Genome Wide Association Study of Karnal Bunt Resistance in a Wheat Germplasm Collection from Afghanistan

 , , ,
, , ,
Abstract
1. Introduction
2. Results
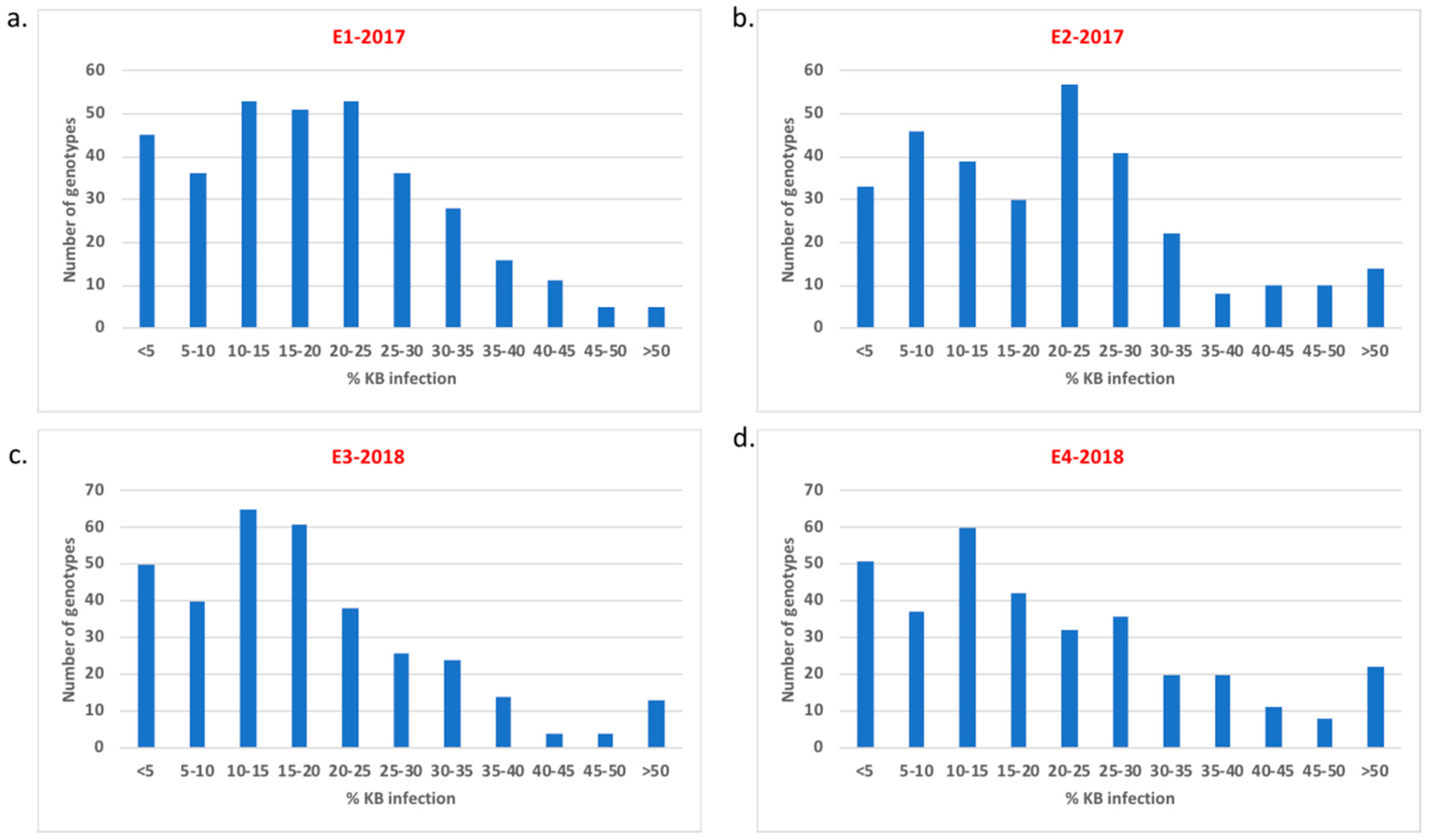
2.1. Karnal Bunt Resistance in the Germplasm Collection
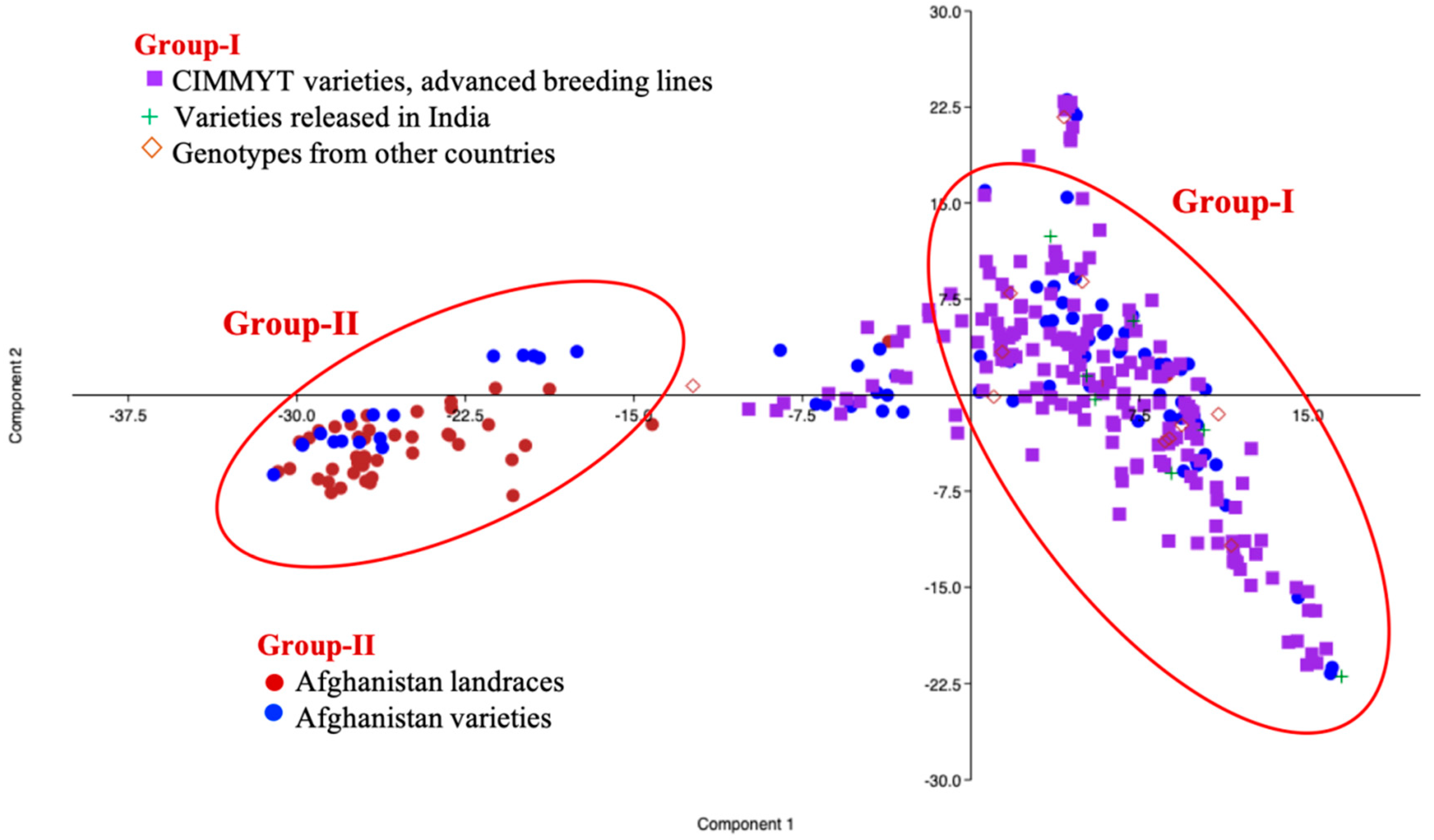
2.2. Population Structure
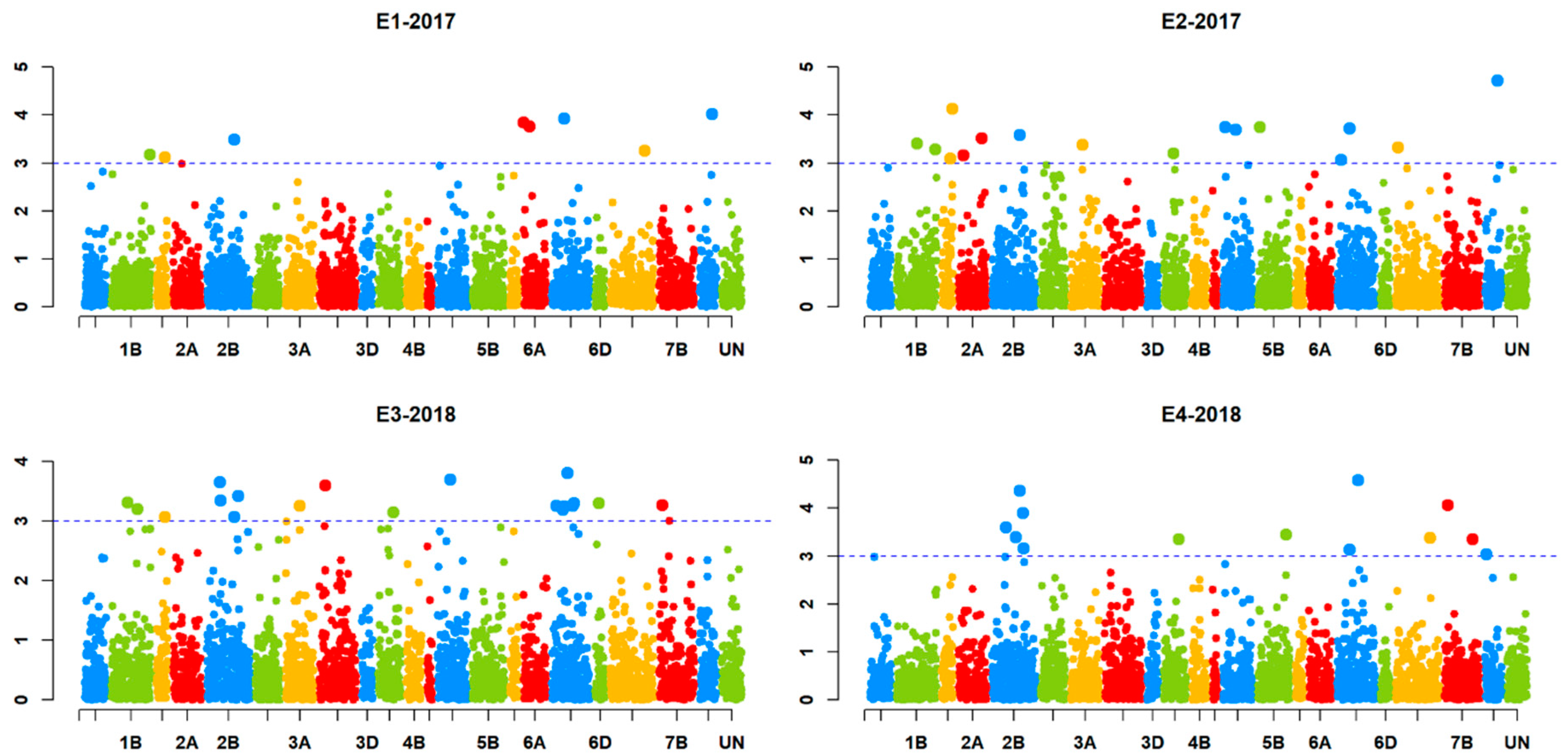
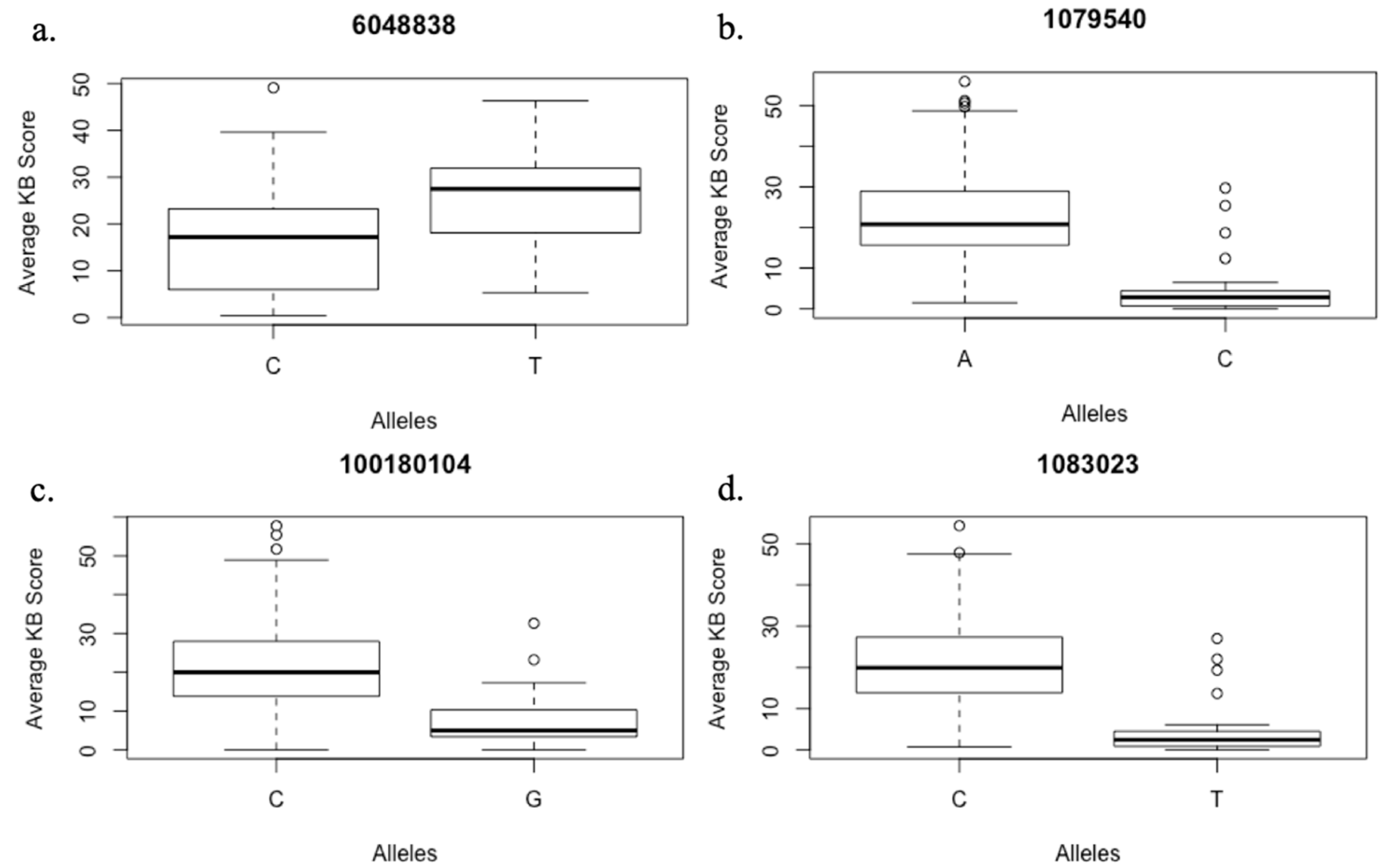
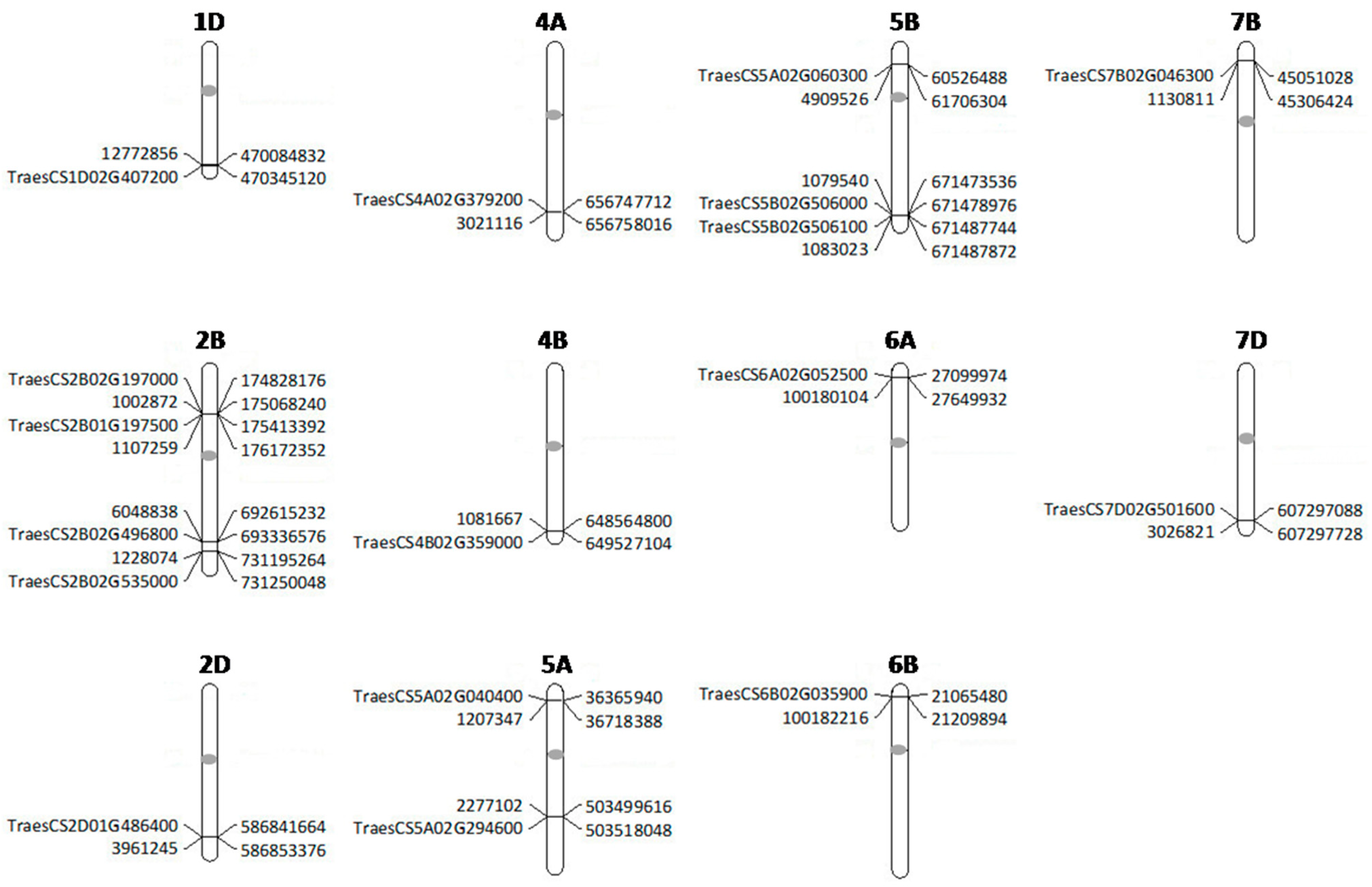
2.3. Markers Significantly Associated with Karnal Bunt
2.4. Putative Genes Associated with MTAs for Karnal Bunt
3. Discussion
4. Materials and Methods
4.1. Experimental Plant Material and Field Trials
4.2. Phenotyping for Karnal Bunt Infection
4.2.1. Inoculum Preparation
4.2.2. Karnal Bunt Inoculations and Disease Scoring
4.3. Statistical Analysis
4.4. Genotypic Data and GWAS
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Kumar, S.; Mishra, C.N.; Gupta, V.; Singh, R.; Sharma, I. Molecular characterization and yield evaluation of near isogenic line (NIL) of wheat cultivar PBW 343 developed for Karnal bunt resistance. Indian Phytopathol. 2016, 69, 119–123. [Google Scholar]
- Mitra, M. A new bunt on wheat in India. Ann. Appl. Biol. 1931, 18, 178–179. [Google Scholar] [CrossRef]
- Singh, D.V. First Report of Tilletia indica on wheat in Nepal. Plant. Dis. 1989, 73, 273. [Google Scholar] [CrossRef]
- Torabi, M.; Mardoukhi, V.; Jaliani, N. First report on the occurrence of partial bunt on wheat in the southern parts of Iran. Seed Plant 1996, 12, 8–9. [Google Scholar]
- Commonwealth Mycological Institute. Distribution Maps Plant Disease, 3rd ed.; Commonwealth Agricultural Bureaux: Kew, UK, 1974. [Google Scholar]
- Locke, C.M.; Watson, A.J. Foreign plant diseases intercepted in quarantine inspection. Plant Dis. Rep. 1955, 39, 518. [Google Scholar]
- Duran, R.; Cromarty, R. Tilletia indica: A heterothallic wheat fungus with multiple alleles controlling incompatibility. Phytopathology 1977, 67, 812–815. [Google Scholar] [CrossRef]
- Ykema, R.E.; Floyd, J.P.; Palm, M.E.; Peterson, G.L. First report of Karnal bunt of wheat in the United States. Plant Dis. 1996, 80, 1207. [Google Scholar] [CrossRef]
