
Culturomics Approaches Expand the Diagnostic Accuracy for Sexually Transmitted Infections

 and
and
Abstract
:1. Introduction
2. Importance of Understanding Microbiota in STIs
3. Role of Microbiota-Driven Microenvironmental Conditions in STIs
4. Consequences for STI Culture Diagnostics
5. State-of-the-Art Culture Diagnostics
5.1. Sample Collection and Transportation
5.2. Sample Processing
5.3. Examination and Identification of Bacteria
6. Limitations and Developments in Routine Culture Diagnostics
7. Culturomics as a New Tool to Improve Diagnostic Standards and Treatment Options
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vos, T.; Allen, C.; Arora, M.; Barber, R.M.; Bhutta, Z.A.; Brown, A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; et al. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990–2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar] [CrossRef] [Green Version]
- Gottlieb, S.L.; Low, N.; Newman, L.M.; Bolan, G.; Kamb, M.; Broutet, N. Toward global prevention of sexually transmitted infections (STIs): The need for STI vaccines. Vaccine 2014, 32, 1527–1535. [Google Scholar] [CrossRef]
- Rowley, J.; Vander Hoorn, S.; Korenromp, E.; Low, N.; Unemo, M.; Abu-Raddad, L.J.; Chico, R.M.; Smolak, A.; Newman, L.; Gottlieb, S.; et al. Chlamydia, gonorrhoea, trichomoniasis and syphilis: Global prevalence and incidence estimates, 2016. Bull. World Health Organ. 2019, 97, 548–562P. [Google Scholar] [CrossRef] [PubMed]
- Hayes, R.; Watson-Jones, D.; Celum, C.; Van De Wijgert, J.; Wasserheit, J. Treatment of sexually transmitted infections for HIV prevention: End of the road or new beginning? AIDS 2010, 24, S15–S26. [Google Scholar] [CrossRef] [PubMed]
- Carroll, K.C.; Pfaller, M.A.; Landry, M.L.; McAdam, A.J.; Patel, R.; Richter, S.S.; Warnock, D.W. Manual of Clinical Microbiology, 12th ed.; ASM Press: Washington, DC, USA, 2019; Volume 1. [Google Scholar]
- Van De Laar, M.J.W.; A Morré, S. Chlamydia: A major challenge for public health. Eurosurveillance 2007, 12, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.D.; Gérard, H.C.; Espinoza, L.R.; Ricca, L.R.; Valeriano-Marcet, J.; Snelgrove, J.; Oszust, C.; Vasey, F.B.; Hudson, A.P. Chlamydiae as etiologic agents in chronic undifferentiated spondylarthritis. Arthritis Rheum. 2009, 60, 1311–1316. [Google Scholar] [CrossRef] [Green Version]
- Aral, S. Determinants of STD epidemics: Implications for phase appropriate intervention strategies. Sex. Transm. Infect. 2002, 78, i3–i13. [Google Scholar] [CrossRef]
- Stewart, E.J. Growing Unculturable Bacteria. J. Bacteriol. 2012, 194, 4151–4160. [Google Scholar] [CrossRef] [Green Version]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef] [Green Version]
- Brotman, R.M.; Klebanoff, M.A.; Nansel, T.; Yu, K.F.; Andrews, W.W.; Zhang, J.; Schwebke, J.R. Bacterial Vaginosis Assessed by Gram Stain and Diminished Colonization Resistance to Incident Gonococcal, Chlamydial, and Trichomonal Genital Infection. J. Infect. Dis. 2010, 202, 1907–1915. [Google Scholar] [CrossRef]
