
Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8


Abstract
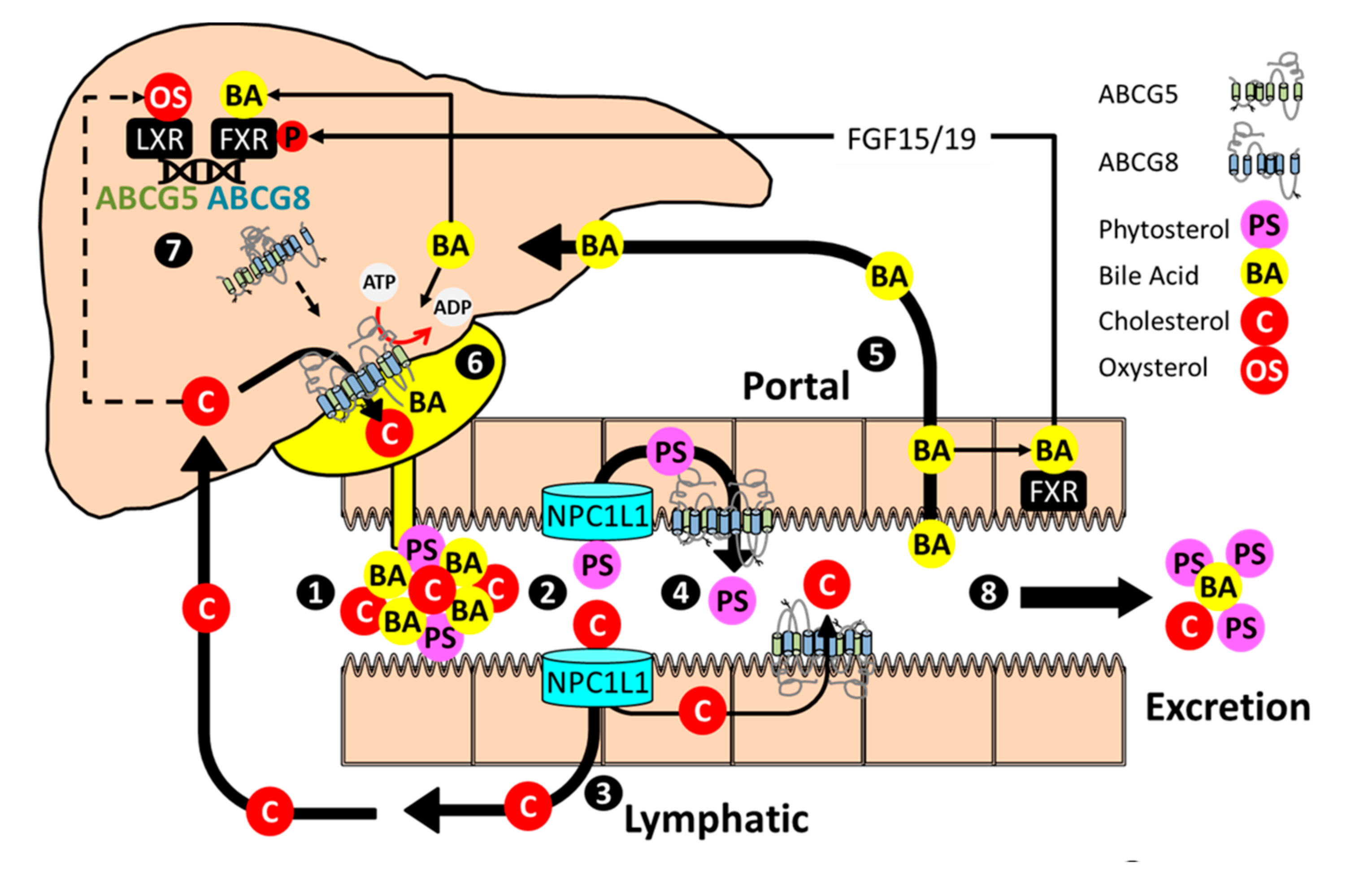
1. Discovery to Therapy
2. Animal Models of Sitosterolemia
3. Heterologous Expression of G5G8
4. Beyond Phytosterols
5. Transcriptional Regulation of G5G8
6. Post-Transcriptional Regulation
7. Sitosterolemia and/or Familial Hypercholesterolemia
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bhattacharyya, A.K.; Connor, W.E. Beta-sitosterolemia and xanthomatosis. A newly described lipid storage disease in two sisters. J. Clin. Investig. 1974, 53, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, T.A. Phytosterolaemia, xanthomatosis and premature atherosclerotic arterial disease: A case with high plant sterol absorption, impaired sterol elimination and low cholesterol synthesis. Eur. J. Clin. Investig. 1980, 10, 27–35. [Google Scholar] [CrossRef]
- Kwiterovich, P.O., Jr.; Bachorik, P.S.; Smith, H.H.; McKusick, V.A.; Connor, W.E.; Teng, B.; Sniderman, A.D. Hyperapobetalipoproteinaemia in two families with xanthomas and phytosterolaemia. Lancet 1981, 1, 466–469. [Google Scholar] [CrossRef]
- Shulman, R.S.; Bhattacharyya, A.K.; Connor, W.E.; Fredrickson, D.S. Beta-sitosterolemia and xanthomatosis. N. Engl. J. Med. 1976, 294, 482–483. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.J.; Wang, C.; Salen, G.; Lam, K.C.; Chan, T.K. Sitosterol and cholesterol metabolism in a patient with coexisting phytosterolemia and cholestanolemia. Metab. Clin. Exp. 1983, 32, 126–133. [Google Scholar] [CrossRef]
- Beaty, T.H.; Kwiterovich, P.O., Jr.; Khoury, M.J.; White, S.; Bachorik, P.S.; Smith, H.H.; Teng, B.; Sniderman, A. Genetic analysis of plasma sitosterol, apoprotein B, and lipoproteins in a large Amish pedigree with sitosterolemia. Am. J. Hum. Genet. 1986, 38, 492–504. [Google Scholar]
- Salen, G.; Horak, I.; Rothkopf, M.; Cohen, J.L.; Speck, J.; Tint, G.S.; Shore, V.; Dayal, B.; Chen, T.; Shefer, S. Lethal atherosclerosis associated with abnormal plasma and tissue sterol composition in sitosterolemia with xanthomatosis. J. Lipid Res. 1985, 26, 1126–1133. [Google Scholar] [CrossRef]
- Nguyen, L.B.; Salen, G.; Shefer, S.; Tint, G.S.; Shore, V.; Ness, G.C. Decreased cholesterol biosynthesis in sitosterolemia with xanthomatosis: Diminished mononuclear leukocyte 3-hydroxy-3-methylglutaryl coenzyme A reductase activity and enzyme protein associated with increased low-density lipoprotein receptor function. Metab. Clin. Exp. 1990, 39, 436–443. [Google Scholar] [CrossRef]
- Bhattacharyya, A.K.; Connor, W.E.; Lin, D.S.; McMurry, M.M.; Shulman, R.S. Sluggish sitosterol turnover and hepatic failure to excrete sitosterol into bile cause expansion of body pool of sitosterol in patients with sitosterolemia and xanthomatosis. Arterioscler. Thromb. A J. Vasc. Biol./Am. Heart Assoc. 1991, 11, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Iolascon, A.; Carella, M.; O’Marcaigh, A.S.; Kendra, J.R.; Jowitt, S.N.; Wales, J.K.; Vora, A.; Makris, M.; Manning, N.; et al. Stomatocytic haemolysis and macrothrombocytopenia (Mediterranean stomatocytosis/macrothrombocytopenia) is the haematological presentation of phytosterolaemia. Br. J. Haematol. 2005, 130, 297–309. [Google Scholar] [CrossRef]
- Mushtaq, T.; Wales, J.K.; Wright, N.P. Adrenal insufficiency in phytosterolaemia. Eur. J. Endocrinol. 2007, 157 (Suppl. 1), S61–S65. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, L.; Su, Y.; Wang, G.; Wang, R.; Yu, Z.; Bai, X.; Ruan, C. Specific macrothrombocytopenia/hemolytic anemia associated with sitosterolemia. Am. J. Hematol. 2014, 89, 320–324. [Google Scholar] [CrossRef] [PubMed]
- Bazerbachi, F.; Conboy, E.E.; Mounajjed, T.; Watt, K.D.; Babovic-Vuksanovic, D.; Patel, S.B.; Kamath, P.S. Cryptogenic Cirrhosis and Sitosterolemia: A Treatable Disease If Identified but Fatal If Missed. Ann. Hepatol. 2017, 16, 970–978. [Google Scholar] [CrossRef]
- Salen, G.; Shore, V.; Tint, G.S.; Forte, T.; Shefer, S.; Horak, I.; Horak, E.; Dayal, B.; Nguyen, L.; Batta, A.K.; et al. Increased sitosterol absorption, decreased removal, and expanded body pools compensate for reduced cholesterol synthesis in sitosterolemia with xanthomatosis. J. Lipid Res. 1989, 30, 1319–1330. [Google Scholar] [CrossRef]
- Cobb, M.M.; Salen, G.; Tint, G.S. Comparative effect of dietary sitosterol on plasma sterols and cholesterol and bile acid synthesis in a sitosterolemic homozygote and heterozygote subject. J. Am. Coll. Nutr. 1997, 16, 605–613. [Google Scholar]
- Nguyen, L.B.; Cobb, M.; Shefer, S.; Salen, G.; Ness, G.C.; Tint, G.S. Regulation of cholesterol biosynthesis in sitosterolemia: Effects of lovastatin, cholestyramine, and dietary sterol restriction. J. Lipid Res. 1991, 32, 1941–1948. [Google Scholar] [CrossRef]
- Patel, S.B.; Honda, A.; Salen, G. Sitosterolemia: Exclusion of genes involved in reduced cholesterol biosynthesis. J. Lipid Res. 1998, 39, 1055–1061. [Google Scholar] [CrossRef]
- Patel, S.B.; Salen, G.; Hidaka, H.; Kwiterovich, P.O.; Stalenhoef, A.F.; Miettinen, T.A.; Grundy, S.M.; Lee, M.H.; Rubenstein, J.S.; Polymeropoulos, M.H.; et al. Mapping a gene involved in regulating dietary cholesterol absorption. The sitosterolemia locus is found at chromosome 2p21. J. Clin. Investig. 1998, 102, 1041–1044. [Google Scholar] [CrossRef] [PubMed]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 93, 693–704. [Google Scholar] [CrossRef]
