
Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Identification of CFTR Activator
2.2. Characterization of Isorhamnetin
2.3. Whole-Cell Patch Clamp in CHO-CFTR Cells
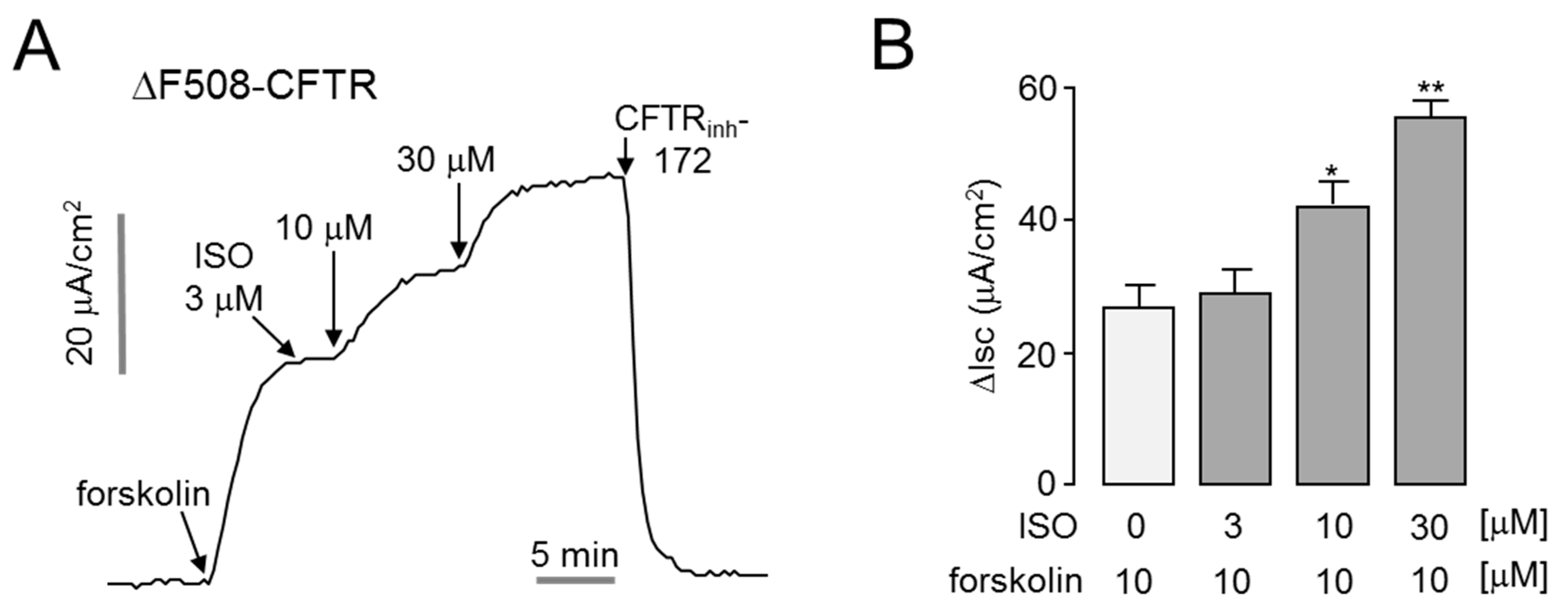
2.4. Isorhamnetin Potentiates Low-Temperature-Rescued ΔF508-CFTR
2.5. Isorhamnetin Increases Ocular Surface Chloride Secretion and Tear Volume
2.6. Isorhamnetin Ameliorates Dry Eye Disease in Scopolamine-Induced Dry Eye Mouse Model
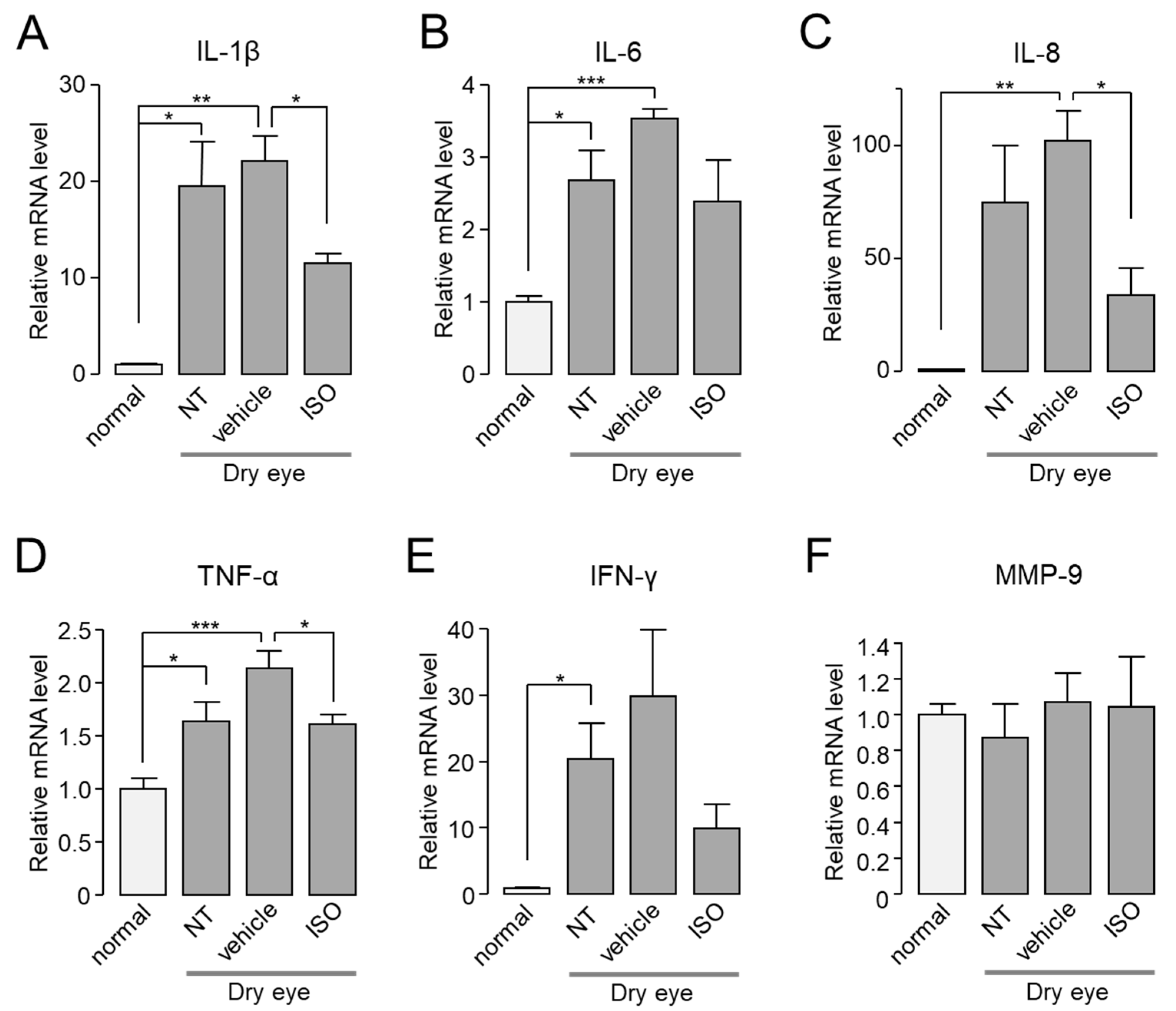
2.7. Isorhamnetin Reduces Ocular Pro-Inflammatory Cytokines in Dry Eye Mouse
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture
4.3. Cell-Based Screening
4.4. Short-Circuit Current
4.5. Patch Clamp
4.6. The cAMP Assay
4.7. FluxOR Potassium Ion Channel Assay
4.8. Cell Proliferation Assays
4.9. Mouse Ocular Potential Difference (PD) Measurement
4.10. Tear Volume
4.11. Fluorescein Staining
4.12. Scopolamine-Induced Dry Eye Mouse Model
4.13. Quantitative PCR Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CFTR | cystic fibrosis transmembrane conductance regulator |
| ANO1 | anoctamin-1 |
| TMEM16A | transmembrane member 16A |
| TNF-α | tumor necrosis factor- α |
| IL | interleukin |
| IFN-γ | interferon- γ |
| ENaC | epithelial sodium channel |
| cAMP | cyclic adenosine monophosphate |
| FRT | fisher rat thyroid |
| CorE | corneal epithelial |
| ConjE | conjunctival epithelial |
| PD | potential difference |
| NF-κB | nuclear factor κ B |
| hERG | human ether-a-go-go-related Gene |
| MMP-9 | matrix metallopeptidase 9 |
References
- O’Brien, P.D.; Collum, L.M. Dry eye: Diagnosis and current treatment strategies. Curr. Allergy Asthma Rep. 2004, 4, 314–319. [Google Scholar] [CrossRef]
- Miljanovic, B.; Dana, R.; Sullivan, D.A.; Schaumberg, D.A. Impact of dry eye syndrome on vision-related quality of life. Am. J. Ophthalmol. 2007, 143, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Uchino, M.; Schaumberg, D.A. Dry Eye Disease: Impact on Quality of Life and Vision. Curr. Ophthalmol. Rep. 2013, 1, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Jani, H. Prevalence and associated factors of dry eye: Our experience in patients above 40 years of age at a Tertiary Care Center. Oman J. Ophthalmol. 2015, 8, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T. Inflammatory Response in Dry Eye. Investig. Opthalmol. Vis. Sci. 2018, 59, DES192–DES199. [Google Scholar] [CrossRef]
- Tatlipinar, S.; Akpek, E.K. Topical ciclosporin in the treatment of ocular surface disorders. Br. J. Ophthalmol. 2005, 89, 1363–1367. [Google Scholar] [CrossRef]
