
Reactions with Proteins of Three Novel Anticancer Platinum(II) Complexes Bearing N-Heterocyclic Ligands

 ,
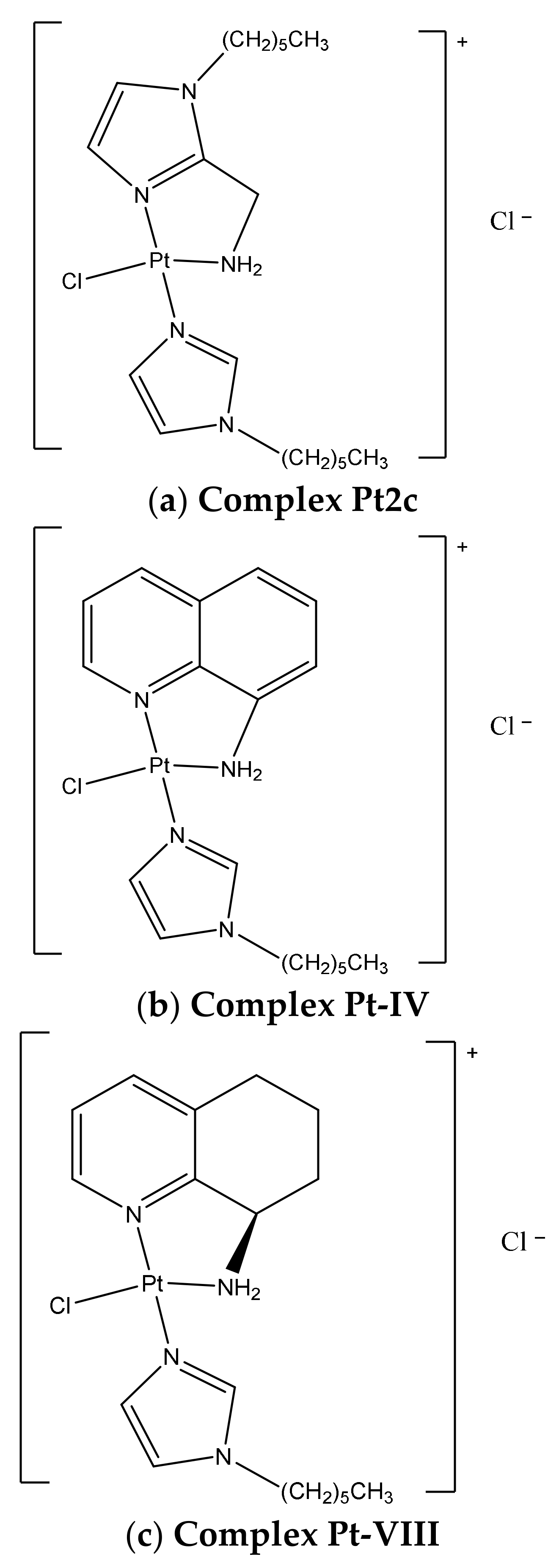
,  ,
,  ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
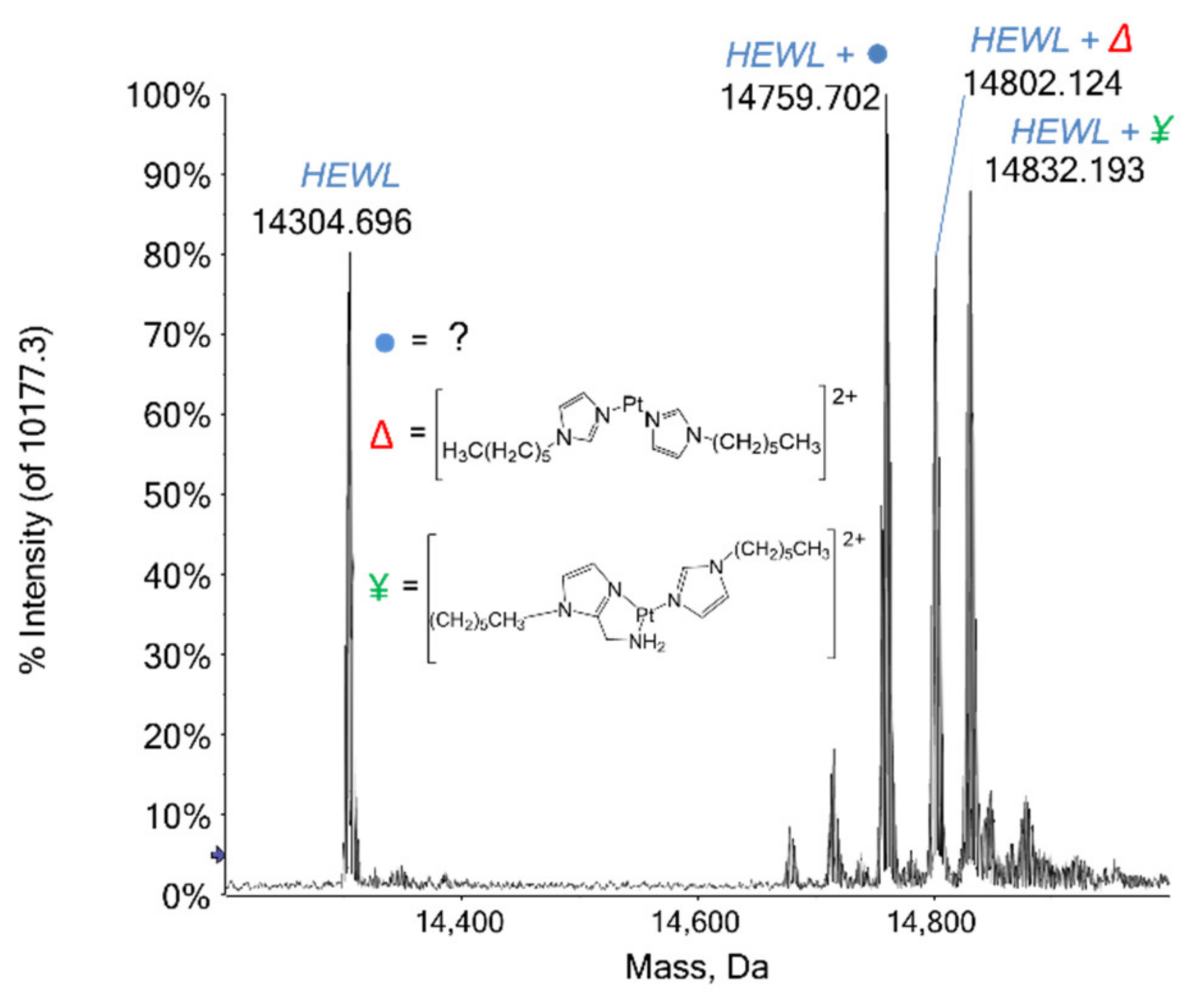
2.1. ESI MS Results
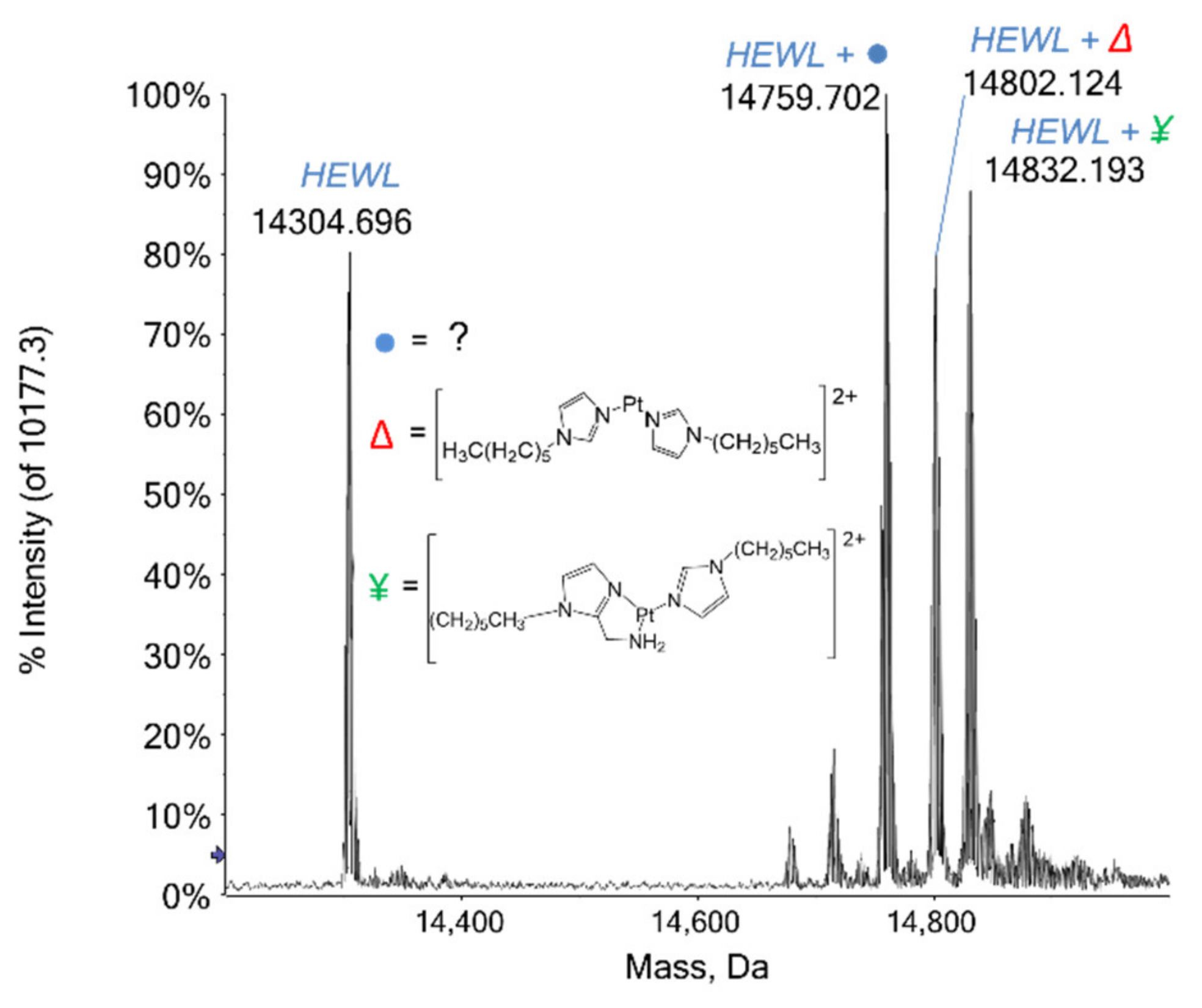
2.1.1. HEWL
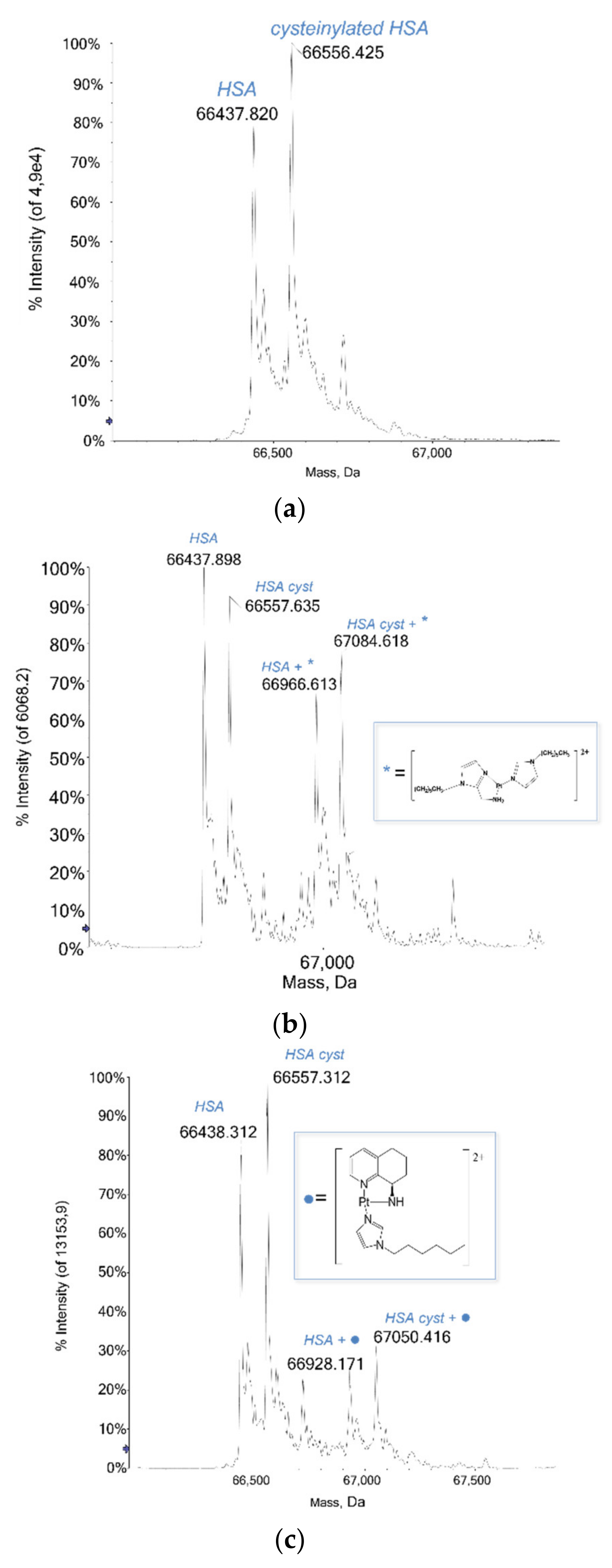
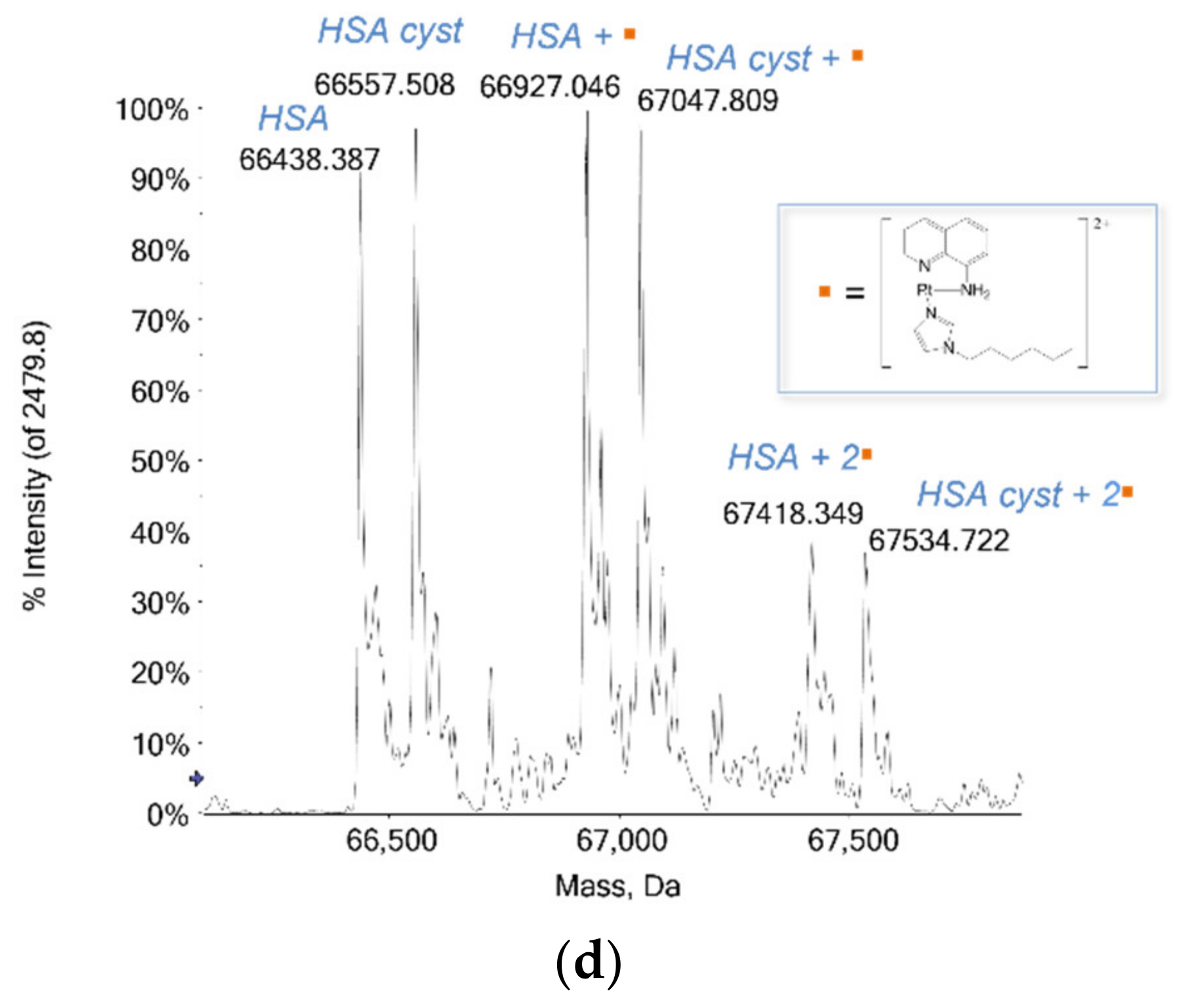
2.1.2. HSA
2.1.3. Cytochrome c
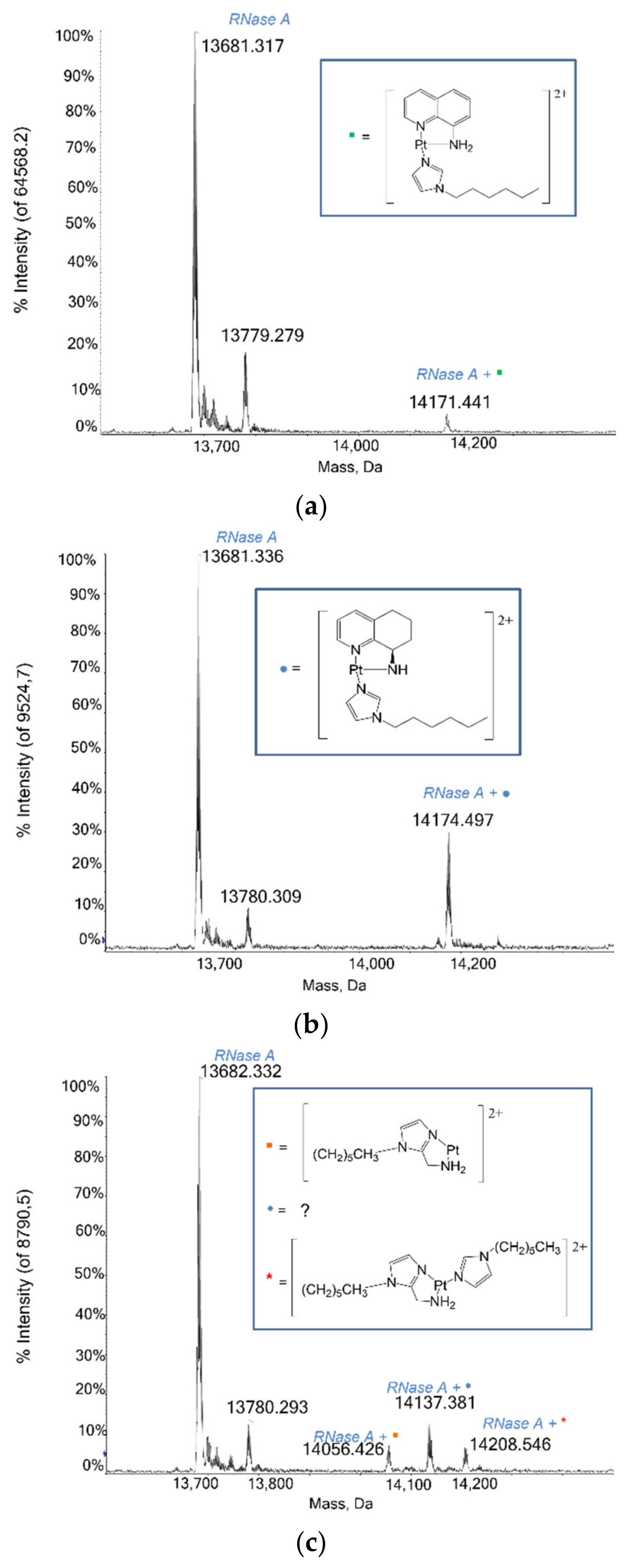
2.1.4. RNase A
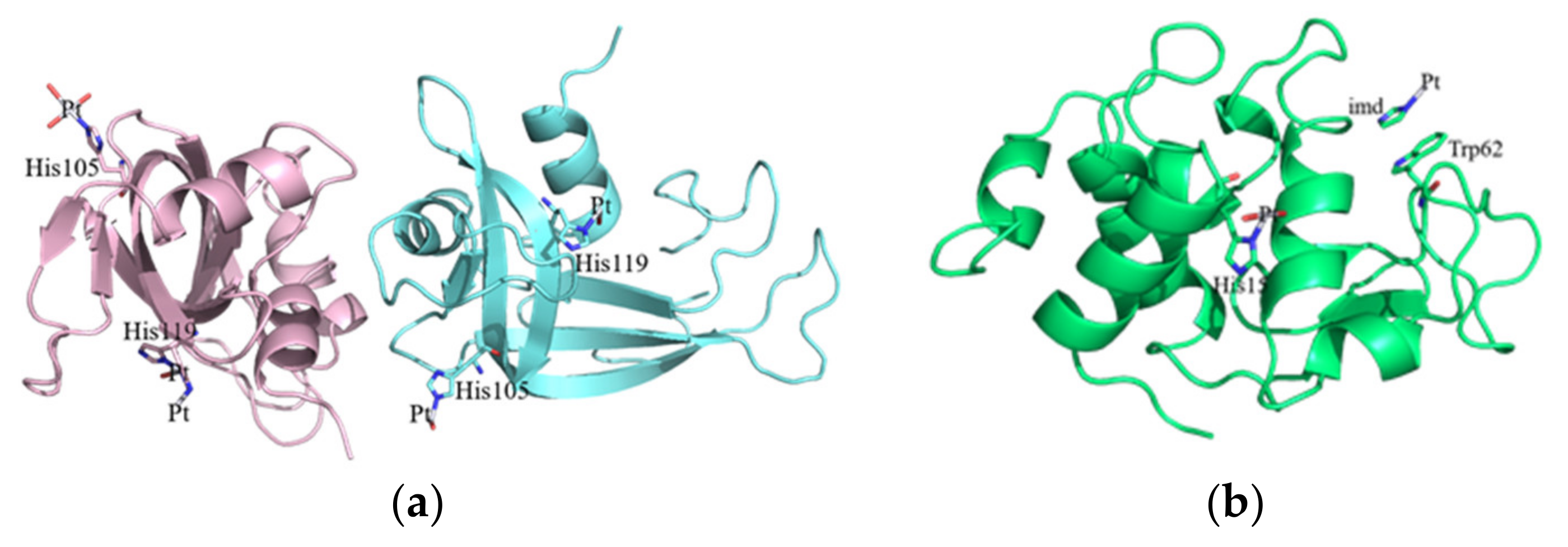
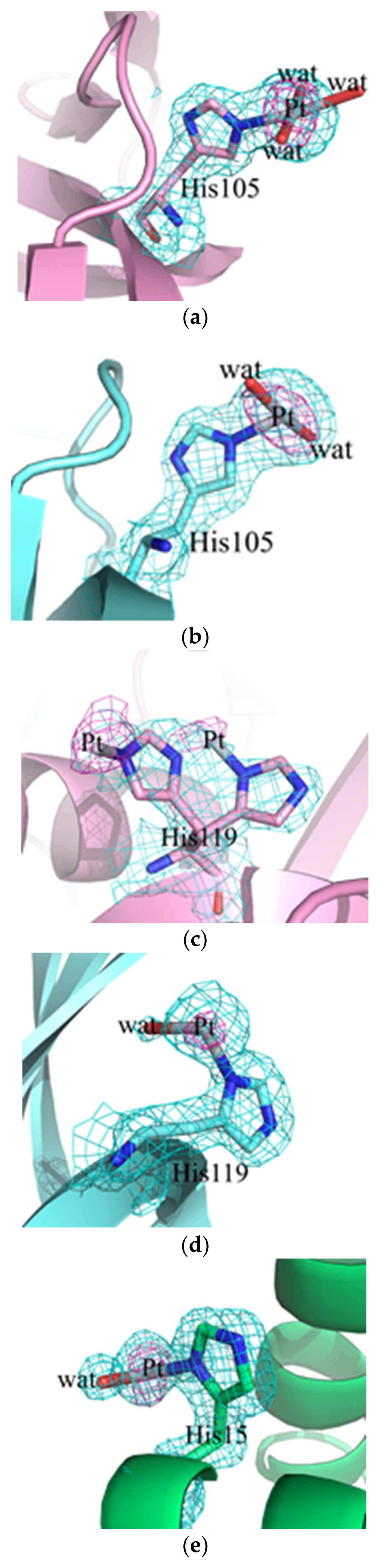
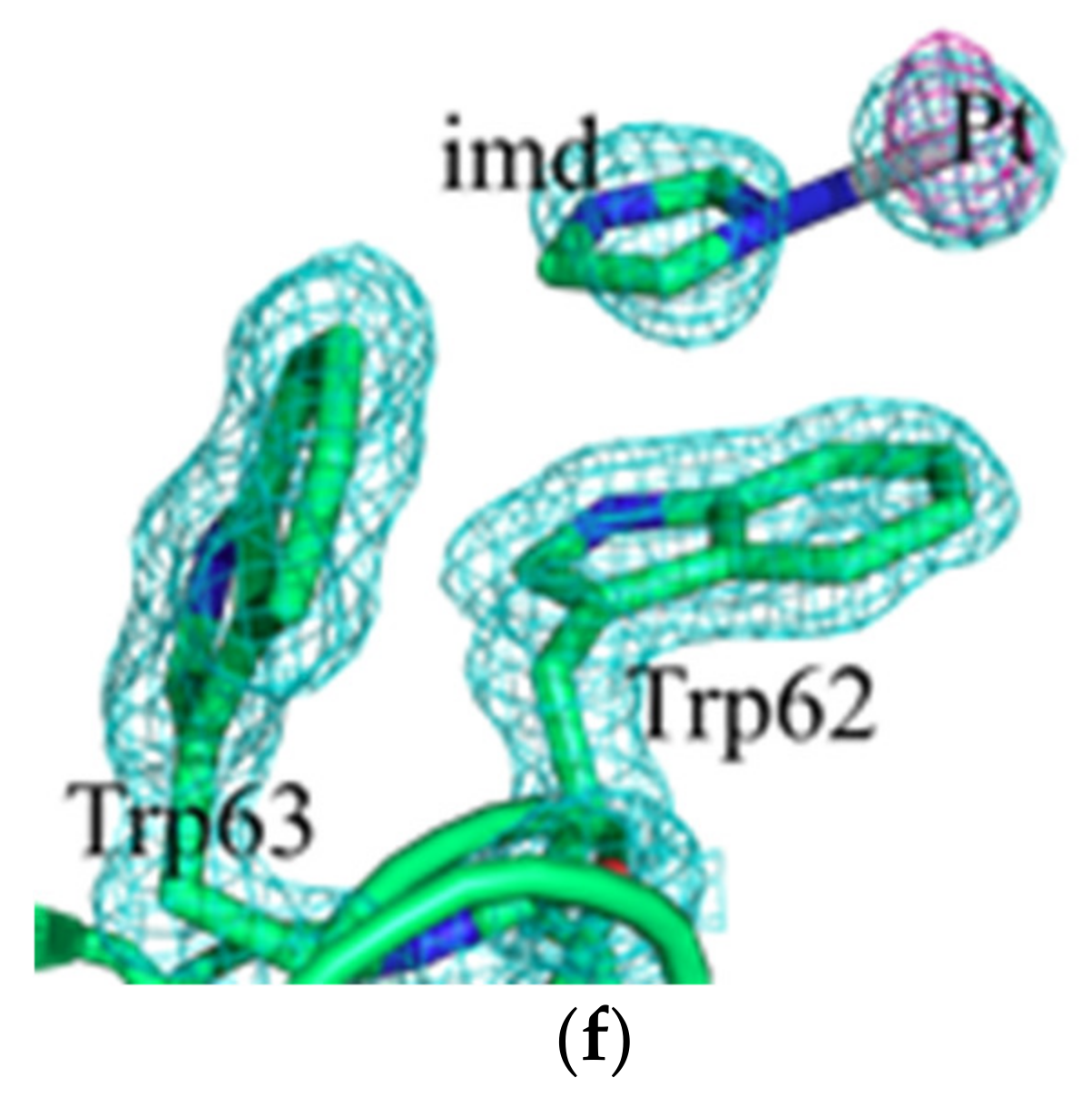
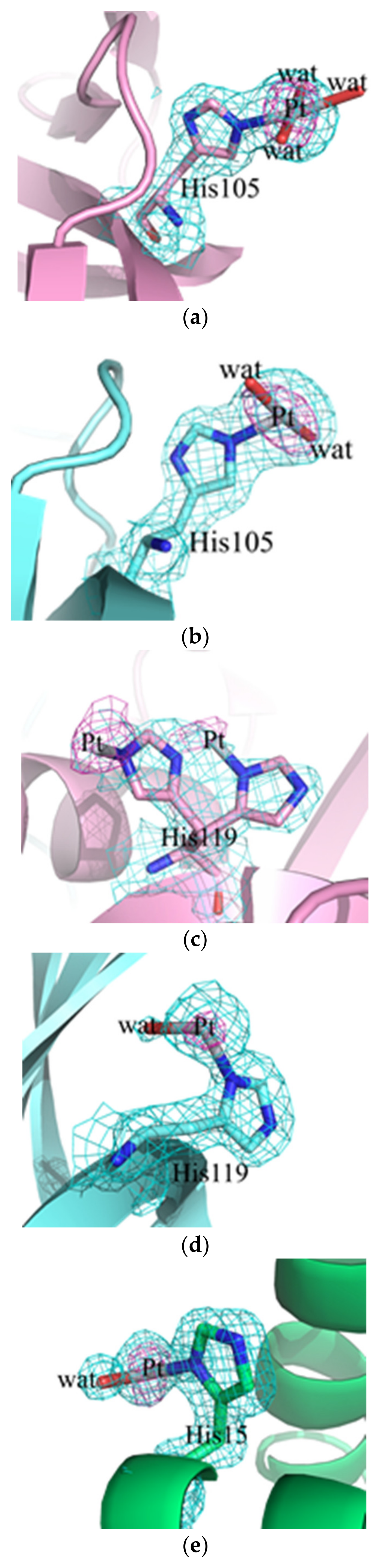
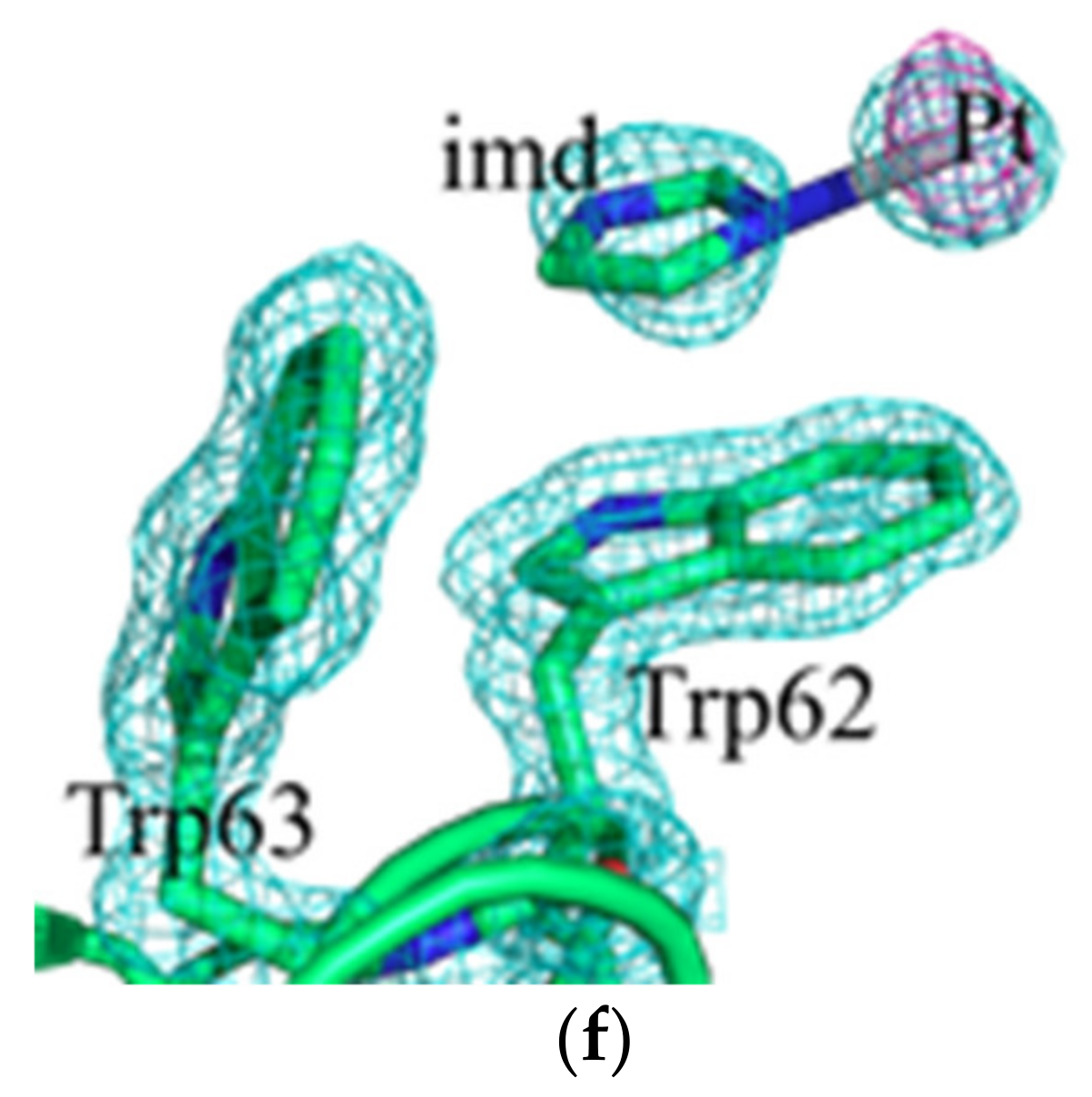
2.2. Crystallographic Results
2.3. RNase A Activity Studies
3. Discussion
3.1. Protein Interaction Studies through ESI MS Experiments
3.2. Crystallographic Studies
4. Materials and Methods
4.1. Materials
4.2. Electrospray Ionization Mass Spectrometry Analysis: Sample Preparation
4.3. Electrospray Ionization Mass Spectrometry Analysis: Final Dilutions
4.4. Electrospray Ionization Mass Spectrometry Analysis: Instrumental Parameters
4.5. Crystallization and X-ray Diffraction Data Collection
