
Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases

 ,
, 
 and
and
Abstract
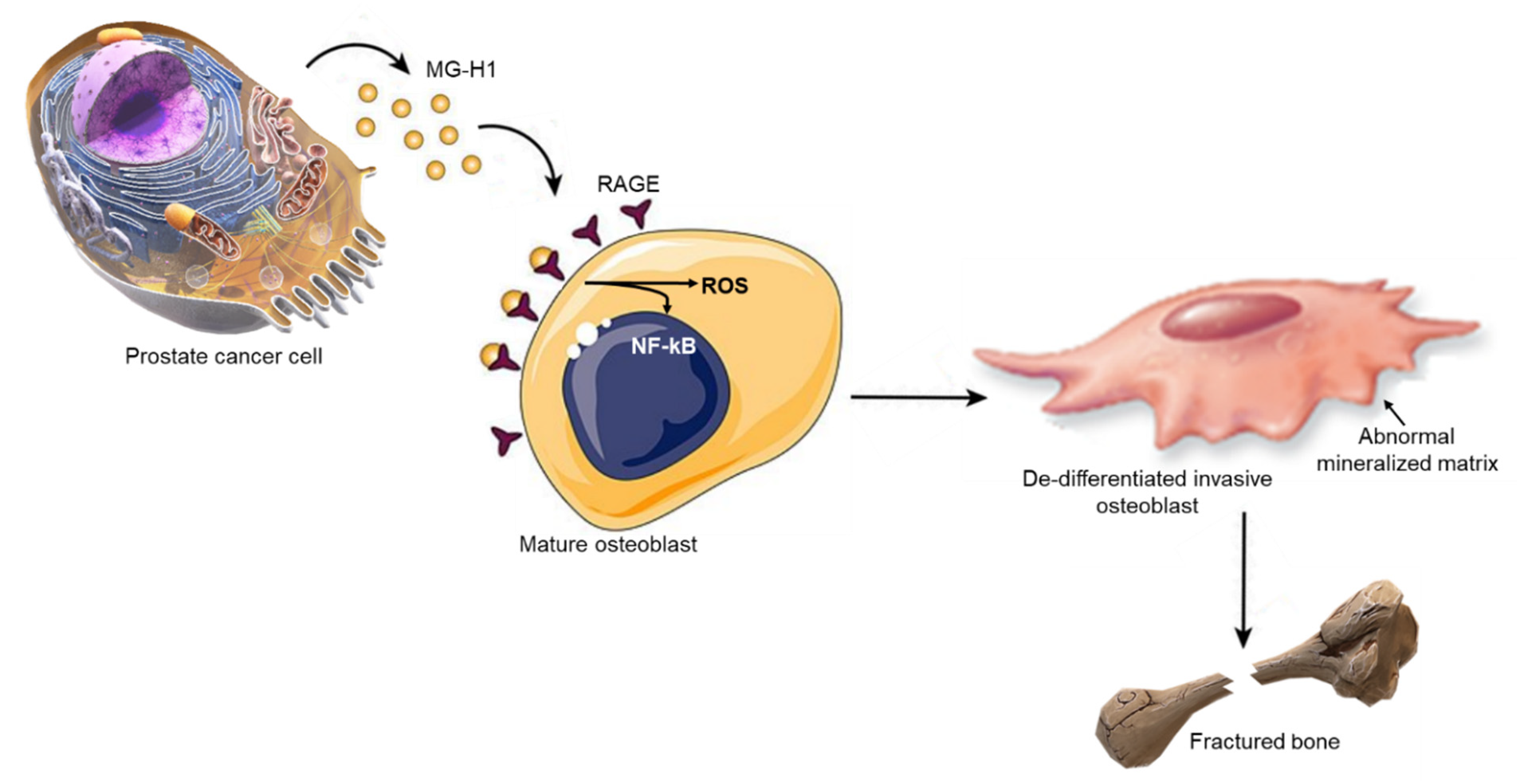
:1. Introduction
2. Results
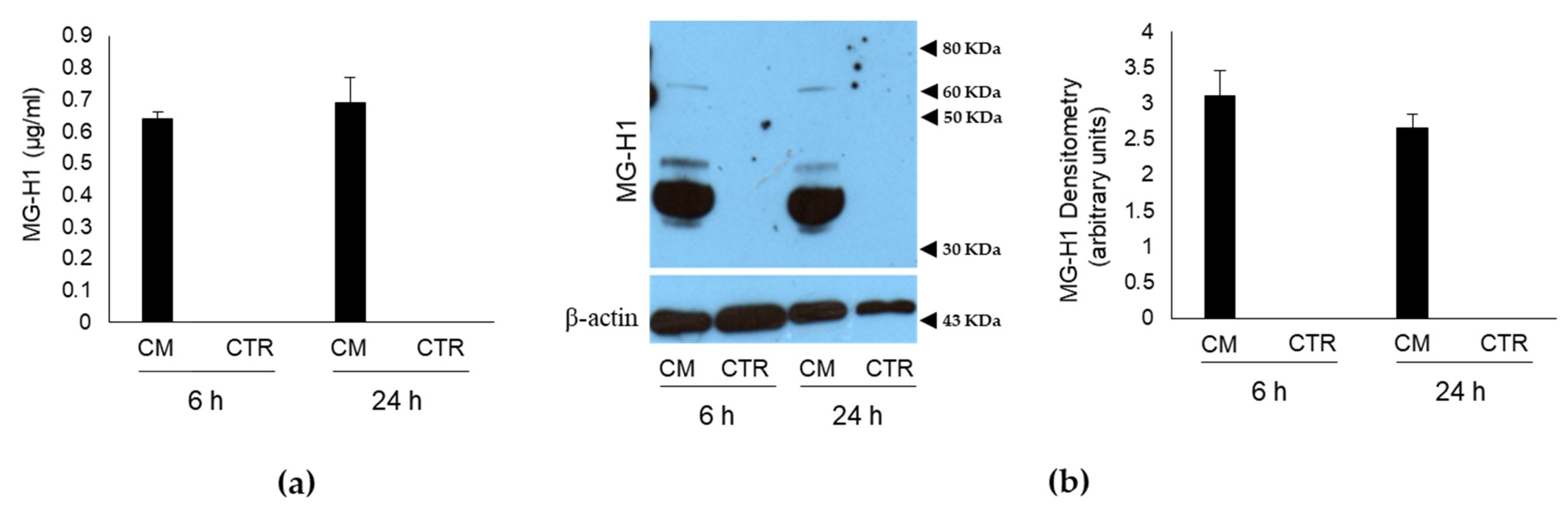
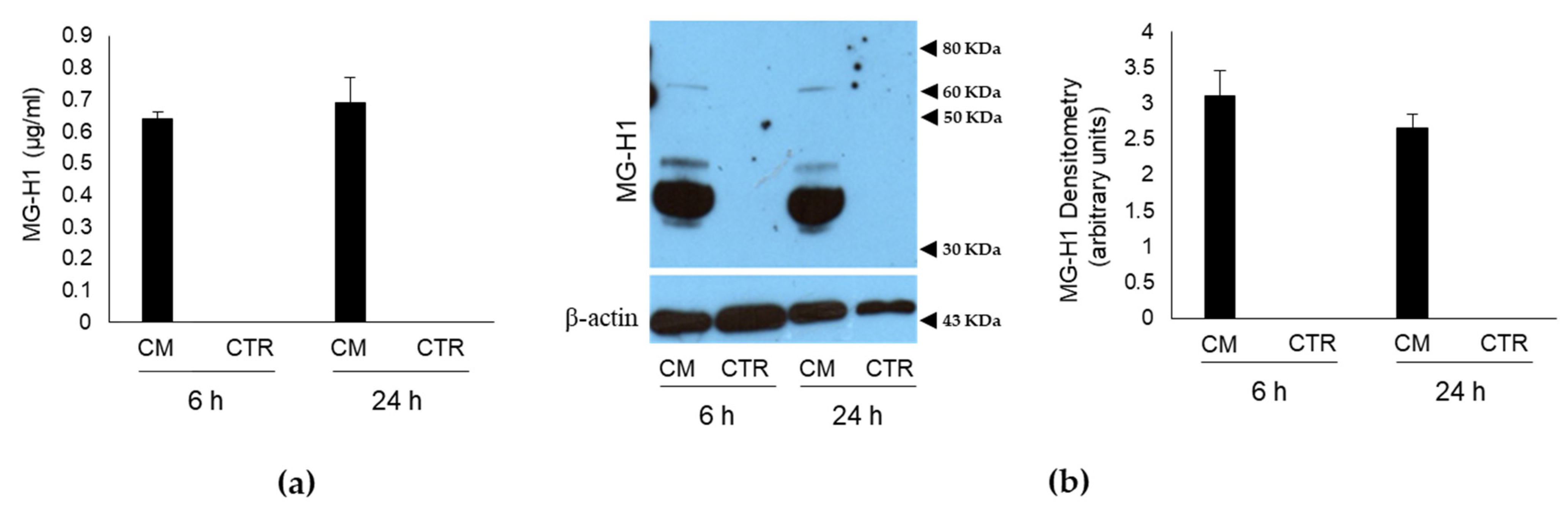
2.1. Conditioned Medium (CM) from PC3 Bone Metastasis-Derived PCa Cells Contains MG-Originated MG-H1
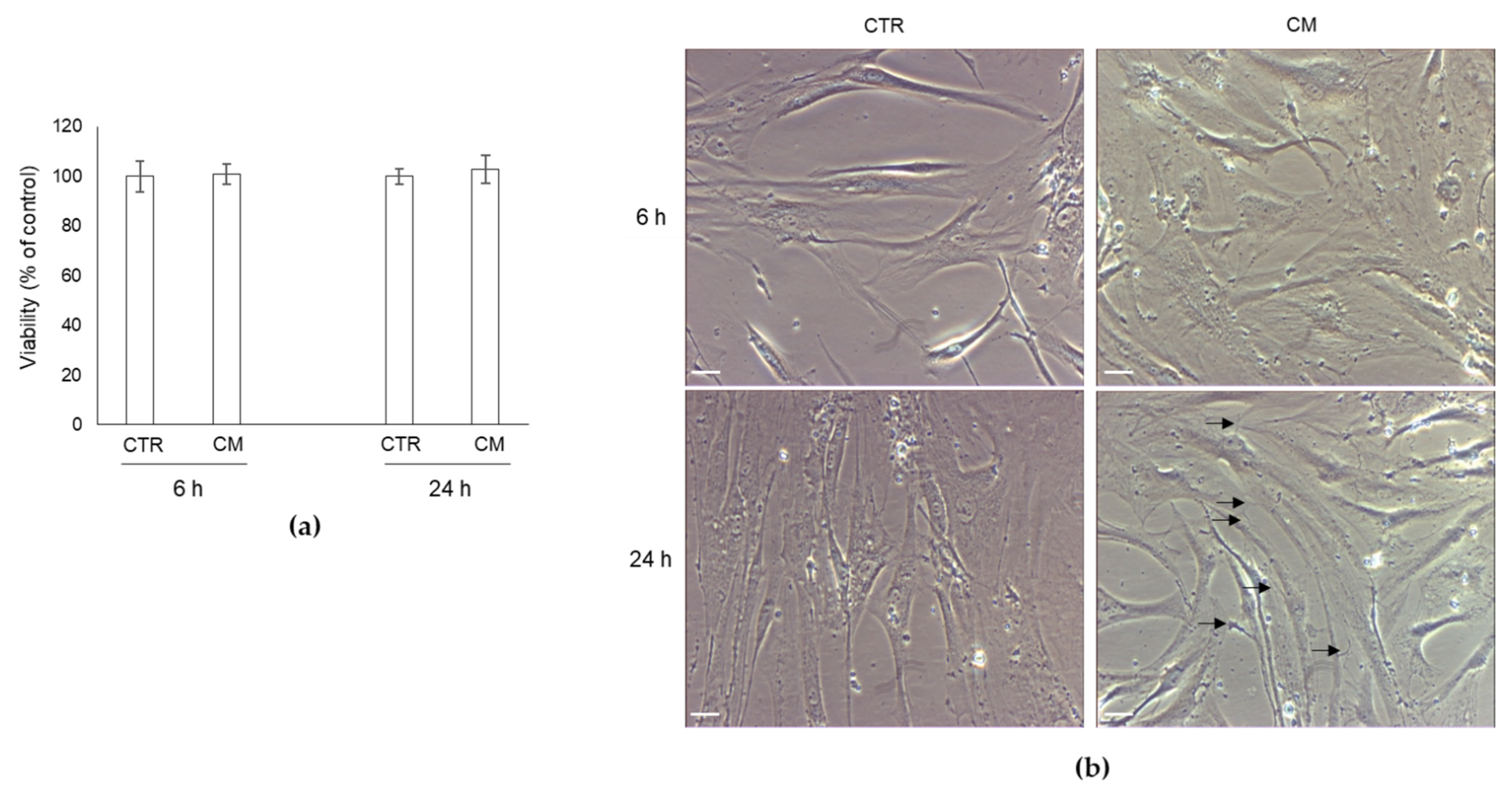
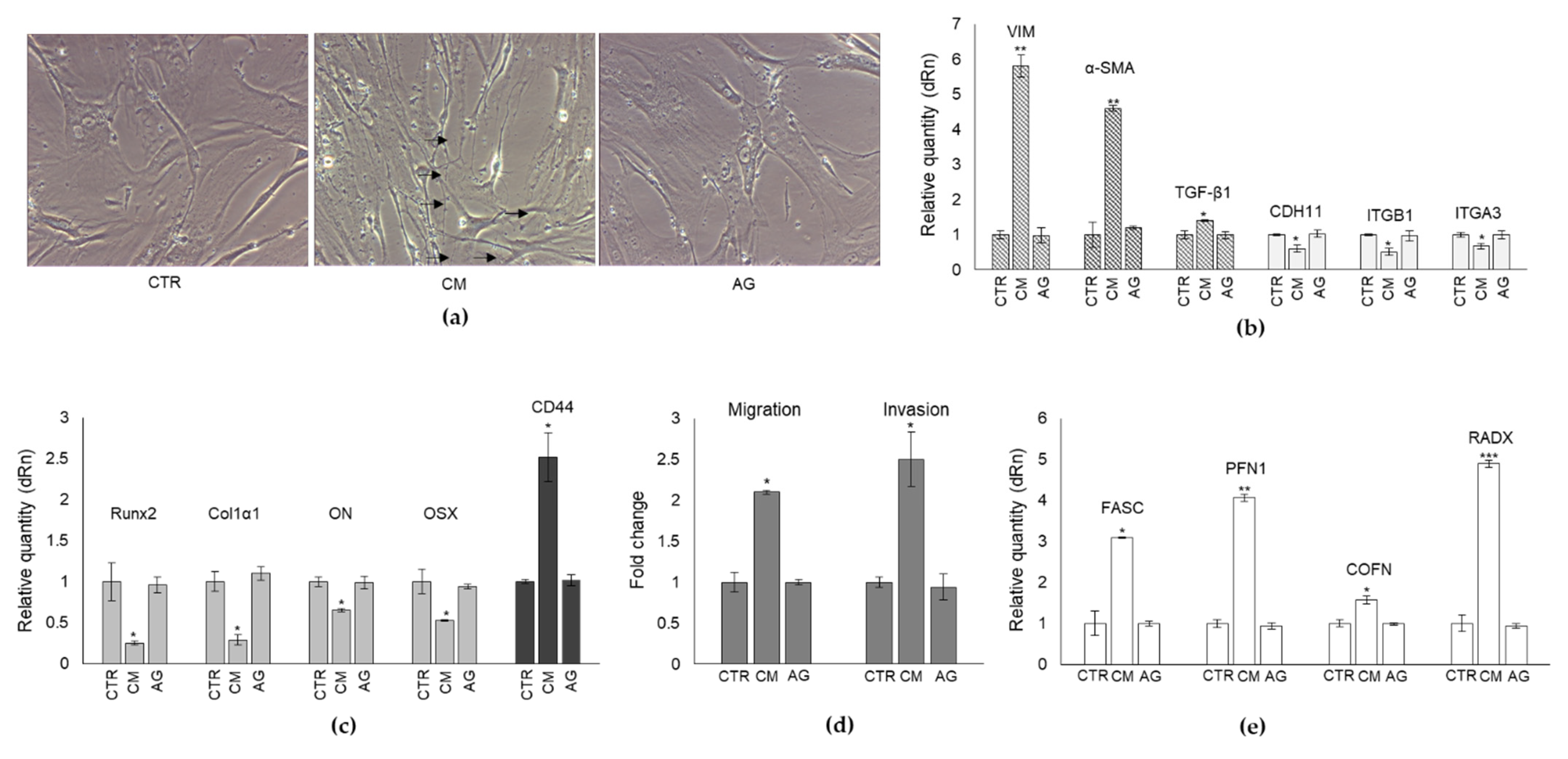
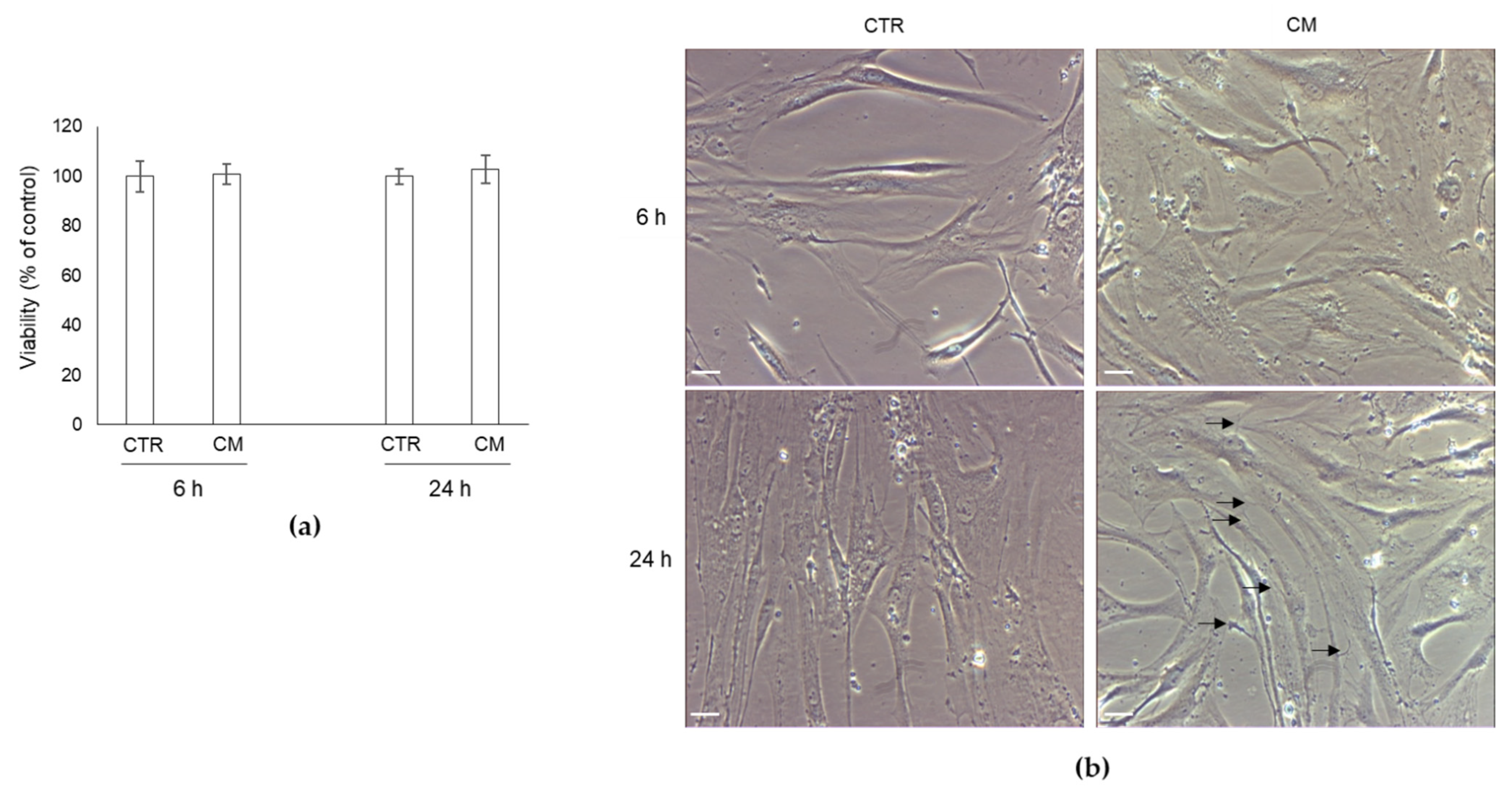
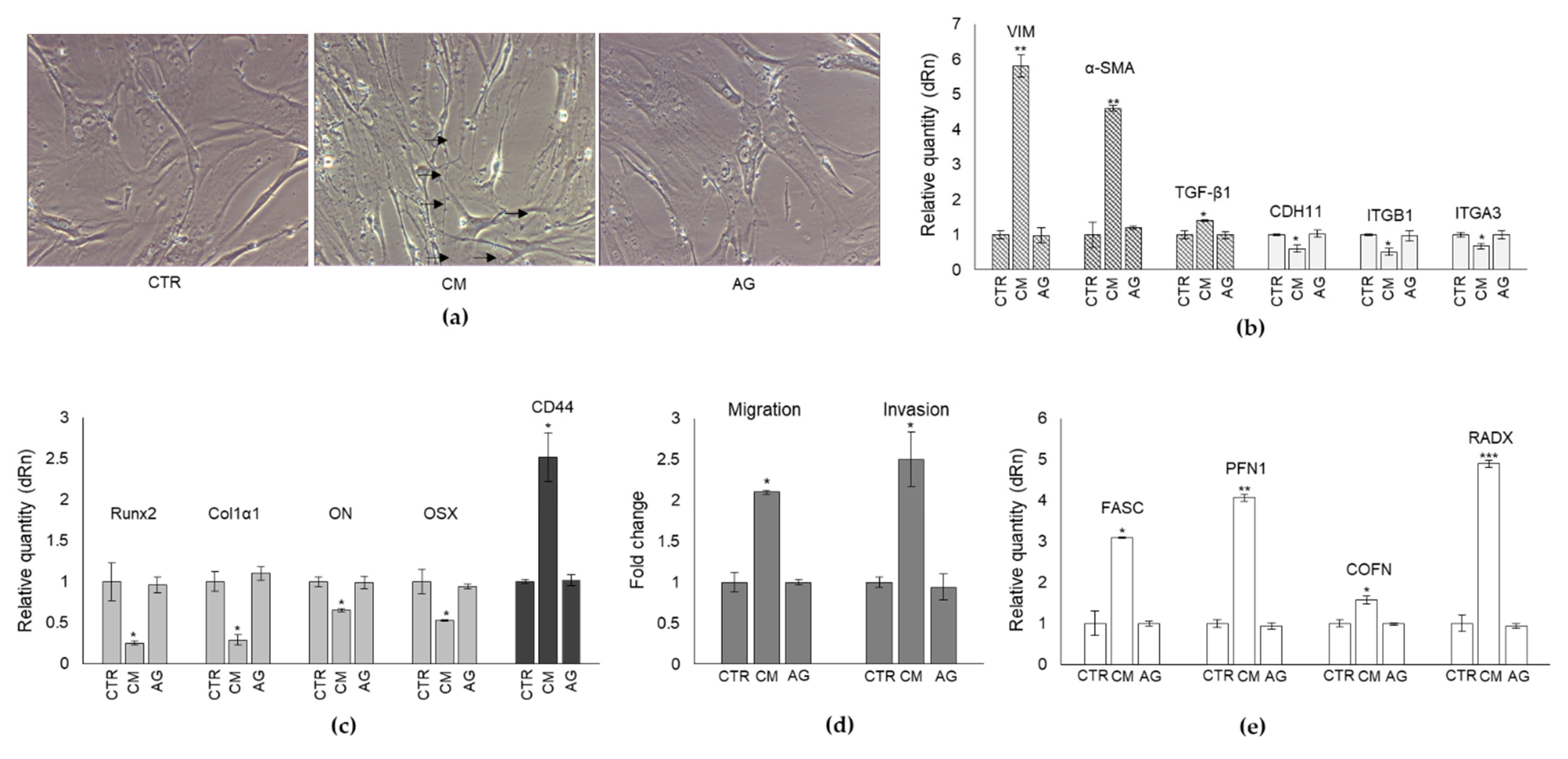
2.2. Effect of CM from PC3 Cells on Human OB Viability and Morphology
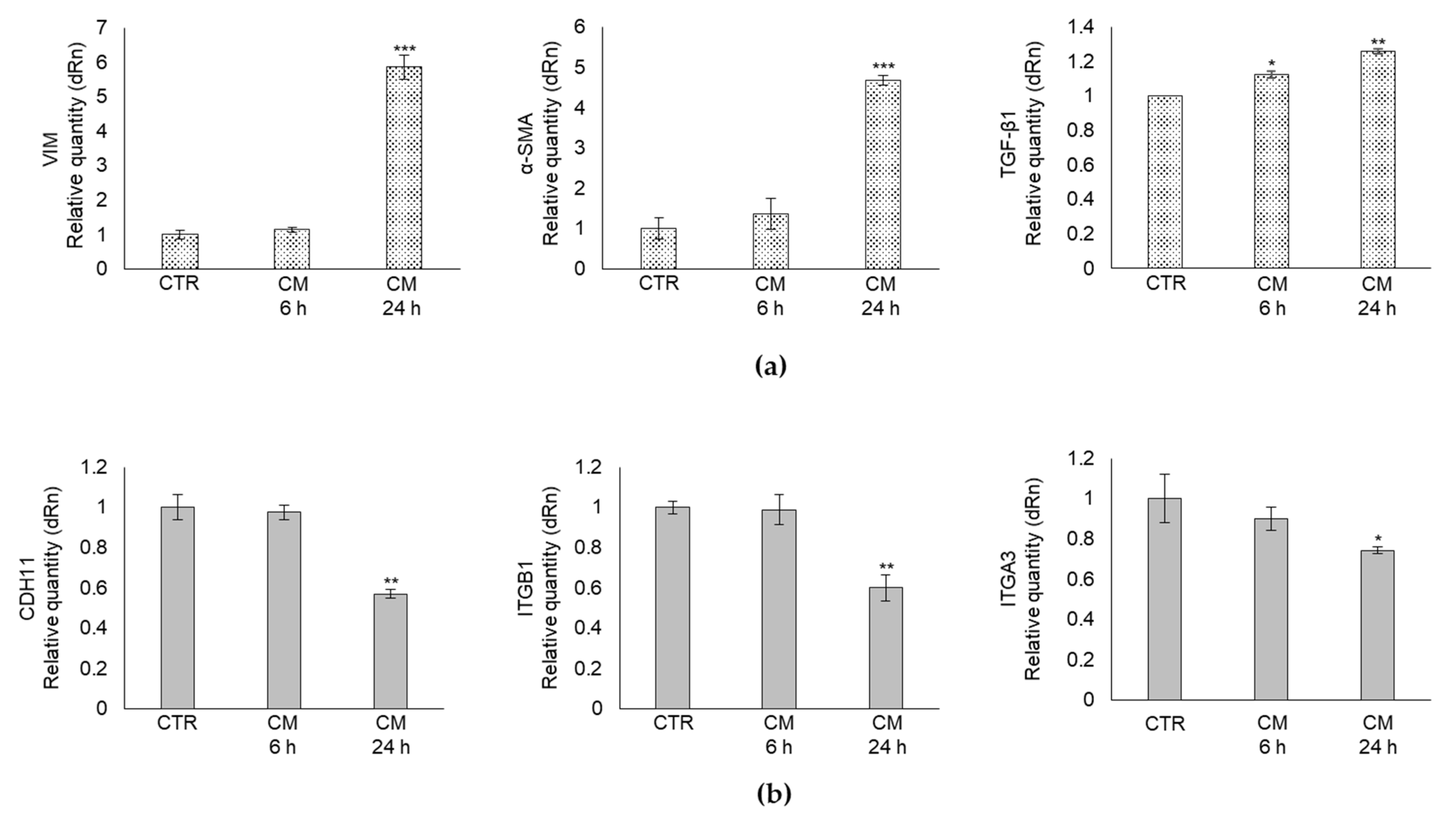
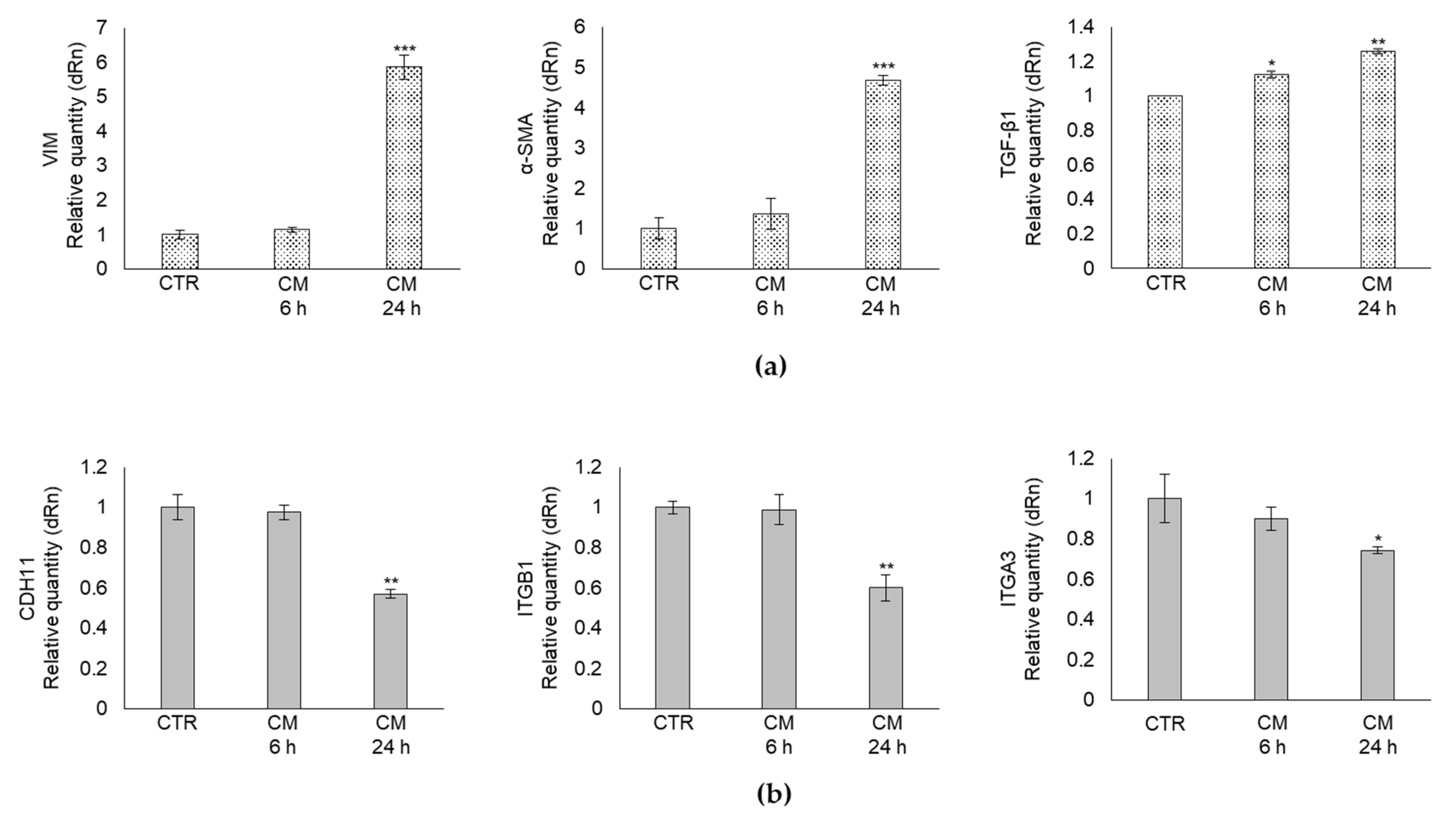
2.3. CM from PC3 Cells Alters in OB the mRNA Expression of Specific Markers Associated with Mesenchymal Trans-Differentiation
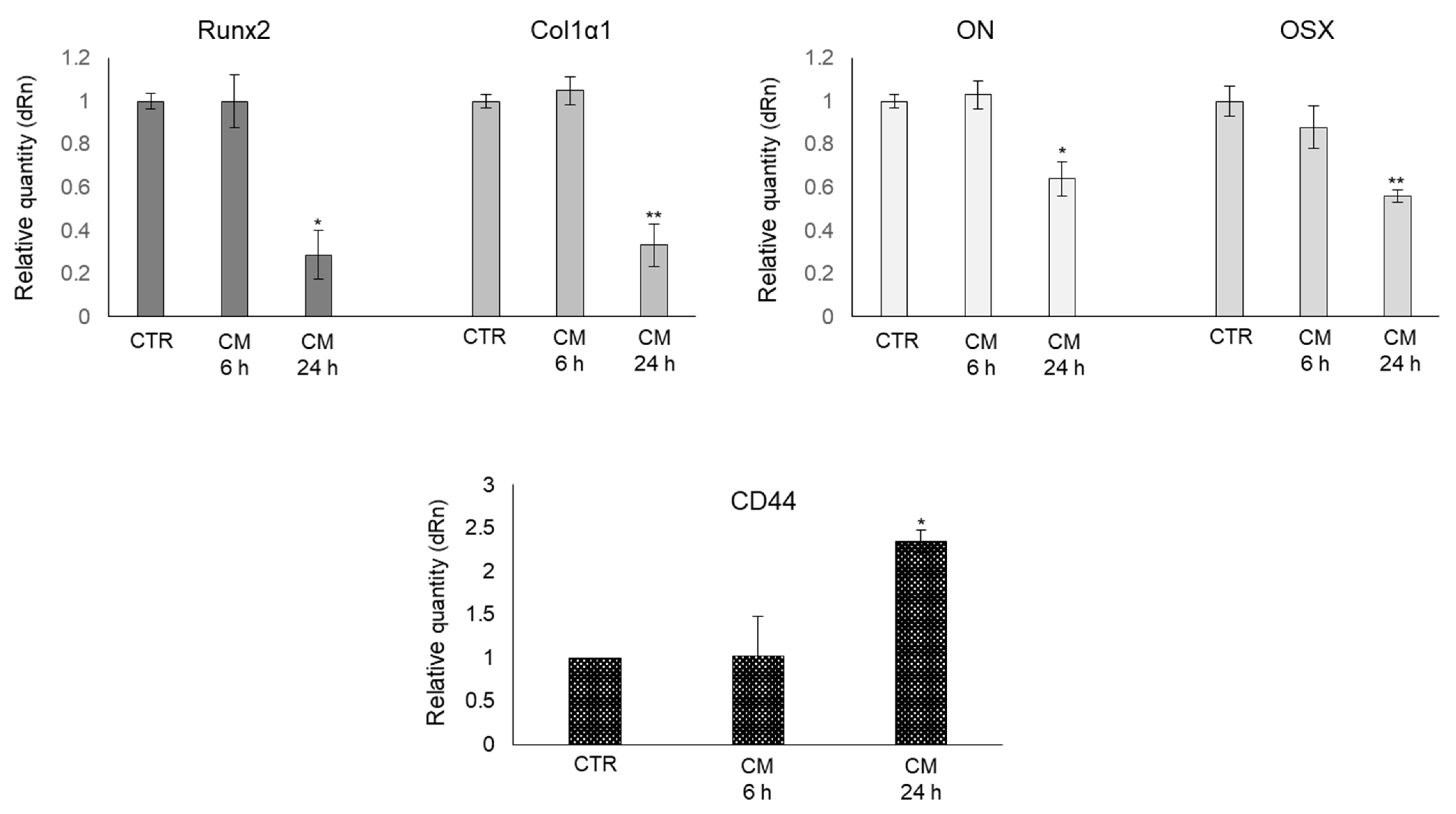
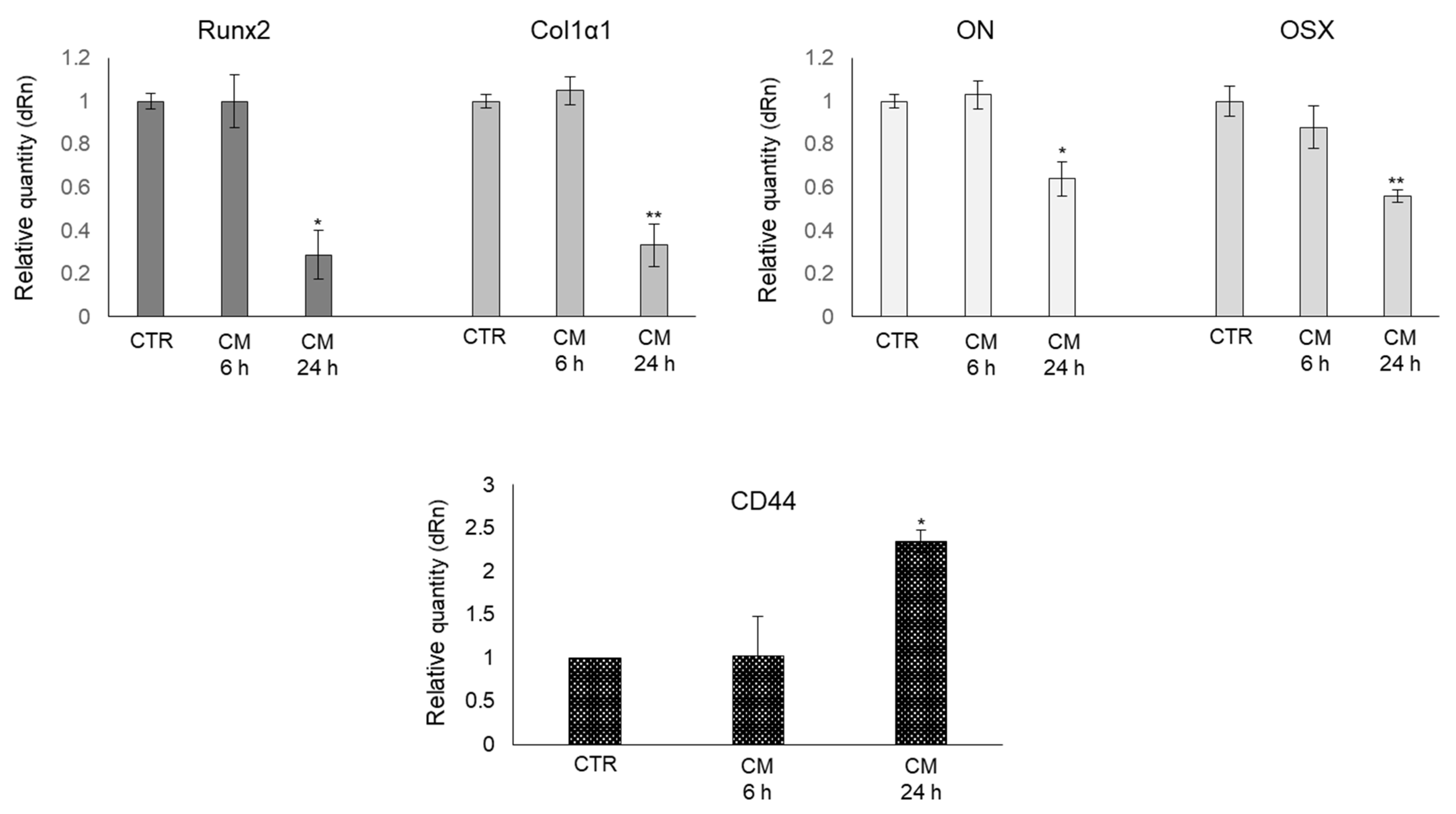
2.4. CM from PC3 Cells Alters the mRNA Expression of Specific Markers Associated with OB Mature Phenotype
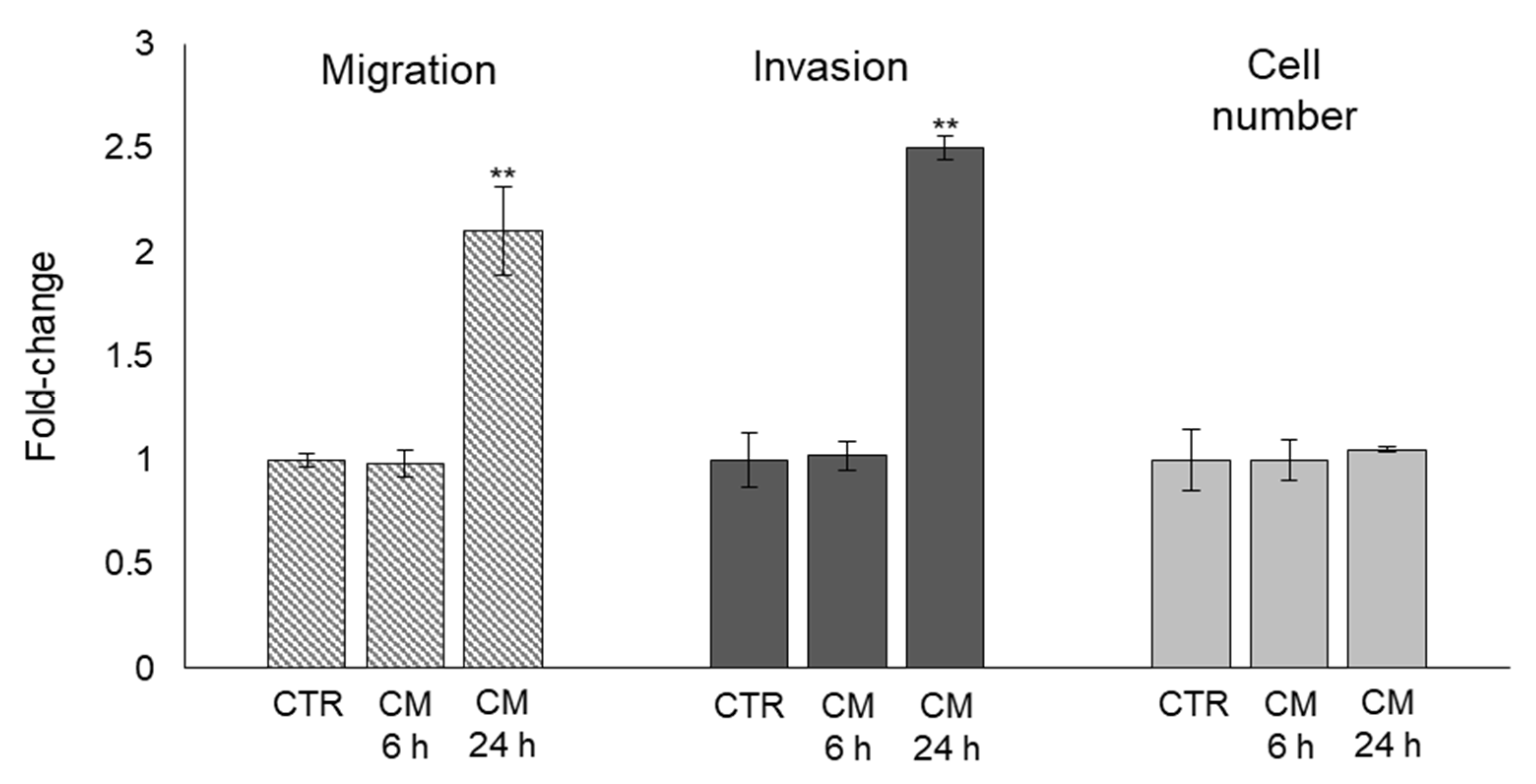
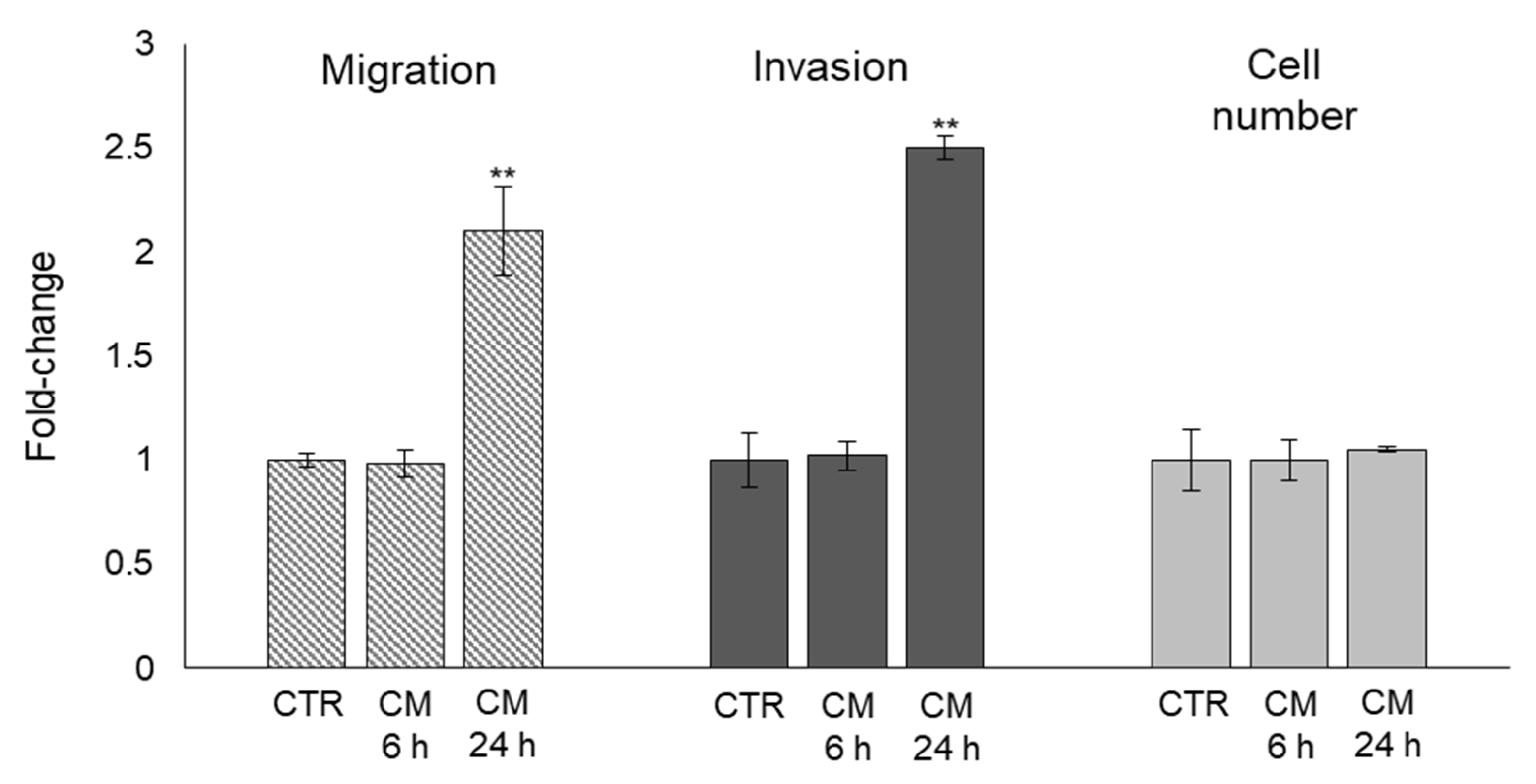
2.5. The Trans-Differentiation of OB Phenotype to A Mesenchymal-like One upon PC3 Cells CM Exposure Is Accompanied by Increased Migration/Invasion but Not Proliferation
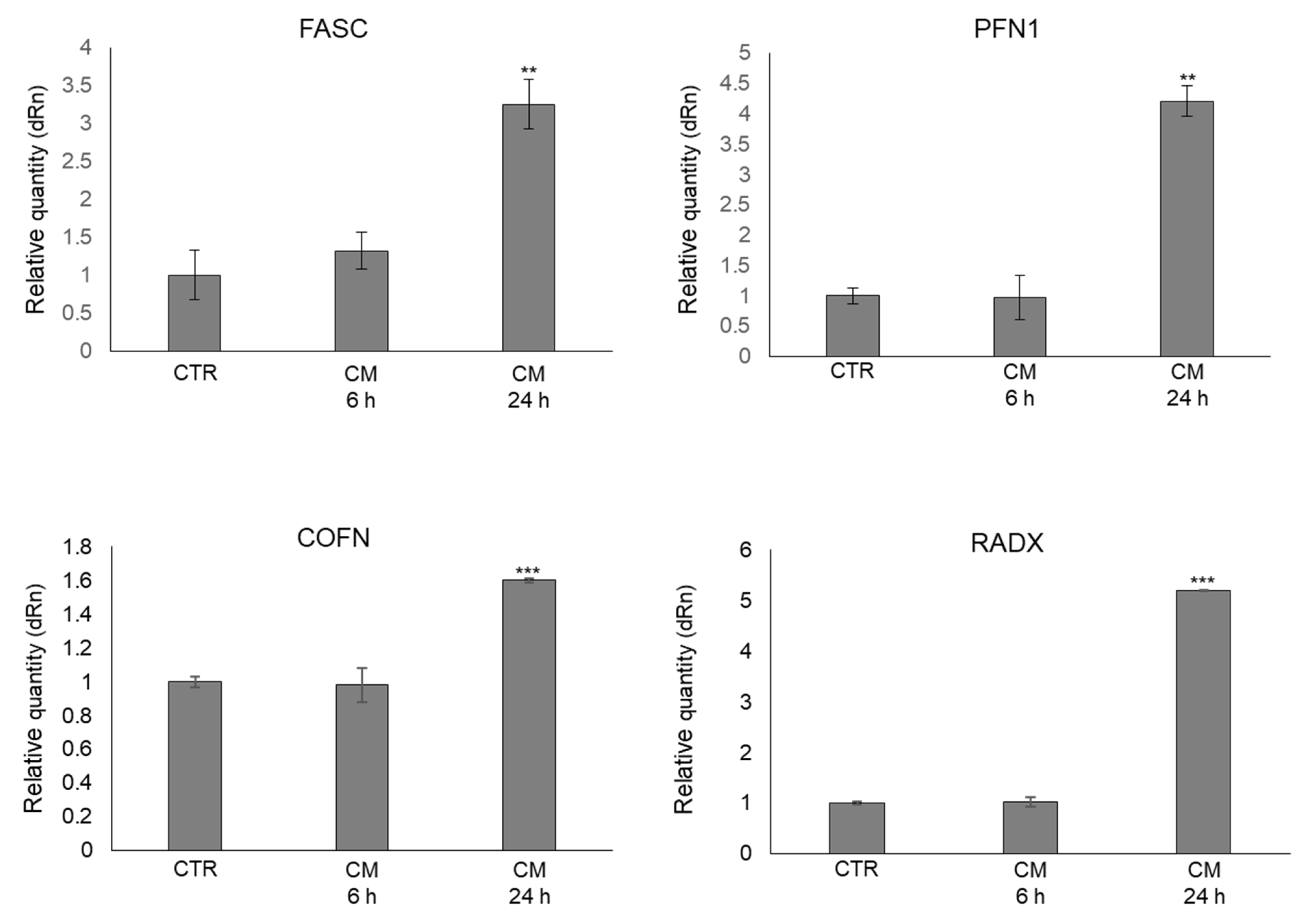
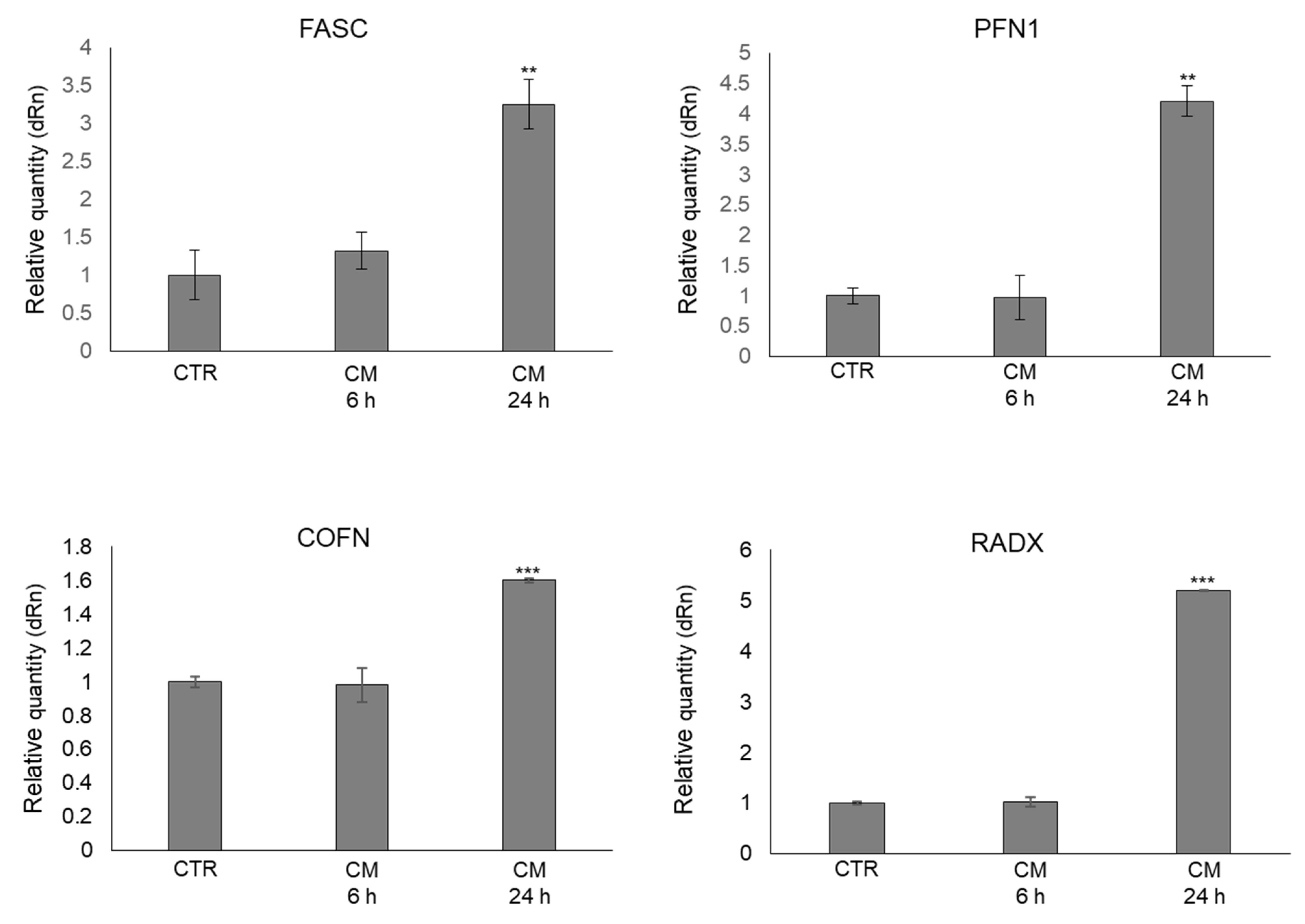
2.6. CM from PC3 Cells Alters in OB the mRNA Expression of Proteins Involved in the Formation of Dynamic Cell Protrusions
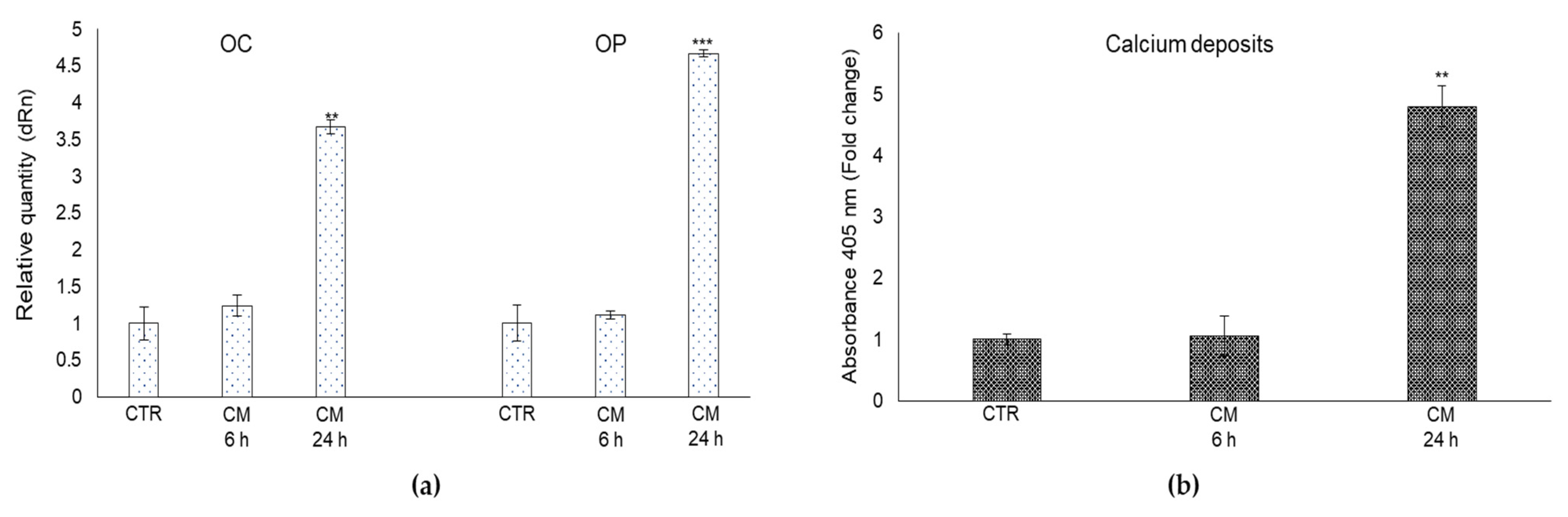
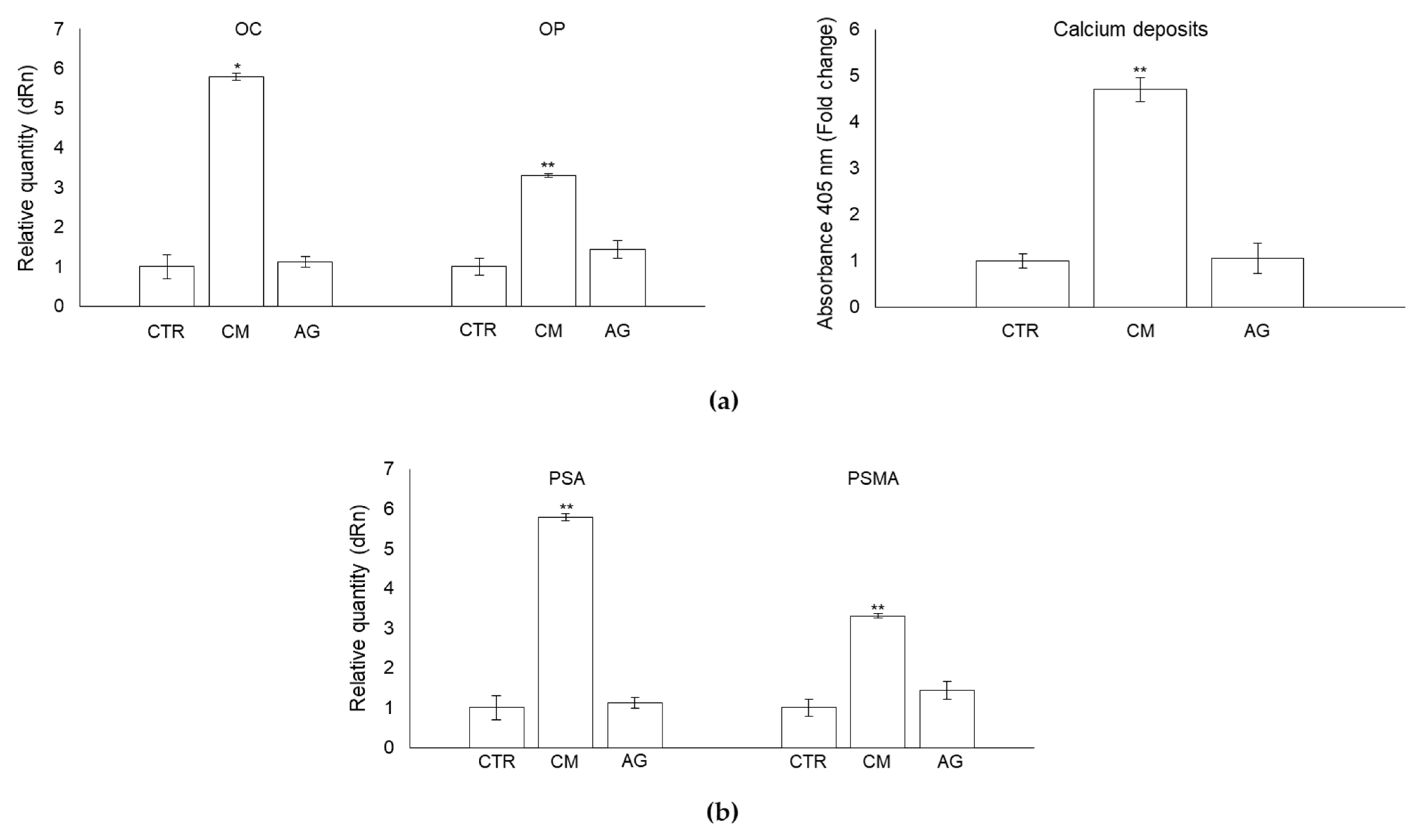
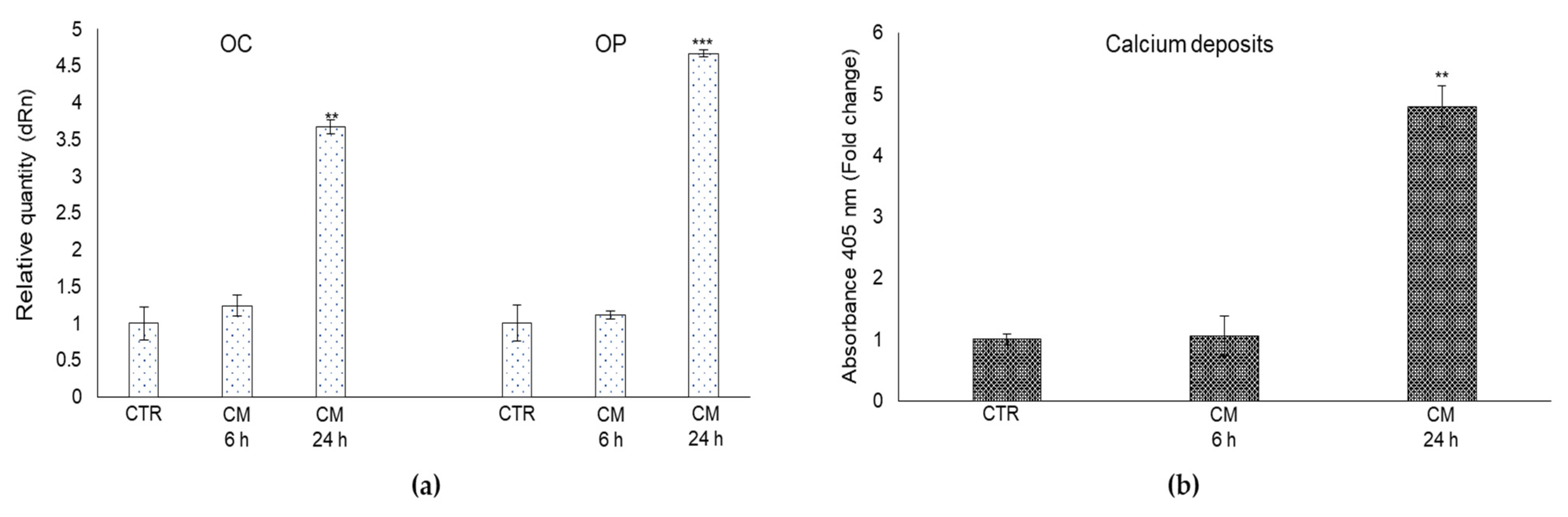
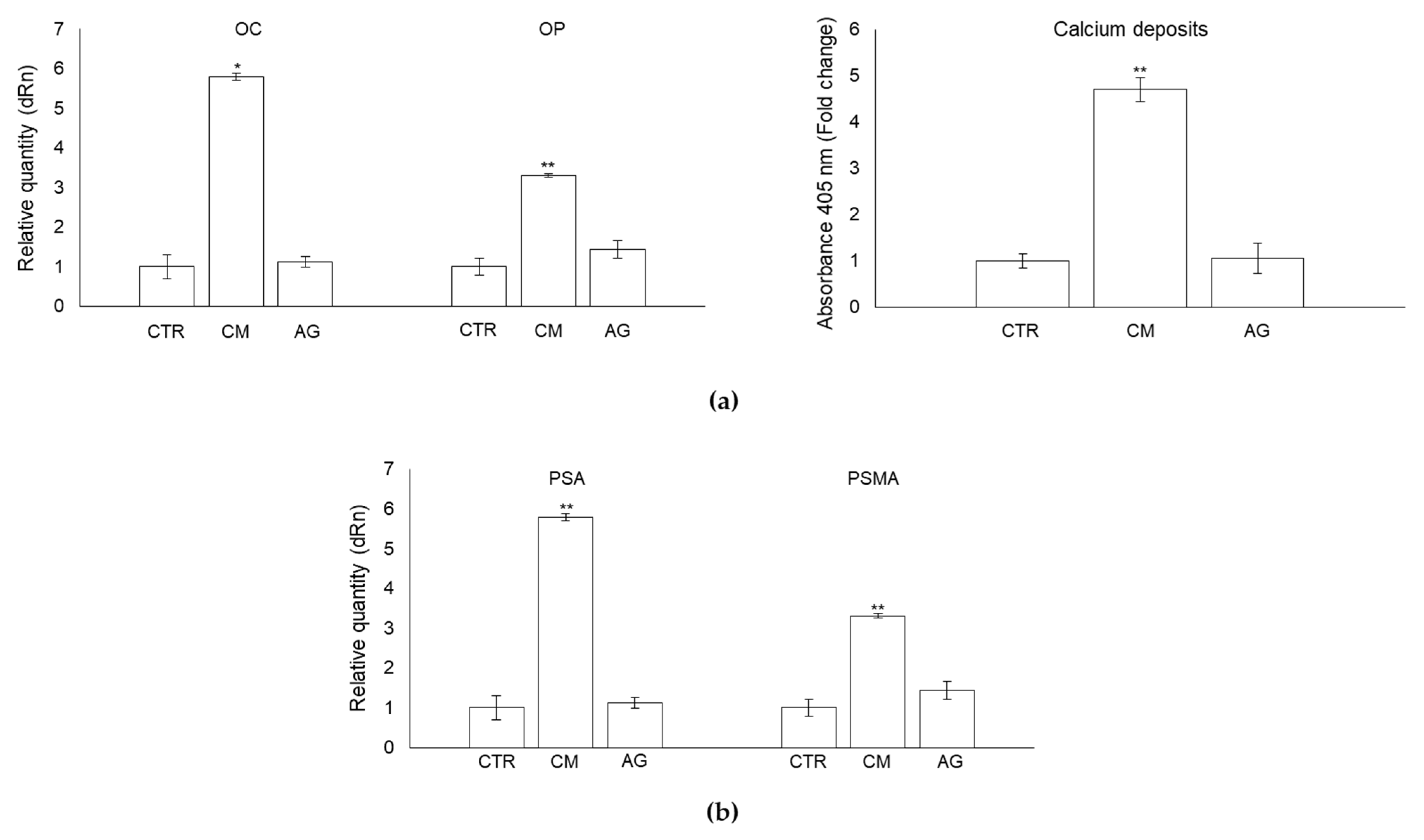
2.7. CM from PC3 Cells Affects OB Mineralization Activity
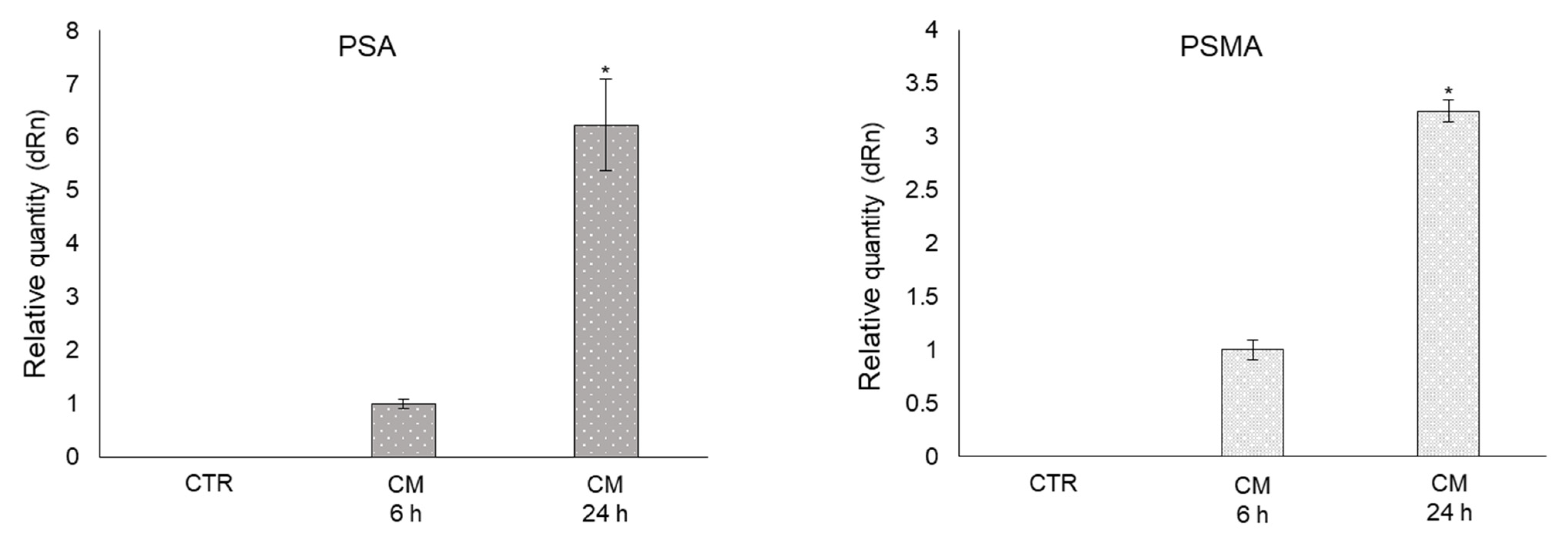
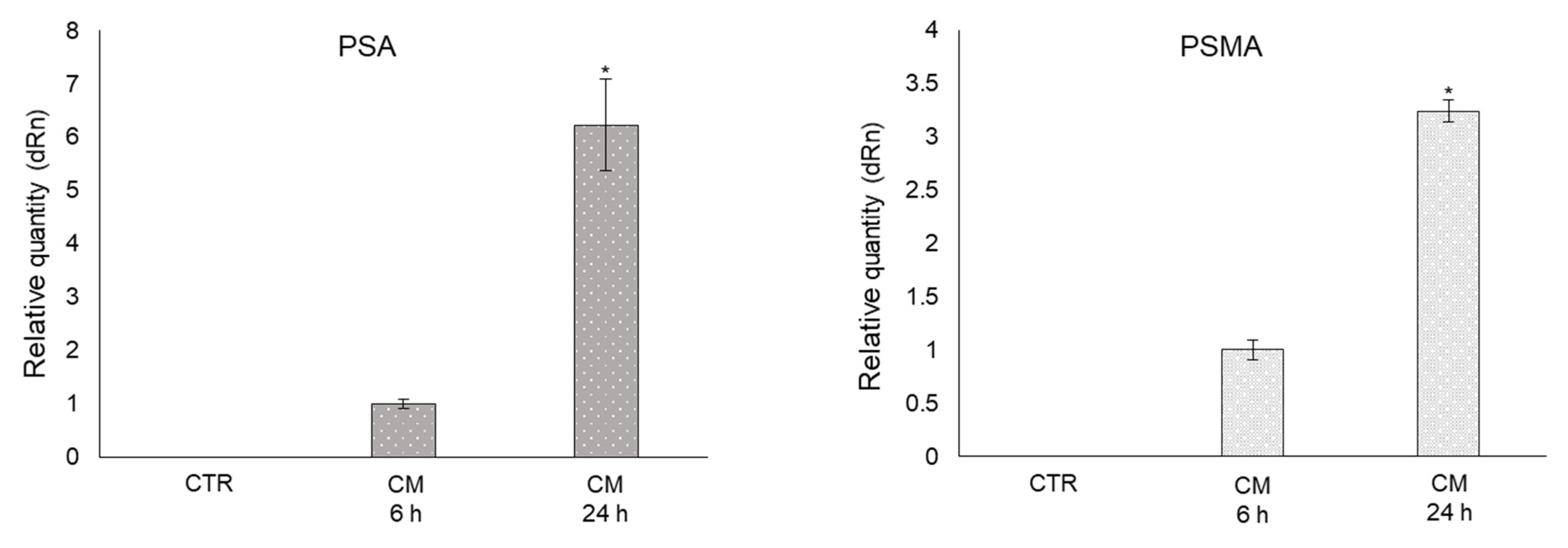
2.8. CM from PC3 Cells Induces De Novo Expression of the Prostate-Specific Antigens PSA and PSMA in OB
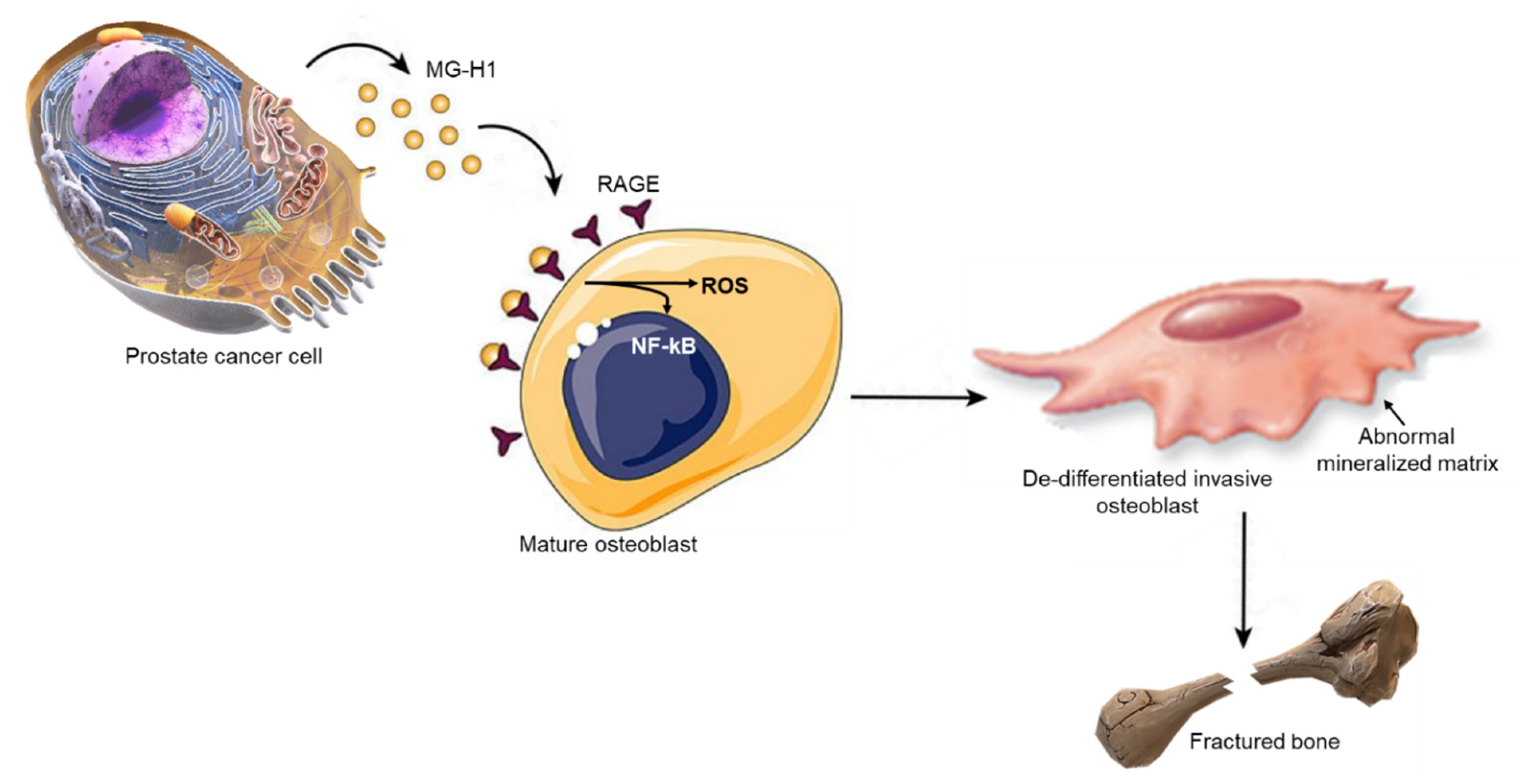
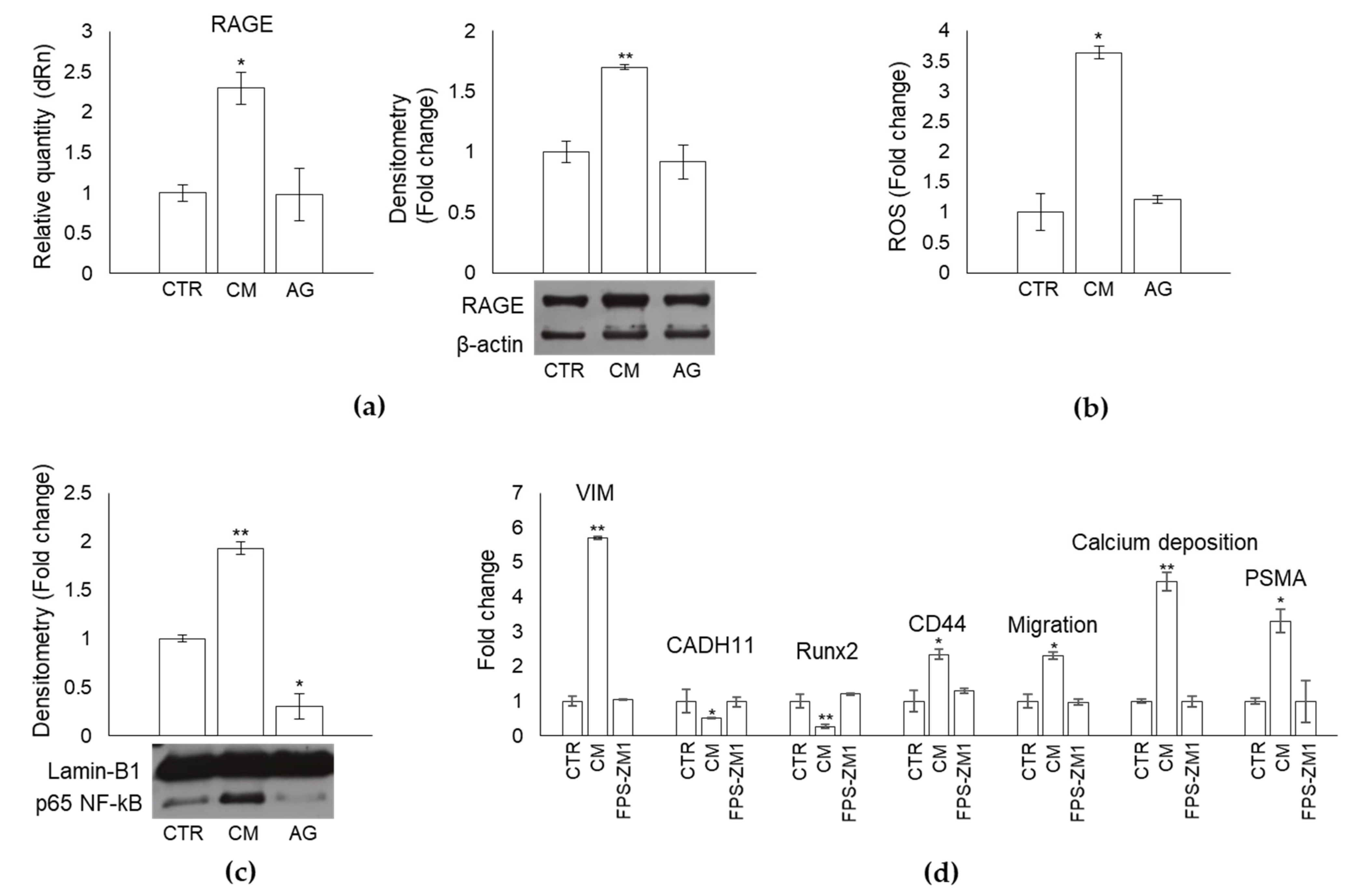
2.9. MG-derived MG-H1 Is a Novel Paracrine Factor Released by Bone Metastasis-Derived PC3 Cells to Reprogram Human OB into a Dedifferentiated, Mesenchymal-, and Malignant-like Phenotype
2.10. MG-Derived MG-H1 Released by Bone Metastasis-Derived PC3 Cells Reprograms Human OB into a Mesenchymal-, Malignant-like Phenotype through a RAGE-Dependent Mechanism with the Involvement of ROS and NF-kB Signaling
2.11. CM from LNCaP Cells, Devoid of MG-H1, Does Not Reprogram OB into a Dedifferentiated Malignant-like Phenotype
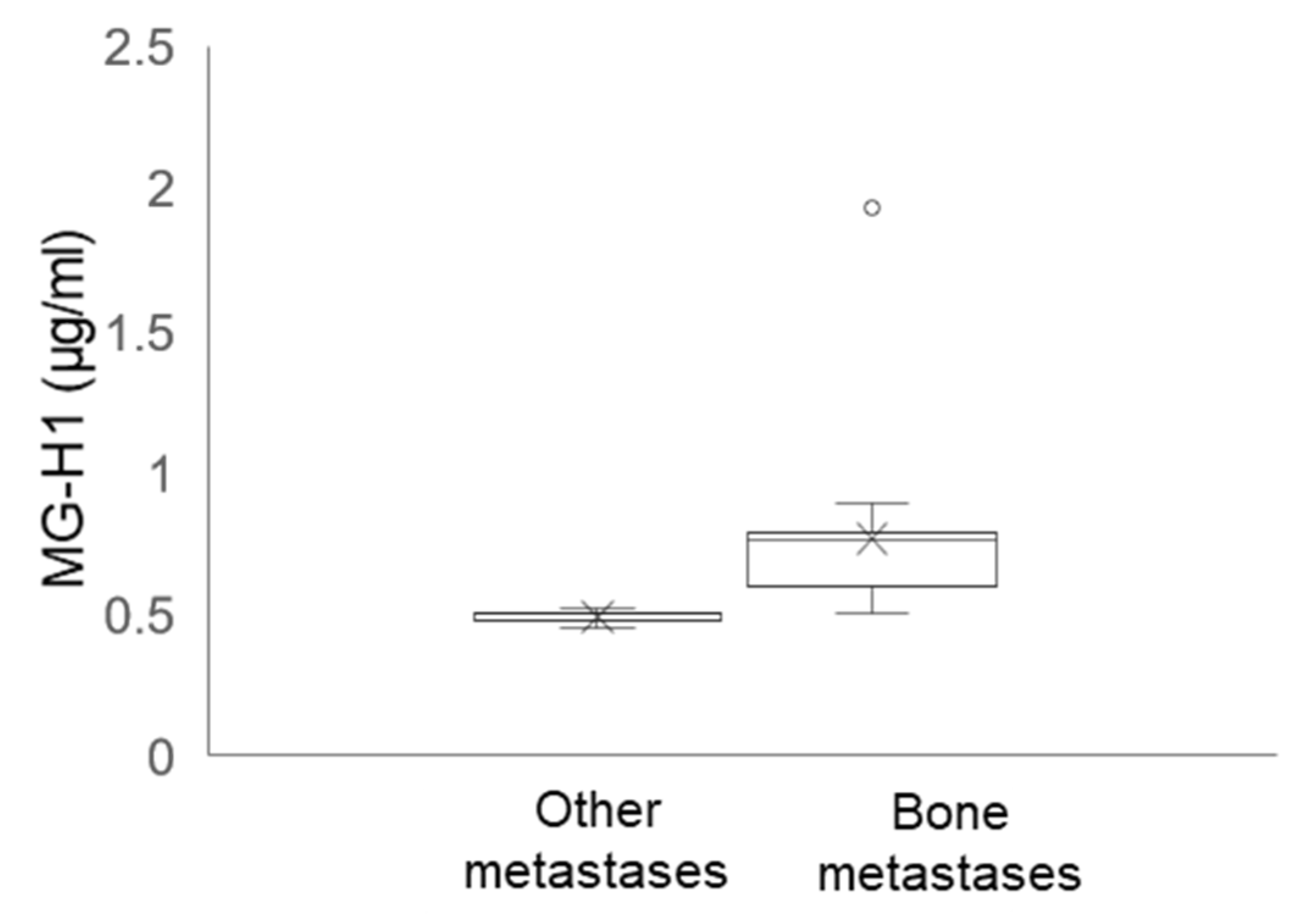
