Nociceptor Signalling through ion Channel Regulation via GPCRs



Abstract
1. Introduction
- ATP as a prototypic nucleotide may activate a subset of ligand-gated ion channels known as P2X receptors [6].
- Cytokines such as various interleukins or tumor necrosis factors (TNFs) target different subtypes of cytokine receptors [7].
- Neurotrophins, in particular nerve growth factor, bind to high affinity tyrosine receptor kinases (trks) and to the low affinity receptor p75 [8].
- All others of the aforementioned inflammatory mediators and ATP elicit their actions on nociceptors via some type of G protein-coupled receptor (GPCR).
2. Ion Channels as Targets of GPCR Signaling in Peripheral Nociceptive Neurons
2.1. TRP Channels Involved in Pain Sensation
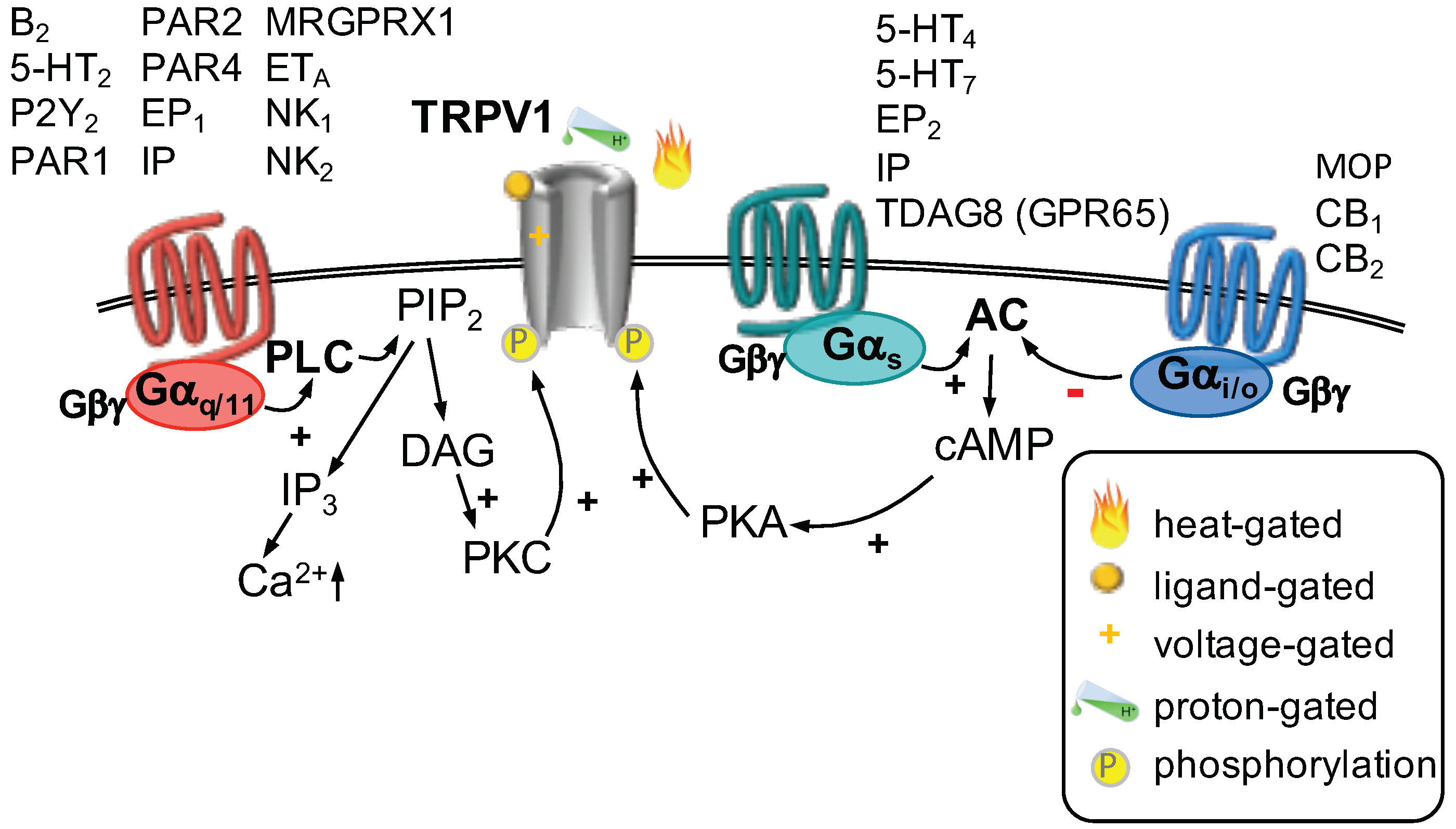
GPCR Regulation of TRP Channels
2.2. Acid-Sensing Ion Channels
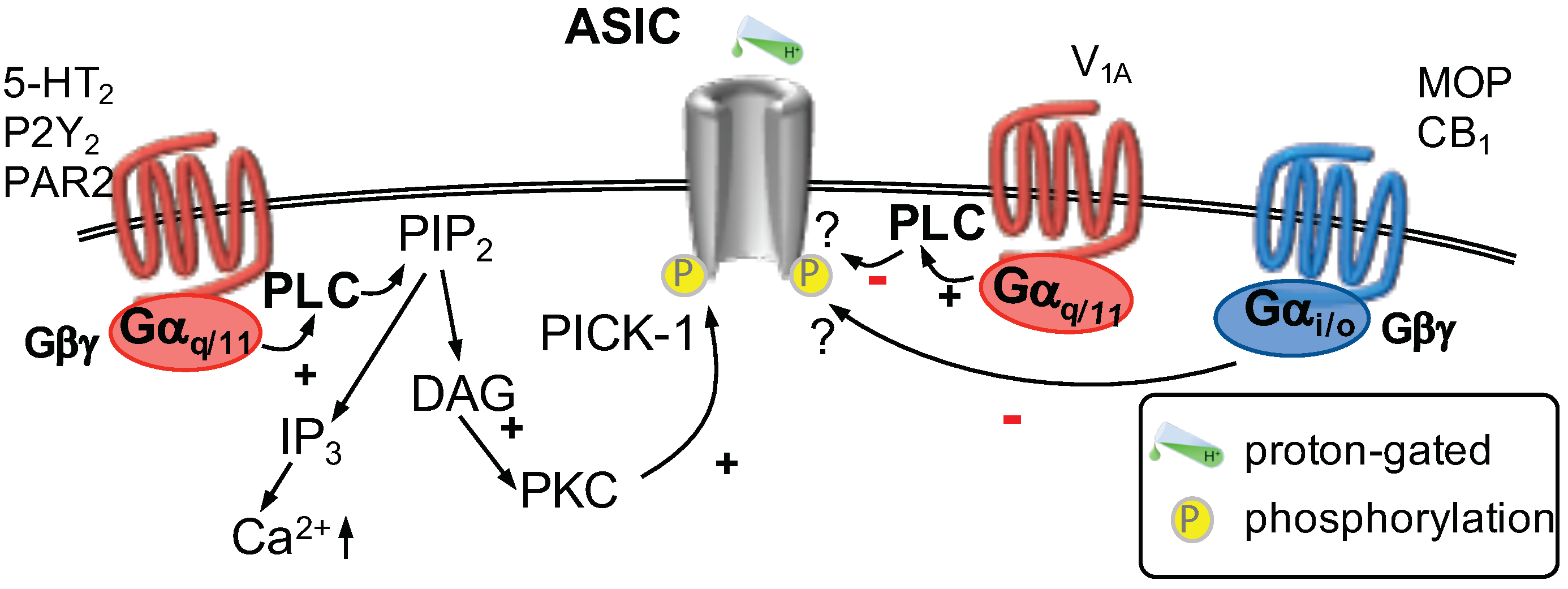
GPCR Regulation of ASICs
2.3. Mechanosensitive Channels in Pain Sensation
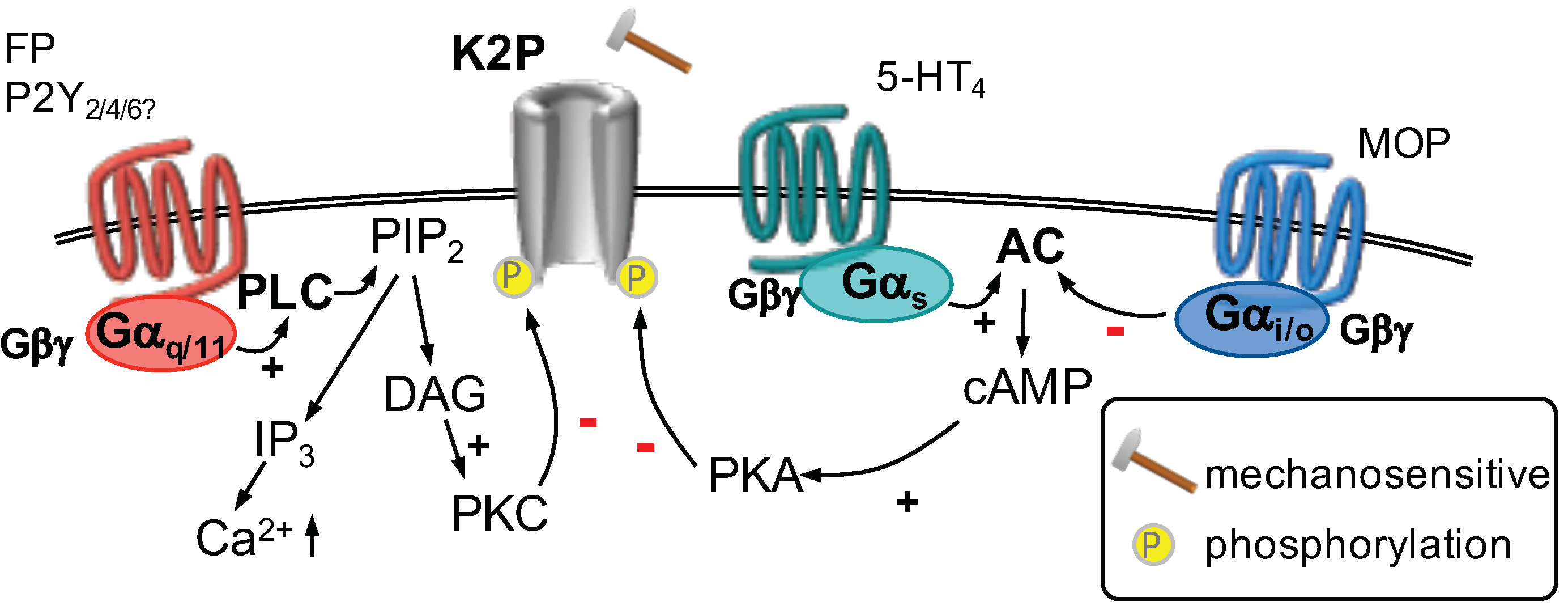
2.3.1. GPCR Regulation of Mechanosensitive Potassium Channels
2.3.2. GPCR Regulation of Piezo Channels
2.4. Calcium-Activated Chloride Channels in Pain Sensation
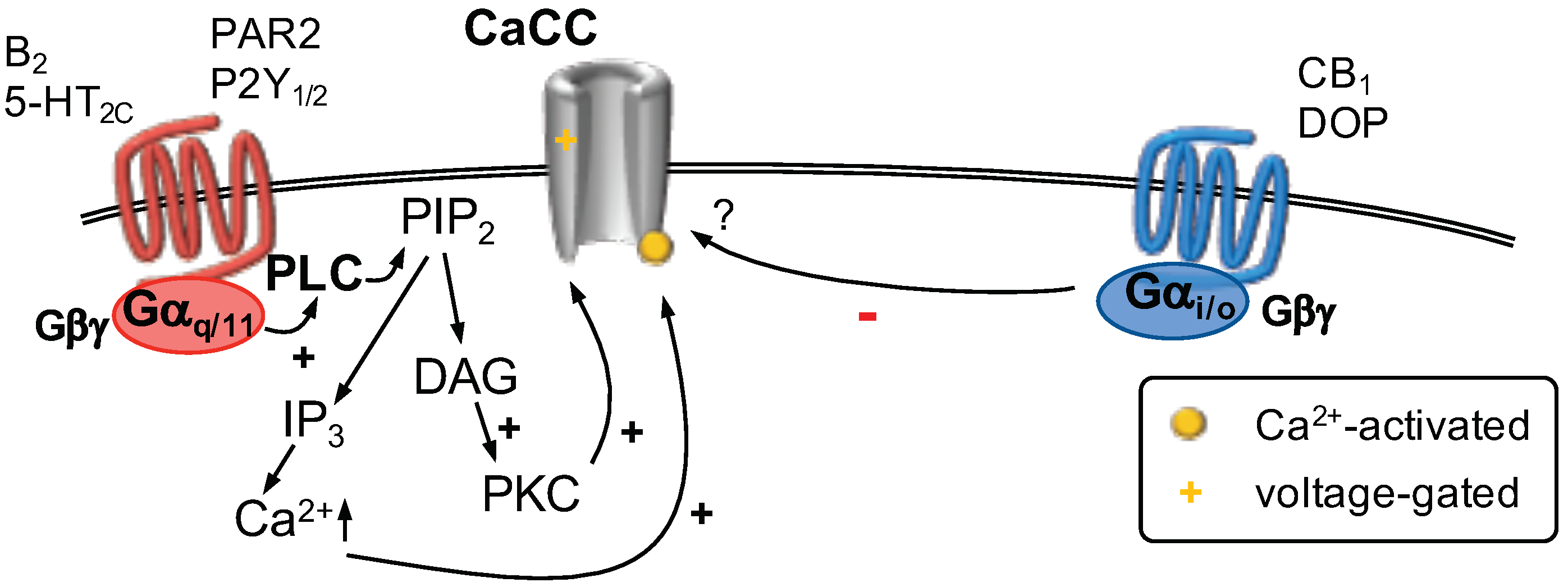
GPCR Regulation of Calcium-Activated Chloride Channels
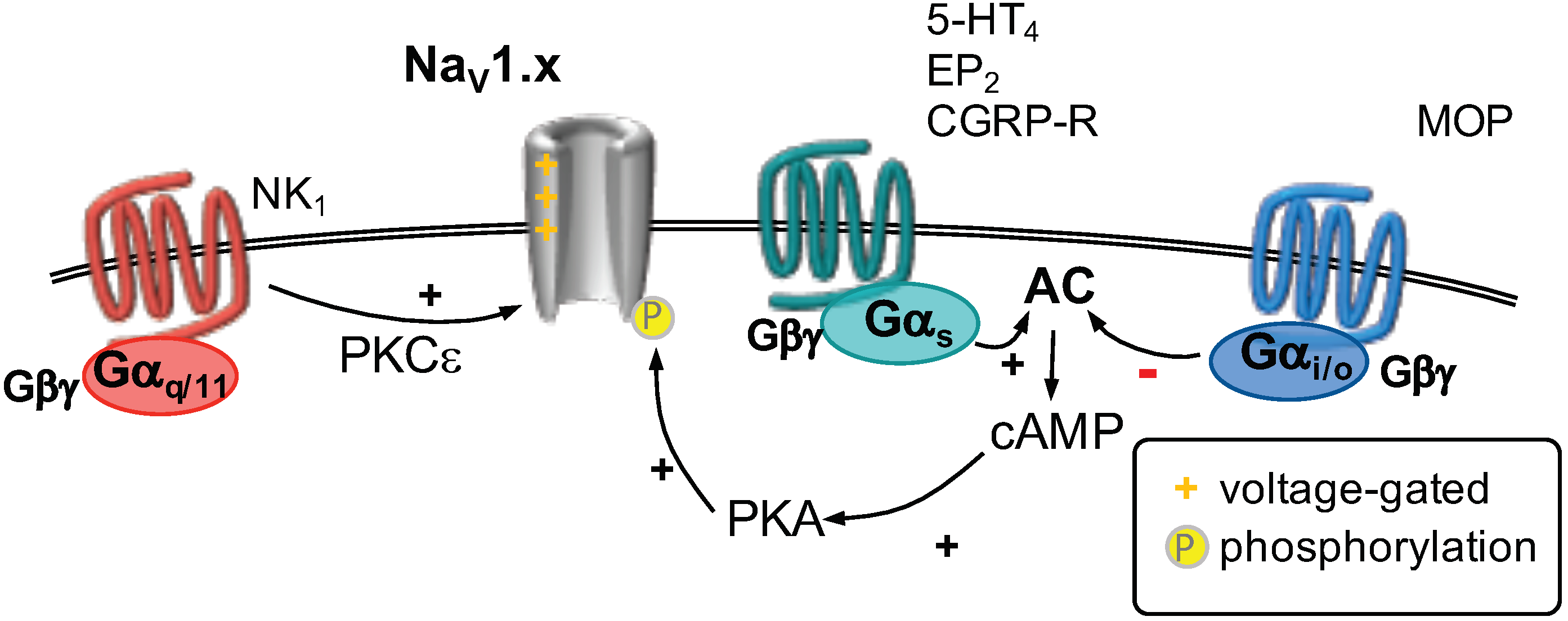
2.5. Voltage-Gated Na Channels
2.6. GPCR Regulation of Voltage-Gated Na Channels
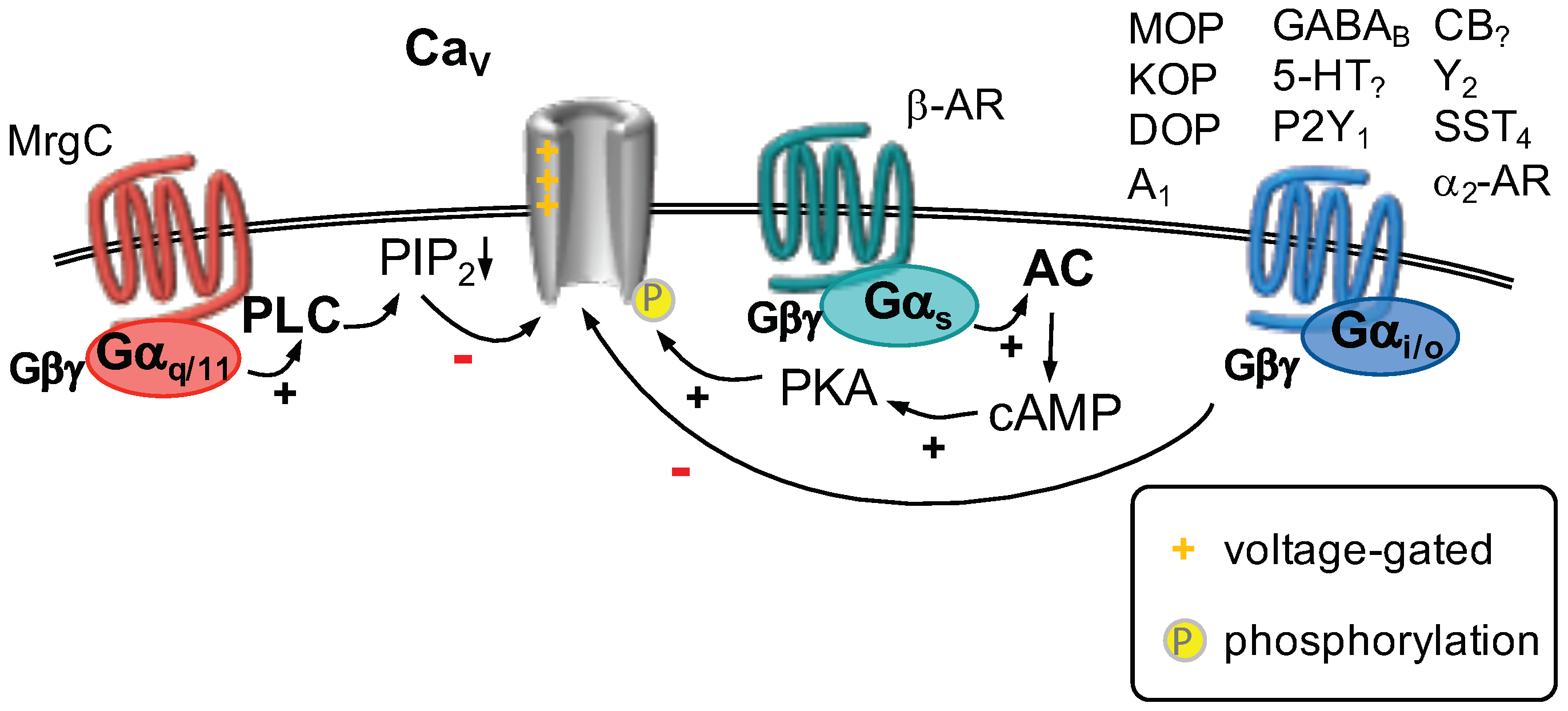
2.7. Voltage-Gated Ca Channels
GPCR Regulation of Voltage-Gated Ca Channels
2.8. Voltage-Gated K Channels
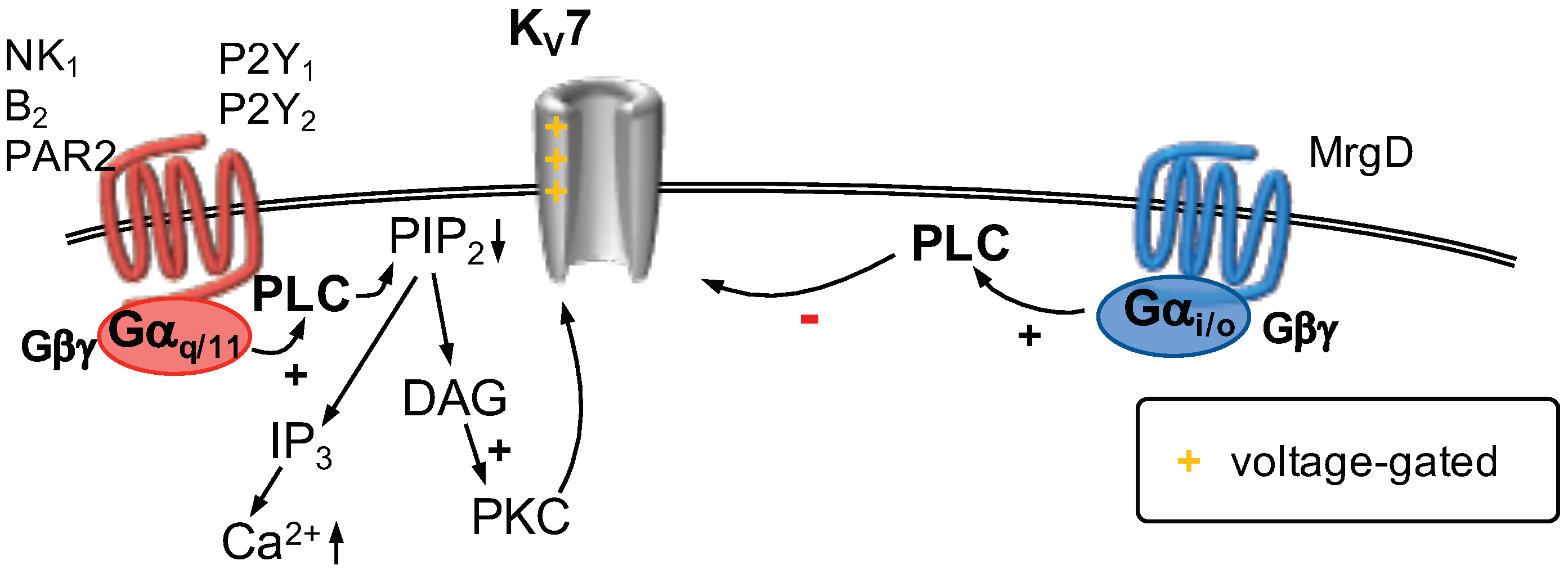
2.8.1. K7 Channels
2.8.2. GPCR Regulation of K7 Channels
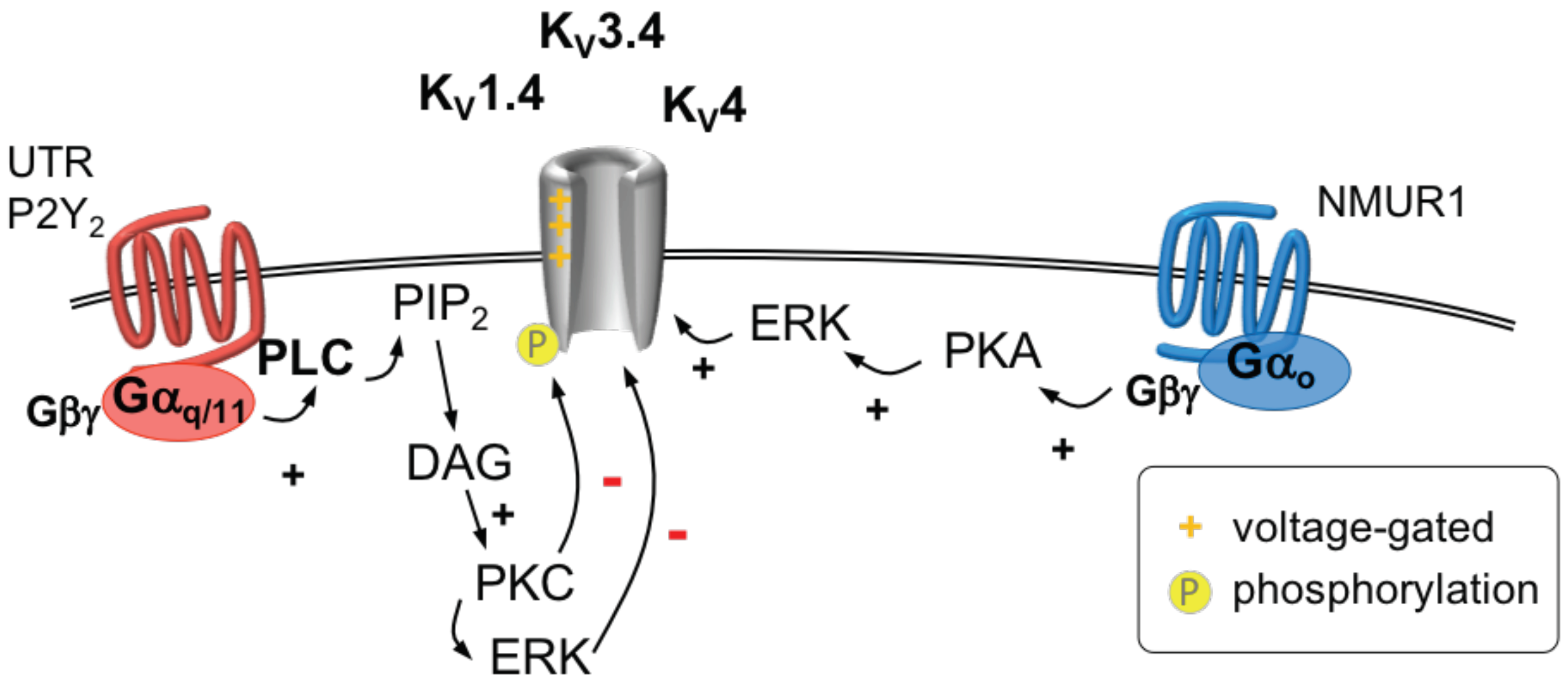
2.8.3. K1.4, K3.4 and K4 Channels
2.8.4. GPCR Regulation of K1.4, K3.4 and K4 Channels
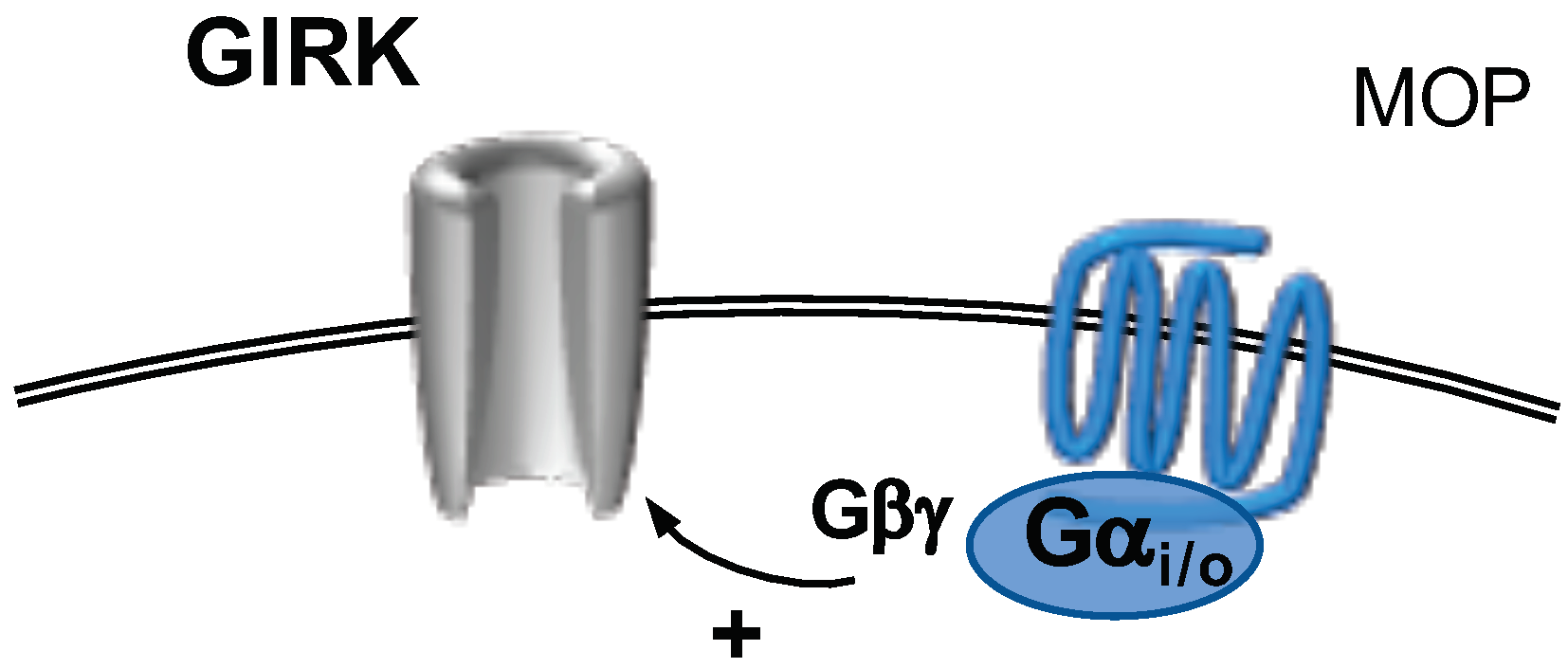
2.9. G-Protein Activated, Inwardly Rectifying Potassium Channels
GPCR Modulation of Girk Channels
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AR | adrenoceptor |
| AR | adrenoceptor |
| -OR | opioid receptor |
| 5-HT | 5-hydroxytryptamine receptor 2 or 4 |
| A | adenosine A receptor |
| AC | adenylyl cyclase |
| ADP | adenosine diphosphate |
| ANO1-10 | anoctamin 1 to 10 |
| AP | action potential |
| ASIC | acid sensing ion channel |
| ATP | adenosine triphosphate |
| B | bradykinin B receptor |
| BAPTA-AM | 1,2-Bis(2-aminophenoxy)ethane-N,N,N,N-tetraacetic acid tetrakis(acetoxymethyl ester) |
| Ca | voltage-gated Ca channel |
| CaCC | Ca-activated Cl channel |
| CamKII | Ca/calmodulin dependent protein kinase II |
| cAMP | cyclic adenosine monophosphate |
| CaN | calcineurin |
| CB | cannabinoid CB or CB receptor |
| cGMP | cyclic guanosine monophosphate |
| CGRP | calcitonin gene-related peptide |
| CGRP-R | calcitonin gene-related peptide receptor |
| CNS | central nervous system |
| Cryo-EM | cryogenic electron microscopy |
| DADLE | [D-Ala2]-enkephalin |
| DAG | diacylglycerol |
| DAMGO | [D-Ala, NMe-Phe, Gly-ol]-enkephalin |
| DI-IV | domain I to IV of Ca and Na channels |
| DOG1 | discovered on GIST 1 |
| DOP | opioid receptor |
| DRG | dorsal root ganglion |
| ECD | extracellular domain |
| EP | prostanoid EP or EP receptor |
| ER | endoplasmic reticulum |
| ERK | extracellular signal-regulated kinase |
| ET | endothelin ET receptor |
| FP | prostanoid FP receptor |
| G | G protein |
| G | G protein |
| G | G protein |
| G | G dimer |
| GABA | -aminobutyric acid receptor B |
| GIRK | G-protein activated, inwardly rectifying potassium channels |
| GIST | gastrointestinal stroma tumor |
| GPCR | G-protein coupled receptor |
| HVA | high voltage activated Ca channel |
| I | A-type K current |
| IP | prostanoid IP receptor |
| IP | inositol 1,4,5 trisphosphate |
| K | two-pore K channel |
| K | inwardly rectifying potassium channel |
| K | voltage-gated K |
| KCNJ | gene name potassium voltage-gated channel subfamily J |
| KCNQ | gene name potassium voltage-gated channel subfamily Q |
| KOP | opioid receptor |
| LVA | low voltage activated Ca channel |
| MOP | opioid receptor |
| MrgC | mas-related G protein coupled receptor type C |
| MrgD | mas-related G protein coupled receptor type D |
| mRNA | messenger ribonucleic acid |
| Na | voltage-gated Na channel |
| NGF | nerve growth factor |
| NK | tachynin NK or NK receptor |
| NKA | neurokinin A |
| NMU | neuromedin U |
| NMUR | NMU receptor |
| P2X | purinergic P2X receptor |
| P2Y | purinergic P2Y receptor |
| PAR2/ 4 | protease-activated receptor type 2 or type 4 |
| PGE | prostaglandin E |
| PGF | prostaglandin F |
| PICK-1 | protein interacting with C-kinase 1 |
| PIP | phosphatitylinositol 4,5 bisphosphate |
