A.K.; Kumar, A.; Singh, V.K.; Yadav, D.; Modi, A.; Wang, W.; Singh, T.; Sitther, V.; Singh, P.K.; et al. Cyanobacterial genome editing toolboxes: Recent advancement and future projections for basic and synthetic biology researches. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.P., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 129–149. [Google Scholar]
- Vitreschak, A.G.; Rodionov, D.A.; Mironov, A.A.; Gelfand, M.S. Regulation of the vitamin B12 metabolism and transport in bacteria by a conserved RNA structural element. RNA 2003, 9, 1084–1097. [Google Scholar] [CrossRef]
- Pérez, A.A.; Rodionov, D.A.; Bryant, D.A. Identification and Regulation of Genes for Cobalamin Transport in the Cyanobacterium Synechococcus sp. Strain PCC 7002. J. Bacteriol. 2016, 198, 2753–2761. [Google Scholar] [CrossRef]
- Johnson, J.E., Jr.; Reyes, F.E.; Polaski, J.T.; Batey, R.T. B12 cofactors directly stabilize an mRNA regulatory switch. Nature 2012, 492, 133–137. [Google Scholar] [CrossRef]
- Helliwell, K.E.; Scaife, M.A.; Sasso, S.; Araujo, A.P.U.; Purton, S.; Smith, A.G. Unraveling vitamin B12-responsive gene regulation in algae. Plant Physiol. 2014, 165, 388–397. [Google Scholar] [CrossRef]
- Gómez-Consarnau, L.; Sachdeva, R.; Gifford, S.M.; Cutter, L.S.; Fuhrman, J.A.; Sañudo-Wilhelmy, S.A.; Moran, M.A. Mosaic patterns of B-vitamin synthesis and utilization in a natural marine microbial community. Environ. Microbiol. 2018, 20, 2809–2823. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.; Lenski, R.E.; Zinser, E.R. The Black Queen Hypothesis: Evolution of Dependencies through Adaptive Gene Loss. mBio 2012, 3, e00036-12. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, K.E.; Wheeler, G.L.; Leptos, K.C.; Goldstein, R.E.; Smith, A.G. Insights into the evolution of vitamin B12 auxotrophy from sequenced algal genomes. Mol. Biol. Evol. 2011, 28, 2921–2933. [Google Scholar] [CrossRef] [PubMed]
- Sousa, F.L.; Shavit-Grievink, L.; Allen, J.F.; Martin, W.F. Chlorophyll biosynthesis gene evolution indicates photosystem gene duplication, not photosystem merger, at the origin of oxygenic photosynthesis. Genome Biol. Evol. 2013, 5, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Rodionov, D.A.; Leyn, S.A.; Tran, D.; Iablokov, S.N.; Ding, H.; Peterson, D.A.; Osterman, A.L.; Peterson, S.N. B-Vitamin Sharing Promotes Stability of Gut Microbial Communities. Front. Microbiol. 2019, 10, 1485. [Google Scholar] [CrossRef] [PubMed]
- González-Montaña, J.-R.; Escalera-Valente, F.; Alonso, A.J.; Lomillos, J.M.; Robles, R.; Alonso, M.E. Relationship between Vitamin B12 and Cobalt Metabolism in Domestic Ruminant: An Update. Animals 2020, 10, 1855. [Google Scholar] [CrossRef]
- Surendran, S.; Adaikalakoteswari, A.; Saravanan, P.; Shatwaan, I.A.; Lovegrove, J.A.; Vimaleswaran, K.S. An update on vitamin B12-related gene polymorphisms and B12 status. Genes Nutr. 2018, 13, 2. [Google Scholar] [CrossRef]
- Riaz, M.; Ansari, Z.A.; Iqbal, F.; Akram, M. Microbial production of vitamin B12 by methanol utilizing strain of Pseudomonas species. Pak. J. Biochem. Mol. Biol. 2007, 40, 5–10. [Google Scholar]