- Cherpes, T.L.; Meyn, L.A.; Krohn, M.A.; Lurie, J.G.; Hillier, S.L. Association between Acquisition of Herpes Simplex Virus Type 2 in Women and Bacterial Vaginosis. Clin. Infect. Dis. 2003, 37, 319–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, J.H.L.; Richardson, B.A.; Nyange, P.M.; Lavreys, L.; Hillier, S.L.; Chohan, B.; Mandaliya, K.; Ndinya-Achola, J.O.; Bwayo, J.; Kreiss, J. Vaginal Lactobacilli, Microbial Flora, and Risk of Human Immunodeficiency Virus Type 1 and Sexually Transmitted Disease Acquisition. J. Infect. Dis. 1999, 180, 1863–1868. [Google Scholar] [CrossRef]
- Wiesenfeld, H.C.; Hillier, S.L.; Krohn, M.A.; Landers, D.V.; Sweet, R.L. Bacterial Vaginosis Is a Strong Predictor of Neisseria gonorrhoeae and Chlamydia trachomatis Infection. Clin. Infect. Dis. 2003, 36, 663–668. [Google Scholar] [CrossRef] [Green Version]
- Brotman, R.M. Vaginal microbiome and sexually transmitted infections: An epidemiologic perspective. J. Clin. Investig. 2011, 121, 4610–4617. [Google Scholar] [CrossRef]
- Donders, G.G.; Van Calsteren, K.; Bellen, G.; Reybrouck, R.; Bosch, T.V.D.; Riphagen, I.; Van Lierde, S. Predictive value for preterm birth of abnormal vaginal flora, bacterial vaginosis and aerobic vaginitis during the first trimester of pregnancy. BJOG Int. J. Obstet. Gynaecol. 2009, 116, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Gupta, K.; Stapleton, A.; Hooton, T.M.; Roberts, P.L.; Fennell, C.L.; Stamm, W.E. Inverse Association of H2O2-Producing Lactobacilli and Vaginal Escherichia coli Colonization in Women with Recurrent Urinary Tract Infections. J. Infect. Dis. 1998, 178, 446–450. [Google Scholar] [CrossRef] [Green Version]
- Ness, R.B.; Kip, K.E.; Hillier, S.L.; Soper, D.E.; Stamm, C.A.; Sweet, R.L.; Rice, P.; Richter, H.E. A Cluster Analysis of Bacterial Vaginosis–associated Microflora and Pelvic Inflammatory Disease. Am. J. Epidemiol. 2005, 162, 585–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Wijgert, J.H.H.M.; Mason, P.R.; Gwanzura, L.; Mbizvo, M.T.; Chirenje, Z.M.; Iliff, V.; Shiboski, S.; Padian, N.S. Intravaginal Practices, Vaginal Flora Disturbances, and Acquisition of Sexually Transmitted Diseases in Zimbabwean Women. J. Infect. Dis. 2000, 181, 587–594. [Google Scholar] [CrossRef]
- Gajer, P.; Brotman, R.; Bai, G.; Sakamoto, J.; Schütte, U.M.E.; Zhong, X.; Koenig, S.S.K.; Fu, L.; Ma, Z.S.; Zhou, X.; et al. Temporal Dynamics of the Human Vaginal Microbiota. Sci. Transl. Med. 2012, 4, 132ra52. [Google Scholar] [CrossRef] [Green Version]
- France, M.T.; Ma, B.; Gajer, P.; Brown, S.; Humphrys, M.S.; Holm, J.B.; Waetjen, L.E.; Brotman, R.M.; Ravel, J. VALENCIA: A nearest centroid classification method for vaginal microbial communities based on composition. Microbiome 2020, 8, 1–15. [Google Scholar] [CrossRef]