- Crous, P.W.; VanJarsveld, A.B.; Castlebury, L.A.; Caris, L.M.; Frederick, R.D.; Pretorius, Z.A. Karnal bunt of wheat newly reported from the African continent. Plant Dis. 2001, 85, 561. [Google Scholar] [CrossRef]
- DaLuz, W.C.; Mendes, M.A.S.; Fereira, M.A.S.V.; Urben, A.F. Tilletia indica on wheat in the southern part of Rio Grande do Sul and means for its eradication. Fitopatol. Bras. 1993, 18, 329. (In Portuguese) [Google Scholar]
- Stansbury, C.D.; McKirdy, S.J.; Diggle, A.J.; Riley, I.T. Modeling the risk of entry, establishment, spread, containment, and economic impact of Tilletia indica, the cause of Karnal bunt of wheat, using an Australian context. Phytopathology 2002, 92, 321–331. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Alam, K.; Rolfe, J. Economics of plant disease outbreaks. Agenda 2006, 13, 133–146. [Google Scholar]
- Singh, S.L. Cultural practices and the Karnal bunt (Neovossia indica) incidence. Indian Phytopathol. 1985, 38, 594. [Google Scholar]
- Babadoost, M.; Mathre, D.E.; Johnston, R.H.; Bonde, M.R. Survival of teliospores of Tilletia indica in soil. Plant Dis. 2004, 88, 56–62. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sansford, C.E.; Baker, R.H.A.; Brennan, J.P.; Ewert, F.; Gioli, B.; Inman, A.; Kinsella, A.; Magnus, H.A.; Miglietta, F.; Murray, G.M.; et al. The new pest risk analysis for Tilletia indica, the cause of Karnal bunt of wheat, continues to support the quarantine status of the pathogen in Europe. Plant Pathol. 2008, 57, 603–611. [Google Scholar] [CrossRef]
- Wei-chuan, Z.; Gui-ming, Z. Prediction of potential epidemiological areas in China prone to Karnal bunt of wheat. J. Plant Pathol. 2010, 92, 367–373. [Google Scholar]
- Brennan, J.P.; Warham, E.J.; Hernandez, J.; Byerlee, D.; Doronel, F. Economic Losses from Karnal Bunt of Wheat in Mexico; 90/02; CIMMYT: Texcoco, Mexico, 1990. [Google Scholar]
- Vocke, G.; Allen, E.W.; Price, J.M. Economic Analysis of Ending the Issuance of Karnal Bunt Phytosanitary Wheat Export Certificates; Wheat Yearbook/WHS-2002; USDA Economic Research Service: Washington, DC, USA, 2002.