- Pucker, A.D.; Ng, S.M.; Nichols, J.J. Over the counter (OTC) artificial tear drops for dry eye syndrome. Cochrane Database Syst. Rev. 2016, 2, Cd009729. [Google Scholar] [CrossRef]
- Lau, O.C.; Samarawickrama, C.; Skalicky, S.E. P2Y2 receptor agonists for the treatment of dry eye disease: A review. Clin. Ophthalmol. 2014, 8, 327–334. [Google Scholar] [CrossRef]
- Itoh, R.; Kawamoto, S.; Miyamoto, Y.; Kinoshita, S.; Okubo, K. Isolation and characterization of a Ca(2+)-activated chloride channel from human corneal epithelium. Curr. Eye Res. 2000, 21, 918–925. [Google Scholar] [CrossRef]
- Turner, H.C.; Bernstein, A.; Candia, O.A. Presence of CFTR in the conjunctival epithelium. Curr. Eye Res. 2002, 24, 182–187. [Google Scholar] [CrossRef]
- Yu, D.; Thelin, W.R.; Rogers, T.D.; Stutts, M.J.; Randell, S.H.; Grubb, B.R.; Boucher, R.C. Regional differences in rat conjunctival ion transport activities. Am. J. Physiol. Cell Physiol. 2012, 303, C767–C780. [Google Scholar] [CrossRef] [PubMed]
- Mrugacz, M.; Minorowska, A.; Bakunowicz-Lazarczyk, A.; Zywalewska, N. Dry eye syndrome in children with cystic fibrosis. Med. Wieku Rozw. 2004, 8, 865–870. [Google Scholar]
- Pasricha, N.D.; Smith, A.J.; Levin, M.H.; Schallhorn, J.M.; Verkman, A.S. Ocular Surface Potential Difference Measured in Human Subjects to Study Ocular Surface Ion Transport. Transl. Vis. Sci. Technol. 2020, 9, 20. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Phuan, P.W.; Felix, C.M.; Tan, J.A.; Levin, M.H.; Verkman, A.S. Nanomolar-Potency Aminophenyl-1,3,5-triazine Activators of the Cystic Fibrosis Transmembrane Conductance Regulator (CFTR) Chloride Channel for Prosecretory Therapy of Dry Eye Diseases. J. Med. Chem. 2017, 60, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Ostedgaard, L.S.; Rogers, C.S.; Dong, Q.; Randak, C.O.; Vermeer, D.W.; Rokhlina, T.; Karp, P.H.; Welsh, M.J. Processing and function of CFTR-DeltaF508 are species-dependent. Proc. Natl. Acad. Sci. USA 2007, 104, 15370–15375. [Google Scholar] [CrossRef] [PubMed]
- Massingale, M.L.; Li, X.; Vallabhajosyula, M.; Chen, D.; Wei, Y.; Asbell, P.A. Analysis of inflammatory cytokines in the tears of dry eye patients. Cornea 2009, 28, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [CrossRef]
- Pflugfelder, S.C.; De Paiva, C.S.; Moore, Q.L.; Volpe, E.A.; Li, D.Q.; Gumus, K.; Zaheer, M.L.; Corrales, R.M. Aqueous Tear Deficiency Increases Conjunctival Interferon-γ (IFN-γ) Expression and Goblet Cell Loss. Investig. Opthalmol. Vis. Sci. 2015, 56, 7545–7550. [Google Scholar] [CrossRef] [PubMed]
- Lanza, N.L.; Valenzuela, F.; Perez, V.L.; Galor, A. The Matrix Metalloproteinase 9 Point-of-Care Test in Dry Eye. Ocul. Surf. 2016, 14, 189–195. [Google Scholar] [CrossRef]