- Graspeuntner, S.; Bohlmann, M.K.; Gillmann, K.; Speer, R.; Kuenzel, S.; Mark, H.; Hoellen, F.; Lettau, R.; Griesinger, G.; König, I.; et al. Microbiota-based analysis reveals specific bacterial traits and a novel strategy for the diagnosis of infectious infertility. PLoS ONE 2018, 13, e0191047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graspeuntner, S.; Loeper, N.; Künzel, S.; Baines, J.F.; Rupp, J. Selection of validated hypervariable regions is crucial in 16S-based microbiota studies of the female genital tract. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Loeper, N.; Graspeuntner, S.; Rupp, J. Microbiota changes impact on sexually transmitted infections and the development of pelvic inflammatory disease. Microbes Infect. 2018, 20, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.C.; McAndrew, T.; Chen, Z.; Harari, A.; Barris, D.M.; Viswanathan, S.; Rodriguez, A.C.; Castle, P.; Herrero, R.; Schiffman, M.; et al. The Cervical Microbiome over 7 Years and a Comparison of Methodologies for Its Characterization. PLoS ONE 2012, 7, e40425. [Google Scholar] [CrossRef]
- O’Hanlon, D.E.; Moench, T.R.; Cone, R.A. Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PLoS ONE 2013, 8, e80074. [Google Scholar] [CrossRef]
- Gong, Z.; Luna, Y.; Yu, P.; Fan, H. Lactobacilli Inactivate Chlamydia trachomatis through Lactic Acid but Not H2O2. PLoS ONE 2014, 9, e107758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, M.I.; Lievens, E.; Malik, S.; Imholz, N.; Lebeer, S. Lactobacillus species as biomarkers and agents that can promote various aspects of vaginal health. Front. Physiol. 2015, 6, 81. [Google Scholar] [CrossRef] [Green Version]
- Nardini, P.; Palomino, R.A.N.; Parolin, C.; Laghi, L.; Foschi, C.; Cevenini, R.; Vitali, B.; Marangoni, A. Lactobacillus crispatus inhibits the infectivity of Chlamydia trachomatis elementary bodies, in vitro study. Sci. Rep. 2016, 6, 29024. [Google Scholar] [CrossRef]
- Witkin, S.S.; Mendes-Soares, H.; Linhares, I.; Jayaram, A.; Ledger, W.J.; Forney, L.J. Influence of Vaginal Bacteria and d- and l-Lactic Acid Isomers on Vaginal Extracellular Matrix Metalloproteinase Inducer: Implications for Protection against Upper Genital Tract Infections. mBio 2013, 4, 10112800460. [Google Scholar] [CrossRef] [Green Version]
- Rahkonen, L.; Rutanen, E.-M.; Unkila-Kallio, L.; Nuutila, M.; Nieminen, P.; Sorsa, T.; Paavonen, J. Factors affecting matrix metalloproteinase-8 levels in the vaginal and cervical fluids in the first and second trimester of pregnancy. Hum. Reprod. 2009, 24, 2693–2702. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, A.; Losacco, A.; Carratelli, C.R. Lactobacillus crispatus modulates epithelial cell defense against Candida albicans through Toll-like receptors 2 and 4, interleukin 8 and human β-defensins 2 and 3. Immunol. Lett. 2013, 156, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the Vaginal Microbiome Alter the Innate Immune Response and Barrier Properties of the Human Vaginal Epithelia in a Species-Specific Manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzo, A.; Fiorentino, M.; Buommino, E.; Donnarumma, G.; Losacco, A.; Bevilacqua, N. Lactobacillus crispatus mediates anti-inflammatory cytokine interleukin-10 induction in response to Chlamydia trachomatis infection in vitro. Int. J. Med. Microbiol. 2015, 305, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zang, Y.; Wang, C.; Li, H.; Fan, A.; Han, C.; Xue, F. The Interaction Between Microorganisms, Metabolites, and Immune System in the Female Genital Tract Microenvironment. Front. Cell. Infect. Microbiol. 2020, 10, 609488. [Google Scholar] [CrossRef]