- Repa, J.J.; Turley, S.D.; Lobaccaro, J.A.; Medina, J.; Li, L.; Lustig, K.; Shan, B.; Heyman, R.A.; Dietschy, J.M.; Mangelsdorf, D.J. Regulation of absorption and ABC1-mediated efflux of cholesterol by RXR heterodimers. Science 2000, 289, 1524–1529. [Google Scholar] [CrossRef]
- Ewart, G.D.; Cannell, D.; Cox, G.B.; Howells, A.J. Mutational analysis of the traffic ATPase (ABC) transporters involved in uptake of eye pigment precursors in Drosophila melanogaster. Implications for structure-function relationships. J. Biol. Chem. 1994, 269, 10370–10377. [Google Scholar] [CrossRef]
- Berge, K.E.; Tian, H.; Graf, G.A.; Yu, L.; Grishin, N.V.; Schultz, J.; Kwiterovich, P.; Shan, B.; Barnes, R.; Hobbs, H.H. Accumulation of dietary cholesterol in sitosterolemia caused by mutations in adjacent ABC transporters. Science 2000, 290, 1771–1775. [Google Scholar] [CrossRef]
- Lee, M.H.; Lu, K.; Hazard, S.; Yu, H.; Shulenin, S.; Hidaka, H.; Kojima, H.; Allikmets, R.; Sakuma, N.; Pegoraro, R.; et al. Identification of a gene, ABCG5, important in the regulation of dietary cholesterol absorption. Nat. Genet. 2001, 27, 79–83. [Google Scholar] [CrossRef]
- Van Heek, M.; France, C.F.; Compton, D.S.; McLeod, R.L.; Yumibe, N.P.; Alton, K.B.; Sybertz, E.J.; Davis, H.R., Jr. In vivo metabolism-based discovery of a potent cholesterol absorption inhibitor, SCH58235, in the rat and rhesus monkey through the identification of the active metabolites of SCH48461. J. Pharm. Exp. Ther. 1997, 283, 157–163. [Google Scholar]
- Davis, H.R., Jr.; Zhu, L.J.; Hoos, L.M.; Tetzloff, G.; Maguire, M.; Liu, J.; Yao, X.; Iyer, S.P.; Lam, M.H.; Lund, E.G.; et al. Niemann-Pick C1 Like 1 (NPC1L1) is the intestinal phytosterol and cholesterol transporter and a key modulator of whole-body cholesterol homeostasis. J. Biol. Chem. 2004, 279, 33586–33592. [Google Scholar] [CrossRef] [PubMed]
- Salen, G.; von Bergmann, K.; Lutjohann, D.; Kwiterovich, P.; Kane, J.; Patel, S.B.; Musliner, T.; Stein, P.; Musser, B.; Multicenter Sitosterolemia Study Group. Ezetimibe effectively reduces plasma plant sterols in patients with sitosterolemia. Circulation 2004, 109, 966–971. [Google Scholar] [CrossRef]
- Lutjohann, D.; von Bergmann, K.; Sirah, W.; Macdonell, G.; Johnson-Levonas, A.O.; Shah, A.; Lin, J.; Sapre, A.; Musliner, T. Long-term efficacy and safety of ezetimibe 10 mg in patients with homozygous sitosterolemia: A 2-year, open-label extension study. Int J. Clin. Pract. 2008, 62, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Escola-Gil, J.C.; Quesada, H.; Julve, J.; Martin-Campos, J.M.; Cedo, L.; Blanco-Vaca, F. Sitosterolemia: Diagnosis, investigation, and management. Curr. Atheroscler. Rep. 2014, 16, 424. [Google Scholar] [CrossRef]
- Plosch, T.; Bloks, V.W.; Terasawa, Y.; Berdy, S.; Siegler, K.; Van Der Sluijs, F.; Kema, I.P.; Groen, A.K.; Shan, B.; Kuipers, F.; et al. Sitosterolemia in ABC-transporter G5-deficient mice is aggravated on activation of the liver-X receptor. Gastroenterology 2004, 126, 290–300. [Google Scholar] [CrossRef]
- Klett, E.L.; Lu, K.; Kosters, A.; Vink, E.; Lee, M.H.; Altenburg, M.; Shefer, S.; Batta, A.K.; Yu, H.; Chen, J.; et al. A mouse model of sitosterolemia: Absence of Abcg8/sterolin-2 results in failure to secrete biliary cholesterol. BMC Med. 2004, 2, 5. [Google Scholar] [CrossRef]
- Yu, L.; Hammer, R.E.; Li-Hawkins, J.; Von Bergmann, K.; Lutjohann, D.; Cohen, J.C.; Hobbs, H.H. Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary cholesterol secretion. Proc. Natl. Acad. Sci. USA 2002, 99, 16237–16242. [Google Scholar] [CrossRef]
- Chen, J.; Batta, A.; Zheng, S.; Fitzgibbon, W.R.; Ullian, M.E.; Yu, H.; Tso, P.; Salen, G.; Patel, S.B. The missense mutation in Abcg5 gene in spontaneously hypertensive rats (SHR) segregates with phytosterolemia but not hypertension. BMC Genet. 2005, 6, 40. [Google Scholar]
- Chase, T.H.; Lyons, B.L.; Bronson, R.T.; Foreman, O.; Donahue, L.R.; Burzenski, L.M.; Gott, B.; Lane, P.; Harris, B.; Ceglarek, U.; et al. The mouse mutation “thrombocytopenia and cardiomyopathy” (trac) disrupts Abcg5: A spontaneous single gene model for human hereditary phytosterolemia/sitosterolemia. Blood 2010, 115, 1267–1276. [Google Scholar] [CrossRef] [PubMed]
- Kruit, J.K.; Drayer, A.L.; Bloks, V.W.; Blom, N.; Olthof, S.G.; Sauer, P.J.; de Haan, G.; Kema, I.P.; Vellenga, E.; Kuipers, F. Plant sterols cause macrothrombocytopenia in a mouse model of sitosterolemia. J. Biol. Chem. 2008, 283, 6281–6287. [Google Scholar] [CrossRef]
- Kanaji, T.; Kanaji, S.; Montgomery, R.R.; Patel, S.B.; Newman, P.J. Platelet hyperreactivity explains the bleeding abnormality and macrothrombocytopenia in a murine model of sitosterolemia. Blood 2013, 122, 2732–2742. [Google Scholar] [CrossRef] [PubMed]
- Solca, C.; Tint, G.S.; Patel, S.B. Dietary xenosterols lead to infertility and loss of abdominal adipose tissue in sterolin-deficient mice. J. Lipid Res. 2013, 54, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; von Bergmann, K.; Lutjohann, D.; Hobbs, H.H.; Cohen, J.C. Ezetimibe normalizes metabolic defects in mice lacking ABCG5 and ABCG8. J. Lipid Res. 2005, 46, 1739–1744. [Google Scholar] [CrossRef]
