
Wnt Modulation Enhances Otic Differentiation by Facilitating the Enucleation Process but Develops Unnecessary Cardiac Structures

 ,
,
Abstract
:1. Introduction
2. Results
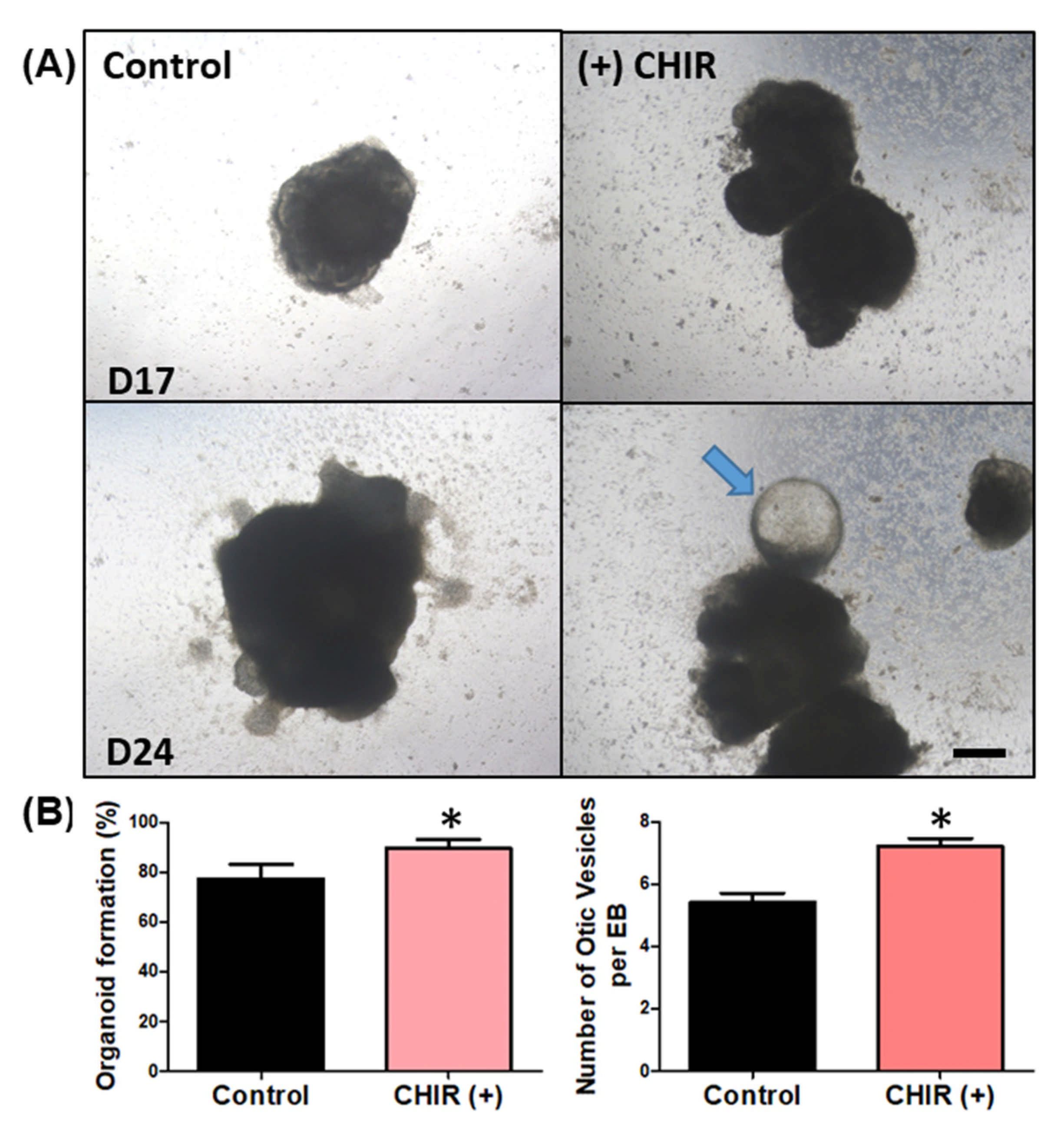
2.1. Development of EBs and Organoids
2.2. Two Different Fates of EBs (Sensory Versus Neural Differentiation)
2.3. Differentiation into Inner Ear Hair Cell-Like Cells and the Effects of Wnt Agonist Application
2.4. Wnt Agonist Application Facilitates Differentiation into Other Lineages
3. Discussion
4. Materials and Methods
4.1. Cells and Cell Culture
4.2. Embryoid Body and Otic Organoid Formation
4.3. EB Morphological Changes
4.4. Characteristics of the EBs Revealed by Epifluorescence Analysis
4.5. RT-qPCR
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Michels, T.C.; Duffy, M.T.; Rogers, D.J. Hearing Loss in Adults: Differential Diagnosis and Treatment. Am. Fam. Phys. 2019, 100, 98–108. [Google Scholar]
- Bowl, M.R.; Dawson, S.J. Age-Related Hearing Loss. Cold Spring Harb. Perspect. Med. 2019, 9, a033217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, S.B.; West, M.B.; Du, X.; Iwasa, Y.; Raphael, Y.; Kopke, R.D. Gene therapy for hair cell regeneration: Review and new data. Hear. Res. 2020, 394, 107981. [Google Scholar] [CrossRef]
- Koehler, K.R.; Hashino, E. 3D mouse embryonic stem cell culture for generating inner ear organoids. Nat. Protoc. 2014, 9, 1229–1244. [Google Scholar] [CrossRef]
- Koehler, K.R.; Nie, J.; Longworth-Mills, E.; Liu, X.-P.; Lee, J.; Holt, J.R.; Hashino, E. Generation of inner ear organoids containing functional hair cells from human pluripotent stem cells. Nat. Biotechnol. 2017, 35, 583–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dejonge, R.E.; Liu, X.-P.; Deig, C.R.; Heller, S.; Koehler, K.R.; Hashino, E. Modulation of Wnt Signaling Enhances Inner Ear Organoid Development in 3D Culture. PLoS ONE 2016, 11, e0162508. [Google Scholar] [CrossRef]
- Yuan, Z.; Li, Q.; Luo, S.; Liu, Z.; Luo, D.; Zhang, B.; Zhang, D.; Rao, P.; Xiao, J. PPARγ and Wnt Signaling in Adipogenic and Osteogenic Differentiation of Mesenchymal Stem Cells. Curr. Stem Cell Res. Ther. 2016, 11, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Wu, L. The non-canonical Wnt pathway leads to aged dendritic cell differentiation. Cell. Mol. Immunol. 2018, 15, 871–872. [Google Scholar] [CrossRef] [Green Version]
- Steinhart, Z.; Angers, S. Wnt signaling in development and tissue homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Tang, Y.; Zhou, Y.; Zhang, J. Deciphering Role of Wnt Signalling in Cardiac Mesoderm and Cardiomyocyte Differentiation from Human iPSCs: Four-dimensional control of Wnt pathway for hiPSC-CMs differentiation. Sci. Rep. 2019, 9, 19389. [Google Scholar] [CrossRef]