4.6. RNase A Enzymatic Activity Assays
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pérez-Herrero, E.; Fernández-Medarde, A. Advanced targeted therapies in cancer: Drug nanocarriers, the future of chemotherapy. Eur. J. Pharm. Biopharm. 2015, 93, 52–57. [Google Scholar] [CrossRef] [Green Version]
- Nabhan, C.; Phillips, E.G.; Feinberg, B.A. Orphan Cancer Drugs in the Era of Precision Medicine. JAMA Oncol. 2018, 4, 1481–1482. [Google Scholar] [CrossRef]
- Brewster, A.M.; Chavez-MacGregor, M.; Brown, P. Epidemiology, biology, and treatment of triple-negative breast cancer in women of African ancestry. Lancet Oncol. 2014, 15, e625–e634. [Google Scholar] [CrossRef] [Green Version]
- Pandy, J.P.; Balolong-Garcia, J.C.; Cruz-Ordinario, M.B.; Que, F.V.F. Triple negative breast cancer and platinum-based systemic treatment: A meta-analysis and systematic review. BMC Cancer 2019, 19, 1065. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Kang, Y.; Chen, L.; Wang, H.; Liu, J.; Zeng, S.; Yu, L. The Drug-Resistance Mechanisms of Five Platinum-Based Antitumor Agents. Front. Pharmacol. 2020, 11, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rottenberg, S.; Disler, C.; Perego, P. The rediscovery of platinum-based cancer therapy. Nat. Rev. Cancer 2021, 21, 37–50. [Google Scholar] [CrossRef]