| PKA | protein kinase A |
| PKC | protein kinase C |
| PKG | protein kinase G |
| PLA | phospholipase A |
| PLC | phospholipase C |
| PPI | protein phosphatase I |
| PTX | pertussis toxin |
| S1–4 | transmembrane segment 1–4 of voltage-gated channels |
| SCI | spinal cord injury |
| siRNA | small interfering RNA |
| SP | substance P |
| SST | somatostatin SST receptor |
| TG | trigeminal ganglion |
| TMEM16 | transmembrane protein of unknown function family 16 |
| TMS | transmembrane segment |
| TNF | tumor necrosis factor |
| TRAAK | TWIK-related arachidonic acid activated K channel = K4.1 |
| TREK1/2 | TWIK-related K channel 1 (K2.1) or TWIK-related K channel 2 (K10.1) |
| TRH | thyrotropin-releasing hormon |
| trk | tyrosine receptor kinase |
| TRPA1 | transient receptor potential channel ankyrin family |
| TRPM8 | transient receptor potential channel melastatin family |
| TRPV | transient receptor potential channel vanilloid family |
| TTX | tetrodotoxin |
| TWIK | tandem of pore domains in a weak inward rectifying K channel |
| UTP | uridine triphosphate |
| UTR | urotensin-II receptor |
| V | vasopressin V receptor |
| Y | neuropeptide Y Y receptor |
References
- Loeser, J.D.; Treede, R.D. The Kyoto protocol of IASP Basic Pain Terminology. Pain 2008, 137, 473–477. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and molecular mechanisms of pain. Cell 2009, 139, 267–284. [Google Scholar] [CrossRef]
- Hucho, T.; Levine, J.D. Signaling pathways in sensitization: Toward a nociceptor cell biology. Neuron 2007, 55, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Venkatachalam, K.; Montell, C. TRP channels. Annu. Rev. Biochem. 2007, 76, 387–417. [Google Scholar] [CrossRef] [PubMed]
- Baron, A.; Lingueglia, E. Pharmacology of acid-sensing ion channels—Physiological and therapeutical perspectives. Neuropharmacology 2015, 94, 19–35. [Google Scholar] [CrossRef]
- North, R.A.; Jarvis, M.F. P2X receptors as drug targets. Mol. Pharmacol. 2013, 83, 759–769. [Google Scholar] [CrossRef]
- Cook, A.D.; Christensen, A.D.; Tewari, D.; McMahon, S.B.; Hamilton, J.A. Immune Cytokines and Their Receptors in Inflammatory Pain. Trends Immunol. 2018, 39, 240–255. [Google Scholar] [CrossRef]
- Denk, F.; Bennett, D.L.; McMahon, S.B. Nerve Growth Factor and Pain Mechanisms. Annu. Rev. Neurosci. 2017, 40, 307–325. [Google Scholar] [CrossRef]
- Kaneko, Y.; Szallasi, A. Transient receptor potential (TRP) channels: A clinical perspective. Br. J. Pharmacol. 2014, 171, 2474–2507. [Google Scholar] [CrossRef]
- Basso, L.; Altier, C. Transient Receptor Potential Channels in neuropathic pain. Curr. Opin. Pharmacol. 2017, 32, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Alexander, S.P.; Striessnig, J.; Kelly, E.; Marrion, N.V.; Peters, J.A.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; Southan, C.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Voltage-gated ion channels. Br. J. Pharmacol. 2017, 174, S160–S194. [Google Scholar] [CrossRef]
- Dai, Y. TRPs and pain. Semin. Immunopathol. 2016, 38, 277–291. [Google Scholar] [CrossRef]
- Mickle, A.D.; Shepherd, A.J.; Mohapatra, D.P. Sensory TRP channels: The key transducers of nociception and pain. Prog. Mol. Biol. Transl. Sci. 2015, 131, 73–118. [Google Scholar]
- Steinberg, X.; Lespay-Rebolledo, C.; Brauchi, S. A structural view of ligand-dependent activation in thermoTRP channels. Front. Physiol. 2014, 5, 171. [Google Scholar] [CrossRef]
- Wang, G.; Wang, K. The Ca2+-Permeable Cation Transient Receptor Potential TRPV3 Channel: An Emerging Pivotal Target for Itch and Skin Diseases. Mol. Pharmacol. 2017, 92, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. Targeting TRP ion channels for itch relief. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2015, 388, 389–399. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kobayashi, K.; Fukuoka, T.; Obata, K.; Yamanaka, H.; Dai, Y.; Tokunaga, A.; Noguchi, K. Distinct expression of TRPM8, TRPA1, and TRPV1 mRNAs in rat primary afferent neurons with adelta/c-fibers and colocalization with trk receptors. J. Comp. Neurol. 2005, 493, 596–606. [Google Scholar] [CrossRef] [PubMed]
- Okazawa, M.; Inoue, W.; Hori, A.; Hosokawa, H.; Matsumura, K.; Kobayashi, S. Noxious heat receptors present in cold-sensory cells in rats. Neurosci. Lett. 2004, 359, 33–36. [Google Scholar] [CrossRef]
- Usoskin, D.; Furlan, A.; Islam, S.; Abdo, H.; Lönnerberg, P.; Lou, D.; Hjerling-Leffler, J.; Haeggström, J.; Kharchenko, O.; Kharchenko, P.V.; et al. Unbiased classification of sensory neuron types by large-scale single-cell RNA sequencing. Nat. Neurosci. 2015, 18, 145–153. [Google Scholar] [CrossRef]