- Hall, H.H.; Benedict, R.G.; Wiesen, C.F.; Smith, C.E.; Jackson, R.W. Studies on vitamin B12 production with Streptomyces olivaceous. Appl. Microbiol. 1953, 1, 124–129. [Google Scholar] [CrossRef]
- Chaudhary, A.K.; Dhakal, D.; Sohng, J.K. An insight into the “omics” based engineering of streptomycetes for secondary metabolite overproduction. BioMed Res. Int. 2013, 2013, 968518. [Google Scholar] [CrossRef]
- Hugenschmidt, S.; Schwenninger, S.M.; Lacroix, C. Concurrent high production of natural folate and vitamin B12 using a co-culture process with Lactobacillus plantarum SM39 and Propionibacterium freudenreichii DF13. Process Biochem. 2011, 46, 1063–1070. [Google Scholar] [CrossRef]
- Kiatpapan, P.; Murooka, Y. Construction of an expression vector for propionibacteria and its use in production of 5-aminolevulinic acid by Propionibacterium freudenreichii. Appl. Microbiol. Biotechnol. 2001, 56, 144–149. [Google Scholar] [CrossRef]
- Piao, Y.; Yamashita, M.; Kawaraichi, N.; Asegawa, R.; Ono, H.; Murooka, Y. Production of vitamin B12 in genetically engineered Propionibacterium freudenreichii. J. Biosci. Bioeng. 2004, 98, 167–173. [Google Scholar] [CrossRef]
- Wang, P.; Wang, Y.; Liu, Y.; Shi, H.; Su, Z. Novel in situ product removal technique for simultaneous production of propionic acid and vitamin B12 by expanded bed adsorption bioreactor. Bioresour. Technol. 2012, 104, 652–659. [Google Scholar] [CrossRef]
- Chamlagain, B.; Sugito, T.A.; Deptula, P.; Edelmann, M.; Kariluoto, S.; Varmanen, P.; Piironen, V. In situ production of active vitamin B12 in cereal matrices using Propionibacterium freudenreichii. Food Sci. Nutr. 2018, 6, 67–76. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, Z.; Jiao, Y.; Liu, S.; Wang, Y. Improved propionic acid and 5,6-dimethylbenzimidazole control strategy for vitamin B12 fermentation by Propionibacterium freudenreichii. J. Biotechnol. 2015, 193, 123–129. [Google Scholar] [CrossRef]
- Thirupathaiah, Y.; Swarupa Rani, C.; Sudhakara Reddy, M.; Sudhakara Reddy, M.; Venkateswar Rao, L. Effect of chemical and microbial vitamin B12 analogues on production of vitamin B12. World J. Microbiol. Biotechnol. 2012, 28, 2267–2271. [Google Scholar] [CrossRef]
- Yu, Y.; Zhu, X.; Shen, Y. Enhancing the vitamin B12 production and growth of Propionibacterium freudenreichii in tofu wastewater via a light-induced vitamin B12 riboswitch. Appl. Microbiol. Biotechnol. 2015, 99, 10481–10488. [Google Scholar] [CrossRef]
- Xie, C.; Coda, R.; Chamlagain, B.; Varmanen, P.; Piironen, V.; Katina, K. Co-fermentation of Propionibacterium freudenreichii and Lactobacillus brevis in Wheat Bran for in situ Production of Vitamin B12. Front. Microbiol. 2019, 10, 1541. [Google Scholar] [CrossRef]
- Li, K.-T.; Liu, D.-H.; Chu, J.; Wang, Y.-H.; Zhuang, Y.-P.; Zhang, S.-L. An effective and simplified pH-stat control strategy for the industrial fermentation of vitamin B12 by Pseudomonas denitrificans. Bioprocess Biosyst. Eng. 2008, 31, 605–610. [Google Scholar] [CrossRef]
- Moore, S.J.; Mayer, M.J.; Biedendieck, R.; Deery, E.; Warren, M.J. Towards a cell factory for vitamin B12 production in Bacillus megaterium: Bypassing of the cobalamin riboswitch control elements. Nat. Biotechnol. 2014, 31, 553–561. [Google Scholar] [CrossRef]