- Tang, W.; Ma, Y.; Jia, L.; Ioannou, Y.A.; Davies, J.P.; Yu, L. Genetic inactivation of NPC1L1 protects against sitosterolemia in mice lacking ABCG5/ABCG8. J. Lipid Res. 2009, 50, 293–300. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, A.L.; Alger, H.M.; Sawyer, J.K.; Kelley, K.L.; Kock, N.D.; Brown, J.M.; Temel, R.E.; Rudel, L.L. Phytosterol feeding causes toxicity in ABCG5/G8 knockout mice. Am. J. Pathol. 2013, 182, 1131–1138. [Google Scholar] [CrossRef]
- Yang, C.; McDonald, J.G.; Patel, A.; Zhang, Y.; Umetani, M.; Xu, F.; Westover, E.J.; Covey, D.F.; Mangelsdorf, D.J.; Cohen, J.C.; et al. Sterol intermediates from cholesterol biosynthetic pathway as liver X receptor ligands. J. Biol. Chem. 2006, 281, 27816–27826. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.A.; Taylor, O.A.; Prendergast, D.R.; Zimmerman, T.L.; Von Furstenberg, R.; Moore, D.D.; Karpen, S.J. Stigmasterol, a soy lipid-derived phytosterol, is an antagonist of the bile acid nuclear receptor FXR. Pediatr. Res. 2007, 62, 301–306. [Google Scholar] [CrossRef]
- Sabeva, N.S.; McPhaul, C.M.; Li, X.; Cory, T.J.; Feola, D.J.; Graf, G.A. Phytosterols differentially influence ABC transporter expression, cholesterol efflux and inflammatory cytokine secretion in macrophage foam cells. J. Nutr. Biochem. 2011, 22, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Nichols, J.A.; Mensink, R.P. Plant sterols and stanols: Effects on mixed micellar composition and LXR (target gene) activation. J. Lipid Res. 2005, 46, 2468–2476. [Google Scholar] [CrossRef]
- Salen, G.; Tint, G.S.; Shefer, S.; Shore, V.; Nguyen, L. Increased sitosterol absorption is offset by rapid elimination to prevent accumulation in heterozygotes with sitosterolemia. Arterioscler. Thromb. A J. Vasc. Biol. Am. Heart Assoc. 1992, 12, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Lütjohann, D.; Björkhem, I.; Ose, L. Phytosterolaemia in a Norwegian family: Diagnosis and characterization of the first Scandinavian case. Scand. J. Clin. Lab. Investig. 1996, 56, 229–240. [Google Scholar] [CrossRef]
- Zhang, L.S.; Xu, M.; Yang, Q.; Lou, D.; Howles, P.N.; Tso, P. ABCG5/G8 deficiency in mice reduces dietary triacylglycerol and cholesterol transport into the lymph. Lipids 2015, 50, 371–379. [Google Scholar] [CrossRef]
- Nguyen, T.M.; Sawyer, J.K.; Kelley, K.L.; Davis, M.A.; Kent, C.R.; Rudel, L.L. ACAT2 and ABCG5/G8 are both required for efficient cholesterol absorption in mice: Evidence from thoracic lymph duct cannulation. J. Lipid Res. 2012, 53, 1598–1609. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Patel, S.B.; Carey, M.C.; Wang, D.Q. Quantifying anomalous intestinal sterol uptake, lymphatic transport, and biliary secretion in Abcg8(-/-) mice. Hepatology 2007, 45, 998–1006. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Li-Hawkins, J.; Hammer, R.E.; Berge, K.E.; Horton, J.D.; Cohen, J.C.; Hobbs, H.H. Overexpression of ABCG5 and ABCG8 promotes biliary cholesterol secretion and reduces fractional absorption of dietary cholesterol. J. Clin. Investig. 2002, 110, 671–680. [Google Scholar] [CrossRef]
- Jakulj, L.; Vissers, M.N.; van Roomen, C.P.; van der Veen, J.N.; Vrins, C.L.J.; Kunne, C.; Stellaard, F.; Kastelein, J.J.P.; Groen, A.K. Ezetimibe stimulates faecal neutral sterol excretion depending on abcg8 function in mice. FEBS Lett. 2010, 584, 3625–3628. [Google Scholar] [CrossRef]
- de Boer, J.F.; Schonewille, M.; Boesjes, M.; Wolters, H.; Bloks, V.W.; Bos, T.; van Dijk, T.H.; Jurdzinski, A.; Boverhof, R.; Wolters, J.C.; et al. Intestinal Farnesoid X Receptor Controls Transintestinal Cholesterol Excretion in Mice. Gastroenterology 2017, 152, 1126–1138.e6. [Google Scholar] [CrossRef] [PubMed]
- Bonde, Y.; Plosch, T.; Kuipers, F.; Angelin, B.; Rudling, M. Stimulation of murine biliary cholesterol secretion by thyroid hormone is dependent on a functional ABCG5/G8 complex. Hepatology 2012, 56, 1828–1837. [Google Scholar] [CrossRef]
- Kosters, A.; Frijters, R.J.; Kunne, C.; Vink, E.; Schneiders, M.S.; Schaap, F.G.; Nibbering, C.P.; Patel, S.B.; Groen, A.K. Diosgenin-induced biliary cholesterol secretion in mice requires Abcg8. Hepatology 2005, 41, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Gupta, S.; Xu, F.; Liverman, A.D.; Moschetta, A.; Mangelsdorf, D.J.; Repa, J.J.; Hobbs, H.H.; Cohen, J.C. Expression of ABCG5 and ABCG8 is required for regulation of biliary cholesterol secretion. J. Biol. Chem. 2005, 280, 8742–8747. [Google Scholar] [CrossRef]
- Yamanashi, Y.; Takada, T.; Yoshikado, T.; Shoda, J.; Suzuki, H. NPC2 regulates biliary cholesterol secretion via stimulation of ABCG5/G8-mediated cholesterol transport. Gastroenterology 2011, 140, 1664–1674. [Google Scholar] [CrossRef]
- Dikkers, A.; de Boer, J.F.; Groen, A.K.; Tietge, U.J. Hepatic ABCG5/G8 overexpression substantially increases biliary cholesterol secretion but does not impact in vivo macrophage-to-feces RCT. Atherosclerosis 2015, 243, 402–406. [Google Scholar] [CrossRef]
- Coy, D.J.; Wooton-Kee, C.R.; Yan, B.; Sabeva, N.; Su, K.; Graf, G.; Vore, M. ABCG5/ABCG8-independent biliary cholesterol excretion in lactating rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G228–G235. [Google Scholar] [CrossRef] [PubMed]