- Palkar, R.; Lippoldt, E.K.; McKemy, D.D. The molecular and cellular basis of thermosensation in mammals. Curr. Opin. Neurobiol. 2015, 34, 14–19. [Google Scholar] [CrossRef]
- White, J.P.M.; Cibelli, M.; Urban, L.; Nilius, B.; McGeown, J.G.; Nagy, I. TRPV4: Molecular Conductor of a Diverse Orchestra. Physiol. Rev. 2016, 96, 911–973. [Google Scholar] [CrossRef]
- Vriens, J.; Owsianik, G.; Hofmann, T.; Philipp, S.E.; Stab, J.; Chen, X.; Benoit, M.; Xue, F.; Janssens, A.; Kerselaers, S.; et al. TRPM3 is a nociceptor channel involved in the detection of noxious heat. Neuron 2011, 70, 482–494. [Google Scholar] [CrossRef]
- Laing, R.J.; Dhaka, A. ThermoTRPs and Pain. Neurosci. A Rev. J. Bringing Neurobiol. Neurol. Psychiatry 2016, 22, 171–187. [Google Scholar] [CrossRef]
- Latorre, R.; Zaelzer, C.; Brauchi, S. Structure-functional intimacies of transient receptor potential channels. Q. Rev. Biophys. 2009, 42, 201–246. [Google Scholar] [CrossRef]
- Jardín, I.; López, J.J.; Diez, R.; Sánchez-Collado, J.; Cantonero, C.; Albarrán, L.; Woodard, G.E.; Redondo, P.C.; Salido, G.M.; Smani, T.; et al. TRPs in Pain Sensation. Front. Physiol. 2017, 8, 392. [Google Scholar] [CrossRef]
- Gao, Y.; Cao, E.; Julius, D.; Cheng, Y. TRPV1 structures in nanodiscs reveal mechanisms of ligand and lipid action. Nature 2016, 534, 347–351. [Google Scholar] [CrossRef]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef]
- Liao, M.; Cao, E.; Julius, D.; Cheng, Y. Structure of the TRPV1 ion channel determined by electron cryo-microscopy. Nature 2013, 504, 107–112. [Google Scholar] [CrossRef]
- Gregorio-Teruel, L.; Valente, P.; Liu, B.; Fernández-Ballester, G.; Qin, F.; Ferrer-Montiel, A. The Integrity of the TRP Domain Is Pivotal for Correct TRPV1 Channel Gating. Biophys. J. 2015, 109, 529–541. [Google Scholar] [CrossRef]
- Rohacs, T. Phosphoinositide regulation of TRPV1 revisited. Pflugers Arch. Eur. J. Physiol. 2015, 467, 1851–1869. [Google Scholar] [CrossRef]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Park, U.; Vastani, N.; Guan, Y.; Raja, S.N.; Koltzenburg, M.; Caterina, M.J. TRP vanilloid 2 knock-out mice are susceptible to perinatal lethality but display normal thermal and mechanical nociception. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 11425–11436. [Google Scholar] [CrossRef]
- Huang, S.M.; Li, X.; Yu, Y.; Wang, J.; Caterina, M.J. TRPV3 and TRPV4 ion channels are not major contributors to mouse heat sensation. Mol. Pain 2011, 7, 37. [Google Scholar] [CrossRef]
- Vandewauw, I.; De Clercq, K.; Mulier, M.; Held, K.; Pinto, S.; Van Ranst, N.; Segal, A.; Voet, T.; Vennekens, R.; Zimmermann, K.; et al. A TRP channel trio mediates acute noxious heat sensing. Nature 2018, 555, 662–666. [Google Scholar] [CrossRef]
- Chen, J.; Kang, D.; Xu, J.; Lake, M.; Hogan, J.O.; Sun, C.; Walter, K.; Yao, B.; Kim, D. Species differences and molecular determinant of TRPA1 cold sensitivity. Nat. Commun. 2013, 4, 2501. [Google Scholar] [CrossRef]
- Moparthi, L.; Survery, S.; Kreir, M.; Simonsen, C.; Kjellbom, P.; Högestätt, E.D.; Johanson, U.; Zygmunt, P.M. Human TRPA1 is intrinsically cold- and chemosensitive with and without its N-terminal ankyrin repeat domain. Proc. Natl. Acad. Sci. USA 2014, 111, 16901–16906. [Google Scholar] [CrossRef]
- Nilius, B.; Appendino, G.; Owsianik, G. The transient receptor potential channel TRPA1: From gene to pathophysiology. Pflugers Arch. Eur. J. Physiol. 2012, 464, 425–458. [Google Scholar] [CrossRef]
- Rohacs, T. Regulation of transient receptor potential channels by the phospholipase C pathway. Adv. Biol. Regul. 2013, 53, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Julius, D. TRP channels and pain. Annu. Rev. Cell Dev. Biol. 2013, 29, 355–384. [Google Scholar] [CrossRef]
- Tang, H.B.; Inoue, A.; Oshita, K.; Nakata, Y. Sensitization of vanilloid receptor 1 induced by bradykinin via the activation of second messenger signaling cascades in rat primary afferent neurons. Eur. J. Pharmacol. 2004, 498, 37–43. [Google Scholar] [CrossRef]
- Ohta, T.; Ikemi, Y.; Murakami, M.; Imagawa, T.; Otsuguro, K.I.; Ito, S. Potentiation of transient receptor potential V1 functions by the activation of metabotropic 5-HT receptors in rat primary sensory neurons. J. Physiol. 2006, 576, 809–822. [Google Scholar] [CrossRef]
- Sugiuar, T.; Bielefeldt, K.; Gebhart, G.F. TRPV1 function in mouse colon sensory neurons is enhanced by metabotropic 5-hydroxytryptamine receptor activation. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 9521–9530. [Google Scholar] [CrossRef]