- Rimoldi, I.; Facchetti, G.; Lucchini, G.; Castiglioni, E.; Marchianò, S.; Ferri, N. In vitro anticancer activity evaluation of new cationic platinum(II) complexes based on imidazole moiety. Bioorg. Med. Chem. 2017, 25, 1907–1913. [Google Scholar] [CrossRef] [PubMed]
- Facchetti, G.; Ferri, N.; Lupo, M.G.; Lucchini, G.; Rimoldi, I. Monofunctional PtII Complexes Based on 8-Aminoquinoline: Synthesis and Pharmacological Characterization. Eur. J. Inorg. Chem. 2019, 29, 3389–3395. [Google Scholar] [CrossRef]
- Rimoldi, I.; Coccè, V.; Facchetti, G.; Alessandri, G.; Brini, A.T.; Sisto, F.; Parati, E.; Cavicchini, L.; Lucchini, G.; Petrella, F.; et al. Uptake-release by MSCs of a cationic platinum(II) complex active in vitro on human malignant cancer cell lines. Biomed. Pharmacother. 2018, 108, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Messori, L.; Merlino, A. Protein Metalation by Metal-Based Drugs: X-ray crystallography and Mass Spectrometry studies. Chem. Commun. 2017, 53, 11622–11633. [Google Scholar] [CrossRef]