- Ziklo, N.; Huston, W.M.; Taing, K.; Katouli, M.; Timms, P. In vitro rescue of genital strains of Chlamydia trachomatis from interferon-γ and tryptophan depletion with indole-positive, but not indole-negative Prevotella spp. BMC Microbiol. 2016, 16, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ziklo, N.; Vidgen, M.E.; Taing, K.; Huston, W.; Timms, P. Dysbiosis of the Vaginal Microbiota and Higher Vaginal Kynurenine/Tryptophan Ratio Reveals an Association with Chlamydia trachomatis Genital Infections. Front. Cell. Infect. Microbiol. 2018, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Hoffman, N.G.; Morgan, M.; Matsen, F.A.; Fiedler, T.L.; Hall, R.W.; Ross, F.J.; McCoy, C.O.; Bumgarner, R.; Marrazzo, J.; et al. Bacterial Communities in Women with Bacterial Vaginosis: High Resolution Phylogenetic Analyses Reveal Relationships of Microbiota to Clinical Criteria. PLoS ONE 2012, 7, e37818. [Google Scholar] [CrossRef] [Green Version]
- France, M.T.; Mendes-Soares, H.; Forney, L.J. Genomic Comparisons of Lactobacillus crispatus and Lactobacillus iners Reveal Potential Ecological Drivers of Community Composition in the Vagina. Appl. Environ. Microbiol. 2016, 82, 7063–7073. [Google Scholar] [CrossRef] [Green Version]
- Mastromarino, P.; Di Pietro, M.; Schiavoni, G.; Nardis, C.; Gentile, M.; Sessa, R. Effects of vaginal lactobacilli in Chlamydia trachomatis infection. Int. J. Med. Microbiol. 2014, 304, 654–661. [Google Scholar] [CrossRef]
- Spurbeck, R.R.; Arvidson, C.G. Inhibition of Neisseria gonorrhoeae Epithelial Cell Interactions by Vaginal Lactobacillus Species. Infect. Immun. 2008, 76, 3124–3130. [Google Scholar] [CrossRef] [Green Version]
- Nagy, E.; Boyanova, L.; Justesen, U.S. How to isolate, identify and determine antimicrobial susceptibility of anaerobic bacteria in routine laboratories. Clin. Microbiol. Infect. 2018, 24, 1139–1148. [Google Scholar] [CrossRef] [Green Version]
- Arbique, J.C.; Forward, K.R.; Leblanc, J. Evaluation of four commercial transport media for the survival of Neisseria Gonorrhoeae. Diagn. Microbiol. Infect. Dis. 2000, 36, 163–168. [Google Scholar] [CrossRef]
- Drake, C.; Barenfanger, J.; Lawhorn, J.; Verhulst, S. Comparison of Easy-Flow Copan Liquid Stuart’s and Starplex Swab Transport Systems for Recovery of Fastidious Aerobic Bacteria. J. Clin. Microbiol. 2005, 43, 1301–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, N.; Contreras, A.; Flynn, J.; Morrison, J.; Slots, J. Proficiencies of three anaerobic culture systems for recov-ering periodontal pathogenic bacteria. J. Clin. Microbiol. 1999, 37, 171–174. [Google Scholar] [CrossRef] [Green Version]
- Baron, E.J.; Citron, D.M. Anaerobic Identification Flowchart Using Minimal Laboratory Resources. Clin. Infect. Dis. 1997, 25, S143–S146. [Google Scholar] [CrossRef]