- Salzer, I.; Gantumur, E.; Yousuf, A.; Boehm, S. Control of sensory neuron excitability by serotonin involves 5HT2C receptors and Ca(2+)-activated chloride channels. Neuropharmacology 2016, 110, 277–286. [Google Scholar] [CrossRef]
- Moriyama, T.; Iida, T.; Kobayashi, K.; Higashi, T.; Fukuoka, T.; Tsumura, H.; Leon, C.; Suzuki, N.; Inoue, K.; Gachet, C.; et al. Possible involvement of P2Y2 metabotropic receptors in ATP-induced transient receptor potential vanilloid receptor 1-mediated thermal hypersensitivity. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 6058–6062. [Google Scholar] [CrossRef]
- Malin, S.A.; Davis, B.M.; Koerber, H.R.; Reynolds, I.J.; Albers, K.M.; Molliver, D.C. Thermal nociception and TRPV1 function are attenuated in mice lacking the nucleotide receptor P2Y2. Pain 2008, 138, 484–496. [Google Scholar] [CrossRef]
- Yousuf, A.; Klinger, F.; Schicker, K.; Boehm, S. Nucleotides control the excitability of sensory neurons via two P2Y receptors and a bifurcated signaling cascade. Pain 2011, 152, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Solinski, H.J.; Zierler, S.; Gudermann, T.; Breit, A. Human sensory neuron-specific Mas-related G protein-coupled receptors-X1 sensitize and directly activate transient receptor potential cation channel V1 via distinct signaling pathways. J. Biol. Chem. 2012, 287, 40956–40971. [Google Scholar] [CrossRef]
- Amadesi, S.; Nie, J.; Vergnolle, N.; Cottrell, G.S.; Grady, E.F.; Trevisani, M.; Manni, C.; Geppetti, P.; McRoberts, J.A.; Ennes, H.; et al. Protease-activated receptor 2 sensitizes the capsaicin receptor transient receptor potential vanilloid receptor 1 to induce hyperalgesia. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 4300–4312. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Moriyama, T.; Higashi, T.; Togashi, K.; Kobayashi, K.; Yamanaka, H.; Tominaga, M.; Noguchi, K. Proteinase-activated receptor 2-mediated potentiation of transient receptor potential vanilloid subfamily 1 activity reveals a mechanism for proteinase-induced inflammatory pain. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 4293–4299. [Google Scholar] [CrossRef] [PubMed]
- Vellani, V.; Kinsey, A.M.; Prandini, M.; Hechtfischer, S.C.; Reeh, P.; Magherini, P.C.; Giacomoni, C.; McNaughton, P.A. Protease activated receptors 1 and 4 sensitize TRPV1 in nociceptive neurones. Mol. Pain 2010, 6, 61. [Google Scholar] [CrossRef]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP reveals peripheral nociceptive mechanism of prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef]
- Schnizler, K.; Shutov, L.P.; Van Kanegan, M.J.; Merrill, M.A.; Nichols, B.; McKnight, G.S.; Strack, S.; Hell, J.W.; Usachev, Y.M. Protein kinase A anchoring via AKAP150 is essential for TRPV1 modulation by forskolin and prostaglandin E2 in mouse sensory neurons. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 4904–4917. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Gao, Y.; Yang, L.; Kong, X.; Yu, J.; Hou, W.; Hua, B. The mechanism of μ-opioid receptor (MOR)-TRPV1 crosstalk in TRPV1 activation involves morphine anti-nociception, tolerance and dependence. Channels 2015, 9, 235–243. [Google Scholar] [CrossRef]
- Plant, T.D.; Zöllner, C.; Mousa, S.A.; Oksche, A. Endothelin-1 potentiates capsaicin-induced TRPV1 currents via the endothelin A receptor. Exp. Biol. Med. 2006, 231, 1161–1164. [Google Scholar]
- Plant, T.D.; Zöllner, C.; Kepura, F.; Mousa, S.S.; Eichhorst, J.; Schaefer, M.; Furkert, J.; Stein, C.; Oksche, A. Endothelin potentiates TRPV1 via ETA receptor-mediated activation of protein kinase C. Mol. Pain 2007, 3, 35. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cang, C.L.; Kawasaki, Y.; Liang, L.L.; Zhang, Y.Q.; Ji, R.R.; Zhao, Z.Q. Neurokinin-1 receptor enhances TRPV1 activity in primary sensory neurons via PKCepsilon: A novel pathway for heat hyperalgesia. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 12067–12077. [Google Scholar] [CrossRef] [PubMed]
- Sculptoreanu, A.; Aura Kullmann, F.; de Groat, W.C. Neurokinin 2 receptor-mediated activation of protein kinase C modulates capsaicin responses in DRG neurons from adult rats. Eur. J. Neurosci. 2008, 27, 3171–3181. [Google Scholar] [CrossRef]
- Chen, Y.J.; Huang, C.W.; Lin, C.S.; Chang, W.H.; Sun, W.H. Expression and function of proton-sensing G-protein-coupled receptors in inflammatory pain. Mol. Pain 2009, 5, 39. [Google Scholar] [CrossRef]
- Vetter, I.; Wyse, B.D.; Monteith, G.R.; Roberts-Thomson, S.J.; Cabot, P.J. The mu opioid agonist morphine modulates potentiation of capsaicin-evoked TRPV1 responses through a cyclic AMP-dependent protein kinase A pathway. Mol. Pain 2006, 2, 22. [Google Scholar] [CrossRef]