- Groen, A.; Kunne, C.; Jongsma, G.; van den Oever, K.; Mok, K.S.; Petruzzelli, M.; Vrins, C.L.; Bull, L.; Paulusma, C.C.; Oude Elferink, R.P. Abcg5/8 independent biliary cholesterol excretion in Atp8b1-deficient mice. Gastroenterology 2008, 134, 2091–2100. [Google Scholar] [CrossRef]
- Wiersma, H.; Gatti, A.; Nijstad, N.; Oude Elferink, R.P.; Kuipers, F.; Tietge, U.J. Scavenger receptor class B type I mediates biliary cholesterol secretion independent of ATP-binding cassette transporter g5/g8 in mice. Hepatology 2009, 50, 1263–1272. [Google Scholar] [CrossRef]
- Wang, H.H.; Li, X.; Patel, S.B.; Wang, D.Q. Evidence that the adenosine triphosphate-binding cassette G5/G8-independent pathway plays a determinant role in cholesterol gallstone formation in mice. Hepatology 2016, 64, 853–864. [Google Scholar] [CrossRef]
- Wilund, K.R.; Yu, L.; Xu, F.; Hobbs, H.H.; Cohen, J.C. High-level expression of ABCG5 and ABCG8 attenuates diet-induced hypercholesterolemia and atherosclerosis in Ldlr-/-mice. J. Lipid Res. 2004, 45, 1429–1436. [Google Scholar] [CrossRef]
- Wu, J.E.; Basso, F.; Shamburek, R.D.; Amar, M.J.; Vaisman, B.; Szakacs, G.; Joyce, C.; Tansey, T.; Freeman, L.; Paigen, B.J.; et al. Hepatic ABCG5 and ABCG8 overexpression increases hepatobiliary sterol transport but does not alter aortic atherosclerosis in transgenic mice. J. Biol. Chem. 2004, 279, 22913–22925. [Google Scholar] [CrossRef] [PubMed]
- Basso, F.; Freeman, L.A.; Ko, C.; Joyce, C.; Amar, M.J.; Shamburek, R.D.; Tansey, T.; Thomas, F.; Wu, J.; Paigen, B.; et al. Hepatic ABCG5/G8 overexpression reduces apoB-lipoproteins and atherosclerosis when cholesterol absorption is inhibited. J. Lipid Res. 2007, 48, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Calpe-Berdiel, L.; Rotllan, N.; Fievet, C.; Roig, R.; Blanco-Vaca, F.; Escola-Gil, J.C. Liver X receptor-mediated activation of reverse cholesterol transport from macrophages to feces in vivo requires ABCG5/G8. J. Lipid Res. 2008, 49, 1904–1911. [Google Scholar] [CrossRef]
- Altemus, J.B.; Patel, S.B.; Sehayek, E. Liver-specific induction of Abcg5 and Abcg8 stimulates reverse cholesterol transport in response to ezetimibe treatment. Metab. Clin. Exp. 2014, 63, 1334–1341. [Google Scholar] [CrossRef]
- Su, K.; Sabeva, N.S.; Wang, Y.; Liu, X.; Lester, J.D.; Liu, J.; Liang, S.; Graf, G.A. Acceleration of biliary cholesterol secretion restores glycemic control and alleviates hypertriglyceridemia in obese db/db mice. Arter. Thromb. Vasc. Biol. 2014, 34, 26–33. [Google Scholar] [CrossRef]
- Wang, J.; Mitsche, M.A.; Lutjohann, D.; Cohen, J.C.; Xie, X.S.; Hobbs, H.H. Relative roles of ABCG5/ABCG8 in liver and intestine. J. Lipid Res. 2015, 56, 319–330. [Google Scholar] [CrossRef] [PubMed]
- Berge, K.E.; von Bergmann, K.; Lutjohann, D.; Guerra, R.; Grundy, S.M.; Hobbs, H.H.; Cohen, J.C. Heritability of plasma noncholesterol sterols and relationship to DNA sequence polymorphism in ABCG5 and ABCG8. J. Lipid Res. 2002, 43, 486–494. [Google Scholar] [CrossRef]
- Buch, S.; Schafmayer, C.; Volzke, H.; Becker, C.; Franke, A.; von Eller-Eberstein, H.; Kluck, C.; Bassmann, I.; Brosch, M.; Lammert, F.; et al. A genome-wide association scan identifies the hepatic cholesterol transporter ABCG8 as a susceptibility factor for human gallstone disease. Nat. Genet. 2007, 39, 995–999. [Google Scholar] [CrossRef]
- Grunhage, F.; Acalovschi, M.; Tirziu, S.; Walier, M.; Wienker, T.F.; Ciocan, A.; Mosteanu, O.; Sauerbruch, T.; Lammert, F. Increased gallstone risk in humans conferred by common variant of hepatic ATP-binding cassette transporter for cholesterol. Hepatology 2007, 46, 793–801. [Google Scholar] [CrossRef]
- Stender, S.; Frikke-Schmidt, R.; Nordestgaard, B.G.; Tybjaerg-Hansen, A. The ABCG5/8 cholesterol transporter and myocardial infarction versus gallstone disease. J. Am. Coll. Cardiol. 2014, 63, 2121–2128. [Google Scholar] [CrossRef]
- Helgadottir, A.; Thorleifsson, G.; Alexandersson, K.F.; Tragante, V.; Thorsteinsdottir, M.; Eiriksson, F.F.; Gretarsdottir, S.; Bjornsson, E.; Magnusson, O.; Sveinbjornsson, G.; et al. Genetic variability in the absorption of dietary sterols affects the risk of coronary artery disease. Eur. Heart J. 2020, 41, 2618–2628. [Google Scholar] [CrossRef]
- Jiang, Z.Y.; Cai, Q.; Chen, E.Z. Association of three common single nucleotide polymorphisms of ATP binding cassette G8 gene with gallstone disease: A meta-analysis. PLoS ONE 2014, 9, e87200. [Google Scholar]
- Viturro, E.; de Oya, M.; Lasuncion, M.A.; Gorgojo, L.; Moreno, J.M.; Benavente, M.; Cano, B.; Garces, C. Cholesterol and saturated fat intake determine the effect of polymorphisms at ABCG5/ABCG8 genes on lipid levels in children. Genet. Med. 2006, 8, 594–599. [Google Scholar] [CrossRef]
- Jakulj, L.; Vissers, M.N.; Tanck, M.W.; Hutten, B.A.; Stellaard, F.; Kastelein, J.J.; Dallinga-Thie, G.M. ABCG5/G8 polymorphisms and markers of cholesterol metabolism: Systematic review and meta-analysis. J. Lipid Res. 2010, 51, 3016–3023. [Google Scholar] [CrossRef] [PubMed]