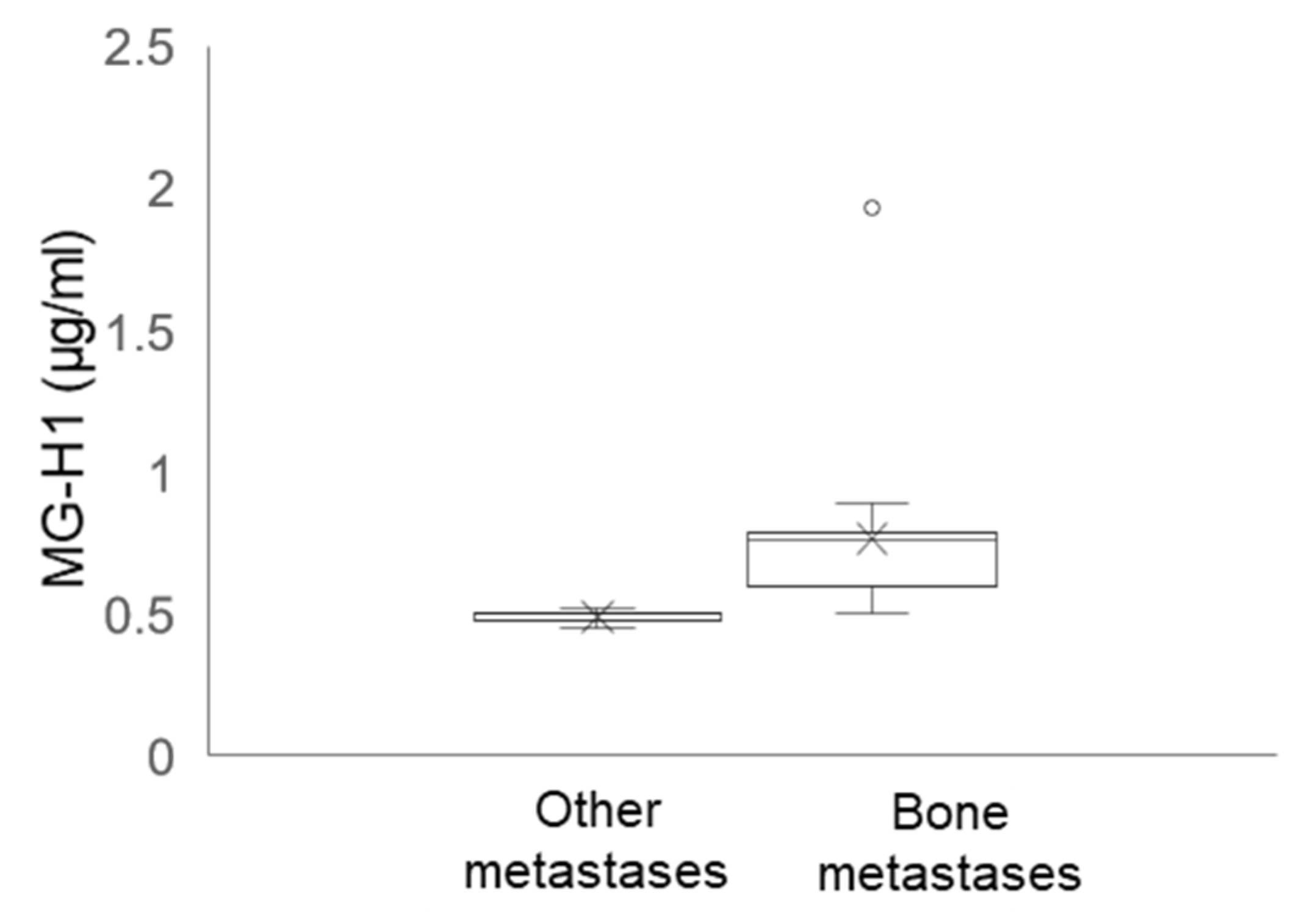
2.12. Circulating Levels of MG-H1 in Bone Metastasis-Bearing Patients
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Cultures
4.3. Preparation of Conditioned Medium (CM) and Treatments
4.4. MG-H1 Detection
4.5. Cell Lysis and Western Blot
4.6. Cell Viability and Morphology
4.7. RNA Isolation, Reverse Transcription, and Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) Analyses
4.8. Migration, Invasion and Cell Counting Assays
4.9. Glo1-Specific Enzyme Activity
4.10. Alizarin Red Staining
4.11. Reactive Oxidative Species (ROS) Measurement
4.12. Circulating Levels of MG-H1 in Samples from Metastasis-bearing Patients
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rucci, N.; Teti, A. Osteomimicry: How the Seed Grows in the Soil. Calcif. Tissue Int. 2017, 102, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Bonollo, F.; Thalmann, G.N.; Julio, M.K.-D.; Karkampouna, S. The Role of Cancer-Associated Fibroblasts in Prostate Cancer Tumorigenesis. Cancers 2020, 12, 1887. [Google Scholar] [CrossRef] [PubMed]
- Paget, S. The distribution of secondary growths in cancer of the breast. Cancer Metastasis Rev. 1989, 8, 98–101. [Google Scholar] [CrossRef] [Green Version]
- Bussard, K.; Gay, C.V.; Mastro, A.M. The bone microenvironment in metastasis; what is special about bone? Cancer Metastasis Rev. 2007, 27, 41–55. [Google Scholar] [CrossRef] [PubMed]
- Shupp, A.B.; Kolb, A.D.; Mukhopadhyay, D.; Bussard, K.M. Cancer Metastases to Bone: Concepts, Mechanisms, and Interactions with Bone Osteoblasts. Cancers 2018, 10, 182. [Google Scholar] [CrossRef] [Green Version]
- Quiroz-Munoz, M.; Izadmehr, S.; Arumugam, D.; Wong, B.; Kirschenbaum, A.; Levine, A.C. Mechanisms of Osteoblastic Bone Metastasis in Prostate Cancer: Role of Prostatic Acid Phosphatase. J. Endocr. Soc. 2019, 3, 655–664. [Google Scholar] [CrossRef] [Green Version]
- Keller, E.T.; Zhang, J.; Cooper, C.R.; Smith, P.C.; McCauley, L.K.; Pienta, K.; Taichman, R.S. Prostate Carcinoma Skeletal Metastases: Cross-talk between Tumor and Bone. Cancer Metastasis Rev. 2001, 20, 333–349. [Google Scholar] [CrossRef] [Green Version]
- Ottewell, P.D. The role of osteoblasts in bone metastasis. J. Bone Oncol. 2016, 5, 124–127. [Google Scholar] [CrossRef] [Green Version]
- Koeneman, K.S.; Yeung, F.; Chung, L.W.K. Osteomimetic properties of prostate cancer cells: A hypothesis supporting the predilection of prostate cancer metastasis and growth in the bone environment. Prostate 1999, 39, 246–261. [Google Scholar] [CrossRef]
