
Differential Polarization Imaging of Plant Cells. Mapping the Anisotropy of Cell Walls and Chloroplasts

 ,
,  ,
, 
Abstract
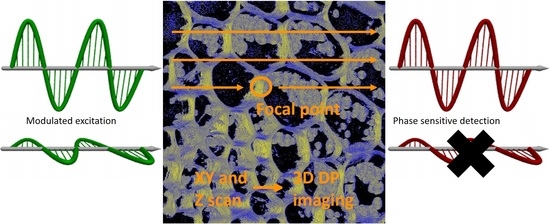
1. Introduction
2. Differential Polarization Imaging Theory and Devices
2.1. Advantages and Need of Spectropolarimetry Techniques in LSMs
2.1.1. Light-Matter Interactions—Mueller Matrix
2.1.2. Polarization State of the Fluorescence
2.1.3. Polarization Selective Interaction
2.2. DP Spectroscopy and Microscopy
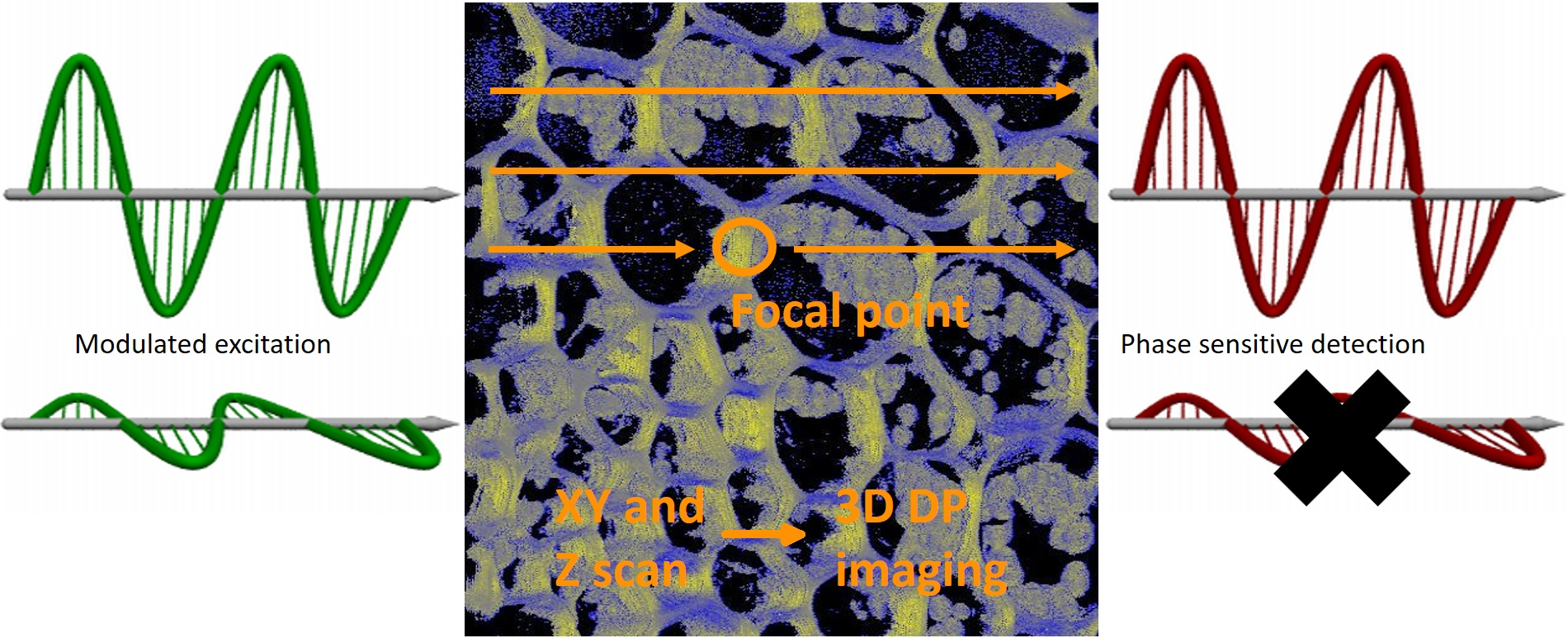
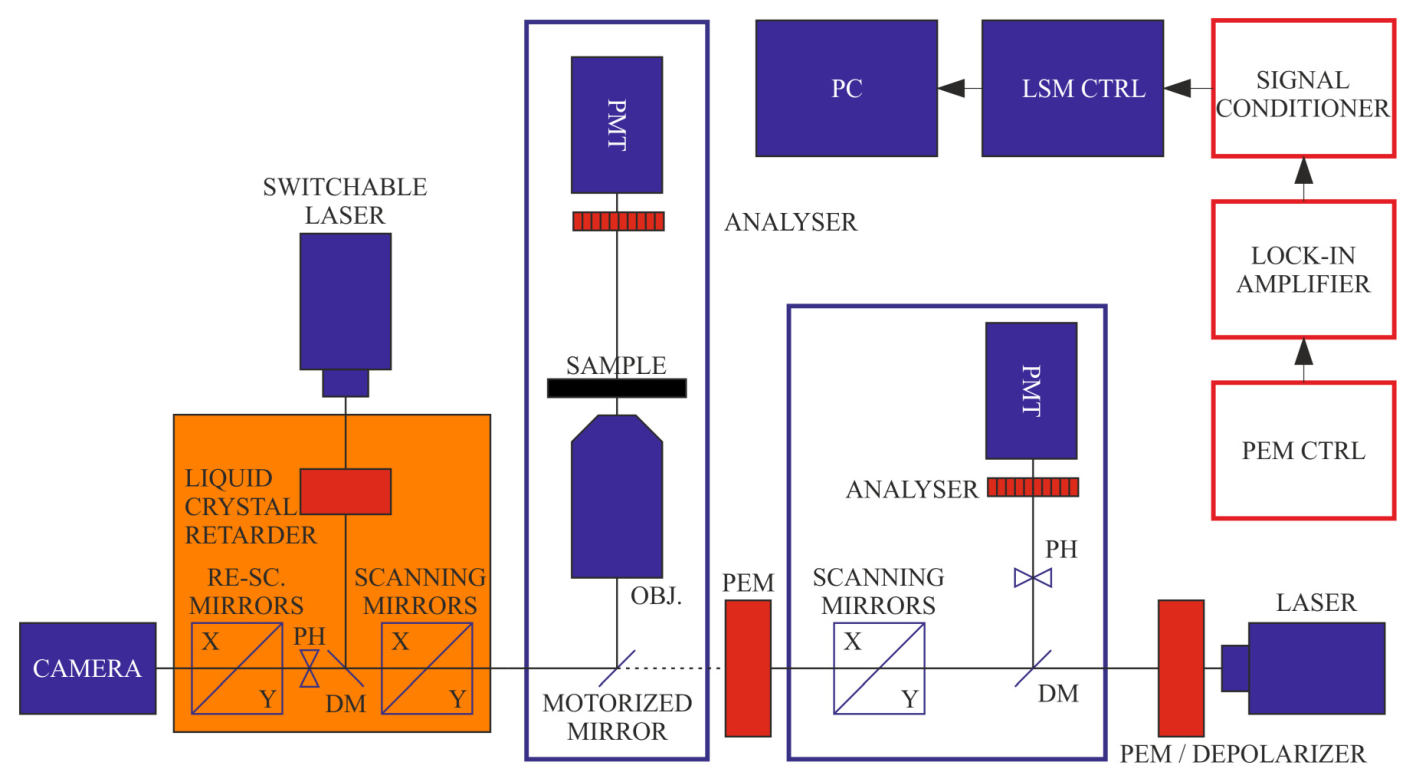
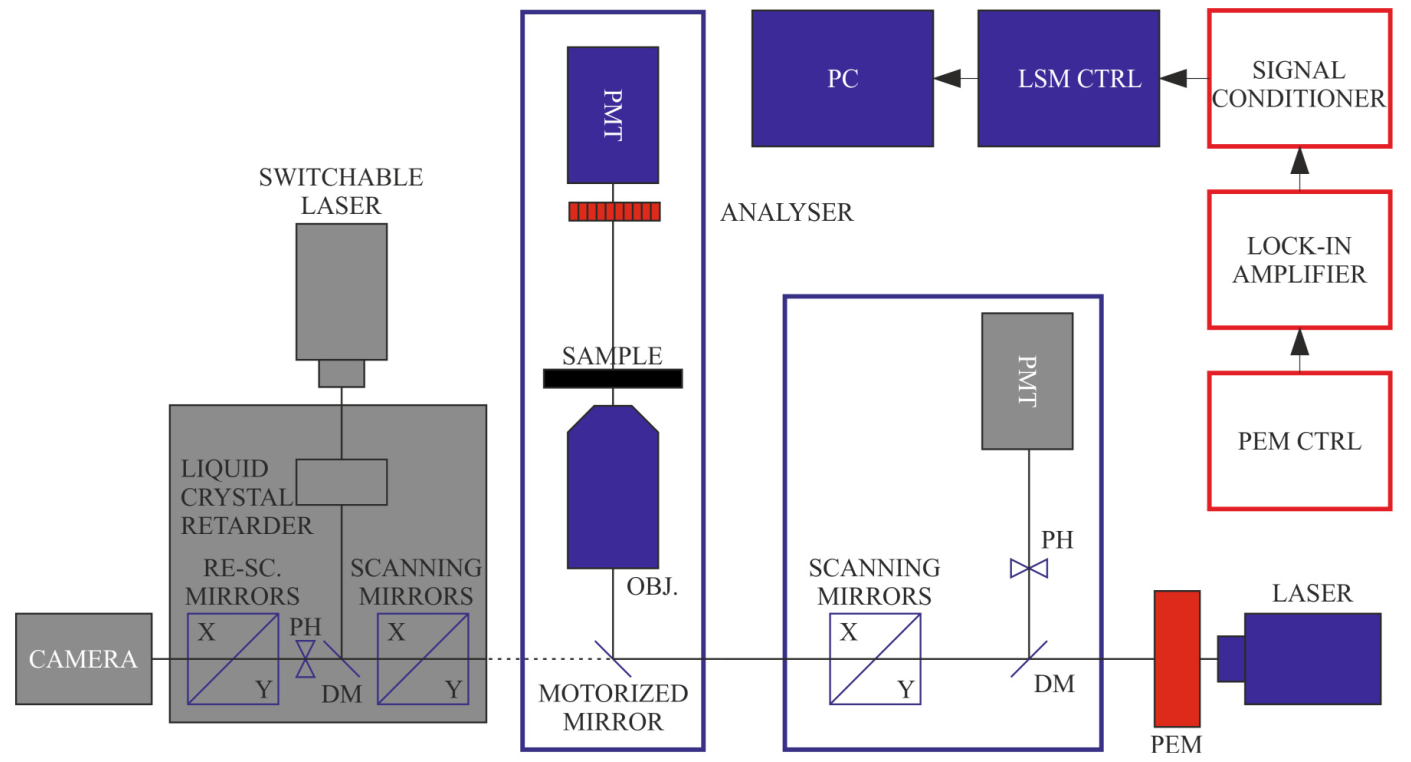
2.3. DP Imaging Devices
2.4. Information Content of DP Images
3. Applications in Plant Cell Research
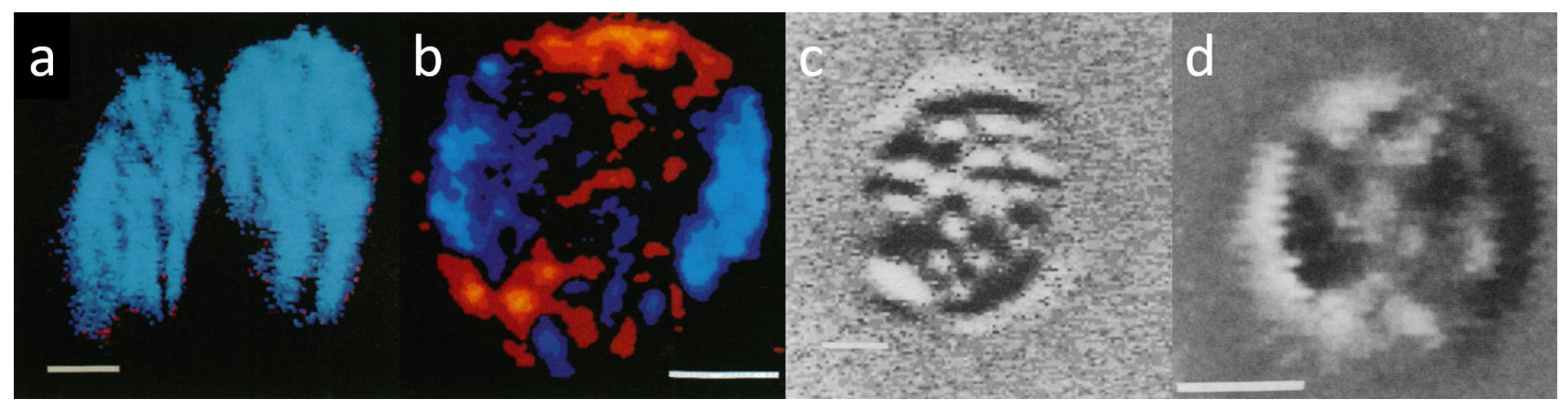
3.1. Anisotropy and DP Imaging of Granal Thylakoid Membranes
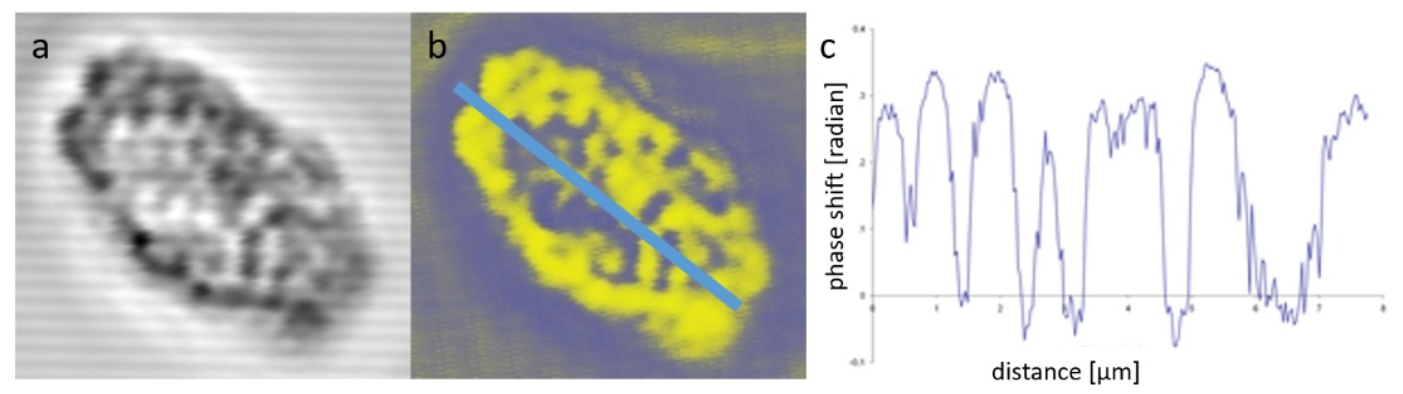
3.2. The Significance of Confocal DP Imaging for Plant Cell Wall Research
3.2.1. Convallaria Majalis Root Cell Walls
3.2.2. Comparison of Cell Wall Structure in Stems of Different Plant Species
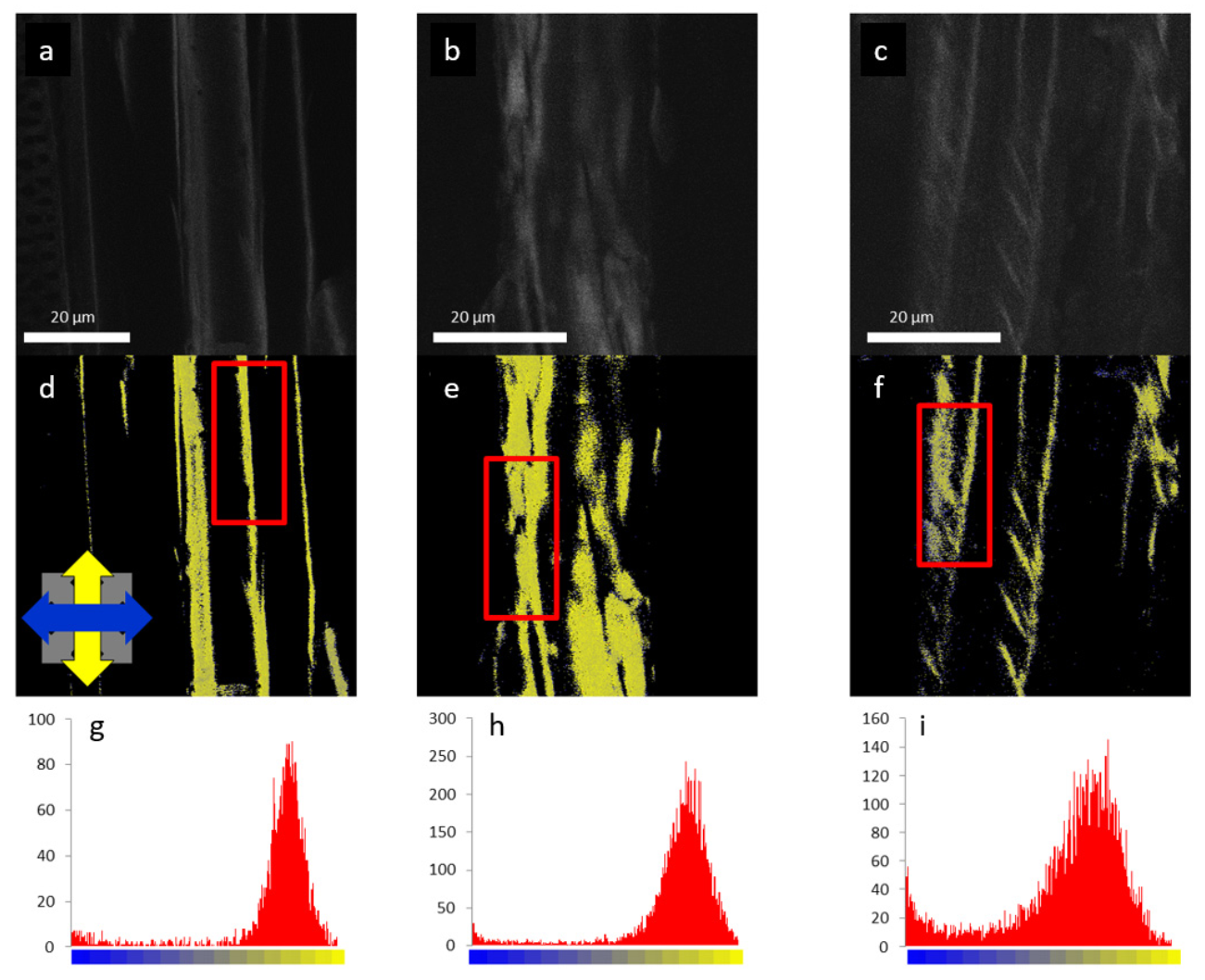
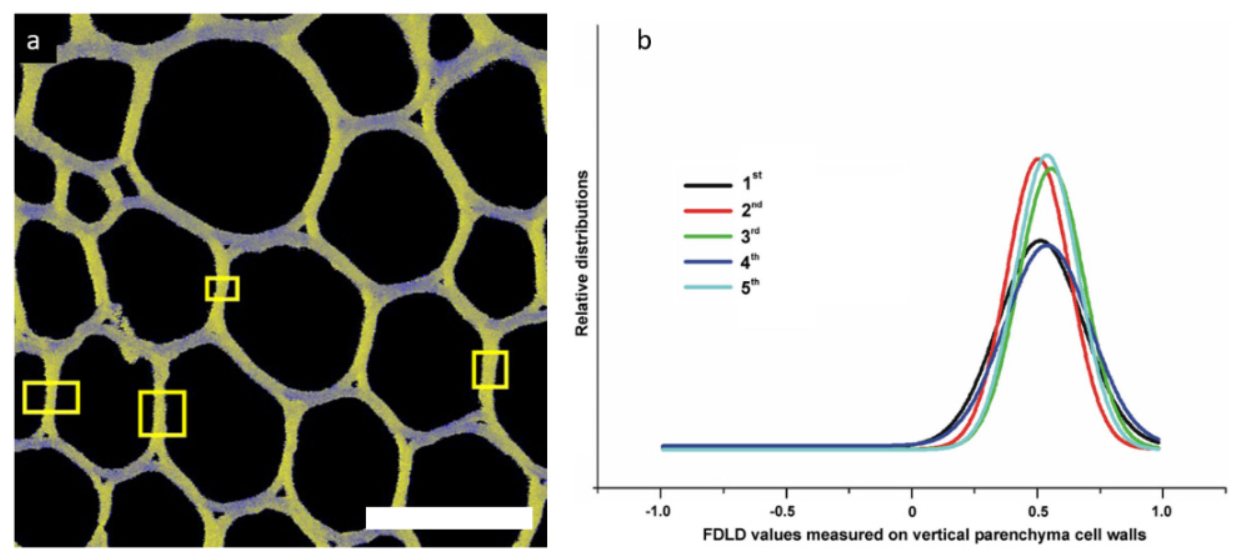
3.2.3. Parenchyma Cell Wall Structure in Liana Plants
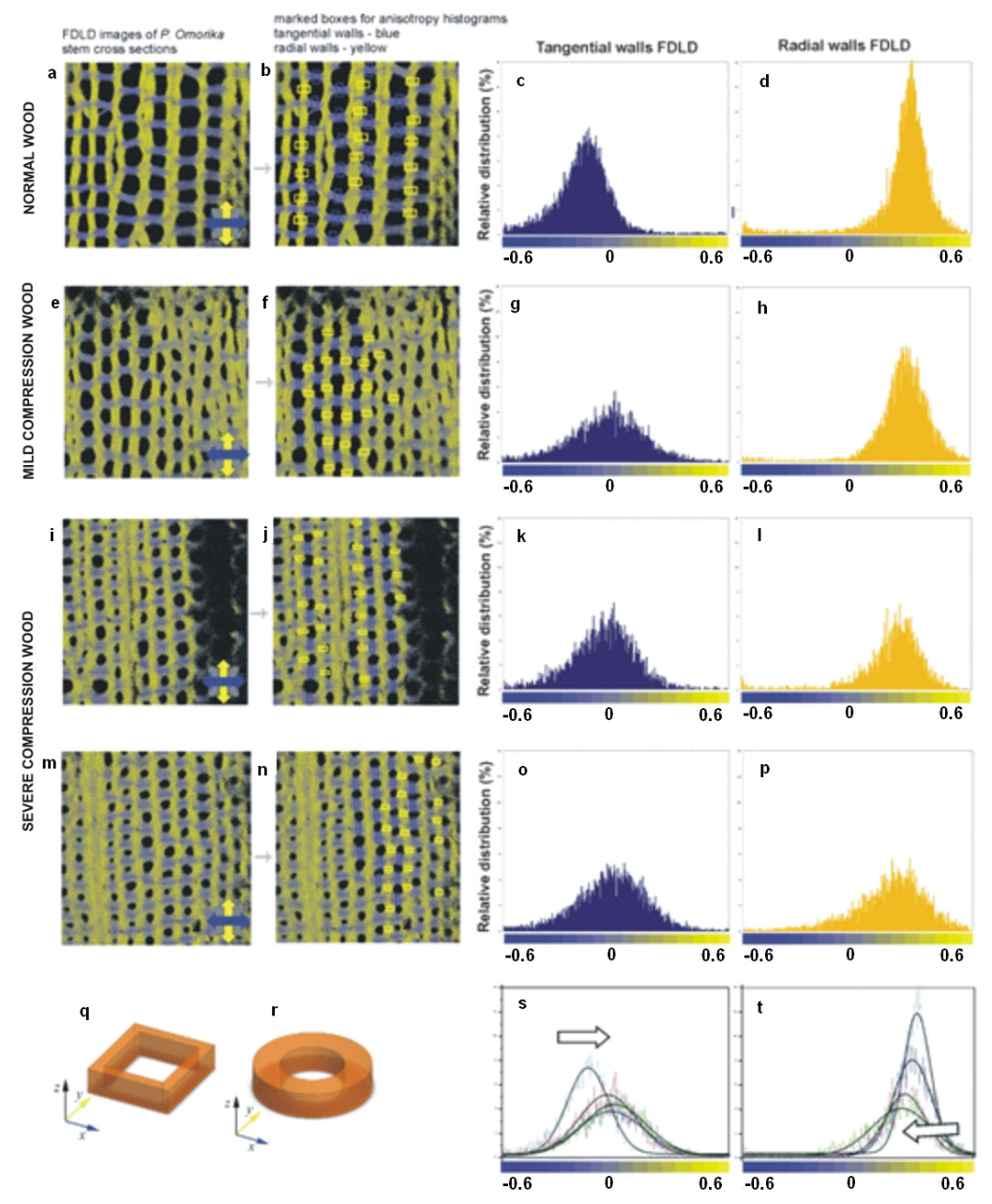
3.2.4. Cell Wall Response to Mechanical Stress
4. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abbe, E. Beiträge zur Theorie des Mikroskops und der mikroskopischen Wahrnehmung. Arch. Für Mikrosk. Anat. 1873, 9, 413–418. [Google Scholar] [CrossRef]
- Airy, G.B. On the diffraction of an object-glass with circular aperture. Trans. Cambridge Phil. Soc. 1835, 5, 283–291. [Google Scholar]
- Webb, R.H. Confocal optical microscopy. Rep. Prog. Phys. 1996, 59, 427–471. [Google Scholar] [CrossRef]
- Diaspro, A.F.M.; Sapuppo, P. Confocal Microscopy; Leica Microsystems CMS GmbH: Mannheim, Germany, 2008. [Google Scholar]
- De Luca, G.M.R.; Breedijk, R.M.P.; Brandt, R.A.J.; Zeelenberg, C.H.C.; de Jong, B.E.; Timmermans, W.; Azar, L.N.; Hoebe, R.A.; Stallinga, S.; Manders, E.M.M. Re-scan confocal microscopy: Scanning twice for better resolution. Biomed. Opt. Express. 2013, 4, 2644–2656. [Google Scholar] [CrossRef]