- von Kampen, O.; Buch, S.; Nothnagel, M.; Azocar, L.; Molina, H.; Brosch, M.; Erhart, W.; von Schonfels, W.; Egberts, J.; Seeger, M.; et al. Genetic and functional identification of the likely causative variant for cholesterol gallstone disease at the ABCG5/8 lithogenic locus. Hepatology 2013, 57, 2407–2417. [Google Scholar] [CrossRef]
- Ma, L.; Yang, J.; Runesha, H.B.; Tanaka, T.; Ferrucci, L.; Bandinelli, S.; Da, Y. Genome-wide association analysis of total cholesterol and high-density lipoprotein cholesterol levels using the Framingham heart study data. BMC Med. Genet. 2010, 11, 55. [Google Scholar] [CrossRef]
- Li, Q.; Yin, R.X.; Wei, X.L.; Yan, T.T.; Aung, L.H.; Wu, D.F.; Wu, J.Z.; Lin, W.X.; Liu, C.W.; Pan, S.L. ATP-binding cassette transporter G5 and G8 polymorphisms and several environmental factors with serum lipid levels. PLoS ONE 2012, 7, e37972. [Google Scholar] [CrossRef]
- Wu, G.; Li, G.B.; Yao, M.; Zhang, D.Q.; Dai, B.; Ju, C.J.; Han, M. ABCG5/8 variants are associated with susceptibility to coronary heart disease. Mol. Med. Rep. 2014, 9, 2512–2520. [Google Scholar] [CrossRef] [PubMed]
- Nomura, A.; Emdin, C.A.; Won, H.H.; Peloso, G.M.; Natarajan, P.; Ardissino, D.; Danesh, J.; Schunkert, H.; Correa, A.; Bown, M.J.; et al. Heterozygous ATP-binding Cassette Transporter G5 Gene Deficiency and Risk of Coronary Artery Disease. Circ. Genom. Precis. Med. 2020, 13, 417–423. [Google Scholar] [CrossRef]
- Chen, Z.C.; Shin, S.J.; Kuo, K.K.; Lin, K.D.; Yu, M.L.; Hsiao, P.J. Significant association of ABCG8:D19H gene polymorphism with hypercholesterolemia and insulin resistance. J. Hum. Genet. 2008, 53, 757–763. [Google Scholar] [CrossRef][Green Version]
- Gok, O.; Karaali, Z.E.; Acar, L.; Kilic, U.; Ergen, A. ABCG5 and ABCG8 gene polymorphisms in type 2 diabetes mellitus in the Turkish population. Can. J. Diabetes 2015, 39, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, A.; Fatima, S.; Lamri, A.; Bellili-Munoz, N.; Halimi, J.M.; Saulnier, P.J.; Hadjadj, S.; Velho, G.; Marre, M.; Roussel, R.; et al. ABCG8 polymorphisms and renal disease in type 2 diabetic patients. Metab. Clin. Exp. 2015, 64, 713–719. [Google Scholar] [CrossRef]
- Sabeva, N.S.; Rouse, E.J.; Graf, G.A. Defects in the leptin axis reduce abundance of the ABCG5-ABCG8 sterol transporter in liver. J. Biol. Chem. 2007, 282, 22397–22405. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Su, K.; Sabeva, N.S.; Ji, A.; van der Westhuyzen, D.R.; Foufelle, F.; Gao, X.; Graf, G.A. GRP78 rescues the ABCG5 ABCG8 sterol transporter in db/db mice. Metab. Clin. Exp. 2015, 64, 1435–1443. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Sabeva, N.S.; Liu, J.; Wang, Y.; Bhatnagar, S.; van der Westhuyzen, D.R.; Graf, G.A. The ABCG5 ABCG8 sterol transporter opposes the development of fatty liver disease and loss of glycemic control independently of phytosterol accumulation. J. Biol. Chem. 2012, 287, 28564–28575. [Google Scholar] [CrossRef]
- Van der Velde, A.E.; Vrins, C.L.J.; van den Oever, K.; Kunne, C.; Oude Elferink, R.P.J.; Kuipers, F.; Groen, A.K. Direct Intestinal Cholesterol Secretion Contributes Significantly to Total Fecal Neutral Sterol Excretion in Mice. Gastroenterology 2007, 133, 967–975. [Google Scholar] [CrossRef] [PubMed]
- Grefhorst, A.; Verkade, H.J.; Groen, A.K. The TICE Pathway: Mechanisms and Lipid-Lowering Therapies. Methodist Debakey Cardiovasc. J. 2019, 15, 70–76. [Google Scholar]
- Nakano, T.; Inoue, I.; Murakoshi, T. A Newly Integrated Model for Intestinal Cholesterol Absorption and Efflux Reappraises How Plant Sterol Intake Reduces Circulating Cholesterol Levels. Nutrients 2019, 11, 310. [Google Scholar] [CrossRef] [PubMed]
- Repa, J.J.; Berge, K.E.; Pomajzl, C.; Richardson, J.A.; Hobbs, H.; Mangelsdorf, D.J. Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by the liver X receptors alpha and beta. J. Biol. Chem. 2002, 277, 18793–18800. [Google Scholar] [CrossRef]
- Freeman, L.A.; Kennedy, A.; Wu, J.; Bark, S.; Remaley, A.T.; Santamarina-Fojo, S.; Brewer, H.B., Jr. The orphan nuclear receptor LRH-1 activates the ABCG5/ABCG8 intergenic promoter. J. Lipid Res. 2004, 45, 1197–1206. [Google Scholar] [CrossRef] [PubMed]
- Sumi, K.; Tanaka, T.; Uchida, A.; Magoori, K.; Urashima, Y.; Ohashi, R.; Ohguchi, H.; Okamura, M.; Kudo, H.; Daigo, K.; et al. Cooperative interaction between hepatocyte nuclear factor 4 alpha and GATA transcription factors regulates ATP-binding cassette sterol transporters ABCG5 and ABCG8. Mol. Cell Biol. 2007, 27, 4248–4260. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Matozel, M.; Boehme, S.; Kong, B.; Nilsson, L.M.; Guo, G.; Ellis, E.; Chiang, J.Y. Overexpression of cholesterol 7alpha-hydroxylase promotes hepatic bile acid synthesis and secretion and maintains cholesterol homeostasis. Hepatology 2011, 53, 996–1006. [Google Scholar] [CrossRef]
- Wang, J.; Einarsson, C.; Murphy, C.; Parini, P.; Bjorkhem, I.; Gafvels, M.; Eggertsen, G. Studies on LXR- and FXR-mediated effects on cholesterol homeostasis in normal and cholic acid-depleted mice. J. Lipid Res. 2006, 47, 421–430. [Google Scholar] [CrossRef]
- Liu, J.; Lu, H.; Lu, Y.F.; Lei, X.; Cui, J.Y.; Ellis, E.; Strom, S.C.; Klaassen, C.D. Potency of individual bile acids to regulate bile acid synthesis and transport genes in primary human hepatocyte cultures. Toxicol. Sci. 2014, 141, 538–546. [Google Scholar] [CrossRef]