- Carducci, M.A.; Nelson, J.B.; Bowling, M.K.; Rogers, T.; Eisenberger, M.A.; Sinibaldi, V.; Donehower, R.; Leahy, T.L.; Carr, R.A.; Isaacson, J.D.; et al. Atrasentan, an Endothelin-Receptor Antagonist for Refractory Adenocarcinomas: Safety and Pharmacokinetics. J. Clin. Oncol. 2002, 20, 2171–2180. [Google Scholar] [CrossRef]
- Carducci, M.A.; Padley, R.J.; Breul, J.; Vogelzang, N.J.; Zonnenberg, B.A.; Daliani, D.D.; Schulman, C.C.; Nabulsi, A.A.; Humerickhouse, R.A.; Weinberg, M.A.; et al. Effect of Endothelin-A Receptor Blockade with Atrasentan on Tumor Progression in Men With Hormone-Refractory Prostate Cancer: A Randomized, Phase II, Placebo-Controlled Trial. J. Clin. Oncol. 2003, 21, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Quinn, D.I.; Tangen, C.M.; Hussain, M.; Lara, P.N.; Goldkorn, A.; Moinpour, C.M.; Garzotto, M.G.; Mack, P.C.; Carducci, M.A.; Monk, J.P.; et al. Docetaxel and atrasentan versus docetaxel and placebo for men with advanced castration-resistant prostate cancer (SWOG S0421): A randomised phase 3 trial. Lancet Oncol. 2013, 14, 893–900. [Google Scholar] [CrossRef] [Green Version]
- Antognelli, C.; Talesa, V.N. Glyoxalases in Urological Malignancies. Int. J. Mol. Sci. 2018, 19, 415. [Google Scholar] [CrossRef] [Green Version]
- Marinucci, L.; Balloni, S.; Fettucciari, K.; Bodo, M.; Talesa, V.N.; Antognelli, C. Nicotine induces apoptosis in human osteoblasts via a novel mechanism driven by H 2 O 2 and entailing Glyoxalase 1-dependent MG-H1 accumulation leading to TG2-mediated NF-kB desensitization: Implication for smokers-related osteoporosis. Free Radic. Biol. Med. 2018, 117, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Ray, R.; Singer, D.; Böhme, D.; Burz, D.S.; Rai, V.; Hoffmann, R.; Shekhtman, A. The Receptor for Advanced Glycation End Products (RAGE) Specifically Recognizes Methylglyoxal-Derived AGEs. Biochemistry 2014, 53, 3327–3335. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Jacobs, K.; Haucke, E.; Santos, A.N.; Grune, T.; Simm, A. Role of advanced glycation end products in cellular signaling. Redox Biol. 2014, 2, 411–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.-R.; Lee, K.-W. Methylglyoxal-Derived Advanced Glycation End Product (AGE4)-Induced Apoptosis Leads to Mitochondrial Dysfunction and Endoplasmic Reticulum Stress through the RAGE/JNK Pathway in Kidney Cells. Int. J. Mol. Sci. 2021, 22, 6530. [Google Scholar] [CrossRef] [PubMed]
