
Xyloglucan Oligosaccharides Hydrolysis by Exo-Acting Glycoside Hydrolases from Hyperthermophilic Microorganism Saccharolobus solfataricus


 ,
,  , ,
, , 
 and
and
Abstract
1. Introduction
2. Results
2.1. Protein Analysis of Recombinant GHs
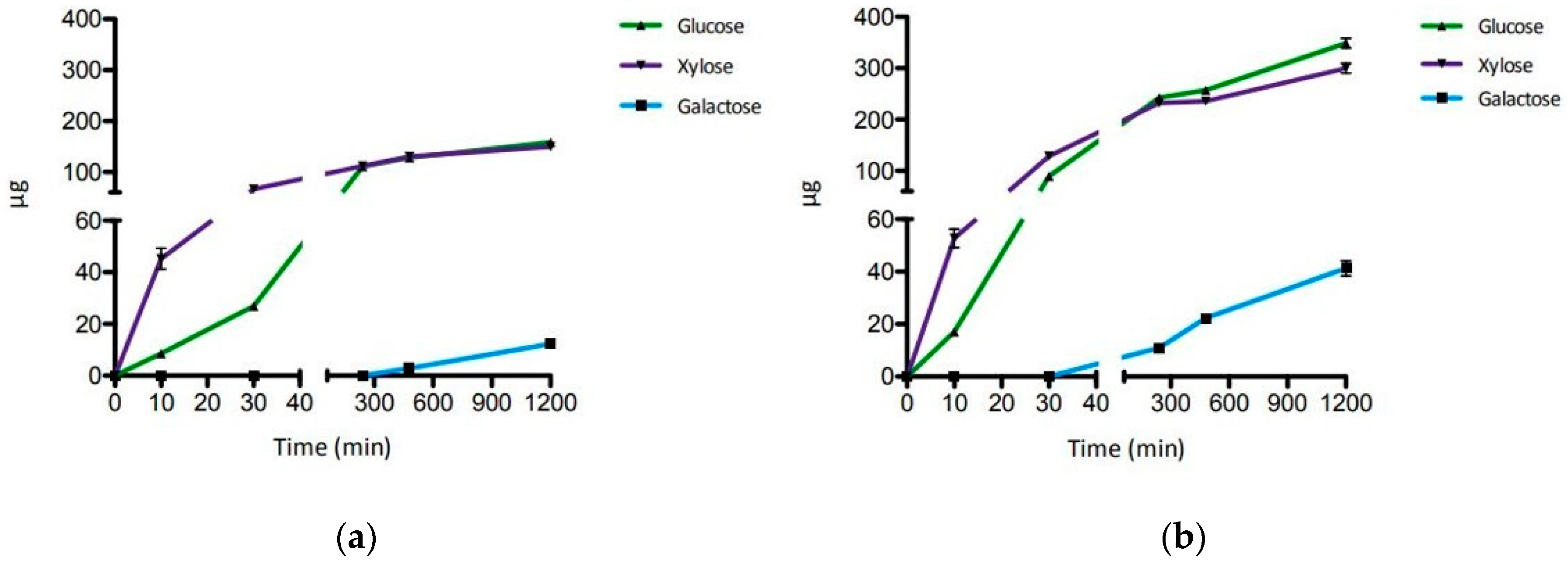
2.2. Hydrolysis of XGO from Tamarind Seeds
2.3. Hydrolysis of XGO from Apple Pomace
3. Discussion
4. Materials and Methods
4.1. Substrates
4.2. Enzyme Expression and Purification
4.3. Protein Analysis of the Recombinant Enzymes
4.4. XGO1 and XGO2 Enzymatic Hydrolysis
4.5. Chemical Hydrolysis
4.6. HPAEC-PAD Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Broeker, J.; Mechelke, M.; Baudrexl, M.; Mennerich, D.; Hornburg, D.; Mann, M.; Schwarz, H.W.; Liebl, W.; Zverlov, V. The hemicellulose-degrading enzyme system of the thermophilic bacterium Clostridium stercorarium: Comparative characterisation and addition of new hemicellulolytic glycoside hydrolases. Biotechnol. Biofuels 2018, 11, 229. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda, R.H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Dai, L.; Ma, L.; Guo, R. Enzymatic degradation of plant biomass and synthetic polymers. Nat. Rev. Chem. 2020, 4, 114–126. [Google Scholar] [CrossRef]
- Strazzulli, A.; Cobucci-Ponzano, B.; Iacono, R.; Giglio, R.; Maurelli, L.; Curci, N.; Schiano-di-Cola, C.; Santangelo, A.; Contursi, P.; Lombard, V.; et al. Discovery of hyperstable carbohydrate-active enzymes through metagenomics of extreme environments. FEBS J. 2019, 287, 1116–1137. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, A.; Krüger, A.; Antranikian, G. Biomass-degrading glycoside hydrolases of archaeal origin. Biotechnol. Biofuels 2020, 13, 153. [Google Scholar] [CrossRef]
- Liu, G.; Qin, Y.; Li, Z.; Qu, Y. Development of highly efficient, low-cost lignocellulolytic enzyme systems in the post-genomic era. Biotechnol. Adv. 2013, 31, 962–975. [Google Scholar] [CrossRef]
- Avanthi, A.; Kumar, S.; Sherpa, K.C.; Banerjee, R. Bioconversion of hemicelluloses of lignocellulosic biomass to ethanol: An attempt to utilize pentose sugars. Biofuels 2017, 8, 431–444. [Google Scholar] [CrossRef]
- Zavyalova, A.V.; Rykova, S.V.; Luninab, N.A.; Sushkovaa, V.I.; Yarotskya, S.V.; Berezina, O.V. Plant Polysaccharide Xyloglucan and Enzymes That Hydrolyze It. Russ. J. Bioorg. Chem. 2019, 45, 845–859. [Google Scholar] [CrossRef]
- Fry, C.; York, S.; Albersheim, P.; Darvill, A.; Hayashi, T.; Joseleau, J.-P.; Kato, Y.; Lorences, P.; Maclachlan, A.; McNeil, M.; et al. An unambiguous nomenclature for xyloglucan-derived oligosaccharides. Physiol. Plant. 1993, 89, 1–3. [Google Scholar] [CrossRef]