- Sharma, A.R.; Nam, J.-S. Kaempferol stimulates WNT/β-catenin signaling pathway to induce differentiation of osteoblasts. J. Nutr. Biochem. 2019, 74, 108228. [Google Scholar] [CrossRef]
- Chang, S.-Y.; Carpena, N.; Mun, S.; Jung, J.Y.; Chung, P.-S.; Shim, H.; Han, K.; Ahn, J.-C.; Lee, M.Y. Enhanced Inner-Ear Organoid Formation from Mouse Embryonic Stem Cells by Photobiomodulation. Mol. Ther. Methods Clin. Dev. 2020, 17, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Rabbani, C.C.; Gao, H.; Steinhart, M.R.; Woodruff, B.M.; Pflum, Z.E.; Kim, A.; Heller, S.; Liu, Y.; Shipchandler, T.Z.; et al. Hair-bearing human skin generated entirely from pluripotent stem cells. Nature 2020, 582, 399–404. [Google Scholar] [CrossRef]
- Koehler, K.; Mikosz, A.; Molosh, A.; Patel, D.; Hashino, E. Generation of inner ear sensory epithelia from pluripotent stem cells in 3D culture. Nature 2013, 500, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arkell, R.M.; Fossat, N.; Tam, P.P. Wnt signalling in mouse gastrulation and anterior development: New players in the pathway and signal output. Curr. Opin. Genet. Dev. 2013, 23, 454–460. [Google Scholar] [CrossRef] [PubMed]
- Hoang, P.; Wang, J.; Conklin, B.R.; Healy, K.E.; Ma, Z. Generation of spatial-patterned early-developing cardiac organoids using human pluripotent stem cells. Nat. Protoc. 2018, 13, 723–737. [Google Scholar] [CrossRef]
- Breckwoldt, K.; Letuffe-Brenière, D.; Mannhardt, I.; Schulze, T.; Ulmer, B.; Werner, T.; Benzin, A.; Klampe, B.; Reinsch, M.C.; Laufer, S.; et al. Differentiation of cardiomyocytes and generation of human engineered heart tissue. Nat. Protoc. 2017, 12, 1177–1197. [Google Scholar] [CrossRef]
- Guo, X.-M.; Zhao, Y.-S.; Chang, H.-X.; Wang, C.-Y.; E, L.-L.; Zhang, X.-A.; Duan, C.-M.; Dong, L.-Z.; Jiang, H.; Li, J.; et al. Creation of Engineered Cardiac Tissue In Vitro From Mouse Embryonic Stem Cells. Circulation 2006, 113, 2229–2237. [Google Scholar] [CrossRef] [Green Version]
- Shkumatov, A.; Baek, K.; Kong, H. Matrix Rigidity-Modulated Cardiovascular Organoid Formation from Embryoid Bodies. PLoS ONE 2014, 9, e94764. [Google Scholar] [CrossRef]
- Sun, X.; Nunes, S.S. Biowire platform for maturation of human pluripotent stem cell-derived cardiomyocytes. Methods 2016, 101, 21–26. [Google Scholar] [CrossRef]
- Mathur, A.; Loskill, P.; Shao, K.; Huebsch, N.; Hong, S.; Marcus, S.G.; Marks, N.; Mandegar, M.; Conklin, B.R.; Lee, L.P.; et al. Human iPSC-based cardiac microphysiological system for drug screening applications. Sci. Rep. 2015, 5, 8883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Koo, S.; Finnegan, M.A.; Loskill, P.; Huebsch, N.; Marks, N.C.; Conklin, B.R.; Grigoropoulos, C.; Healy, K.E. Three-dimensional filamentous human diseased cardiac tissue model. Biomaterials 2014, 35, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinson, J.T.; Chopra, A.K.; Nafissi, N.; Polacheck, W.J.; Benson, C.C.; Swist, S.; Gorham, J.M.; Yang, L.; Schafer, S.; Sheng, C.C.; et al. Titin mutations in iPS cells define sarcomere insufficiency as a cause of dilated cardiomyopathy. Science 2015, 349, 982–986. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Host | Supplier | Catalog No. | Dilution |