- Huszka, G.; Gijs, M.A.M. Super-resolution optical imaging: A comparison. Micro. Nano. Eng. 2019, 2, 7–28. [Google Scholar] [CrossRef]
- Datta, R.; Heaster, T.M.; Sharick, J.T.; Gillette, A.A.; Skala, M.C. Fluorescence lifetime imaging microscopy: Fundamentals and advances in instrumentation, analysis, and applications. J. Biomed. Opt. 2020, 25, 071203. [Google Scholar] [CrossRef]
- Fish, K.N. Total Internal Reflection Fluorescence. Curr. Protoc. Cytom. 2009, 50, 12–18. [Google Scholar]
- Gierlinger, N.; Schwanninger, M. The potential of Raman microscopy and Raman imaging in plant research. Spectrosc. Int. J. 2007, 21, 69–89. [Google Scholar] [CrossRef]
- Steinbach, G.; Pawlak, K.; Pomozi, I.; Toth, E.A.; Molnar, A.; Matko, J.; Garab, G. Mapping microscopic order in plant and mammalian cells and tissues: Novel differential polarization attachment for new generation confocal microscopes (DP-LSM). Methods Appl. Fluores. 2014, 2, 015005. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, G.; Pomozi, I.; Zsiros, O.; Menczel, L.; Garab, G. Imaging anisotropy using differential polarization laser scanning confocal microscopy. Acta Histochem. 2009, 111, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, G.; Nagy, D.; Sipka, G.; Manders, E.; Garab, G.; Zimanyi, L. Fluorescence-detected linear dichroism imaging in a re-scan confocal microscope equipped with differential polarization attachment. Eur. Biophys. J. Biophy. 2019, 48, 457–463. [Google Scholar] [CrossRef]
- Kim, M.; Keller, D.; Bustamante, C. Differential Polarization Imaging.1. Theory. Biophys. J. 1987, 52, 911–927. [Google Scholar] [CrossRef]
- Kim, M.; Bustamante, C. Differential Polarization Imaging.4. Images in Higher Born Approximations. Biophys. J. 1991, 59, 1171–1182. [Google Scholar] [CrossRef]
- Mueller, H. The foundation of optics. J. Opt. Soc. Am. 1948, 38, 661. [Google Scholar]
- Tinoco, I.; Mickols, W.; Maestre, M.F.; Bustamante, C. Absorption, Scattering, and Imaging of Biomolecular Structures with Polarized-Light. Annu. Rev. Biophys. Bio. 1987, 16, 319–349. [Google Scholar] [CrossRef]