- Villareal, R.L.; Fuentes-Davila, G.; Mujeeb-Kazi, A.; Rajaram, S. Inheritance of resistance to Tilletia indica (Mitra) in Synthetic Hexaploid Wheat x Triticum aestivum crosses. Plant Breed. 1995, 114, 547–548. [Google Scholar] [CrossRef]
- Fuentes-Davila, G.; Rajaram, S.; Singh, G. Inheritance of resistance to Karnal bunt (Tilletia indica Mitra) in bread wheat (Triticum aestivum L.). Plant Breed. 1995, 114, 250–252. [Google Scholar] [CrossRef]
- Singh, H.; Grewal, T.S.; Pannu, P.P.S.; Dhaliwal, H.S. Genetics of resistance to Karnal bunt disease of wheat. Euphytica 1999, 105, 125–131. [Google Scholar] [CrossRef]
- Sharma, I.; Bains, N.S.; Nanda, G.S. Inheritance of Karnal bunt-free trait in bread wheat. Plant Breed. 2004, 123, 96–97. [Google Scholar] [CrossRef]
- Sharma, I.; Bains, N.S.; Singh, K.; Nanda, G.S. Additive genes at nine loci govern Karnal bunt resistance in a set of common wheat cultivars. Euphytica 2005, 142, 301–307. [Google Scholar] [CrossRef]
- Bala, R.; Kumar, S.; Bains, N.S.; Sharma, I. Development of disease resistant bread wheat (Triticum aestivum) line in background of PBW 343 and genetics of Karnal bunt free trait. Indian Phytopathol. 2015, 68, 42–44. [Google Scholar]
- Fuentes-Davila, G.; Rajaram, S. Sources of resistance to Tilletia indica in wheat. Crop Protect. 1994, 13, 20–24. [Google Scholar] [CrossRef]
- Kumar, S.; Singroha, G.; Bhardwaj, S.C.; Bala, R.; Saharan, M.S.; Gupta, V.; Khan, A.; Mahapatra, S.; Sivasamy, M.; Rana, V.; et al. Multi-environmental evaluation of wheat (Triticum aestivum L.) germplasm identifies donors with multiple fungal disease resistance. Genet. Resour. Crop Evol. 2019, 66, 797–808. [Google Scholar] [CrossRef]
- Chhuneja, P.; Kaur, S.; Singh, K.; Dhaliwal, H.S. Evaluation of Aegilops tauschii (Coss.) germplasm for Karnal bunt resistance in a screen house with simulated environmental conditions. Plant Genet. Resour. 2008, 6, 79–84. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A.; Fuentes-Davilla, G.; Gul, A.; Mirza, J.I. Karnal bunt resistance in Synthetic Hexaploid Wheats (SH) derived from Durum Wheat x Aegilops tauschii combinations and in some SH x bread wheat derivatives. Cereal Res. Commun. 2006, 34, 1199–1205. [Google Scholar] [CrossRef]