- Lee, E.H.L.; Degener, J.E.; Welling, G.W.; Veloo, A.C.M. Evaluation of the Vitek 2 ANC Card for Identification of Clinical Isolates of Anaerobic Bacteria. J. Clin. Microbiol. 2011, 49, 1745–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justesen, U.S.; Skov, M.N.; Knudsen, E.; Holt, H.M.; Søgaard, P.; Justesen, T. 16S rRNA Gene Sequencing in Routine Identification of Anaerobic Bacteria Isolated from Blood Cultures. J. Clin. Microbiol. 2010, 48, 946–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ligozzi, M.; Bernini, C.; Bonora, M.G.; de Fatima, M.; Zuliani, J.; Fontana, R. Evaluation of the VITEK 2 System for Identification and Antimicrobial Susceptibility Testing of Medically Relevant Gram-Positive Cocci. J. Clin. Microbiol. 2002, 40, 1681–1686. [Google Scholar] [CrossRef] [Green Version]
- Lévesque, S.; Dufresne, P.J.; Soualhine, H.; Domingo, M.-C.; Bekal, S.; Lefebvre, B.; Tremblay, C. A Side by Side Comparison of Bruker Biotyper and VITEK MS: Utility of MALDI-TOF MS Technology for Microorganism Identification in a Public Health Reference Laboratory. PLoS ONE 2015, 10, e0144878. [Google Scholar] [CrossRef]
- Singhal, N.; Kumar, M.; Kanaujia, P.K.; Virdi, J.S. MALDI-TOF mass spectrometry: An emerging technology for microbial identification and diagnosis. Front. Microbiol. 2015, 6, 791. [Google Scholar] [CrossRef] [Green Version]
- Fredericks, D.N.; Relman, D.A. Sequence-based identification of microbial pathogens: A reconsideration of Koch’s postulates. Clin. Microbiol. Rev. 1996, 9, 18–33. [Google Scholar] [CrossRef]
- Govender, K.N.; Street, T.L.; Sanderson, N.D.; Eyre, D.W. Metagenomic Sequencing as a Pathogen-Agnostic Clinical Diagnostic Tool for Infectious Diseases: A Systematic Review and Meta-analysis of Diagnostic Test Accuracy Studies. J. Clin. Microbiol. 2021, 59, 910112802916. [Google Scholar] [CrossRef]
- Citron, D.M.; Warren, Y.A.; Hudspeth, M.K.; Goldstein, E.J.C. Survival of Aerobic and Anaerobic Bacteria in Purulent Clinical Specimens Maintained in the Copan Venturi Transystem and Becton Dickinson Port-a-Cul Transport Systems. J. Clin. Microbiol. 2000, 38, 892–894. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMarco, A.L.; Rabe, L.K.; Austin, M.N.; Stoner, K.A.; Avolia, H.A.; Meyn, L.A.; Hillier, S.L. Survival of vaginal microorganisms in three commercially available transport systems. Anaerobe 2017, 45, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Demuyser, T.; De Geyter, D.; Van Dorpe, D.; Vandoorslaer, K.; Wybo, I. Extensive evaluation of fastidious anaerobic bacteria recovery from the Copan eSwab® transport system. J. Microbiol. Methods 2018, 144, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Nys, S.; Vijgen, S.; Magerman, K.; Cartuyvels, R. Comparison of Copan eSwab with the Copan Venturi Transystem for the quantitative survival of Escherichia coli, Streptococcus agalactiae and Candida albicans. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 453–456. [Google Scholar] [CrossRef]
- Rishmawi, N.; Ghneim, R.; Kattan, R.; Ghneim, R.; Zoughbi, M.; Abu-Diab, A.; Turkuman, S.; Dauodi, R.; Shomali, I.; Issa, A.E.-R.; et al. Survival of Fastidious and Nonfastidious Aerobic Bacteria in Three Bacterial Transport Swab Systems. J. Clin. Microbiol. 2007, 45, 1278–1283. [Google Scholar] [CrossRef] [Green Version]