- Patty, C.H.L.; Luo, D.A.; Snik, F.; Ariese, F.; Buma, W.J.; ten Kate, I.L.; van Spanning, R.J.M.; Sparks, W.B.; Germer, T.A.; Garab, G.; et al. Imaging linear and circular polarization features in leaves with complete Mueller matrix polarimetry. Bba-Gen Subj. 2018, 1862, 1350–1363. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy; Springer: New York, NY, USA, 2006. [Google Scholar]
- Garab, G. Linear and Circular Dichroism; Kluwer Academic Publishers: Amsterdam, The Netherlands, 1996. [Google Scholar]
- Gombos, I.; Steinbach, G.; Pomozi, I.; Balogh, A.; Vamosi, G.; Gansen, A.; Laszlo, G.; Garab, G.; Matko, J. Some new faces of membrane microdomains: A complex confocal fluorescence, differential polarization, and FCS imaging study on live immune cells. Cytom. Part A 2008, 73, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Kuball, H.G. CD and ACD spectroscopy on anisotropic samples: Chirality of oriented molecules and anisotropic phases—A critical analysis. Enantiomer 2002, 7, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, P.; Lindorfer, D.; Lingvay, M.; Pawlak, K.; Zsiros, O.; Siligardi, G.; Javorfi, T.; Dorogi, M.; Ughy, B.; Garab, G.; et al. Anisotropic Circular Dichroism of Light-Harvesting Complex II in Oriented Lipid Bilayers: Theory Meets Experiment. J. Phys. Chem. B 2019, 123, 1090–1098. [Google Scholar] [CrossRef]
- Mickols, W.; Tinoco, I.; Katz, J.E.; Maestre, M.F.; Bustamante, C. Imaging Differential Polarization Microscope with Electronic Readout. Rev. Sci. Instrum. 1985, 56, 2228–2236. [Google Scholar] [CrossRef]
- Finzi, L.; Bustamante, C.; Garab, G.; Juang, C.B. Direct Observation of Large Chiral Domains in Chloroplast Thylakoid Membranes by Differential Polarization Microscopy. Proc. Natl. Acad. Sci. USA 1989, 86, 8748–8752. [Google Scholar] [CrossRef]
- Gupta, V.K.; Kornfield, J.A. Polarization Modulation Laser-Scanning Microscopy—A Powerful Tool to Image Molecular-Orientation and Order. Rev. Sci. Instrum. 1994, 65, 2823–2828. [Google Scholar] [CrossRef][Green Version]