- The definition and classification of dry eye disease: Report of the Definition and Classification Subcommittee of the International Dry Eye WorkShop (2007). Ocul. Surf. 2007, 5, 75–92. [CrossRef]
- Stern, M.E.; Schaumburg, C.S.; Dana, R.; Calonge, M.; Niederkorn, J.Y.; Pflugfelder, S.C. Autoimmunity at the ocular surface: Pathogenesis and regulation. Mucosal Immunol. 2010, 3, 425–442. [Google Scholar] [CrossRef]
- Ooi, K.G.; Galatowicz, G.; Calder, V.L.; Lightman, S.L. Cytokines and chemokines in uveitis: Is there a correlation with clinical phenotype? Clin. Med. Res. 2006, 4, 294–309. [Google Scholar] [CrossRef]
- Zhou, J.; Wang, S.; Xia, X. Role of intravitreal inflammatory cytokines and angiogenic factors in proliferative diabetic retinopathy. Curr. Eye Res. 2012, 37, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, S.; Sakamoto, T.; Yamashita, T.; Shirasawa, M.; Otsuka, H.; Sonoda, Y. Retinal morphologic changes and concentrations of cytokines in eyes with diabetic macular edema. Retina 2014, 34, 741–748. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-Inflammatory Activity of Natural Products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Lee, C.S. Flavonoid myricetin inhibits TNF-α-stimulated production of inflammatory mediators by suppressing the Akt, mTOR and NF-κB pathways in human keratinocytes. Eur. J. Pharmacol. 2016, 784, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Lee, S.; Son, H.Y.; Park, S.B.; Kim, M.S.; Choi, E.J.; Singh, T.S.; Ha, J.H.; Lee, M.G.; Kim, J.E.; et al. Flavonoids inhibit histamine release and expression of proinflammatory cytokines in mast cells. Arch. Pharm. Res. 2008, 31, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Min, Y.D.; Choi, C.H.; Bark, H.; Son, H.Y.; Park, H.H.; Lee, S.; Park, J.W.; Park, E.K.; Shin, H.I.; Kim, S.H. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-kappaB and p38 MAPK in HMC-1 human mast cell line. Inflamm. Res. 2007, 56, 210–215. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Kanashiro, A.; Fontanari, C.; da Silva, E.V.; Lucisano-Valim, Y.M.; Soares, E.G.; Faccioli, L.H. Anti-inflammatory activity of quercetin and isoquercitrin in experimental murine allergic asthma. Inflamm. Res. 2007, 56, 402–408. [Google Scholar] [CrossRef]
- Majumdar, S.; Srirangam, R. Potential of the bioflavonoids in the prevention/treatment of ocular disorders. J. Pharm. Pharmacol. 2010, 62, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Oh, H.N.; Kim, C.E.; Lee, J.H.; Yang, J.W. Effects of Quercetin in a Mouse Model of Experimental Dry Eye. Cornea 2015, 34, 1130–1136. [Google Scholar] [CrossRef]