- Tuomivaara, S.T.; Yao, K.; O’Neill, M.A.; York, W.S. Generation and structural validation of library of diverse xyloglucan-derived oligosaccharides, including an update on xyloglucan nomenclature. Carbohydr. Res. 2015, 402, 56–66. [Google Scholar] [CrossRef] [PubMed]
- van der Brink, J.; de Vries, R.P. Fungal enzyme sets for plant polysaccharide degradation. Appl. Microbiol. Biotechnol. 2011, 91, 1477–1492. [Google Scholar] [CrossRef] [PubMed]
- Larsbrink, J.; Rogers, T.E.; Hemsworth, G.R.; McKee, L.S.; Tauzin, A.S.; Spadiut, O.; Klinter, S.; Pudlo, N.A.; Urs, K.; Koropatkin, N.M.; et al. A discrete genetic locus confers xyloglucan metabolism in select human gut Bacteroidetes. Nature 2014, 506, 498–502. [Google Scholar] [CrossRef]
- Gardner, J.C. Polysaccharide degradation systems of the saprophytic bacterium Cellvibrio japonicus. World J. Microbiol. Biotechnol. 2016, 32, 121. [Google Scholar] [CrossRef] [PubMed]
- Ravachol, J.; de Philip, P.; Borne, R.; Mansuelle, P.; Maté, M.J.; Perret, S.; Fierobe, H.-P. Mechanisms involved in xyloglucan catabolism by the cellulosome-producing bacterium Ruminiclostridium cellulolyticum. Sci. Rep. 2016, 6, 22770. [Google Scholar] [CrossRef]
- Attia, M.A.; Brumer, H. Recent structural insights into the enzymology of the ubiquitous plant cell wall glycan xyloglucan. Curr. Opin. Struct. Biol. 2016, 40, 43–53. [Google Scholar] [CrossRef]
- Ulas, T.; Riemer, A.; Zaparty, M.; Siebers, B.; Schomburg, D. Genome-Scale Reconstruction and Analysis of the Metabolic Network in the Hyperthermophilic Archaeon Sulfolobus Solfataricus. PLoS ONE 2012, 7, e43401. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.D.; Norio, K.N. Saccharolobus caldissimus gen. nov., sp. nov., a facultatively anaerobic iron-reducing hyperthermophilic archaeon isolated from an acidic terrestrial hot spring, and reclassification of Sulfolobus solfataricus as Saccharolobus solfataricus comb. nov. and Sulfolobus shibatae as Saccharolobus shibatae comb. nov. Int. J. Syst. Evol. Microbiol. 2018, 68, 1271–1278. [Google Scholar]
- Wolf, J.; Stark, H.; Fafenrot, K.; Albersmeier, A.; Pham, T.K.; Müller, K.B.; Meyer, B.H.; Hoffmann, L.; Shen, L.; Albaum, S.P.; et al. A systems biology approach reveals major metabolic changes in the thermoacidophilic archaeon Sulfolobus solfataricus in response to the carbon source L-fucose versus D-glucose. Mol. Microbiol. 2016, 102, 882–908. [Google Scholar] [CrossRef] [PubMed]