- Stoner, K.A.; Rabe, L.K.; Austin, M.N.; Meyn, L.A.; Hillier, S.L. Quantitative Survival of Aerobic and Anaerobic Microorganisms in Port-A-Cul and Copan Transport Systems. J. Clin. Microbiol. 2008, 46, 2739–2744. [Google Scholar] [CrossRef] [Green Version]
- Tyrrell, K.L.; Citron, D.M.; Leoncio, E.S.; Goldstein, E.J.C. Comparison of the Copan eSwab System with an Agar Swab Transport System for Maintenance of Fastidious Anaerobic Bacterium Viability. J. Clin. Microbiol. 2016, 54, 1364–1367. [Google Scholar] [CrossRef] [Green Version]
- Van Horn, K.G.; Audette, C.D.; Tucker, K.A.; Sebeck, D. Comparison of 3 swab transport systems for direct release and recovery of aerobic and anaerobic bacteria. Diagn. Microbiol. Infect. Dis. 2008, 62, 471–473. [Google Scholar] [CrossRef]
- Jamal, W.Y.; Shahin, M.; Rotimi, V.O. Comparison of two matrix-assisted laser desorption/ionization-time of flight (MALDI-TOF) mass spectrometry methods and API 20AN for identification of clinically relevant anaerobic bacteria. J. Med. Microbiol. 2013, 62, 540–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veloo, A.; Elgersma, P.; Friedrich, A.; Nagy, E.; van Winkelhoff, A. The influence of incubation time, sample preparation and exposure to oxygen on the quality of the MALDI-TOF MS spectrum of anaerobic bacteria. Clin. Microbiol. Infect. 2014, 20, O1091–O1097. [Google Scholar] [CrossRef] [Green Version]
- Coltella, L.; Mancinelli, L.; Onori, M.; Lucignano, B.; Menichella, D.; Sorge, R.; Raponi, M.; Mancini, R.; Russo, C. Advancement in the routine identification of anaerobic bacteria by MALDI-TOF mass spectrometry. Eur. J. Clin. Microbiol. Infect. Dis. 2013, 32, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Fettweis, J.M.; Serrano, M.G.; Sheth, N.U.; Mayer, C.M.; Glascock, A.L.; Brooks, J.P.; Jefferson, K.K.; Buck, G.A.; Vaginal Microbiome Consortium (additional members). Species-level classification of the vaginal microbiome. BMC Genom. 2012, 13, S17. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-M.S.; Burnham, C.-A.D. MALDI-TOF MS identification of anaerobic bacteria: Assessment of pre-analytical variables and specimen preparation techniques. Diagn. Microbiol. Infect. Dis. 2014, 79, 144–148. [Google Scholar] [CrossRef]
- Jensen, A.; Fagö-Olsen, H.; Sørensen, C.H.; Kilian, M. Molecular Mapping to Species Level of the Tonsillar Crypt Microbiota Associated with Health and Recurrent Tonsillitis. PLoS ONE 2013, 8, e56418. [Google Scholar] [CrossRef] [Green Version]
- Lagier, J.-C.; Dubourg, G.; Million, M.; Cadoret, F.; Bilen, M.; Fenollar, F.; Levasseur, A.; Rolain, J.-M.; Fournier, P.-E.; Raoult, D. Culturing the human microbiota and culturomics. Nat. Rev. Microbiol. 2018, 16, 540–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Tun, H.M.; Jahan, M.; Zhang, Z.; Kumar, A.; Fernando, W.G.D.; Farenhorst, A.; Khafipour, E. Comparison of DNA-, PMA-, and RNA-based 16S rRNA Illumina sequencing for detection of live bacteria in water. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Nagy, E.; Becker, S.; Kostrzewa, M.; Barta, N.; Urbán, E. The value of MALDI-TOF MS for the identification of clinically relevant anaerobic bacteria in routine laboratories. J. Med. Microbiol. 2012, 61, 1393–1400. [Google Scholar] [CrossRef] [Green Version]