- Endres-Becker, J.; Heppenstall, P.A.; Mousa, S.A.; Labuz, D.; Oksche, A.; Schäfer, M.; Stein, C.; Zöllner, C. Mu-opioid receptor activation modulates transient receptor potential vanilloid 1 (TRPV1) currents in sensory neurons in a model of inflammatory pain. Mol. Pharmacol. 2007, 71, 12–18. [Google Scholar] [CrossRef]
- Anand, U.; Otto, W.R.; Sanchez-Herrera, D.; Facer, P.; Yiangou, Y.; Korchev, Y.; Birch, R.; Benham, C.; Bountra, C.; Chessell, I.P.; et al. Cannabinoid receptor CB2 localisation and agonist-mediated inhibition of capsaicin responses in human sensory neurons. Pain 2008, 138, 667–680. [Google Scholar] [CrossRef]
- Sántha, P.; Jenes, A.; Somogyi, C.; Nagy, I. The endogenous cannabinoid anandamide inhibits transient receptor potential vanilloid type 1 receptor-mediated currents in rat cultured primary sensory neurons. Acta Physiol. Hung. 2010, 97, 149–158. [Google Scholar] [CrossRef]
- Bonnington, J.K.; McNaughton, P.A. Signalling pathways involved in the sensitisation of mouse nociceptive neurones by nerve growth factor. J. Physiol. 2003, 551, 433–446. [Google Scholar] [CrossRef]
- Kim, B.M.; Lee, S.H.; Shim, W.S.; Oh, U. Histamine-induced Ca(2+) influx via the PLA(2)/lipoxygenase/ TRPV1 pathway in rat sensory neurons. Neurosci. Lett. 2004, 361, 159–162. [Google Scholar] [CrossRef]
- Shim, W.S.; Tak, M.H.; Lee, M.H.; Kim, M.; Kim, M.; Koo, J.Y.; Lee, C.H.; Kim, M.; Oh, U. TRPV1 mediates histamine-induced itching via the activation of phospholipase A2 and 12-lipoxygenase. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 2331–2337. [Google Scholar] [CrossRef]
- Kajihara, Y.; Murakami, M.; Imagawa, T.; Otsuguro, K.; Ito, S.; Ohta, T. Histamine potentiates acid-induced responses mediating transient receptor potential V1 in mouse primary sensory neurons. Neuroscience 2010, 166, 292–304. [Google Scholar] [CrossRef]
- Dembla, S.; Behrendt, M.; Mohr, F.; Goecke, C.; Sondermann, J.; Schneider, F.M.; Schmidt, M.; Stab, J.; Enzeroth, R.; Leitner, M.G.; et al. Anti-nociceptive action of peripheral mu-opioid receptors by G-beta-gamma protein-mediated inhibition of TRPM3 channels. eLife 2017, 6, 1–32. [Google Scholar] [CrossRef]
- Quallo, T.; Alkhatib, O.; Gentry, C.; Andersson, D.A.; Bevan, S. G protein βγ subunits inhibit TRPM3 ion channels in sensory neurons. eLife 2017, 6, 1–22. [Google Scholar] [CrossRef]
- Badheka, D.; Yudin, Y.; Borbiro, I.; Hartle, C.M.; Yazici, A.; Mirshahi, T.; Rohacs, T. Inhibition of Transient Receptor Potential Melastatin 3 ion channels by G-protein βγ subunits. eLife 2017, 6, 1–21. [Google Scholar] [CrossRef]
- Wang, S.; Dai, Y.; Fukuoka, T.; Yamanaka, H.; Kobayashi, K.; Obata, K.; Cui, X.; Tominaga, M.; Noguchi, K. Phospholipase C and protein kinase A mediate bradykinin sensitization of TRPA1: A molecular mechanism of inflammatory pain. Brain A J. Neurol. 2008, 131, 1241–1251. [Google Scholar] [CrossRef]
- Meotti, F.C.; Figueiredo, C.P.; Manjavachi, M.; Calixto, J.B. The transient receptor potential ankyrin-1 mediates mechanical hyperalgesia induced by the activation of B1 receptor in mice. Biochem. Pharmacol. 2017, 125, 75–83. [Google Scholar] [CrossRef]
- Andrade, E.L.; Luiz, A.P.; Ferreira, J.; Calixto, J.B. Pronociceptive response elicited by TRPA1 receptor activation in mice. Neuroscience 2008, 152, 511–520. [Google Scholar] [CrossRef]
- Brozmanova, M.; Mazurova, L.; Ru, F.; Tatar, M.; Hu, Y.; Yu, S.; Kollarik, M. Mechanisms of the adenosine A2A receptor-induced sensitization of esophageal C fibers. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G215–223. [Google Scholar] [CrossRef]
- Taylor-Clark, T.E.; Undem, B.J.; Macglashan, D.W.; Ghatta, S.; Carr, M.J.; McAlexander, M.A. Prostaglandin-induced activation of nociceptive neurons via direct interaction with transient receptor potential A1 (TRPA1). Mol. Pharmacol. 2008, 73, 274–281. [Google Scholar] [CrossRef]
- Weng, Y.; Batista-Schepman, P.A.; Barabas, M.E.; Harris, E.Q.; Dinsmore, T.B.; Kossyreva, E.A.; Foshage, A.M.; Wang, M.H.; Schwab, M.J.; Wang, V.M.; et al. Prostaglandin metabolite induces inhibition of TRPA1 and channel-dependent nociception. Mol. Pain 2012, 8, 75. [Google Scholar] [CrossRef]
- Linte, R.M.; Ciobanu, C.; Reid, G.; Babes, A. Desensitization of cold- and menthol-sensitive rat dorsal root ganglion neurones by inflammatory mediators. Exp. Brain Res. 2007, 178, 89–98. [Google Scholar] [CrossRef]
- Premkumar, L.S.; Raisinghani, M.; Pingle, S.C.; Long, C.; Pimentel, F. Downregulation of transient receptor potential melastatin 8 by protein kinase C-mediated dephosphorylation. J. Neurosci. Off. J. Soc. Neurosci. 2005, 25, 11322–11329. [Google Scholar] [CrossRef]