- Merlino, A.; Marzo, T.; Messori, L. Protein metalation by anticancer metallodrugs: Joint ESI MS and XRD studies. Chemistry 2017, 29, 6942–6947. [Google Scholar] [CrossRef] [PubMed]
- Schoch, S.; Gajewski, S.; Rothfuß, I.; Hartwig, A.; Köberle, B. Comparative Study of the Mode of Action of Clinically Approved Platinum-Based Chemotherapeutics. Int. J. Mol. Sci. 2020, 21, 6928. [Google Scholar] [CrossRef]
- Ferraro, G.; Marzo, T.; Cucciolito, M.E.; Ruffo, F.; Messori, L.; Merlino, A. Reaction with Proteins of a Five-Coordinate Platinum(II) Compound. Int. J. Mol. Sci. 2019, 20, 520. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, G.; Marzo, T.; Infrasca, T.; Cilibrizzi, A.; Vilar, R.; Messori, L.; Merlino, A. A case of extensive protein platination: The reaction of lysozyme with a Pt(ii)-terpyridine complex. Dalton Trans. 2018, 47, 8716–8723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timerbaev, A.R.; Hartinger, C.G.; Aleksenko, S.S.; Keppler, B.K. Interactions of antitumor metallo-drugs with serum proteins: Advances in characterization using modern analytical methodology. Chem. Rev. 2006, 106, 2224–2248. [Google Scholar] [CrossRef]