- Nelson, J.C.; Autrique, J.E.; Fuentes-Dávila, G.; Sorrells, M.E. Chromosomal location of genes for resistance to Karnal bunt in wheat. Crop Sci. 1998, 38, 231. [Google Scholar] [CrossRef]
- Sukhwinder-Singh; Brown-Guedira, G.; Grewal, T.; Dhaliwal, H.; Nelson, J.; Singh, H.; Gill, B. Mapping of a resistance gene effective against Karnal bunt pathogen of wheat. Theor. Appl. Genet. 2003, 106, 287–292. [Google Scholar] [CrossRef]
- Brooks, S.A.; See, D.R.; Brown-Guedira, G. SNP-based improvement of a microsatellite marker associated with Karnal bunt resistance in wheat. Crop Sci. 2006, 46, 1467. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, I.; Sehgal, S.K.; Bains, N.S.; Guo, Z.; Nelson, J.C.; Bowden, R.L. Molecular mapping of QTLs for Karnal bunt resistance in two recombinant inbred populations of bread wheat. Theor. Appl. Genet. 2007, 116, 147–154. [Google Scholar] [CrossRef]
- Kumar, S.; Chawla, V.; Yadav, N.R.; Sharma, I.; Yadav, P.K.; Kumar, S.; Behl, R.K. Identification and validation of SSR markers for Karnal bunt (Neovossia indica) resistance in wheat (Triticum aestivum). Indian Indian J. Agric. Sci. 2015, 85, 712–717. [Google Scholar]
- Kaur, M.; Singh, R.; Kumar, S.; Mandhan, R.P.; Sharma, I. Identification of QTL conferring Karnal bunt resistance in bread wheat. Indian J. Biotechnol. 2016, 15, 34–38. [Google Scholar]
- Bala, R.; Sharma, A.; Kashyap, L.; Rana, B.; Bains, N.S.; Sharma, I. Molecular mapping of QTLs for Karnal bunt resistance in six near isogenic (NILs) populations of bread wheat. Indian Phytopathol. 2016, 69, 242–246. [Google Scholar]
- Brar, G.S.; Fuentes-Dávila, G.; He, X.; Sansaloni, C.P.; Singh, R.P.; Singh, P.K. Genetic mapping of resistance in hexaploid wheat for a quarantine Disease: Karnal bunt. Front. Plant Sci. 2018, 9, 1497. [Google Scholar] [CrossRef] [PubMed]
- Emebiri, L.; Singh, S.; Tan, M.-K.; Singh, P.K.; Fuentes-Dávila, G.; Ogbonnaya, F. Unravelling the complex genetics of Karnal bunt (Tilletia indica) resistance in common wheat (Triticum aestivum) by genetic linkage and genome-wide association analyses. G3 2019, 9, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Rafalski, J.A. Association genetics in crop improvement. Curr. Opin. Plant Biol. 2010, 13, 174–180. [Google Scholar] [CrossRef]