- Gilbard, J.P.; Rossi, S.R.; Heyda, K.G.; Dartt, D.A. Stimulation of tear secretion and treatment of dry-eye disease with 3-isobutyl-1-methylxanthine. Arch. Ophthalmol. 1991, 109, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Levin, M.H.; Verkman, A.S. CFTR-regulated chloride transport at the ocular surface in living mice measured by potential differences. Investig. Opthalmol. Vis. Sci. 2005, 46, 1428–1434. [Google Scholar] [CrossRef] [PubMed]
- Shiue, M.H.; Gukasyan, H.J.; Kim, K.J.; Loo, D.D.; Lee, V.H. Characterization of cyclic AMP-regulated chloride conductance in the pigmented rabbit conjunctival epithelial cells. Can. J. Physiol. Pharmacol. 2002, 80, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Morkeberg, J.C.; Edmund, C.; Prause, J.U.; Lanng, S.; Koch, C.; Michaelsen, K.F. Ocular findings in cystic fibrosis patients receiving vitamin A supplementation. Graefe’s Arch. Clin. Exp. Ophthalmol. 1995, 233, 709–713. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.Y.; Zhang, Z.L.; Rahman, K.; Wang, S.J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Jin, C.Y.; Kim, C.H.; Yoo, Y.H.; Choi, S.H.; Kim, G.Y.; Yoon, H.M.; Park, H.T.; Choi, Y.H. Isorhamnetin alleviates lipopolysaccharide-induced inflammatory responses in BV2 microglia by inactivating NF-κB, blocking the TLR4 pathway and reducing ROS generation. Int. J. Mol. Med. 2019, 43, 682–692. [Google Scholar] [CrossRef]
- Li, Y.; Chi, G.; Shen, B.; Tian, Y.; Feng, H. Isorhamnetin ameliorates LPS-induced inflammatory response through downregulation of NF-κB signaling. Inflammation 2016, 39, 1291–1301. [Google Scholar] [CrossRef] [PubMed]
- Chi, G.; Zhong, W.; Liu, Y.; Lu, G.; Lü, H.; Wang, D.; Sun, F. Isorhamnetin protects mice from lipopolysaccharide-induced acute lung injury via the inhibition of inflammatory responses. Inflamm. Res. 2016, 65, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Braghiroli, M.; Iovino, C.; Demarinis, G.; Fossarello, M. A study of refractory cases of persistent epithelial defects associated with dry eye syndrome and recurrent corneal erosions successfully treated with cyclosporine A 0.05% eye drops. Drug Des. Dev. Ther. 2019, 13, 2001–2008. [Google Scholar] [CrossRef] [PubMed]
- Solomon, A.; Dursun, D.; Liu, Z.; Xie, Y.; Macri, A.; Pflugfelder, S.C. Pro- and anti-inflammatory forms of interleukin-1 in the tear fluid and conjunctiva of patients with dry-eye disease. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2283–2292. [Google Scholar]
- Yoon, K.C.; Jeong, I.Y.; Park, Y.G.; Yang, S.Y. Interleukin-6 and tumor necrosis factor-alpha levels in tears of patients with dry eye syndrome. Cornea 2007, 26, 431–437. [Google Scholar] [CrossRef]