- Garab, G.; Pomozi, I.; Weiss, G.; Jörgens, R. Method and Apparatus for Determining the Polarization Properties of Light Emitted, Reflected or Transmitted by a Material Using a Laser Scanning Microscope. U.S. Patent No. 6,856,391, 15 February 2005. [Google Scholar]
- Gorjanacz, M.; Torok, I.; Pomozi, I.; Garab, G.; Szlanka, T.; Kiss, I.; Mechler, B.M. Domains of importin-alpha 2 required for ring canal assembly during Drosophila oogenesis. J. Struct. Biol. 2006, 154, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Ulibarri, L.; Bustamante, C. Differential Polarization Imaging.2. Symmetry Properties and Calculations. Biophys. J. 1987, 52, 929–946. [Google Scholar] [CrossRef]
- Steinbach, G.; Besson, F.; Pomozi, I.; Garab, G. Differential polarization laser scanning microscopy: Biological applications. In Photonic Applications in Biosensing and Imaging; International Society for Optics and Photonics: San Diego, CA, USA, 2005; Volume 5969, pp. 566–575. [Google Scholar]
- Lazar, J.; Bondar, A.; Timr, S.; Firestein, S.J. Two-photon polarization microscopy reveals protein structure and function. Nat. Methods 2011, 8, 684–690. [Google Scholar] [CrossRef]
- Wang, X.; Kress, A.; Brasselet, S.; Ferrand, P. High frame-rate fluorescence confocal angle-resolved linear dichroism microscopy. Rev. Sci. Instrum. 2013, 84, 053708. [Google Scholar] [CrossRef] [PubMed]
- Hafi, N.; Grunwald, M.; van den Heuvel, L.S.; Aspelmeier, T.; Chen, J.H.; Zagrebelsky, M.; Schutte, O.M.; Steinem, C.; Korte, M.; Munk, A.; et al. Fluorescence nanoscopy by polarization modulation and polarization angle narrowing. Nat. Methods 2014, 11, 579–584. [Google Scholar] [CrossRef] [PubMed]
- Mazumder, N.; Qiu, J.J.; Kao, F.J.; Diaspro, A. Mueller matrix signature in advanced fluorescence microscopy imaging. J. Opt. UK 2017, 19, 025301. [Google Scholar] [CrossRef]