- Sonah, H.; O’Donoughue, L.; Cober, E.; Rajcan, I.; Belzile, F. Identification of loci governing eight agronomic traits using a GBS-GWAS approach and validation by QTL mapping in soya bean. Plant Biotechnol. J. 2015, 13, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Buckler, E.S.; Thornsberry, J.M. Plant Molecular diversity and applications to genomics. Curr. Opin. Plant Biol. 2002, 5, 107–111. [Google Scholar] [CrossRef]
- Emebiri, L.; Singh, P.K.; Tan, M.-K.; Fuentes-Dávila, G.; He, X.; Singh, R.P. Reaction of Australian durum, common wheat and triticale varieties to Karnal bunt (Tilletia indica) infection under artificial inoculation in the field. Crop. Pasture Sci. 2019, 70, 107–112. [Google Scholar] [CrossRef]
- Dhaliwal, H.S.; Singh, D.V. Up-to-date life cycle of Neovossia indica (Mitra) Mundkur. Curr. Sci. 1989, 57, 675–677. [Google Scholar]
- Albert, E.; Segura, V.; Gricourt, J.; Bonnefoi, J.; Derivot, L.; Causse, M. Association mapping reveals the genetic architecture of tomato response to water deficit: Focus on major fruit quality traits. J. Exp. Bot. 2016, 67, 6413–6430. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, Z.; Ding, J.; Wu, Y.; Zhou, B.; Wang, R.; Ma, J.; Wang, S.; Zhang, X.; Xia, Z.; et al. Combined linkage and association mapping reveals QTL and candidate genes for plant and ear height in maize. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Raman, H.; Stodart, B.; Ryan, P.R.; Delhaize, E.; Emebiri, L.; Raman, R.; Coombes, N.; Milgate, A. Genome-wide association analyses of common wheat (Triticum aestivum L.) germplasm identifies multiple loci for aluminium resistance This Article Is One of a Selection of Papers from the Conference “Exploiting Genome-Wide Association in Oilseed Brassicas: A Model for Genetic Improvement of Major OECD Crops for Sustainable Farming”. Genome 2010, 53, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Mulki, M.A.; Jighly, A.; Ye, G.; Emebiri, L.C.; Moody, D.; Ansari, O.; Ogbonnaya, F.C. Association mapping for soilborne pathogen resistance in synthetic hexaploid wheat. Mol. Breed. 2013, 31, 299–311. [Google Scholar] [CrossRef]