- Pendergast, W.; Yerxa, B.R.; Douglass, J.G., 3rd; Shaver, S.R.; Dougherty, R.W.; Redick, C.C.; Sims, I.F.; Rideout, J.L. Synthesis and P2Y receptor activity of a series of uridine dinucleoside 5’-polyphosphates. Bioorg. Med. Chem. Lett. 2001, 11, 157–160. [Google Scholar] [CrossRef]
- Fujihara, T.; Murakami, T.; Fujita, H.; Nakamura, M.; Nakata, K. Improvement of corneal barrier function by the P2Y(2) agonist INS365 in a rat dry eye model. Investig. Ophthalmol. Vis. Sci. 2001, 42, 96–100. [Google Scholar]
- Fujihara, T.; Murakami, T.; Nagano, T.; Nakamura, M.; Nakata, K. INS365 suppresses loss of corneal epithelial integrity by secretion of mucin-like glycoprotein in a rabbit short-term dry eye model. J. Ocul. Pharmacol. Ther. 2002, 18, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Thelin, W.R.; Johnson, M.R.; Hirsh, A.J.; Kublin, C.L.; Zoukhri, D. Effect of topically applied epithelial sodium channel inhibitors on tear production in normal mice and in mice with induced aqueous tear deficiency. J. Ocul. Pharmacol. Ther. 2012, 28, 433–438. [Google Scholar] [CrossRef]
- Gipson, I.K.; Spurr-Michaud, S.; Argüeso, P.; Tisdale, A.; Ng, T.F.; Russo, C.L. Mucin gene expression in immortalized human corneal-limbal and conjunctival epithelial cell lines. Opthalmol. Vis. Sci. 2003, 44, 2496–2506. [Google Scholar] [CrossRef]
- Bang, S.P.; Yeon, C.Y.; Adhikari, N.; Neupane, S.; Kim, H.; Lee, D.C.; Son, M.J.; Lee, H.G.; Kim, J.Y.; Jun, J.H. Cyclosporine A eyedrops with self-nanoemulsifying drug delivery systems have improved physicochemical properties and efficacy against dry eye disease in a murine dry eye model. PLoS ONE 2019, 14, e0224805. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.K.; Park, J.; Kim, B.-R.; Jun, I.; Kim, T.-i.; Namkung, W. Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. Int. J. Mol. Sci. 2021, 22, 3954. https://doi.org/10.3390/ijms22083954
Lee HK, Park J, Kim B-R, Jun I, Kim T-i, Namkung W. Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. International Journal of Molecular Sciences. 2021; 22(8):3954. https://doi.org/10.3390/ijms22083954
Chicago/Turabian StyleLee, Ho K., Jinhong Park, Bo-Rahm Kim, Ikhyun Jun, Tae-im Kim, and Wan Namkung. 2021. "Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice" International Journal of Molecular Sciences 22, no. 8: 3954. https://doi.org/10.3390/ijms22083954
APA StyleLee, H. K., Park, J., Kim, B.-R., Jun, I., Kim, T.-i., & Namkung, W. (2021). Isorhamnetin Ameliorates Dry Eye Disease via CFTR Activation in Mice. International Journal of Molecular Sciences, 22(8), 3954. https://doi.org/10.3390/ijms22083954