- Loison, O.; Weitkunat, M.; Kaya-Copur, A.; Alves, C.N.; Matzat, T.; Spletter, M.L.; Luschnig, S.; Brasselet, S.; Lenne, P.F.; Schnorrer, F. Polarization-resolved microscopy reveals a muscle myosin motor-independent mechanism of molecular actin ordering during sarcomere maturation. Plos. Biol. 2018, 16, e2004718. [Google Scholar] [CrossRef]
- Blankenship, R.E. Molecular Mechanisms of Photosynthesis; Wiley-Blackwell: Oxford, UK, 2014. [Google Scholar]
- Garab, G.; van Amerongen, H. Linear dichroism and circular dichroism in photosynthesis research. Photosynth. Res. 2009, 101, 135–146. [Google Scholar] [CrossRef]
- Keller, D.; Bustamante, C. Theory of the Interaction of Light with Large Inhomogeneous Molecular Aggregates.2. Psi-Type Circular-Dichroism. J. Chem. Phys. 1986, 84, 2972–2980. [Google Scholar] [CrossRef]
- Mustardy, L.; Garab, G. Granum revisited. A three-dimensional model—Where things fall into place. Trends Plant Sci. 2003, 8, 117–122. [Google Scholar] [CrossRef]
- Mustardy, L.; Buttle, K.; Steinbach, G.; Garab, G. The Three-Dimensional Network of the Thylakoid Membranes in Plants: Quasihelical Model of the Granum-Stroma Assembly. Plant Cell 2008, 20, 2552–2557. [Google Scholar] [CrossRef]
- Garab, G.; Galajda, P.; Pomozi, I.; Finzi, L.; Praznovszky, T.; Ormos, P.; van Amerongen, H. Alignment of biological microparticles by a polarized laser beam. Eur. Biophys. J. Biophy. 2005, 34, 335–343. [Google Scholar] [CrossRef]
- Verbelen, J.P.; Kerstens, S. Polarization confocal microscopy and Congo Red fluorescence: A simple and rapid method to determine the mean cellulose fibril orientation in plants. J. Microsc. 2000, 198, 101–107. [Google Scholar] [CrossRef]
- Harris, P.J. Primary and secondary plant cell walls: A comparative overview. N. Z. J. For. Sci. 2006, 36, 36–53. [Google Scholar]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 204. [Google Scholar] [CrossRef] [PubMed]
- Timell, T.E. Compression Wood in Gymnosperms; Springer: Heidelberg, Germany, 1986. [Google Scholar]
- Donaldson, L.; Xu, P. Microfibril orientation across the secondary cell wall of Radiata pine tracheids. Trees-Struct. Funct. 2005, 19, 644–653. [Google Scholar] [CrossRef]