- Loreto, D.; Ferraro, G.; Merlino, A. Protein-metallodrugs interactions: Effects on the overall protein structure and characterization of Au, Ru and Pt binding sites. Inter. J. Biol. Macromol. 2020, 163, 970–976. [Google Scholar] [CrossRef]
- Ferraro, G.; Loreto, D.; Merlino, A. Interaction of platinum-based drugs with proteins: An overview of representative crystallographic studies. Curr. Top. Med. Chem. 2021, 21, 6–27. [Google Scholar] [CrossRef]
- Russo Krauss, I.; Ferraro, G.; Pica, A.; Márquez, J.; Helliwell, J.R.; Merlino, A. Principles and methods used to grow and optimize crystals of protein-metallodrug adducts, to determine metal binding sites and to assign metal ligands. Metallomics 2017, 9, 1534–1547. [Google Scholar] [CrossRef]
- Marasco, D.; Messori, L.; Marzo, T.; Merlino, A. Oxaliplatin vs. cisplatin: Competition experiments on their binding to lysozyme. Dalton Trans. 2015, 44, 10392–10398. [Google Scholar] [CrossRef] [Green Version]
- Tanley, S.W.; Helliwell, J.R. Structural dynamics of cisplatin binding to histidine in a protein. Struct. Dyn. 2014, 1, 034701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Vaney, M.C.; Maignan, S.; Ries-Kautt, M.; Ducruix, A. High-resolution