- Byun, S.; Jung, H.; Chen, J.; Kim, Y.C.; Kim, D.H.; Kong, B.; Guo, G.; Kemper, B.; Kemper, J.K. Phosphorylation of hepatic farnesoid X receptor by FGF19 signaling-activated Src maintains cholesterol levels and protects from atherosclerosis. J. Biol. Chem. 2019, 294, 8732–8744. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shen, J.; Wu, T.; Kuang, J.; Liu, Q.; Cheng, S.; Pu, S.; Chen, L.; Li, R.; Li, Y.; et al. Irisin Is Controlled by Farnesoid X Receptor and Regulates Cholesterol Homeostasis. Front. Pharm. 2019, 10, 548. [Google Scholar] [CrossRef]
- Cheng, S.; Zou, M.; Liu, Q.; Kuang, J.; Shen, J.; Pu, S.; Chen, L.; Li, H.; Wu, T.; Li, R.; et al. Activation of Constitutive Androstane Receptor Prevents Cholesterol Gallstone Formation. Am. J. Pathol. 2017, 187, 808–818. [Google Scholar] [CrossRef]
- Sberna, A.L.; Assem, M.; Gautier, T.; Grober, J.; Guiu, B.; Jeannin, A.; Pais de Barros, J.P.; Athias, A.; Lagrost, L.; Masson, D. Constitutive androstane receptor activation stimulates faecal bile acid excretion and reverse cholesterol transport in mice. J. Hepatol. 2011, 55, 154–161. [Google Scholar] [CrossRef]
- Gooijert, K.E.; Havinga, R.; Wolters, H.; Wang, R.; Ling, V.; Tazuma, S.; Verkade, H.J. The mechanism of increased biliary lipid secretion in mice with genetic inactivation of bile salt export pump. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G450–G457. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Pijut, S.S.; Li, J.; Horn, J.; Bradford, E.M.; Leggas, M.; Barrett, T.A.; Graf, G.A. The combination of ezetimibe and ursodiol promotes fecal sterol excretion and reveals a G5G8-independent pathway for cholesterol elimination. J. Lipid Res. 2015, 56, 810–820. [Google Scholar] [CrossRef]
- Wang, H.H.; Portincasa, P.; Mendez-Sanchez, N.; Uribe, M.; Wang, D.Q. Effect of ezetimibe on the prevention and dissolution of cholesterol gallstones. Gastroenterology 2008, 134, 2101–2110. [Google Scholar] [CrossRef] [PubMed]
- Kamisako, T.; Ogawa, H.; Yamamoto, K. Effect of cholesterol, cholic acid and cholestyramine administration on the intestinal mRNA expressions related to cholesterol and bile acid metabolism in the rat. J. Gastroenterol. Hepatol. 2007, 22, 1832–1837. [Google Scholar] [CrossRef] [PubMed]
- Galman, C.; Bonde, Y.; Matasconi, M.; Angelin, B.; Rudling, M. Dramatically increased intestinal absorption of cholesterol following hypophysectomy is normalized by thyroid hormone. Gastroenterology 2008, 134, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Aleksunes, L.M.; Xu, J.; Lin, E.; Wen, X.; Goedken, M.J.; Slitt, A.L. Pregnancy represses induction of efflux transporters in livers of type I diabetic mice. Pharm. Res. 2013, 30, 2209–2220. [Google Scholar] [CrossRef]
- Biddinger, S.B.; Haas, J.T.; Yu, B.B.; Bezy, O.; Jing, E.; Zhang, W.; Unterman, T.G.; Carey, M.C.; Kahn, C.R. Hepatic insulin resistance directly promotes formation of cholesterol gallstones. Nat. Med. 2008, 14, 778–782. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Liu, H.; Guo, H.; Zhang, M.; Mei, D.; Liu, C.; He, L.; Liu, L.; Liu, X. Impaired hepatic and intestinal ATP-binding cassette transporter G5/8 was associated with high exposure of beta-sitosterol and the potential risks to blood-brain barrier integrity in diabetic rats. J. Pharm. Pharm. 2014, 66, 428–436. [Google Scholar] [CrossRef]
- Molusky, M.M.; Hsieh, J.; Lee, S.X.; Ramakrishnan, R.; Tascau, L.; Haeusler, R.A.; Accili, D.; Tall, A.R. Metformin and AMP Kinase Activation Increase Expression of the Sterol Transporters ABCG5/8 (ATP-Binding Cassette Transporter G5/G8) With Potential Antiatherogenic Consequences. Arter. Thromb. Vasc. Biol. 2018, 38, 1493–1503. [Google Scholar] [CrossRef]
- Pan, X.; Bradfield, C.A.; Hussain, M.M. Global and hepatocyte-specific ablation of Bmal1 induces hyperlipidaemia and enhances atherosclerosis. Nat. Commun. 2016, 7, 13011. [Google Scholar] [CrossRef]
- Nishimoto, T.; Pellizzon, M.A.; Aihara, M.; Stylianou, I.M.; Billheimer, J.T.; Rothblat, G.; Rader, D.J. Fish oil promotes macrophage reverse cholesterol transport in mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 1502–1508. [Google Scholar] [CrossRef]
- Kamisako, T.; Tanaka, Y.; Ikeda, T.; Yamamoto, K.; Ogawa, H. Dietary fish oil regulates gene expression of cholesterol and bile acid transporters in mice. Hepatol. Res. Off. J. Jpn. Soc. Hepatol. 2012, 42, 321–326. [Google Scholar] [CrossRef]
- Kim, E.H.; Bae, J.S.; Hahm, K.B.; Cha, J.Y. Endogenously synthesized n-3 polyunsaturated fatty acids in fat-1 mice ameliorate high-fat diet-induced non-alcoholic fatty liver disease. Biochem. Pharm. 2012, 84, 1359–1365. [Google Scholar] [CrossRef]
- De Vogel-van den Bosch, H.M.; de Wit, N.J.; Hooiveld, G.J.; Vermeulen, H.; van der Veen, J.N.; Houten, S.M.; Kuipers, F.; Muller, M.; van der Meer, R. A cholesterol-free, high-fat diet suppresses gene expression of cholesterol transporters in murine small intestine. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1171–G1180. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Hashizume, T.; Morioka, H.; Sadamitsu, S.; Ikari, A.; Miwa, M.; Sugatani, J. Diet-induced lipid accumulation in liver enhances ATP-binding cassette transporter g5/g8 expression in bile canaliculi. Drug Metab. Pharm. 2011, 26, 442–450. [Google Scholar] [CrossRef]