- Suh, K.S.; Chon, S.; Choi, E.M. Protective effects of honokiol against methylglyoxal-induced osteoblast damage. Chem. Interact. 2016, 244, 169–177. [Google Scholar] [CrossRef]
- Franke, S.; Rüster, C.; Pester, J.; Hofmann, G.; Oelzner, P.; Wolf, G. Advanced glycation end products affect growth and function of osteoblasts. Clin. Exp. Rheumatol. 2011, 29, 650–660. [Google Scholar]
- Antognelli, C.; Mezzasoma, L.; Fettucciari, K.; Talesa, V.N. A novel mechanism of methylglyoxal cytotoxicity in prostate cancer cells. Int. J. Biochem. Cell Biol. 2013, 45, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Mezzasoma, L.; Fettucciari, K.; Mearini, E.; Talesa, V.N. Role of glyoxalase I in the proliferation and apoptosis control of human LNCaP and PC3 prostate cancer cells. Prostate 2013, 73, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Talesa, V.N.; Ferri, I.; Bellezza, G.; Love, H.D.; Sidoni, A.; Antognelli, C. Glyoxalase 2 Is Involved in Human Prostate Cancer Progression as Part of a Mechanism Driven By PTEN/PI3K/AKT/mTOR Signaling with Involvement of PKM2 and ERα. Prostate 2017, 77, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Cecchetti, R.; Riuzzi, F.; Peirce, M.J.; Talesa, V.N. Glyoxalase 1 sustains the metastatic phenotype of prostate cancer cells via EMT control. J. Cell. Mol. Med. 2018, 22, 2865–2883. [Google Scholar] [CrossRef]
- Antognelli, C.; Mandarano, M.; Prosperi, E.; Sidoni, A.; Talesa, V. Glyoxalase-1-Dependent Methylglyoxal Depletion Sustains PD-L1 Expression in Metastatic Prostate Cancer Cells: A Novel Mechanism in Cancer Immunosurveillance Escape and a Potential Novel Target to Overcome PD-L1 Blockade Resistance. Cancers 2021, 13, 2965. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.K.; Jeon, J.M.; Sung, E.; Na, A.-Y.; Kim, S.J.; Lee, S. Comparative Secretome Profiling and Mutant Protein Identification in Metastatic Prostate Cancer Cells by Quantitative Mass Spectrometry-based Proteomics. Cancer Genom.-Proteom. 2018, 15, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Bear, J.E.; Haugh, J.M. Directed migration of mesenchymal cells: Where signaling and the cytoskeleton meet. Curr. Opin. Cell Biol. 2014, 30, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, M.E.; Rodriguez, A.G.; Speckl, K.F.; Walker, C.J.; Midekssa, F.S.; Grim, J.C.; Weiss, R.M.; Anseth, K.S. Collagen networks within 3D PEG hydrogels support valvular interstitial cell matrix mineralization. Acta Biomater. 2021, 119, 197–210. [Google Scholar] [CrossRef]