- Marone, D.; Russo, M.; Laidò, G.; De Leonardis, A.; Mastrangelo, A. Plant nucleotide binding site–leucine-rich repeat (NBS-LRR) genes: Active guardians in host defense responses. Int. J. Mol. Sci. 2013, 14, 7302–7326. [Google Scholar] [CrossRef] [PubMed]
- Aujla, S.S.; Grewal, A.S.; Sharma, I. Artificial creation of Karnal bunt disease of wheat. Cereal Res. Commun. 1982, 10, 171–176. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Osman, M.; He, X.; Singh, R.P.; Duveiller, E.; Lillemo, M.; Pereyra, S.A.; Westerdijk-Hoks, I.; Kurushima, M.; Yau, S.-K.; Benedettelli, S.; et al. Phenotypic and genotypic characterization of CIMMYT’s 15th International Fusarium Head Blight screening nursery of wheat. Euphytica 2015, 205, 521–537. [Google Scholar] [CrossRef]
- Li, H.; Vikram, P.; Singh, R.P.; Kilian, A.; Carling, J.; Song, J.; Burgueno-Ferreira, J.A.; Bhavani, S.; Huerta-Espino, J.; Payne, T.; et al. A high density GBS map of bread wheat and its application for dissecting complex disease resistance traits. BMC Genom. 2015, 16, 216. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Chromosome | Linked Marker/ Interval | Reference |
|---|---|---|---|
| Altar 84 | 3BS, 5AL | RFLP | [30] |
| HD 29 | 4BL | Xgwm 538 | [31] |
| HD 29 | 4BL | SNP 52bp fragment of interest (gwm538 snp) | [32] |
| HD29 | Qkb.ksu-5BL.1 | Xgdm116-Xmc 235 | [33] |
| HD29 | Qkb.ksu-6BS.1 | Xwmc105- Xgwm 88 | |
| W485 | Qkb.ksu-4BL.1 | Xgwm 6- Xwmc 349 | |
| H567.71 | 4B | Xgwm 6 | [34] |
| ALDAN | Qkb.dwr-5BL.1 | Xwmc 235 and Xbarc 140 | [35] |
| HD29 | 5B | Xgdm116 - Xwmc235 | [36] |
| HD29 | 6B | Xwmc105 – Xgwm 88 | |
| W485 | 4B | Xgwm 6 – Xwmc 349 | |
| WKCBW | QKb.cim-2BL | 1086228 - 1092041 | [37] |
| WKCBW | QKb.cim-3DL | 7487658- 2252592 | |
| Huirivis#1 | QKb.cim-3BS1 | 1079551- 100010977 | |
| Mutus | QKb.cim-5BS2 | 2253589 - 1011847 | |
| HD29 | 3B | IWB57185 | [38] |
| WH542 | 1A | IWA1644 | |
| WH542 | 1D | IWB2650 | |
| W485 | 1B | IWB59865 |
| SN | Panel ID | Pedigree | E1-2017 | E2-2017 | E3-2018 | E4-2018 | Mean | *QTL |
|---|---|---|---|---|---|---|---|---|
| 1 | 498 | FAHAD-8-2*2//PTR/PND-T/3/ERIZO-11/YOGUI-3 | 0 | 0 | 0 | 0 | 0.0 | 1, 2, 3 |