structure (1.33 A) of a HEW lysozyme tetragonal crystal grown in the APCF apparatus. Data and structural comparison with a crystal grown under microgravity from SpaceHab-01 mission. Acta Crystallogr. Sect. D Biol. Crystallogr. 1996, 52, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Vitagliano, L.; Merlino, A.; Zagari, A.; Mazzarella, L. Reversible substrate-induced domain motions in ribonuclease A. Proteins 2002, 46, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Kunitz, M. A spectrophotometric method for the measurement of ribonuclease activity. J. Biol. Chem. 1946, 164, 563–568. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HEWL | RNase A | |
|---|---|---|
| Data collection | ||
| Soaking time | 12 days | 12 days |
| Space group | P43 21 2 | C 2 |
| a (Å) | 77.20 | 100.14 |
| b (Å) | 77.20 | 31.79 |
| c (Å) | 38.53 | 73.74 |
| α β γ (°) | 90.0-90.0-90.0 | 90.0-90.3-90.0 |
| Resolution range (Å) | 34.52–1.22 (1.24–1.22) | 28.04–2.13(2.17–2.13) |
| Observations | 819,608 (42,148) | 80,993 (3210) |
| Unique reflections | 35,283 (1737) | 13,153 (649) |
| Completeness (%) | 100.0 (100.0) | 99.0 (97.3) |