- Elferink, M.G.L.; Albers, S.V.; Konings, W.N.; Driessen, A.J.M. Sugar transport in Sulfolobus solfataricus is mediated by two families of binding protein-dependent ABC transporters. Mol. Microbiol. 2008, 39, 1494–1503. [Google Scholar] [CrossRef]
- Lalithambika, S.; Peterson, L.; Dana, K.; Blum, P. Carbohydrate hydrolysis and transport in the extreme thermoacidophile Sulfolobus solfataricus. Appl. Environ. Microbiol. 2012, 78, 7931–7938. [Google Scholar] [CrossRef]
- Moracci, M.; Cobucci-Ponzano, B.; Trincone, A.; Fusco, S.; De Rosai, M.; van der Oost, J.; Sensen, C.W.; Charlebois, R.L.; Rossi, M. Identification and Molecular Characterization of the first α-Xylosidase from an Archaeon. J. Biol. Chem. 2000, 275, 22082–22089. [Google Scholar] [CrossRef] [PubMed]
- Limauro, D.; Cannio, R.; Fiorentino, G.; Rossi, M.; Bartolucci, S. Identification and molecular characterization of an endoglucanase gene, celS, from the extremely thermophilic archaeon Sulfolobus solfataricus. Extremophiles 2001, 5, 213–219. [Google Scholar] [CrossRef]
- Moracci, M.; Ciaramella, M.; Rossi, M. Beta-glycosidase from Sulfolobus solfataricus. Methods Enzymol. 2001, 330, 201–215. [Google Scholar] [PubMed]
- She, Q.; Singh, R.K.; Confalonieri, F.; Zivanovic, Y.; Allard, G.; Awayez, M.J.; Chan-Weiher, C.C.-Y.; Clausen, I.G.; Curtis, B.A.; De Moors, A.; et al. The complete genome of the crenarchaeon Sulfolobus solfataricus P2. Proc. Natl. Acad. Sci. USA 2001, 98, 7835–7840. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Trincone, A.; Giordano, A.; Rossi, M.; Moracci, M. Identification of an archaeal α-L-fucosidase encoded by an interrupted gene. J. Biol. Chem. 2003, 278, 14622–14631. [Google Scholar] [CrossRef] [PubMed]
- Cannio, R.; Di Prizito, N.; Rossi, M.; Morana, A. A xylan-degrading strain of Sulfolobus solfataricus: Isolation and characterization of the xylanase activity. Extremophiles 2004, 8, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Krauss, G.; Cottaz, S.; Driguez, H.; Lipps, G. A highly acid-stable and thermostable endo-beta-glucanase from the thermoacidophilic archaeon Sulfolobus solfataricus. Biochem. J. 2005, 385, 581–588. [Google Scholar] [CrossRef]
- Brouns, S.J.J.; Smits, N.; Wu, H.; Snijders, A.P.L.; Wright, P.C.; de Vos, W.M.; van der Oost, J. Identification of a Novel α-Galactosidase from the Hyperthermophilic Archaeon Sulfolobus solfataricus. J. Bacteriol. 2006, 188, 2392–2399. [Google Scholar] [CrossRef] [PubMed]