| 2 | 516 | CMH80A.542/CNO79 | 0 | 0 | 0 | 0 | 0.0 | 1, 3, 4 |
| 3 | 540 | ANOAS-5/STIER-13/5/274/320//BGL.3.MUSX/ | 0 | 0.48 | 0 | 0 | 0.1 | 1, 2, 3, 4 |
| 4 | 506 | GAUR-2/HARE-3//JLO97/CIVET/5/DIS B5/3/SPHD/ | 0.65 | 0 | 0 | 0 | 0.2 | 1, 2, 3, 4 |
| 5 | 402 | ZCL/3/PGFN//CNO67/SON64(ES86-8)/4/SERI/5/UA-2827 | 0.61 | 0.53 | 0 | 0 | 0.3 | 1, 3 |
| 6 | 554 | GNU/ASAD//ARDI/3/MANATI-1/4/FAHAD-5 | 1.78 | 0 | 0 | 0 | 0.4 | 1, 2, 3, 4 |
| 7 | 509 | PRESTO//2*TESMO-1/MUSX 603/4/ARDI-1/ | 0 | 0 | 0 | 1.78 | 0.4 | 1, 2, 4, 5, 6 |
| 8 | 280 | GK.ZOMBOR/ATTILA | 1.64 | 0 | 0 | 0.42 | 0.5 | 1, 2, 4, 5, 7 |
| 9 | 547 | STIER-13/FAHAD-4//MANATI-1/3/POLLMER-1.1 | 2.05 | 0 | 0.4 | 0 | 0.6 | 1, 2, 4, 5 |
| 10 | 558 | FD-693/2*FAHAD-4//POLLMER-4/3/POLLMER-2.1 | 0 | 2.17 | 0.96 | 0 | 0.8 | 1, 2, 4 |
| 11 | 6 | AFGHAN Wheat Collection #6 | 0 | NA | 1.52 | 1.03 | 0.9 | 1, 2, 5, 8, 9, 10, 11, 12 |
| Source of Variation | Degree of Freedom | Mean Square | p Value | Heritability |
|---|---|---|---|---|
| Genotype | 338 | 550.71 | <0.001 | 0.80 |
| Experiment | 3 | 437.69 | <0.001 | |
| Genotype x Experiment | 1352 | 184.01 | <0.001 | |
| Error | 1014 | 61.78 |
| SN | Marker | Allele | Chromosome | Physical Position | E1- 2017 | E2- 2017 | E3- 2018 | E4- 2018 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Additve Effect | R2 | Additive Effect | R2 | Additive Effect | R2 | Additive Effect | R2 | |||||
| 1 | 12772856 | C/T | 1DL | 470084827 | −0.43 | 0.06 | - | - | −2.59 | 0.06 | - | - |
| 2 | 1002872 | A/C | 2BS | 175068236 | - | - | - | - | −5.72 | 0.06 | −6.29 | 0.04 |
| 3 | 1107259 | A/G | 2BS | 176172344 | - | - | - | - | −5.55 | 0.06 | −6.48 | 0.06 |
| 4 | 1228074 | A/G | 2BL | 731195296 | - | - | - | - | −3.47 | 0.06 | −1.25 | 0.05 |
| 5 | 6048838 | T/C | 2BL | 692615259 | −1.90 | 0.13 | −4.64 | 0.15 | −4.27 | 0.11 | −4.96 | 0.20 |
| 6 | 3961245 | T/C | 2DL | 586853396 | - | - | −1.75 | 0.06 | −1.54 | 0.05 | - | - |
| 7 | 3021116 | A/C | 4AL | 656758037 | - | - | - | - | 0.75 | 0.06 | 1.80 | 0.07 |
| 8 | 1081667 | A/G | 4BL | 648564812 | −4.13 | 0.05 | −2.96 | 0.05 | - | - | - | - |
| 9 | 1207347 | G/A | 5AS | 36718388 | −3.55 | 0.05 | −4.05 | 0.07 | - | - | - | - |