- Kinner, B.; Spector, M. Expression of smooth muscle actin in osteoblasts in human bone. J. Orthop. Res. 2002, 20, 622–632. [Google Scholar] [CrossRef]
- Torii, Y.; Hitomi, K.; Tsukagoshi, N. Synergistic effect of BMP-2 and ascorbate on the phenotypic expression of osteoblastic MC3T3-E1 cells. Mol. Cell. Biochem. 1996, 165, 25–29. [Google Scholar] [CrossRef]
- Bonneau, C.; Eliès, A.; Kieffer, Y.; Bourachot, B.; Ladoire, S.; Pelon, F.; Hequet, D.; Guinebretière, J.-M.; Blanchet, C.; Vincent-Salomon, A.; et al. A subset of activated fibroblasts is associated with distant relapse in early luminal breast cancer. Breast Cancer Res. 2020, 22, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Brunner, M.; Mandier, N.; Gautier, T.; Chevalier, G.; Ribba, A.-S.; Guardiola, P.; Block, M.R.; Bouvard, D. β1 integrins mediate the BMP2 dependent transcriptional control of osteoblast differentiation and osteogenesis. PLoS ONE 2018, 13, e0196021. [Google Scholar] [CrossRef] [Green Version]
- Gioia, M.; Michaletti, A.; Scimeca, M.; Marini, M.; Tarantino, U.; Zolla, L.; Coletta, M. Simulated microgravity induces a cellular regression of the mature phenotype in human primary osteoblasts. Cell Death Discov. 2018, 4, 59. [Google Scholar] [CrossRef] [Green Version]
- Molagoda, I.M.N.; Karunarathne, W.A.H.M.; Choi, Y.H.; Park, E.K.; Jeon, Y.-J.; Lee, B.-J.; Kang, C.-H.; Kim, G.-Y. Fermented Oyster Extract Promotes Osteoblast Differentiation by Activating the Wnt/β-Catenin Signaling Pathway, Leading to Bone Formation. Biomolecules 2019, 9, 711. [Google Scholar] [CrossRef] [Green Version]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef]
- Shi, G.; Yang, F. Krüppel-like factor 1 (KLF1) promoted the proliferation, migration and invasion of human lens epithelial cells by enhancing the expression of Zinc Finger and BTB Domain Containing 7A (ZBTB7A) and activating Wnt/β-catenin pathway. Bioengineered 2021, 12, 4374–4384. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Ezura, Y.; Izu, Y.; Aryal, S.A.C.; Kawasaki, M.; Chantida, P.N.M.; Moriyama, K.; Noda, M.; Smriti, A.; Pawaputanon, C. Profilin Expression Is Regulated by Bone Morphogenetic Protein (BMP) in Osteoblastic Cells. J. Cell. Biochem. 2015, 117, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Gu, Y.; Jiang, C.; Chen, L. Osteonectin regulates the extracellular matrix mineralization of osteoblasts through P38 signaling pathway. J. Cell. Physiol. 2020, 235, 2220–2231. [Google Scholar] [CrossRef]