- Mohammed, Y.; Lee, B.; Kang, Z.; Du, G. Development of a two-step cultivation strategy for the production of vitamin B12 by Bacillus megaterium. Microb. Cell Fact. 2014, 13, 102. [Google Scholar] [CrossRef]
- Ko, Y.; Ashok, S.; Ainala, S.K.; Sankaranarayanan, M.; Chun, A.Y.; Jung, G.Y.; Park, S. Coenzyme B 12 can be produced by engineered Escherichia coli under both anaerobic and aerobic conditions. Biotechnol. J. 2014, 9, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.; Li, D.; Kang, J.; Jiang, P.; Sun, J.; Zhang, D. Metabolic engineering of Escherichia coli for de novo biosynthesis of vitamin B12. Nat. Commun. 2018, 9, 4917. [Google Scholar] [CrossRef] [PubMed]
- Maekawa, T.; Zhang, Z. Method for Producing Vitamin B12 from Hydrogen-Metabolizing Methane Bacterium. U.S. Patent US2005227332A1, 13 October 2005. [Google Scholar]
- Zhang, D.; Li, S.; Dong, H.; Xia, M.; Fang, H.; Zhou, W.; Zheng, P. A kind of Sinorhizobium meliloti strain combinations thereof and application. U.S. Patent CN104342390B, 15 March 2017. [Google Scholar]
- Tanaka, Y.; Kasahara, K.; Izawa, M.; Ochi, K. Applicability of ribosome engineering to vitamin B12 production by Propionibacterium shermanii. Biosci. Biotechnol. Biochem. 2017, 81, 1636–1641. [Google Scholar] [CrossRef] [PubMed]
- Poblete-Castro, I.; Escapa, I.F.; Jager, C.; Jacek, P.; Puchalka, J.; Lam, C.M.C.; Schomburg, D.; Prieto, M.A.; dos Santos, V.A.P.M. The metabolic response of P. putida KT2442 producing high levels of polyhydroxyalkanoate under single- and multiple-nutrient-limited growth: Highlights from a multi-level omics approach. Microb. Cell Fact. 2012, 11, 34. [Google Scholar] [CrossRef]
- Cameron, B.; Briggs, K.; Pridmore, S.; Brefort, G.; Crouzet, J. Cloning and analysis of genes involved in coenzyme B12 biosynthesis in Pseudomonas denitrificans. J. Bacteriol. 1989, 171, 547–557. [Google Scholar] [CrossRef]
- Palacios, O.; Bashan, Y.; de-Bashan, L. Proven and potential involvement of vitamins in interactions of plants with plant growth-promoting bacteria—an overview. Biol. Fertil. Soils 2014, 50, 415–432. [Google Scholar] [CrossRef]
- Falentin, H.; Deutsch, S.M.; Jan, G.; Loux, V.; Thierry, A.; Parayre, S.; Maillard, M.B.; Dherbécourt, J.; Cousin, F.J.; Jardin, J.; et al. The complete genome of Propionibacterium freudenreichii CIRM-BIA1T, a hardy Actinobacterium with food and probiotic applications. PLoS ONE 2010, 5, e11748. [Google Scholar] [CrossRef]
- Loux, V.; Mariadassou, M.; Almeida, S.; Chiapello, H.; Hammani, A.; Buratti, J.; Gendrault, A.; Barbe, V.; Aury, J.-M.; Deutsch, S.-M.; et al. Mutations and genomic islands can explain the strain dependency of sugar utilization in 21 strains of Propionibacterium freudenreichii. BMC Genom. 2015, 16, 296. [Google Scholar] [CrossRef]
- Koskinen, P.; Deptula, P.; Smolander, O.P.; Tamene, F.; Kammonen, J.; Savijoki, K.; Paulin, L.; Piironen, V.; Auvinen, P.; Varmanen, P. Complete genome sequence of Propionibacterium freudenreichii DSM 20271(T). Stand. Genom. Sci. 2015, 10, 83. [Google Scholar] [CrossRef]