- Babes, A.; Ciobanu, A.C.; Neacsu, C.; Babes, R.M. TRPM8, a sensor for mild cooling in mammalian sensory nerve endings. Curr. Pharm. Biotechnol. 2011, 12, 78–88. [Google Scholar] [CrossRef]
- Yudin, Y.; Lukacs, V.; Cao, C.; Rohacs, T. Decrease in phosphatidylinositol 4,5-bisphosphate levels mediates desensitization of the cold sensor TRPM8 channels. J. Physiol. 2011, 589, 6007–6027. [Google Scholar] [CrossRef]
- Zhang, X.; Mak, S.; Li, L.; Parra, A.; Denlinger, B.; Belmonte, C.; McNaughton, P.A. Direct inhibition of the cold-activated TRPM8 ion channel by Gαq. Nat. Cell Biol. 2012, 14, 851–858. [Google Scholar] [CrossRef]
- Abdelhamid, R.E.; Sluka, K.A. ASICs Mediate Pain and Inflammation in Musculoskeletal Diseases. Physiology 2015, 30, 449–459. [Google Scholar] [CrossRef]
- Hwang, S.W.; Oh, U. Current concepts of nociception: Nociceptive molecular sensors in sensory neurons. Curr. Opin. Anaesthesiol. 2007, 20, 427–434. [Google Scholar] [CrossRef]
- Alexander, S.P.; Peters, J.A.; Kelly, E.; Marrion, N.V.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; Southan, C.; Davies, J.A.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: Ligand-gated ion channels. Br. J. Pharmacol. 2017, 174, S130–S159. [Google Scholar] [CrossRef]
- Deval, E.; Lingueglia, E. Acid-Sensing Ion Channels and nociception in the peripheral and central nervous systems. Neuropharmacology 2015, 94, 49–57. [Google Scholar] [CrossRef]
- Ugawa, S.; Ueda, T.; Yamamura, H.; Shimada, S. In situ hybridization evidence for the coexistence of ASIC and TRPV1 within rat single sensory neurons. Brain Research. Mol. Brain Res. 2005, 136, 125–133. [Google Scholar] [CrossRef]
- Wemmie, J.A.; Taugher, R.J.; Kreple, C.J. Acid-sensing ion channels in pain and disease. Nat. Rev. Neurosci. 2013, 14, 461–471. [Google Scholar] [CrossRef]
- Yoder, N.; Gouaux, E. Divalent cation and chloride ion sites of chicken acid sensing ion channel 1a elucidated by X-ray crystallography. PLoS ONE 2018, 13, e0202134. [Google Scholar]
- Rash, L.D. Acid-Sensing Ion Channel Pharmacology, Past, Present, and Future …. Adv. Pharmacol. 2017, 79, 35–66. [Google Scholar]
- Krauson, A.J.; Carattino, M.D. The Thumb Domain Mediates Acid-sensing Ion Channel Desensitization. J. Biol. Chem. 2016, 291, 11407–11419. [Google Scholar] [CrossRef]
- Cheng, Y.R.; Jiang, B.Y.; Chen, C.C. Acid-sensing ion channels: Dual function proteins for chemo-sensing and mechano-sensing. J. Biomed. Sci. 2018, 25, 46. [Google Scholar] [CrossRef] [PubMed]
- Nagaeva, E.I.; Tikhonova, T.B.; Magazanik, L.G.; Tikhonov, D.B. Histamine selectively potentiates acid-sensing ion channel 1a. Neurosci. Lett. 2016, 632, 136–140. [Google Scholar] [CrossRef]
- Ren, C.; Gan, X.; Wu, J.; Qiu, C.Y.; Hu, W.P. Enhancement of acid-sensing ion channel activity by metabotropic P2Y UTP receptors in primary sensory neurons. Purinergic Signal. 2016, 12, 69–78. [Google Scholar] [CrossRef]
- Qiu, F.; Qiu, C.Y.; Liu, Y.Q.; Wu, D.; Li, J.D.; Hu, W.P. Potentiation of acid-sensing ion channel activity by the activation of 5-HT2 receptors in rat dorsal root ganglion neurons. Neuropharmacology 2012, 63, 494–500. [Google Scholar] [CrossRef]
- Deval, E.; Salinas, M.; Baron, A.; Lingueglia, E.; Lazdunski, M. ASIC2b-dependent regulation of ASIC3, an essential acid-sensing ion channel subunit in sensory neurons via the partner protein PICK-1. J. Biol. Chem. 2004, 279, 19531–19539. [Google Scholar] [CrossRef]
- Gu, Q.; Lee, L.Y. Effect of protease-activated receptor 2 activation on single TRPV1 channel activities in rat vagal pulmonary sensory neurons. Exp. Physiol. 2009, 94, 928–936. [Google Scholar] [CrossRef]
- Zhao, P.; Metcalf, M.; Bunnett, N.W. Biased signaling of protease-activated receptors. Front. Endocrinol. 2014, 5, 67. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Qiu, F.; Qiu, C.Y.; Cai, Q.; Zou, P.; Wu, H.; Hu, W.P. Cannabinoids inhibit acid-sensing ion channel currents in rat dorsal root ganglion neurons. PLoS ONE 2012, 7, e45531. [Google Scholar] [CrossRef]
- Cai, Q.; Qiu, C.Y.; Qiu, F.; Liu, T.T.; Qu, Z.W.; Liu, Y.M.; Hu, W.P. Morphine inhibits acid-sensing ion channel currents in rat dorsal root ganglion neurons. Brain Res. 2014, 1554, 12–20. [Google Scholar] [CrossRef]
- Kubo, A.; Shinoda, M.; Katagiri, A.; Takeda, M.; Suzuki, T.; Asaka, J.; Yeomans, D.C.; Iwata, K. Oxytocin alleviates orofacial mechanical hypersensitivity associated with infraorbital nerve injury through vasopressin-1A receptors of the rat trigeminal ganglia. Pain 2017, 158, 649–659. [Google Scholar] [CrossRef]