- White, K.; Chalaby, R.; Lowe, G.; Berlin, J.; Glackin, C.; Olabisi, R. Calcein Binding to Assess Mineralization in Hydrogel Microspheres. Polymers 2021, 13, 2274. [Google Scholar] [CrossRef] [PubMed]
- Cumming, A.P.; Hopmans, S.N.; Vukmirović-Popović, S.; Duivenvoorden, W.C. PSA affects prostate cancer cell invasion in vitro and induces an osteoblastic phenotype in bone in vivo. Prostate Cancer Prostatic Dis. 2011, 14, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Xiang, S.; Hu, X.; Mo, M.; Zhao, C.; Cai, Y.; Tong, S.; Jiang, H.; Chen, L.; Wang, Z.; et al. Prostate-specific antigen modulates the osteogenic differentiation of MSCs via the cadherin 11-Akt axis. Clin. Transl. Med. 2020, 10, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Viryasova, G.; Golenkina, E.; Hianik, T.; Soshnikova, N.; Dolinnaya, N.; Gaponova, T.; Romanova, Y.; Sud’Ina, G. Magic Peptide: Unique Properties of the LRR11 Peptide in the Activation of Leukotriene Synthesis in Human Neutrophils. Int. J. Mol. Sci. 2021, 22, 2671. [Google Scholar] [CrossRef]
- Mao, Y.X.; Cai, W.J.; Sun, X.Y.; Dai, P.P.; Li, X.M.; Wang, Q.; Huang, X.L.; He, B.; Wang, P.P.; Wu, G.; et al. RAGE-dependent mitochondria pathway: A novel target of silibinin against apoptosis of osteoblastic cells induced by advanced glycation end products. Cell Death Dis. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hinz, N.; Jücker, M. AKT in Bone Metastasis of Solid Tumors: A Comprehensive Review. Cancers 2021, 13, 2287. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Yu, J. Role of the bone microenvironment in bone metastasis of malignant tumors—Therapeutic implications. Cell. Oncol. 2020, 43, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Kessel, K.; Bernemann, C.; Bögemann, M.; Rahbar, K. Evolving Castration Resistance and Prostate Specific Membrane Antigen Expression: Implications for Patient Management. Cancers 2021, 13, 3556. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.; Gomez, A.W.; Armstrong, B.E.; Henner, A.; Stankunas, K. Sequential and Opposing Activities of Wnt and BMP Coordinate Zebrafish Bone Regeneration. Cell Rep. 2014, 6, 482–498. [Google Scholar] [CrossRef] [Green Version]