|---|---|---|---|---|
| Brn3c | Mouse | Santa Cruz | Sc81980 | 1:25 |
| CTPI | Rabbit | Abcam | ab47003 | 1:100 |
| F-actin (Fluorescein Phalloidin) | Thermo Fisher Scientific | F432 | 1:100 | |
| GFAP | Chicken | Abcam | ab4674 | 1:1000 |
| MyosinVIIa | Rabbit | Proteus | 256790 | 1:100 |
| Nestin | Mouse | Abcam | ab11306 | 1:100 |
| SOX2 | Mouse | BD Biosciences | 561469 | 1:100 |
| TAGLN (SM22) | Rabbit | Abcam | ab14106 | 1:200 |
| TuJ1 | Mouse | Biolegend | 836504 | 1:100 |
| Oligomer | Forward Sequence | Reverse Sequence |
|---|---|---|
| Atoh1 | 5′-TCCTATGAAGGAGGTGCGGG-3′ | 5′-TTAGGGCCCTGTCCTCGAAG-3′ |
| E-cadherin | 5′-TCTTAGGCACCCAGTAGGCC-3′ | 5′-TTCCAGGGAGACTGCTAGGC-3′ |
| GAPDH | 5′-AGGTCGGTGTGAACGGATTTG-3′ | 5′-GTAGACCATGTAGTTGAGGTCA-3′ |
| Laminin-β1 | 5′-CACCCCTAGCCAACTTGCTG-3′ | 5′-CTTTGTTCTCCTCACCCGGC-3′ |
| Myo7a | 5’-CACCAAGGGAGATTGTGGCC-3´ | 5´-CCTTGGACACCATGACACGG-3´ |
| Pax2 | 5′-GACAGCACCAGACAAGAGGC-3′ | 5′-TAGCCAAAAAGCCTCGGCAG-3′ |
| Pax8 | 5′-CTTTGCAGTCCCCAGCTCAG-3′ | 5′-GCCAAGTGCTCTCCTGTGTC-3′ |
| Sox2 | 5´-CACCCCTAGCCAACTTGCTG-3´ | 5´-CTTTGTTCTCCTCACCCGGC-3´ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carpena, N.T.; Chang, S.-Y.; Choi, J.-E.; Jung, J.Y.; Lee, M.Y. Wnt Modulation Enhances Otic Differentiation by Facilitating the Enucleation Process but Develops Unnecessary Cardiac Structures. Int. J. Mol. Sci. 2021, 22, 10306. https://doi.org/10.3390/ijms221910306
Carpena NT, Chang S-Y, Choi J-E, Jung JY, Lee MY. Wnt Modulation Enhances Otic Differentiation by Facilitating the Enucleation Process but Develops Unnecessary Cardiac Structures. International Journal of Molecular Sciences. 2021; 22(19):10306. https://doi.org/10.3390/ijms221910306
Chicago/Turabian StyleCarpena, Nathaniel T., So-Young Chang, Ji-Eun Choi, Jae Yun Jung, and Min Young Lee. 2021. "Wnt Modulation Enhances Otic Differentiation by Facilitating the Enucleation Process but Develops Unnecessary Cardiac Structures" International Journal of Molecular Sciences 22, no. 19: 10306. https://doi.org/10.3390/ijms221910306
APA StyleCarpena, N. T., Chang, S.-Y., Choi, J.-E., Jung, J. Y., & Lee, M. Y. (2021). Wnt Modulation Enhances Otic Differentiation by Facilitating the Enucleation Process but Develops Unnecessary Cardiac Structures. International Journal of Molecular Sciences, 22(19), 10306. https://doi.org/10.3390/ijms221910306