- Kambourova., M.; Mandeva, R.; Fiume, I.; Maurelli, L.; Rossi, M.; Morana, A. Hydrolysis of xylan at high temperature by co-action of the xylanase from Anoxybacillus flavithermus BC and the beta-xylosidase/alpha-arabinosidase from Sulfolobus solfataricus Oalpha. J. Appl. Microbiol. 2007, 102, 1586–1593. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Aurilia, V.; Riccio, G.; Henrissat, B.; Coutinho, P.M.; Strazzulli, A.; Padula, A.; Corsaro, M.M.; Pieretti, G.; Pocsfalvi, G.; et al. A New Archaeal β-Glycosidase from Sulfolobus solfataricus. Seeding a novel retaining β-glycan-specific glycoside hydrolase family along with the human non-lysosomal glucosylceramidase GBA2. J. Biol. Chem. 2010, 285, 20691–20703. [Google Scholar] [CrossRef]
- Cobucci-Ponzano, B.; Conte, F.; Strazzulli, A.; Capasso, C.; Fiume, I.; Pocsfalvi, G.; Rossi, M.; Moracci, M. The molecular characterization of a novel GH38 α-mannosidase from the crenarchaeon Sulfolobus solfataricus revealed its ability in de-mannosylating glycoproteins. Biochimie 2010, 92, 1895–1907. [Google Scholar] [CrossRef] [PubMed]
- Honarbakhsh, M.; Villafane, A.A.; Ruhl, I.; Sannino, D.; Bini, E. Development of a thermostable β-glucuronidase-based reporter system for monitoring gene expression in hyperthermophiles. Biotechnol. Bioeng. 2012, 109, 1181–1186. [Google Scholar] [CrossRef]
- Briggs, D.C.; Yoshida-Moriguchi, T.; Zheng, T.; Venzke, D.; Anderson, M.E.; Strazzulli, A.; Moracci, M.; Yu, L.; Hohenester, E.; Campbell, K.P. Structural basis of laminin binding to the LARGE glycans on dystroglycan. Nat. Chem. Biol. 2016, 12, 810–814. [Google Scholar] [CrossRef]
- Larsbrink, J.; Izumi, A.; Ibatullin, F.M.; Nakhai, A.; Gilbert, H.J.; Davies, G.J.; Brumer, H. Structural and enzymatic characterization of a glycoside hydrolase family 31 α-xylosidase from Cellvibrio japonicus involved in xyloglucan saccharification. Biochem. J. 2011, 436, 567–580. [Google Scholar] [CrossRef]
- Kumar, R.; Henrissat, B.; Coutinho, P.M. Intrinsic dynamic behavior of enzyme: Substrate complexes govern the catalytic action of β-galactosidases across clan GH-A. Sci. Rep. 2019, 9, 10346. [Google Scholar] [CrossRef]
- Chen, J.M.; Xia, Y.; Wan, H.; Wang, H.; Liu, X. A complete specific cleavage of glucosyl ester linkages of stevioside for preparing steviol with β-galactosidase from Sulfolobus solfataricus. J. Mol. Catal. B Enzym. 2014, 105, 126–131. [Google Scholar] [CrossRef]
- Bauer, S.; Vasu, P.; Persson, S.; Mort, A.J.; Somerville, C.R. Development and application of a suite of polysaccharide-degrading enzymes for analyzing plant cell walls. Proc. Natl. Acad. Sci. USA 2006, 103, 11417–11422. [Google Scholar] [CrossRef]
- Rogowski, A.; Briggs, J.A.; Mortimer, J.C.; Tryfona, T.; Terrapon, N.; Lowe, E.C.; Baslé, A.; Morland, C.; Day, A.M.; Zheng, H.; et al. Glycan complexity dictates microbial resource allocation in the large intestine. Nat. Commun. 2015, 6, 7481. [Google Scholar] [CrossRef] [PubMed]