- Apro, J.; Beckman, L.; Angelin, B.; Rudling, M. Influence of dietary sugar on cholesterol and bile acid metabolism in the rat: Marked reduction of hepatic Abcg5/8 expression following sucrose ingestion. Biochem. Biophys. Res. Commun. 2015, 461, 592–597. [Google Scholar] [CrossRef]
- Duan, L.P.; Wang, H.H.; Ohashi, A.; Wang, D.Q. Role of intestinal sterol transporters Abcg5, Abcg8, and Npc1l1 in cholesterol absorption in mice: Gender and age effects. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G269–G276. [Google Scholar] [CrossRef] [PubMed]
- Dieter, M.Z.; Maher, J.M.; Cheng, X.; Klaassen, C.D. Expression and regulation of the sterol half-transporter genes ABCG5 and ABCG8 in rats. Comp. Biochem. Physiol. C Toxicol. Pharm. 2004, 139, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, I.; Kudo, M.; Hamada, T.; Nagao, K.; Oshiro, Y.; Kato, M.; Sugawara, T.; Yamahira, T.; Ito, H.; Tamaru, S.; et al. Dietary soy protein isolate and its undigested high molecular fraction upregulate hepatic ATP-binding cassette transporter G5 and ATP-binding cassette transporter G8 mRNA and increase biliary secretion of cholesterol in rats. J. Nutr. Sci. Vitaminol. 2009, 55, 252–256. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhong, C.Y.; Sun, W.W.; Ma, Y.; Zhu, H.; Yang, P.; Wei, H.; Zeng, B.H.; Zhang, Q.; Liu, Y.; Li, W.X.; et al. Microbiota prevents cholesterol loss from the body by regulating host gene expression in mice. Sci. Rep. 2015, 5, 10512. [Google Scholar] [CrossRef]
- Prasnicka, A.; Cermanova, J.; Hroch, M.; Dolezelova, E.; Rozkydalova, L.; Smutny, T.; Carazo, A.; Chladek, J.; Lenicek, M.; Nachtigal, P.; et al. Iron depletion induces hepatic secretion of biliary lipids and glutathione in rats. Biochim. Biophys. Acta 2017, 1862, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.Y.; Yang, N.; Jiao, R.; Peng, C.; Guan, L.; Huang, Y.; Chen, Z.Y. Dietary calcium decreases plasma cholesterol by down-regulation of intestinal Niemann-Pick C1 like 1 and microsomal triacylglycerol transport protein and up-regulation of CYP7A1 and ABCG 5/8 in hamsters. Mol. Nutr. Food Res. 2011, 55, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Li, J.; Chao, Y.; Bi, Y.; Zhang, W.; Zhang, Y.; Ji, T.; Fu, Y.; Chen, Q.; Zhang, Q.; et al. beta-estradiol adjusts intestinal function via ERbeta and GPR30 mediated PI3K/AKT signaling activation to alleviate postmenopausal dyslipidemia. Biochem. Pharm. 2020, 180, 114134. [Google Scholar] [CrossRef]
- Li, R.; Liu, Y.; Shi, J.; Yu, Y.; Lu, H.; Yu, L.; Liu, Y.; Zhang, F. Diosgenin regulates cholesterol metabolism in hypercholesterolemic rats by inhibiting NPC1L1 and enhancing ABCG5 and ABCG8. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 1124–1133. [Google Scholar] [CrossRef]
- Graf, G.A.; Li, W.P.; Gerard, R.D.; Gelissen, I.; White, A.; Cohen, J.C.; Hobbs, H.H. Coexpression of ATP-binding cassette proteins ABCG5 and ABCG8 permits their transport to the apical surface. J. Clin. Investig. 2002, 110, 659–669. [Google Scholar] [CrossRef]
- Graf, G.A.; Yu, L.; Li, W.P.; Gerard, R.; Tuma, P.L.; Cohen, J.C.; Hobbs, H.H. ABCG5 and ABCG8 are obligate heterodimers for protein trafficking and biliary cholesterol excretion. J. Biol. Chem. 2003, 278, 48275–48282. [Google Scholar] [CrossRef] [PubMed]
- Okiyoneda, T.; Kono, T.; Niibori, A.; Harada, K.; Kusuhara, H.; Takada, T.; Shuto, T.; Suico, M.A.; Sugiyama, Y.; Kai, H. Calreticulin facilitates the cell surface expression of ABCG5/G8. Biochem. Biophys. Res. Commun. 2006, 347, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Graf, G.A.; Cohen, J.C.; Hobbs, H.H. Missense mutations in ABCG5 and ABCG8 disrupt heterodimerization and trafficking. J. Biol. Chem. 2004, 279, 24881–24888. [Google Scholar] [CrossRef]
- Hirata, T.; Okabe, M.; Kobayashi, A.; Ueda, K.; Matsuo, M. Molecular mechanisms of subcellular localization of ABCG5 and ABCG8. Biosci. Biotechnol. Biochem. 2009, 73, 619–626. [Google Scholar] [CrossRef]
- Suzuki, S.; Shuto, T.; Sato, T.; Kaneko, M.; Takada, T.; Suico, M.A.; Cyr, D.M.; Suzuki, H.; Kai, H. Inhibition of post-translational N-glycosylation by HRD1 that controls the fate of ABCG5/8 transporter. Sci. Rep. 2014, 4, 4258. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, Y.; Yasui, K.; Hashizume, T.; Suto, A.; Mori, A.; Murata, Y.; Yamaguchi, M.; Ikari, A.; Sugatani, J. Involvement of a cyclic adenosine monophosphate-dependent signal in the diet-induced canalicular trafficking of adenosine triphosphate-binding cassette transporter g5/g8. Hepatology 2015, 62, 1215–1226. [Google Scholar] [CrossRef]
- Tachibana, S.; Hirano, M.; Hirata, T.; Matsuo, M.; Ikeda, I.; Ueda, K.; Sato, R. Cholesterol and plant sterol efflux from cultured intestinal epithelial cells is mediated by ATP-binding cassette transporters. Biosci. Biotechnol. Biochem. 2007, 71, 1886–1895. [Google Scholar] [CrossRef] [PubMed]
- Vrins, C.; Vink, E.; Vandenberghe, K.E.; Frijters, R.; Seppen, J.; Groen, A.K. The sterol transporting heterodimer ABCG5/ABCG8 requires bile salts to mediate cholesterol efflux. FEBS Lett. 2007, 581, 4616–4620. [Google Scholar] [CrossRef]