- Ling, Z.; Kong, J.; Liu, F.; Zhu, H.; Chen, X.; Wang, Y.; Li, L.; E Nelson, K.; Xia, Y.; Xiang, C. Molecular analysis of the diversity of vaginal microbiota associated with bacterial vaginosis. BMC Genom. 2010, 11, 488. [Google Scholar] [CrossRef] [Green Version]
- Shipitsyna, E.; Roos, A.; Datcu, R.; Hallén, A.; Fredlund, H.; Jensen, J.S.; Engstrand, L.; Unemo, M. Composition of the Vaginal Microbiota in Women of Reproductive Age—Sensitive and Specific Molecular Diagnosis of Bacterial Vaginosis Is Possible? PLoS ONE 2013, 8, e60670. [Google Scholar] [CrossRef] [Green Version]
- Browne, H.P.; Forster, S.C.; Anonye, B.O.; Kumar, N.; Neville, B.A.; Stares, M.D.; Goulding, D.; Lawley, T.D. Culturing of ‘unculturable’ human microbiota reveals novel taxa and extensive sporulation. Nature 2016, 533, 543–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greub, G. Culturomics: A new approach to study the human microbiome. Clin. Microbiol. Infect. 2012, 18, 1157–1159. [Google Scholar] [CrossRef] [Green Version]
- Diop, A.; Diop, K.; Tomei, E.; Armstrong, N.; Bretelle, F.; Raoult, D.; Fenollar, F.; Fournier, P.-E. Collinsella vaginalis sp. nov. strain Marseille-P2666T, a new member of the Collinsella genus isolated from the genital tract of a patient suffering from bacterial vaginosis. Int. J. Syst. Evol. Microbiol. 2019, 69, 949–956. [Google Scholar] [CrossRef]
- Klein, S.; Nurjadi, D.; Horner, S.; Heeg, K.; Zimmermann, S.; Burckhardt, I. Significant increase in cultivation of Gardnerella vaginalis, Alloscardovia omnicolens, Actinotignum schaalii, and Actinomyces spp. in urine samples with total laboratory automation. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1305–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, I.; Jerchel, S.; Szaszák, M.; Shima, K.; Rupp, J. When oxygen runs short: The microenvironment drives host–pathogen interactions. Microbes Infect. 2012, 14, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Dione, N.; Khelaifia, S.; La Scola, B.; Lagier, J.; Raoult, D. A quasi-universal medium to break the aerobic/anaerobic bacterial culture dichotomy in clinical microbiology. Clin. Microbiol. Infect. 2016, 22, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weimann, A.; Mooren, K.; Frank, J.; Pope, P.B.; Bremges, A.; McHardy, A.C. From Genomes to Phenotypes: Traitar, the Microbial Trait Analyzer. mSystems 2016, 1, e00101-16. [Google Scholar] [CrossRef] [Green Version]
- Holm, J.B.; France, M.T.; Ma, B.; McComb, E.; Robinson, C.K.; Mehta, A.; Tallon, L.J.; Brotman, R.M.; Ravel, J. Comparative Metagenome-Assembled Genome Analysis of “Candidatus Lachnocurva vaginae”, Formerly Known as Bacterial Vaginosis-Associated Bacterium−1 (BVAB1). Front. Cell. Infect. Microbiol. 2020, 10, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burckhardt, I. Laboratory Automation in Clinical Microbiology. Bioengineering 2018, 5, 102. [Google Scholar] [CrossRef] [Green Version]
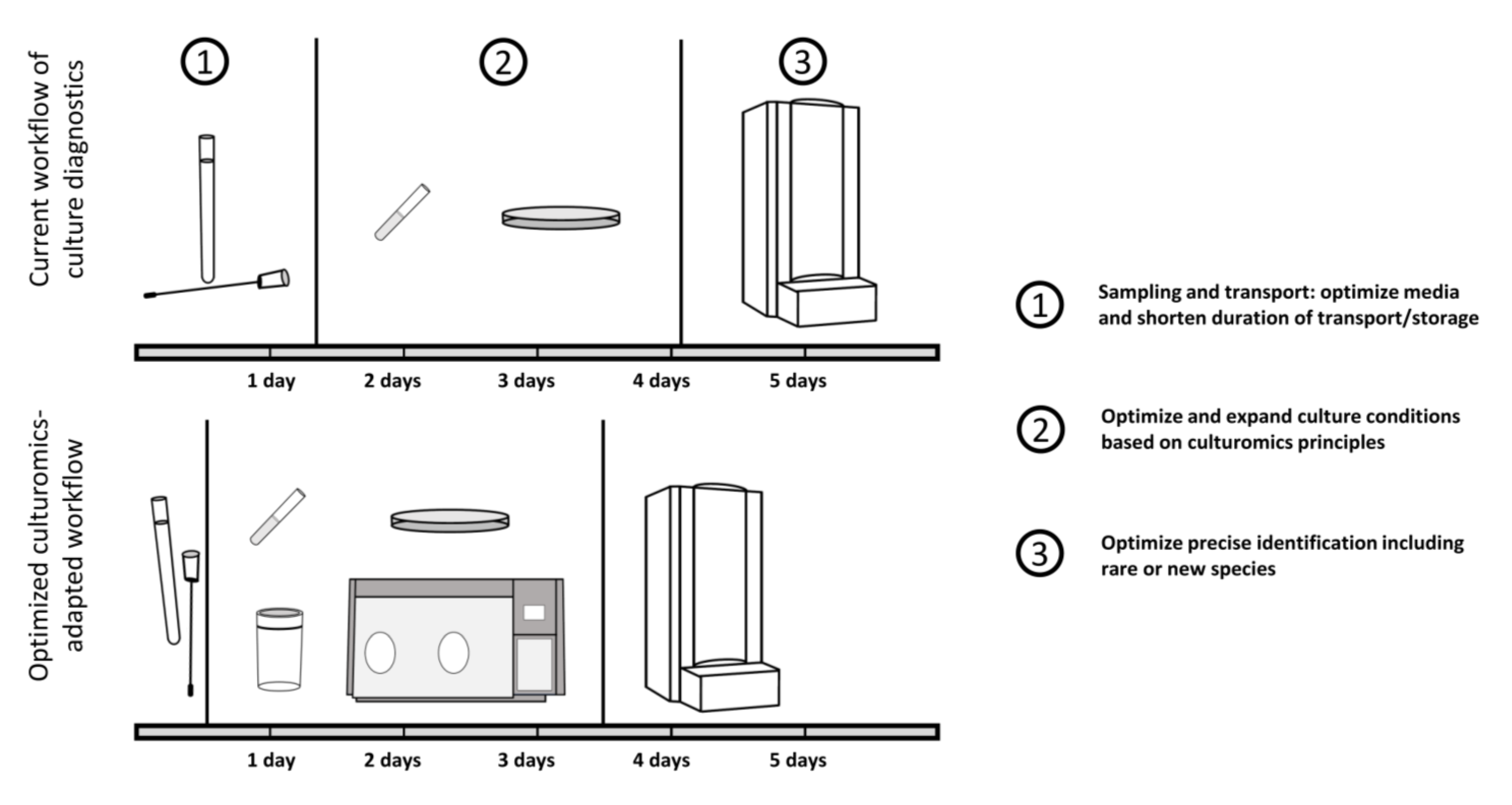