- Sakurama, H.; Tsutsumi, E.; Ashida, H.; Katayama, T.; Yamamoto, K.; Kumagai, H. Differences in the Substrate Specificities and Active-Site Structures of Two α-L-Fucosidases (Glycoside Hydrolase Family 29) from Bacteroides thetaiotaomicron. Biosci. Biotechnol. Biochem. 2012, 76, 1022–1024. [Google Scholar] [CrossRef] [PubMed]
- Paper, J.M.; Scott-Craig, J.S.; Cavalier, D.; Faik, A.; Wiemels, R.E.; Borrusch, M.S.; Bongers, M.; Walton, J.D. α-Fucosidases with different substrate specificities from two species of Fusarium. Appl. Microbiol. Biotechnol. 2013, 97, 5371–5380. [Google Scholar] [CrossRef] [PubMed]
- Vecchio, G.; Parascandolo, A.; Allocca, C.; Ugolini, C.; Basolo, F.; Moracci, M.; Strazzulli, A.; Cobucci-Ponzano, B.; Laukkanen, M.; Castellone, M.; et al. Human α-L-fucosidase-1 attenuates the invasive properties of thyroid cancer. Oncotarget 2017, 8, 27075–27092. [Google Scholar] [CrossRef]
- Ashida, H.; Miyake, A.; Kiyohara, M.; Wada, J.; Yoshida, E.; Kumagai, H.; Katayama, T.; Yamamoto, K. Two distinct α-L-fucosidases from Bifidobacterium bifidum are essential for the utilization of fucosylated milk oligosaccharides and glycoconjugates. Glycobiology 2009, 19, 1010–1017. [Google Scholar] [CrossRef]
- Cao, H.; Walton, J.D.; Brumm, P.; Phillips, G.N., Jr. Crystal structure of α-xylosidase from Aspergillus niger in complex with a hydrolyzed xyloglucan product and new insights in accurately predicting substrate specificities of GH31 family glycosidases. ACS Sustain. Chem. Eng. 2020, 8, 2540–2547. [Google Scholar] [CrossRef] [PubMed]
- Chaillou, S.; Lokman, B.C.; Leer, R.J.; Posthuma, C.; Postma, P.W.; Pouwels, P.H. Cloning, sequence analysis, and characterization of the genes involved in isoprimeverose metabolism in Lactobacillus pentosus. J. Bacteriol. 1998, 180, 2312–2320. [Google Scholar] [CrossRef]
- Matsuzawa, T.; Kameyama, A.; Yaoi, K. Identification and characterization of α-xylosidase involved in xyloglucan degradation in Aspergillus oryzae. Appl. Microbiol. Biotechnol. 2019, 104, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, M.; Kang, M.-S.; Mitsuishi, Y.; Mori, H.; Kimura, A. Substrate recognition of Escherichia coli YicI (α-Xylosidase). J. Appl. Glycosci. 2008, 55, 111–118. [Google Scholar] [CrossRef][Green Version]