- Barnett, J.R.; Bonham, V.A. Cellulose microfibril angle in the cell wall of wood fibres. Biol. Rev. 2004, 79, 461–472. [Google Scholar] [CrossRef]
- Schaffer, J.B.H. Test Preparation for Microscopes. U.S. Patent 20040180384, 12 December 2002. [Google Scholar]
- Baskin, T.I.; Meekes, H.T.H.M.; Liang, B.M.; Sharp, R.E. Regulation of growth anisotropy in well-watered and water-stressed maize roots. II. Role of cortical microtubules and cellulose microfibrils. Plant Physiol. 1999, 119, 681–692. [Google Scholar] [CrossRef]
- Steinbach, G.; Pomozi, I.; Zsiros, O.; Pay, A.; Horvath, G.V.; Garab, G. Imaging fluorescence detected linear dichroism of plant cell walls in laser scanning confocal microscope. Cytom. Part A 2008, 73, 202–208. [Google Scholar] [CrossRef]
- Kerstens, S.; Verbelen, J.P. Cellulose orientation in the outer epidermal wall of angiosperm roots: Implications for biosystematics. Ann. Bot. Lond. 2002, 90, 669–676. [Google Scholar] [CrossRef]
- Djikanovic, D.; Devecerski, A.; Steinbach, G.; Simonovic, J.; Matovic, B.; Garab, G.; Kalauzi, A.; Radotic, K. Comparison of macromolecular interactions in the cell walls of hardwood, softwood and maize by fluorescence and FTIR spectroscopy, differential polarization laser scanning microscopy and X-ray diffraction. Wood Sci. Technol. 2016, 50, 547–566. [Google Scholar] [CrossRef]
- Radosavljevic, J.S.; Pristov, J.B.; Mitrovic, A.L.; Steinbach, G.; Mouille, G.; Tufegdzic, S.; Maksimovic, V.; Mutavdzic, D.; Janosevic, D.; Vukovic, M.; et al. Parenchyma cell wall structure in twining stem of Dioscorea balcanica. Cellulose 2017, 24, 4653–4669. [Google Scholar] [CrossRef]
- Savic, A.; Mitrovic, A.; Donaldson, L.; Radosavljevic, J.S.; Pristov, J.B.; Steinbach, G.; Garab, G.; Radotic, K. Fluorescence-Detected Linear Dichroism of Wood Cell Walls in Juvenile Serbian Spruce: Estimation of Compression Wood Severity. Microsc. Microanal. 2016, 22, 361–367. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Chappaz-Gillot, C.; Marek, P.L.; Blaive, B.J.; Canard, G.; Burck, J.; Garab, G.; Hahn, H.; Javorfi, T.; Kelemen, L.; Krupke, R.; et al. Anisotropic Organization and Microscopic Manipulation of Self-Assembling Synthetic Porphyrin Microrods That Mimic Chlorosomes: Bacterial Light-Harvesting Systems. J. Am. Chem. Soc. 2012, 134, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Garab, G. Hierarchical organization and structural flexibility of thylakoid membranes. Biochim. Biophys. Acta (BBA) Bioenerg. 2014, 1837, 481–494. [Google Scholar] [CrossRef]
- Banas, A.K.; Aggarwal, C.; Labuz, J.; Sztatelman, O.; Gabrys, H. Blue light signalling in chloroplast movements. J. Exp. Bot. 2012, 63, 1559–1574. [Google Scholar] [CrossRef]
- Wada, M. Chloroplast movement. Plant Sci 2013, 210, 177–182. [Google Scholar] [CrossRef]
- Kost, B.; Chua, N.H. The plant cytoskeleton: Vacuoles and cell walls make the difference. Cell 2002, 108, 9–12. [Google Scholar] [CrossRef]
- Friese, M.E.J.; Nieminen, T.A.; Heckenberg, N.R.; Rubinsztein-Dunlop, H. Optical alignment and spinning of laser-trapped microscopic particles. Nature 1998, 394, 348–350. [Google Scholar] [CrossRef]
- Steinbach, G.; Pomozi, I.; Janosa, D.P.; Makovitzky, J.; Garab, G. Confocal Fluorescence Detected Linear Dichroism Imaging of Isolated Human Amyloid Fibrils. Role of Supercoiling. J. Fluoresc. 2011, 21, 983–989. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DP Quantity | Laser Beam Polarization | Detection | Definition | Information Content |
|---|---|---|---|---|
| LD | Modulated | Nonpolarized. Transmission: non-confocal | Linear dichroism A1 − A2 ≈ (I2 − I1)/2.3 I0 | Anisotropic organization of molecular macro-assemblies (absorption dipoles) |
| FDLD | Modulated | Nonpolarized. Fluorescence: confocal | Fluorescence-detected linear dichroism I1(A‖) − I2(A⟂) = ΔI | LD of fluorophores (absorption dipoles) |
| r | Unpolarized | Modulated. Fluorescence: confocal | Anisotropy of the emission dipoles: (I1 − I2)/Ia | Anisotropic organization of molecular macro-assemblies (emitting dipoles) |
| P | Linearly polarized | Modulated. Fluorescence: confocal | Degree of polarization of the fluorescence emission: (I1 − I2)/(I1 + I2) | Microviscosity, fluorescence lifetime, energy transfer |