| Redundancy | 23.2 (24.3) | 6.2 (4.9) |
| †Rmerge (%) | 0.056 (1.531) | 0.078 (0.643) |
| Rmeas | 0.057 (1.564) | 0.086 (0.721) |
| Rpim | 0.012 (0.315) | 0.034 (0.318) |
| Average I/σ(I) | 24.5 (2.2) | 14.5 (2.3) |
| CC1/2 | 1.000 (0.861) | 0.998 (0.898) |
| Anom. completeness (%) | 100.0 (100.0) | 98.1 (94.7) |
| Anom. Multiplicity | 12.3 (12.6) | 3.3 (2.6) |
| Refinement | ||
| Resolution range (Å) | 34.55–1.22 | 28.04–2.13 |
| N. of reflections (working set) | 32,310 | 12,454 |
| N. of reflections (test set) | ||
| R-factor/R-free (%) | 12.87/16.97 | 19.75/27.43 |
| N. of atoms | 1276 | 2074 |
| Average B-factors (Å2) | ||
| All atoms | 20.05 | 39.56 |
| Pt atoms (occupancy) | 0.20–0.25 | 0.20/0.25–0.35–0.50–0.55 |
| R.m.s. deviations | ||
| Bond lengths (Å) | 0.019 | 0.013 |
| Bond angles (°) | 1.674 | 1.675 |
| Ramachandran statistics (Validation Report) | ||
| In preferred regions | 113 (98.26%) | 211 (93.36%) |
| In allowed regions | 2 (1.74%) | 10 (4.42%) |
| OutliersPDB code | 0 (0.00%)7PNH | 5 (2.21%)7PNI |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sacco, F.; Tarchi, M.; Ferraro, G.; Merlino, A.; Facchetti, G.; Rimoldi, I.; Messori, L.; Massai, L. Reactions