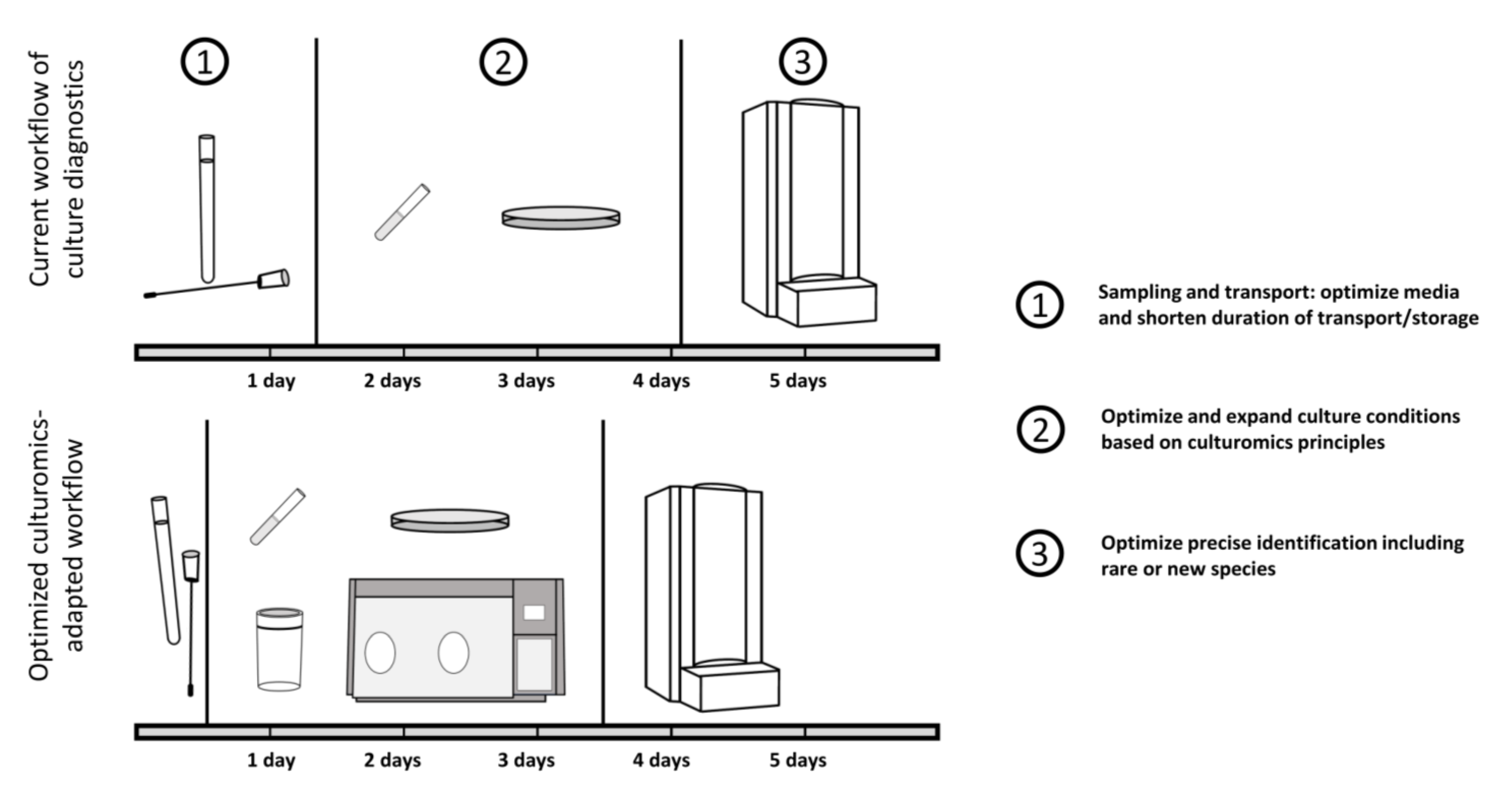{kind=link}
| Genital Tract Infection | Usual Bacterial Etiologies |
|---|---|
| Ulcers | Haemophilus ducreyi, Chlamydia trachomatis (Lymphogranuloma venereum), Treponema pallidum, Klebsiella granulomatis |
| Vulvovaginitis, cervicitis | Neisseria gonorrhoeae, C. trachomatis |
| Bacterial vaginosis (BV) | Overgrowth of vaginal microbiota with anaerobic and facultative endogenous bacteria |
| Endometritis | Enterobacteriaceae, streptococci (groups A and B), enterococci, mixed anaerobic microbes |
| Salpingitis, oophoritis | N. gonorrhoeae, C. trachomatis, mixed aerobic and anaerobic microbiota |
| Pelvic abscess following infection | Mixed aerobic and anaerobic microbes |
| Identification Method | Strengths | Weaknesses |
|---|---|---|
| Morphology (colony and Gram-stain) | very low cost and time investment | identification only possible for some species examiner-dependent |
| Biochemical spot tests | very low cost and time investment | low reliability for anaerobes |
| Antibiotic susceptibility testing | important information for clinical decision making | takes 24 to 48 h low reliability for anaerobes |
| Automated systems such as VITEK® 2 (see also Box 1) | automated identification plus antimicrobial testing | high costs |
| MALDI-TOF MS (see also Box 1) | early and reliable identification | initially high cost of instrument sample preparation influences spectrum quality |
| Partial 16S rRNA gene sequencing analysis (see also Box 1) | reliable identification | time and costs |
| Metagenomics/next-generation sequencing (see also Box 1) | no bacterial growth necessary discovery of new, uncultured taxa association of microbial signatures with diseases | time and costs limited discrimination between live bacteria and transient DNA results depend on used methodologies |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolf, E.A.; Rettig, H.C.; Lupatsii, M.; Schlüter, B.; Schäfer, K.; Friedrich, D.; Graspeuntner, S.; Rupp, J. Culturomics Approaches Expand the Diagnostic Accuracy for Sexually Transmitted Infections. Int. J. Mol. Sci. 2021, 22, 10815. https://doi.org/10.3390/ijms221910815
Wolf EA, Rettig HC, Lupatsii M, Schlüter B, Schäfer K, Friedrich D, Graspeuntner S, Rupp J. Culturomics Approaches Expand the Diagnostic Accuracy for Sexually Transmitted Infections. International Journal of Molecular Sciences. 2021; 22(19):10815. https://doi.org/10.3390/ijms221910815
Chicago/Turabian StyleWolf, Ellinor Anna, Hannah Clara Rettig, Mariia Lupatsii, Britta Schlüter, Kathrin Schäfer, Dirk Friedrich, Simon Graspeuntner, and Jan Rupp. 2021. "Culturomics Approaches Expand the Diagnostic Accuracy for Sexually Transmitted Infections" International Journal of Molecular Sciences 22, no. 19: 10815. https://doi.org/10.3390/ijms221910815
APA StyleWolf, E. A., Rettig, H. C., Lupatsii, M., Schlüter, B., Schäfer, K., Friedrich, D., Graspeuntner, S., & Rupp, J. (2021). Culturomics Approaches Expand the Diagnostic Accuracy for Sexually Transmitted Infections. International Journal of Molecular Sciences, 22(19), 10815. https://doi.org/10.3390/ijms221910815