- Sampedro, J.; Sieiro, C.; Revilla, G.; Gonzalez-Villa, T.; Zarra, I. Cloning and expression pattern of a gene encoding an alpha-xylosidase active against xyloglucan oligosaccharides from Arabidopsis. Plant Physiol. 2001, 126, 910–920. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wang, Q.C.; Zhao, X.; Pu, J.H.; Luan, X.H. Influences of acid reaction and hydrolytic conditions on monosaccharide composition analysis of acid, neutral and basic polysaccharides. Carbohydr. Polym. 2016, 143, 296–300. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Galactose (µg) | Glucose (µg) | Xylose (µg) |
|---|---|---|---|
| TFA hydrolysis 1 | 365 ± 16 | 835 ± 26 | 803 ± 9 |
| LacS 2 + XylS | 12.3 ± 1.1 | 158.3 ± 1.3 | 149.7 ± 1 |
| LacS 3 + XylS | 41.3 ± 2.9 | 348.9 ± 9.3 | 299.9 ± 9.6 |
| LacS1 | ND | ND | ND |
| LacS2 | ND | ND | ND |
| XylS | ND | ND | 89.1 ± 9 |
| Enzymes | Fucose (µg) | Galactose (µg) | Glucose (µg) | Xylose (µg) |
|---|---|---|---|---|
| TFA hydrolysis 1 | 83 ± 2 | 220 ± 18 | 346 ± 25 | 306 ± 11 |
| LacS + XylS + SsαFuc | 75.5 ± 5 | 6.6 ± 1 | 34.7 ± 5.8 | 115 ± 13 |
| LacS + XylS | ND | 3.15 ± 0.1 | 35 ± 0.5 | 118 ± 3.9 |
| XylS +SsαFuc | 80.1 ± 14 | ND | ND | 72.8 ± 9 |
| LacS + SsαFuc | 79.5 ± 9 | 4.7 ± 0.3 | 24.2 ± 4 | ND |
| SsαFuc | 73.3 ± 11 | ND | ND | ND |
| LacS | ND | 1.5 | 13.6 | ND |
| XylS | ND | ND | ND | 73.5 ± 11 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curci, N.; Strazzulli, A.; Iacono, R.; De Lise, F.; Maurelli, L.; Di Fenza, M.; Cobucci-Ponzano, B.; Moracci, M. Xyloglucan Oligosaccharides Hydrolysis by Exo-Acting Glycoside Hydrolases from Hyperthermophilic Microorganism Saccharolobus solfataricus. Int. J. Mol. Sci. 2021, 22, 3325. https://doi.org/10.3390/ijms22073325
Curci N, Strazzulli A, Iacono R, De Lise F, Maurelli L, Di Fenza M, Cobucci-Ponzano B, Moracci M. Xyloglucan Oligosaccharides Hydrolysis by Exo-Acting Glycoside Hydrolases from Hyperthermophilic Microorganism Saccharolobus solfataricus. International Journal of Molecular Sciences. 2021; 22(7):3325. https://doi.org/10.3390/ijms22073325
Chicago/Turabian StyleCurci, Nicola, Andrea Strazzulli, Roberta Iacono, Federica De Lise, Luisa Maurelli, Mauro Di Fenza, Beatrice Cobucci-Ponzano, and Marco Moracci. 2021. "Xyloglucan Oligosaccharides Hydrolysis by Exo-Acting Glycoside Hydrolases from Hyperthermophilic Microorganism Saccharolobus solfataricus" International Journal of Molecular Sciences 22, no. 7: 3325. https://doi.org/10.3390/ijms22073325
APA StyleCurci, N., Strazzulli, A., Iacono, R., De Lise, F., Maurelli, L., Di Fenza, M., Cobucci-Ponzano, B., & Moracci, M. (2021). Xyloglucan Oligosaccharides Hydrolysis by Exo-Acting Glycoside Hydrolases from Hyperthermophilic Microorganism Saccharolobus solfataricus. International Journal of Molecular Sciences, 22(7), 3325. https://doi.org/10.3390/ijms22073325