- Johnson, B.J.; Lee, J.Y.; Pickert, A.; Urbatsch, I.L. Bile acids stimulate ATP hydrolysis in the purified cholesterol transporter ABCG5/G8. Biochemistry 2010, 49, 3403–3411. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, D.W.; Lei, Y.; Xu, F.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Purification and reconstitution of sterol transfer by native mouse ABCG5 and ABCG8. Biochemistry 2008, 47, 5194–5204. [Google Scholar] [CrossRef][Green Version]
- Wang, J.; Sun, F.; Zhang, D.W.; Ma, Y.; Xu, F.; Belani, J.D.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Sterol transfer by ABCG5 and ABCG8: In vitro assay and reconstitution. J. Biol. Chem. 2006, 281, 27894–27904. [Google Scholar] [CrossRef]
- Muller, M.; Klein, I.; Kopacsi, S.; Remaley, A.T.; Rajnavolgyi, E.; Sarkadi, B.; Varadi, A. Co-expression of human ABCG5 and ABCG8 in insect cells generates an androstan stimulated membrane ATPase activity. FEBS Lett. 2006, 580, 6139–6144. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zhang, D.W.; Graf, G.A.; Gerard, R.D.; Cohen, J.C.; Hobbs, H.H. Functional asymmetry of nucleotide-binding domains in ABCG5 and ABCG8. J. Biol. Chem. 2006, 281, 4507–4516. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kinch, L.N.; Borek, D.M.; Wang, J.; Wang, J.; Urbatsch, I.L.; Xie, X.S.; Grishin, N.V.; Cohen, J.C.; Otwinowski, Z.; et al. Crystal structure of the human sterol transporter ABCG5/ABCG8. Nature 2016, 533, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Grishin, N.; Kinch, L.; Cohen, J.C.; Hobbs, H.H.; Xie, X.S. Sequences in the nonconsensus nucleotide-binding domain of ABCG5/ABCG8 required for sterol transport. J. Biol. Chem. 2011, 286, 7308–7314. [Google Scholar] [CrossRef]
- Patel, S.B.; Graf, G.A.; Temel, R.E. ABCG5 and ABCG8: More than a defense against xenosterols. J. Lipid Res. 2018, 59, 1103–1113. [Google Scholar] [CrossRef] [PubMed]
- Bastida, J.M.; Benito, R.; Gonzalez-Porras, J.R.; Rivera, J. ABCG5 and ABCG8 gene variations associated with sitosterolemia and platelet dysfunction. Platelets 2020. [Google Scholar] [CrossRef]
- Wang, J.; Joy, T.; Mymin, D.; Frohlich, J.; Hegele, R.A. Phenotypic heterogeneity of sitosterolemia. J. Lipid Res. 2004, 45, 2361–2367. [Google Scholar] [CrossRef]
- Lee, J.H.; Song, D.Y.; Jun, S.H.; Song, S.H.; Shin, C.H.; Ki, C.S.; Lee, K.; Song, J. High prevalence of increased sitosterol levels in hypercholesterolemic children suggest underestimation of sitosterolemia incidence. PLoS ONE 2020, 15, e0238079. [Google Scholar] [CrossRef]
- Tada, H.; Okada, H.; Nomura, A.; Yashiro, S.; Nohara, A.; Ishigaki, Y.; Takamura, M.; Kawashiri, M.A. Rare and Deleterious Mutations in ABCG5/ABCG8 Genes Contribute to Mimicking and Worsening of Familial Hypercholesterolemia Phenotype. Circ. J. 2019, 83, 1917–1924. [Google Scholar] [CrossRef] [PubMed]
- Lek, M.; Karczewski, K.J.; Minikel, E.V.; Samocha, K.E.; Banks, E.; Fennell, T.; O’Donnell-Luria, A.H.; Ware, J.S.; Hill, A.J.; Cummings, B.B.; et al. Analysis of protein-coding genetic variation in 60,706 humans. Nature 2016, 536, 285–291. [Google Scholar] [CrossRef]
- Vauthier, V.; Ben Saad, A.; Elie, J.; Oumata, N.; Durand-Schneider, A.M.; Bruneau, A.; Delaunay, J.L.; Housset, C.; Ait-Slimane, T.; Meijer, L.; et al. Structural analogues of roscovitine rescue the intracellular traffic and the function of ER-retained ABCB4 variants in cell models. Sci. Rep. 2019, 9, 6653. [Google Scholar] [CrossRef]
- Davis, P.B.; Yasothan, U.; Kirkpatrick, P. Ivacaftor. Nat. Rev. Drug Discov. 2012, 11, 349–350. [Google Scholar] [CrossRef] [PubMed]
- Delaunay, J.L.; Durand-Schneider, A.M.; Dossier, C.; Falguieres, T.; Gautherot, J.; Davit-Spraul, A.; Ait-Slimane, T.; Housset, C.; Jacquemin, E.; Maurice, M. A functional classification of ABCB4 variations causing progressive familial intrahepatic cholestasis type 3. Hepatology 2016, 63, 1620–1631. [Google Scholar] [CrossRef]


{kind=link}
| Typical | No Mutation | ABCG5 | ABCG8 |
|---|---|---|---|
| Class I | Nonsense, Frameshift, Deletions | 57 known or predicted | 58 known or predicted |
| Class II | Maturation | R389H, R419P, R419H, N437K | R189H, P231T, R263Q, L501P, G574E, L596R |
| Class III | Activity | ||
| Class IV | Stability | ||
| Class V | No Detectible Defect | ||
| Unclassified | Inconclusive Results | E146Q | R543S, G574R |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, K.; Segard, A.; Graf, G.A. Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. Int. J. Mol. Sci. 2021, 22, 2641. https://doi.org/10.3390/ijms22052641
Williams K, Segard A, Graf GA. Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. International Journal of Molecular Sciences. 2021; 22(5):2641. https://doi.org/10.3390/ijms22052641
Chicago/Turabian StyleWilliams, Kori, Allison Segard, and Gregory A. Graf. 2021. "Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8" International Journal of Molecular Sciences 22, no. 5: 2641. https://doi.org/10.3390/ijms22052641
APA StyleWilliams, K., Segard, A., & Graf, G. A. (2021). Sitosterolemia: Twenty Years of Discovery of the Function of ABCG5ABCG8. International Journal of Molecular Sciences, 22(5), 2641. https://doi.org/10.3390/ijms22052641