with Proteins of Three Novel Anticancer Platinum(II) Complexes Bearing N-Heterocyclic Ligands. Int. J. Mol. Sci. 2021, 22, 10551. https://doi.org/10.3390/ijms221910551
Sacco F, Tarchi M, Ferraro G, Merlino A, Facchetti G, Rimoldi I, Messori L, Massai L. Reactions with Proteins of Three Novel Anticancer Platinum(II) Complexes Bearing N-Heterocyclic Ligands. International Journal of Molecular Sciences. 2021; 22(19):10551. https://doi.org/10.3390/ijms221910551
Chicago/Turabian StyleSacco, Francesca, Matteo Tarchi, Giarita Ferraro, Antonello Merlino, Giorgio Facchetti, Isabella Rimoldi, Luigi Messori, and Lara Massai. 2021. "Reactions with Proteins of Three Novel Anticancer Platinum(II) Complexes Bearing N-Heterocyclic Ligands" International Journal of Molecular Sciences 22, no. 19: 10551. https://doi.org/10.3390/ijms221910551
APA StyleSacco, F., Tarchi, M., Ferraro, G., Merlino, A., Facchetti, G., Rimoldi, I., Messori, L., & Massai, L. (2021). Reactions with Proteins of Three Novel Anticancer Platinum(II) Complexes Bearing N-Heterocyclic Ligands. International Journal of Molecular Sciences, 22(19), 10551. https://doi.org/10.3390/ijms221910551