- Miyano, K.; Ye, K.; Shimizu, K. Improvement of vitamin B12 fermentation by reducing in the inhibitory metabolites by cell recycle system and mixed culture. J. Biochem. Eng. 2000, 6, 207–214. [Google Scholar] [CrossRef]
- Peng, W.; Cheng, X.; Zhang, H.-Y.; Li, K.-T. The metabolic character- istics of high-production vitamin B12 by Pseudomonas denitrificans under dissolved oxygen step-wise reduction. J. Chem. Technol. Biotechnol. 2014, 89, 1396–1401. [Google Scholar] [CrossRef]
- Wang, Z.-J.; Wang, H.-Y.; Wang, P.; Zhang, Y.-M.; Chu, J.; Zhuang, Y.-P.; Zhang, S.-L. Enhance Vitamin B12 Production by Online CO2 Concentration Control Optimization in 120 m3 Fermentation. J. Bioprocess Biotechniq. 2014, 4, 4. [Google Scholar]





| Species/Strain * | Isolation Source ** | GeneBank Accession ** |
|---|---|---|
| Acetonema longum APO-1 DSM 6540 | gut, termite, Pterotermes occidentis | AFGF00000000 |
| Anaeroarcus burkinensis DSM 6283 | rice field soil | AUMI00000000 |
| Anaeromusa acidaminophila DSM 3853 | anaerobic purification plant | ARGA00000000 |
| Dendrosporobacter quercicolus DSM 1736 | discolored tissue in living oak tree | FNHB00000000 |
| Dialister succinatiphilus YIT 11850 | human faeces | CABKRA000000000 |
| Pelobacter seleniigenes DSM 18267 | freshwater wetland system | QKBM00000000 |
| Pelosinus fermentans R7 DSM 17108 | kaolin deposit Zhuravlinii Log | DJJR00000000 |
| Pelosinus propionicus DSM 13327 | gut of the termite Thoracotermes macrothorax Sjöstedt | FOTS00000000 |
| Sporomusa acidovorans DSM 3132 | effluent from alcohol-distillery plant | FNAM00000000 |
| Sporomusa malonica DSM 5090 | freshwater mud | FWXI00000000 |
| Veillonella atypica KON ATCC 17744 | human mouth | CP020566 |
| Veillonella montpellierensis DSM 17217 | human, gastric fluid of a newborn | AUFY00000000 |
| Veillonella parvula *** DSM 2008 | intestinal tract | CP001820 |
| Veillonella tobetsuensis ATCC BAA-2400 | tongue biofilm of healthy human adult | BBXI00000000 |
| Sporomusa ovate *** DSM 2662 | sugar beet leaf silage | ASXP00000000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balabanova, L.; Averianova, L.; Marchenok, M.; Son, O.; Tekutyeva, L. Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology. Int. J. Mol. Sci. 2021, 22, 4522. https://doi.org/10.3390/ijms22094522
Balabanova L, Averianova L, Marchenok M, Son O, Tekutyeva L. Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology. International Journal of Molecular Sciences. 2021; 22(9):4522. https://doi.org/10.3390/ijms22094522
Chicago/Turabian StyleBalabanova, Larissa, Liudmila Averianova, Maksim Marchenok, Oksana Son, and Liudmila Tekutyeva. 2021. "Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology" International Journal of Molecular Sciences 22, no. 9: 4522. https://doi.org/10.3390/ijms22094522
APA StyleBalabanova, L., Averianova, L., Marchenok, M., Son, O., & Tekutyeva, L. (2021). Microbial and Genetic Resources for Cobalamin (Vitamin B12) Biosynthesis: From Ecosystems to Industrial Biotechnology. International Journal of Molecular Sciences, 22(9), 4522. https://doi.org/10.3390/ijms22094522