| LB | Modulated | Polarized | Phase-shift determination | Anisotropic ultrastructure or texture |
| DP Quantity | Materials/Objects | Plant Biology | Publications |
|---|---|---|---|
| LD | Convallaria majalis root cell wall | yes | [49] |
| FDLD | Acer platanoides L., Picea omorika, Zea mays L. | yes | [51] |
| Dioscorea balcanica | yes | [52] | |
| Isolated human amyloid | no | [61] | |
| Synthetic porphyrin microrods (Zn-C2) | no | [55] | |
| r | Synthetic porphyrin microrods (Zn-C2) | no | [55] |
| Drosophila melanogaster nurse cells ring canals | no | [27] | |
| P | JY human B lymphoblasts | no | [20] |
| LB | Chloroplasts isolated from Pisum sativum | yes | [29] |
| FDLD (on DP-RCM) | Convallaria majalis root cell wall | yes | [12] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Radosavljević, J.S.; Mitrović, A.L.; Radotić, K.; Zimányi, L.; Garab, G.; Steinbach, G. Differential Polarization Imaging of Plant Cells. Mapping the Anisotropy of Cell Walls and Chloroplasts. Int. J. Mol. Sci. 2021, 22, 7661. https://doi.org/10.3390/ijms22147661
Radosavljević JS, Mitrović AL, Radotić K, Zimányi L, Garab G, Steinbach G. Differential Polarization Imaging of Plant Cells. Mapping the Anisotropy of Cell Walls and Chloroplasts. International Journal of Molecular Sciences. 2021; 22(14):7661. https://doi.org/10.3390/ijms22147661
Chicago/Turabian StyleRadosavljević, Jasna Simonović, Aleksandra Lj. Mitrović, Ksenija Radotić, László Zimányi, Győző Garab, and Gábor Steinbach. 2021. "Differential Polarization Imaging of Plant Cells. Mapping the Anisotropy of Cell Walls and Chloroplasts" International Journal of Molecular Sciences 22, no. 14: 7661. https://doi.org/10.3390/ijms22147661
APA StyleRadosavljević, J. S., Mitrović, A. L., Radotić, K., Zimányi, L., Garab, G., & Steinbach, G. (2021). Differential Polarization Imaging of Plant Cells. Mapping the Anisotropy of Cell Walls and Chloroplasts. International Journal of Molecular Sciences, 22(14), 7661. https://doi.org/10.3390/ijms22147661