- Qiu, F.; Qiu, C.Y.; Cai, H.; Liu, T.T.; Qu, Z.W.; Yang, Z.; Li, J.D.; Zhou, Q.Y.; Hu, W.P. Oxytocin inhibits the activity of acid-sensing ion channels through the vasopressin, V1A receptor in primary sensory neurons. Br. J. Pharmacol. 2014, 171, 3065–3076. [Google Scholar] [CrossRef] [PubMed]
- Delmas, P.; Hao, J.; Rodat-Despoix, L. Molecular mechanisms of mechanotransduction in mammalian sensory neurons. Nat. Rev. Neurosci. 2011, 12, 139–153. [Google Scholar] [CrossRef]
- Ranade, S.S.; Syeda, R.; Patapoutian, A. Mechanically Activated Ion Channels. Neuron 2015, 87, 1162–1179. [Google Scholar] [CrossRef] [PubMed]
- Brohawn, S.G. How ion channels sense mechanical force: Insights from mechanosensitive K2P channels TRAAK, TREK1, and TREK2. Ann. N. Y. Acad. Sci. 2015, 1352, 20–32. [Google Scholar] [CrossRef]
- Enyedi, P.; Czirják, G. Molecular background of leak K+ currents: Two-pore domain potassium channels. Physiol. Rev. 2010, 90, 559–605. [Google Scholar] [CrossRef] [PubMed]
- Viatchenko-Karpinski, V.; Ling, J.; Gu, J.G. Characterization of temperature-sensitive leak K+ currents and expression of TRAAK, TREK-1, and TREK2 channels in dorsal root ganglion neurons of rats. Mol. Brain 2018, 11, 40. [Google Scholar] [CrossRef]
- Li, X.Y.; Toyoda, H. Role of leak potassium channels in pain signaling. Brain Res. Bull. 2015, 119, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, Y.; Geng, J.; Zhou, S.; Xiao, B. Mechanically Activated Piezo Channels Mediate Touch and Suppress Acute Mechanical Pain Response in Mice. Cell Rep. 2019, 26, 1419–1431.e4. [Google Scholar] [CrossRef] [PubMed]
- Alexander, S.P.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Marrion, N.V.; Peters, J.A.; Faccenda, E.; Harding, S.D.; Pawson, A.J.; Sharman, J.L.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2017/18: G protein-coupled receptors. Br. J. Pharmacol. 2017, 174, S17–S129. [Google Scholar] [CrossRef] [PubMed]
- Pereira, V.; Busserolles, J.; Christin, M.; Devilliers, M.; Poupon, L.; Legha, W.; Alloui, A.; Aissouni, Y.; Bourinet, E.; Lesage, F.; et al. Role of the TREK2 potassium channel in cold and warm thermosensation and in pain perception. Pain 2014, 155, 2534–2544. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.J.; Honoré, E.; Maingret, F.; Lesage, F.; Fink, M.; Duprat, F.; Lazdunski, M. A mammalian two pore domain mechano-gated S-like K+ channel. EMBO J. 1998, 17, 4283–4290. [Google Scholar] [CrossRef] [PubMed]
- Srisomboon, Y.; Zaidman, N.A.; Maniak, P.J.; Deachapunya, C.; O’Grady, S.M. P2Y receptor regulation of K2P channels that facilitate K+ secretion by human mammary epithelial cells. Am. J. Physiol. Cell Physiol. 2018, 314, C627–C639. [Google Scholar] [CrossRef]
- Woo, D.H.; Bae, J.Y.; Nam, M.H.; An, H.; Ju, Y.H.; Won, J.; Choi, J.H.; Hwang, E.M.; Han, K.S.; Bae, Y.C.; et al. Activation of Astrocytic μ-opioid Receptor Elicits Fast Glutamate Release Through TREK-1-Containing K2P Channel in Hippocampal Astrocytes. Front. Cell. Neurosci. 2018, 12, 319. [Google Scholar] [CrossRef]
- Devilliers, M.; Busserolles, J.; Lolignier, S.; Deval, E.; Pereira, V.; Alloui, A.; Christin, M.; Mazet, B.; Delmas, P.; Noel, J.; et al. Activation of TREK-1 by morphine results in analgesia without adverse side effects. Nat. Commun. 2013, 4, 2941. [Google Scholar] [CrossRef]
- Cho, P.S.; Lee, H.K.; Lee, S.H.; Im, J.Z.; Jung, S.J. DAMGO modulates two-pore domain K(+) channels in the substantia gelatinosa neurons of rat spinal cord. Korean J. Physiol. Pharmacol. Off. J. Korean Physiol. Soc. Korean Soc. Pharmacol. 2016, 20, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Nockemann, D.; Rouault, M.; Labuz, D.; Hublitz, P.; McKnelly, K.; Reis, F.C.; Stein, C.; Heppenstall, P.A. The K(+) channel GIRK2 is both necessary and sufficient for peripheral opioid-mediated analgesia. EMBO Mol. Med. 2013, 5, 1263–1277. [Google Scholar] [CrossRef]
- Dubin, A.E.; Schmidt, M.; Mathur, J.; Petrus, M.J.; Xiao, B.; Coste, B.; Patapoutian, A. Inflammatory signals enhance piezo2-mediated mechanosensitive currents. Cell Rep. 2012, 2, 511–517. [Google Scholar] [CrossRef]
- Lechner, S.G.; Lewin, G.R. Peripheral sensitisation of nociceptors via G-protein-dependent potentiation of mechanotransduction currents. J. Physiol. 2009, 587, 3493–3503. [Google Scholar] [CrossRef]
- Jia, Z.; Ikeda, R.; Ling, J.; Gu, J.G. GTP-dependent run-up of Piezo2-type mechanically activated currents in rat dorsal root ganglion neurons. Mol. Brain 2013, 6, 57. [Google Scholar] [CrossRef]
- Borbiro, I.; Badheka, D.; Rohacs, T. Activation of TRPV1 channels inhibits mechanosensitive Piezo channel activity by depleting membrane phosphoinositides. Sci. Signal. 2015, 8, ra15. [Google Scholar] [CrossRef]