- Almeida, M.; Han, L.; Martin-Millan, M.; O’Brien, C.A.; Manolagas, S.C. Oxidative Stress Antagonizes Wnt Signaling in Osteoblast Precursors by Diverting β-Catenin from T Cell Factor- to Forkhead Box O-mediated Transcription. J. Biol. Chem. 2007, 282, 27298–27305. [Google Scholar] [CrossRef] [Green Version]
- Antognelli, C.; Ferri, I.; Bellezza, G.; Siccu, P.; Love, H.D.; Talesa, V.N.; Sidoni, A. Glyoxalase 2 drives tumorigenesis in human prostate cells in a mechanism involving androgen receptor and p53-p21 axis. Mol. Carcinog. 2017, 56, 2112–2126. [Google Scholar] [CrossRef]
- Antognelli, C.; Trapani, E.; Monache, S.D.; Perrelli, A.; Daga, M.; Pizzimenti, S.; Barrera, G.; Cassoni, P.; Angelucci, A.; Trabalzini, L.; et al. KRIT1 loss-of-function induces a chronic Nrf2-mediated adaptive homeostasis that sensitizes cells to oxidative stress: Implication for Cerebral Cavernous Malformation disease. Free Radic. Biol. Med. 2018, 115, 202–218. [Google Scholar] [CrossRef]
- Antognelli, C.; Trapani, E.; Monache, S.D.; Perrelli, A.; Fornelli, C.; Retta, F.; Cassoni, P.; Talesa, V.N.; Retta, S.F. Data in support of sustained upregulation of adaptive redox homeostasis mechanisms caused by KRIT1 loss-of-function. Data Brief 2018, 16, 929–938. [Google Scholar] [CrossRef]
- Monache, S.D.; Pulcini, F.; Frosini, R.; Mattei, V.; Talesa, V.; Antognelli, C. Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants 2021, 10, 716. [Google Scholar] [CrossRef] [PubMed]
- Antognelli, C.; Frosini, R.; Santolla, M.F.; Peirce, M.J.; Talesa, V.N. Oleuropein-Induced Apoptosis Is Mediated by Mitochondrial Glyoxalase 2 in NSCLC A549 Cells: A Mechanistic Inside and a Possible Novel Nonenzymatic Role for an Ancient Enzyme. Oxidative Med. Cell. Longev. 2019, 2019, 8576961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antognelli, C.; Mancuso, F.; Frosini, R.; Arato, I.; Calvitti, M.; Calafiore, R.; Talesa, V.N.; Luca, G. Testosterone and Follicle Stimulating Hormone–Dependent Glyoxalase 1 Up-Regulation Sustains the Viability of Porcine Sertoli Cells through the Control of Hydroimidazolone– and Argpyrimidine-Mediated NF-κB Pathway. Am. J. Pathol. 2018, 188, 2553–2563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antognelli, C.; Mezzasoma, L.; Mearini, E.; Talesa, V.N. Glyoxalase 1−419C>A Variant Is Associated with Oxidative Stress: Implications in Prostate Cancer Progression. PLoS ONE 2013, 8, e74014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Xia, F.; Wei, Y.; Wei, X. Molecular mechanisms and clinical management of cancer bone metastasis. Bone Res. 2020, 8, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kreps, L.; Addison, C. Targeting Intercellular Communication in the Bone Microenvironment to Prevent Disseminated Tumor Cell Escape from Dormancy and Bone Metastatic Tumor Growth. Int. J. Mol. Sci. 2021, 22, 2911. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| VIM | GCACACAGCAAGGCGATGG | GGAGCGAGAGTGGCAGAGG |
| α-SMA | GGCATCATCACCAACTGGGACGAC | AGCACCGCCTGGATAGCCACATAC |
| TGF-β1 | GGCGACCCACAGAGAGGAAATAG | AGGCAGAAATTGGCGTGGTAGC |
| CDH11 | TGGCAGCAAGTATCCAATGG | TTTGGTTACGTGGTAGGCAC |
| ITGB1 | TGATTGGCTGGAGGAATGTTA | GTTTCTGGACAAGGTGAGCAA |
| ITGA3 | GGACCTTACAACGCCGAGTG | GGAGGCTCTTTGGCTTGTTTT |
| Runx2 | GCTCTTCCCAAAGCCAGAGT | ATCCTGAC-GAAGTGCCAT |
| Col1α1 | GAGGGCCAAGACGAAGACATC | CAGATCACGTCATCGCACAAC |
| ON | CCTGGAGACAAGGTGCTAACAT | CGAGTTCTCAGCCTGTGAGA |
| OSX | CCACCTACCCATCTGACT | GTTTGGCTCCACCACTCC |
| CD44 | AGTCCCTGGATCACCGA | CCTCTTGGTTGCTGTCTCA |
| FASC | CTGGCTACACGCTGGAGTTC | CTGAGTCCCCTGCTGTCTCC |
| PFN1 | TGGAGCAAACCCTACCCTT | AGCCCAGACACCGAACTTT |
| COFN | ATGCCCTCTATGATGCAACC | GCTTGATCCCTGTCAGCTTC |
| RADX | GAATCAGGAGCAGCTAGCAGCAGAACTT | TTGGTCTTTTCCAAGTCTTCCTGGGCTGCA |
| OC | TCACACTCCTCGCCCTATTGG | TCACACTCCTCGCCCTATTGG |
| OP | AGACCCCAAAAGTAAGGAAGAAG | GACAACCGTGGGAAAACAAATAAG |
| Glo1 | CTCTCCAGAAAAGCTACACTTTGAG | CGAGGGTCTGAATTGCCATTG |
| β-actin | CACTCTTCCAGCCTTCCTTCC | ACAGCACTGTGTTGGCGTAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Antognelli, C.; Marinucci, L.; Frosini, R.; Macchioni, L.; Talesa, V.N. Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases. Int. J. Mol. Sci. 2021, 22, 10191. https://doi.org/10.3390/ijms221910191
Antognelli C, Marinucci L, Frosini R, Macchioni L, Talesa VN. Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases. International Journal of Molecular Sciences. 2021; 22(19):10191. https://doi.org/10.3390/ijms221910191
Chicago/Turabian StyleAntognelli, Cinzia, Lorella Marinucci, Roberta Frosini, Lara Macchioni, and Vincenzo Nicola Talesa. 2021. "Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases" International Journal of Molecular Sciences 22, no. 19: 10191. https://doi.org/10.3390/ijms221910191
APA StyleAntognelli, C., Marinucci, L., Frosini, R., Macchioni, L., & Talesa, V. N. (2021). Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases. International Journal of Molecular Sciences, 22(19), 10191. https://doi.org/10.3390/ijms221910191