| 10 | 2277102 | T/C | 5AL | 503499615 | - | - | 0.46 | 0.06 | −0.57 | 0.06 | - | - |
| 11 | 1079540 | C/A | 5BL | 671473536 | 1.25 | 0.05 | 1.69 | 0.06 | 2.75 | 0.06 | - | - |
| 12 | 1083023 | C/T | 5BL | 671487862 | NaN | 0.06 | - | - | NaN | 0.06 | NaN | 0.08 |
| 13 | 4909526 | C/G | 5BS | 61706304 | 5.83 | 0.07 | - | - | - | - | 7.76 | 0.08 |
| 14 | 100180104 | G/C | 6AS | 27649931 | 4.69 | 0.07 | 3.32 | 0.06 | - | - | - | - |
| 15 | 4394191 | G/A | 6BL | 500595153 | - | - | 7.69 | 0.10 | 0.91 | 0.06 | - | - |
| 16 | 100182216 | T/G | 6BS | 21209894 | - | - | - | - | 4.87 | 0.07 | 7.08 | 0.09 |
| 17 | 1130811 | G/C | 7BS | 45306426 | - | - | - | - | −1.58 | 0.05 | −1.78 | 0.06 |
| 18 | 3026821 | C/G | 7DL | 607297738 | 3.10 | 0.06 | 2.96 | 0.08 | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gupta, V.; He, X.; Kumar, N.; Fuentes-Davila, G.; Sharma, R.K.; Dreisigacker, S.; Juliana, P.; Ataei, N.; Singh, P.K. Genome Wide Association Study of Karnal Bunt Resistance in a Wheat Germplasm Collection from Afghanistan. Int. J. Mol. Sci. 2019, 20, 3124. https://doi.org/10.3390/ijms20133124
Gupta V, He X, Kumar N, Fuentes-Davila G, Sharma RK, Dreisigacker S, Juliana P, Ataei N, Singh PK. Genome Wide Association Study of Karnal Bunt Resistance in a Wheat Germplasm Collection from Afghanistan. International Journal of Molecular Sciences. 2019; 20(13):3124. https://doi.org/10.3390/ijms20133124
Chicago/Turabian StyleGupta, Vikas, Xinyao He, Naresh Kumar, Guillermo Fuentes-Davila, Rajiv K. Sharma, Susanne Dreisigacker, Philomin Juliana, Najibeh Ataei, and Pawan K. Singh. 2019. "Genome Wide Association Study of Karnal Bunt Resistance in a Wheat Germplasm Collection from Afghanistan" International Journal of Molecular Sciences 20, no. 13: 3124. https://doi.org/10.3390/ijms20133124
APA StyleGupta, V., He, X., Kumar, N., Fuentes-Davila, G., Sharma, R. K., Dreisigacker, S., Juliana, P., Ataei, N., & Singh, P. K. (2019). Genome Wide Association Study of Karnal Bunt Resistance in a Wheat Germplasm Collection from Afghanistan. International Journal of Molecular Sciences, 20(13), 3124. https://doi.org/10.3390/ijms20133124