- Oh, U.; Jung, J. Cellular functions of TMEM16/anoctamin. Pflugers Arch. Eur. J. Physiol. 2016, 468, 443–453. [Google Scholar] [CrossRef]
- Schroeder, B.C.; Cheng, T.; Jan, Y.N.; Jan, L.Y. Expression cloning of TMEM16A as a calcium-activated chloride channel subunit. Cell 2008, 134, 1019–1029. [Google Scholar] [CrossRef] [PubMed]
- Caputo, A.; Caci, E.; Ferrera, L.; Pedemonte, N.; Barsanti, C.; Sondo, E.; Pfeffer, U.; Ravazzolo, R.; Zegarra-Moran, O.; Galietta, L.J.V. TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity. Science 2008, 322, 590–594. [Google Scholar] [CrossRef]
- Yang, Y.D.; Cho, H.; Koo, J.Y.; Tak, M.H.; Cho, Y.; Shim, W.S.; Park, S.P.; Lee, J.; Lee, B.; Kim, B.M.; et al. TMEM16A confers receptor-activated calcium-dependent chloride conductance. Nature 2008, 455, 1210–1215. [Google Scholar] [CrossRef] [PubMed]
- Falzone, M.E.; Malvezzi, M.; Lee, B.C.; Accardi, A. Known structures and unknown mechanisms of TMEM16 scramblases and channels. J. Gen. Physiol. 2018, 150, 933–947. [Google Scholar] [CrossRef]
- Huang, F.; Wong, X.; Jan, L.Y. International Union of Basic and Clinical Pharmacology. LXXXV: calcium-activated chloride channels. Pharmacol. Rev. 2012, 64, 1–15. [Google Scholar] [CrossRef]
- Suzuki, J.; Fujii, T.; Imao, T.; Ishihara, K.; Kuba, H.; Nagata, S. Calcium-dependent phospholipid scramblase activity of TMEM16 protein family members. J. Biol. Chem. 2013, 288, 13305–13316. [Google Scholar] [CrossRef] [PubMed]
- Scudieri, P.; Caci, E.; Venturini, A.; Sondo, E.; Pianigiani, G.; Marchetti, C.; Ravazzolo, R.; Pagani, F.; Galietta, L.J.V. Ion channel and lipid scramblase activity associated with expression of TMEM16F/ANO6 isoforms. J. Physiol. 2015, 593, 3829–3848. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Li, L.I.; Ma, K.T.; Wang, Y.; Li, J.; Shi, W.Y.; Zhu, H.E.; Zhang, Z.S.; Si, J.Q. NSAIDs modulate GABA-activated currents via Ca2+-activated Cl- channels in rat dorsal root ganglion neurons. Exp. Ther. Med. 2016, 11, 1755–1761. [Google Scholar] [CrossRef]
- Huang, F.; Wang, X.; Ostertag, E.M.; Nuwal, T.; Huang, B.; Jan, Y.N.; Basbaum, A.I.; Jan, L.Y. TMEM16C facilitates Na(+)-activated K+ currents in rat sensory neurons and regulates pain processing. Nat. Neurosci. 2013, 16, 1284–1290. [Google Scholar] [CrossRef]
- Cho, H.; Yang, Y.D.; Lee, J.; Lee, B.; Kim, T.; Jang, Y.; Back, S.K.; Na, H.S.; Harfe, B.D.; Wang, F.; et al. The calcium-activated chloride channel anoctamin 1 acts as a heat sensor in nociceptive neurons. Nat. Neurosci. 2012, 15, 1015–1021. [Google Scholar] [CrossRef]
- Lee, B.; Cho, H.; Jung, J.; Yang, Y.D.; Yang, D.J.; Oh, U. Anoctamin 1 contributes to inflammatory and nerve-injury induced hypersensitivity. Mol. Pain 2014, 10, 5. [Google Scholar] [CrossRef]
- Dang, S.; Feng, S.; Tien, J.; Peters, C.J.; Bulkley, D.; Lolicato, M.; Zhao, J.; Zuberbühler, K.; Ye, W.; Qi, L.; et al. Cryo-EM structures of the TMEM16A calcium-activated chloride channel. Nature 2017, 552, 426–429. [Google Scholar] [CrossRef]
- Paulino, C.; Kalienkova, V.; Lam, A.K.M.; Neldner, Y.; Dutzler, R. Activation mechanism of the calcium-activated chloride channel TMEM16A revealed by cryo-EM. Nature 2017, 552, 421–425. [Google Scholar] [CrossRef]
- Paulino, C.; Neldner, Y.; Lam, A.K.; Kalienkova, V.; Brunner, J.D.; Schenck, S.; Dutzler, R. Structural basis for anion conduction in the calcium-activated chloride channel TMEM16A. eLife 2017, 6, 1–23. [Google Scholar] [CrossRef]
- Brunner, J.D.; Lim, N.K.; Schenck, S.; Duerst, A.; Dutzler, R. X-ray structure of a calcium-activated TMEM16 lipid scramblase. Nature 2014, 516, 207–212. [Google Scholar] [CrossRef]
- Fallah, G.; Römer, T.; Detro-Dassen, S.; Braam, U.; Markwardt, F.; Schmalzing, G. TMEM16A(a)/anoctamin-1 shares a homodimeric architecture with CLC chloride channels. Mol. Cell. Proteom. MCP 2011, 10, M110.004697. [Google Scholar] [CrossRef]
- Sheridan, J.T.; Worthington, E.N.; Yu, K.; Gabriel, S.E.; Hartzell, H.C.; Tarran, R. Characterization of the oligomeric structure of the Ca(2+)-activated Cl- channel Ano1/TMEM16A. J. Biol. Chem. 2011, 286, 1381–1388. [Google Scholar] [CrossRef]
- Xiao, Q.; Yu, K.; Perez-Cornejo, P.; Cui, Y.; Arreola, J.; Hartzell, H.C. Voltage- and calcium-dependent gating of TMEM16A/Ano1 chloride channels are physically coupled by the first intracellular loop. Proc. Natl. Acad. Sci. USA 2011, 108, 8891–8896. [Google Scholar] [CrossRef]
- Ma, K.; Wang, H.; Yu, J.; Wei, M.; Xiao, Q. New Insights on the Regulation of Ca2+ -Activated Chloride Channel TMEM16A. J. Cell. Physiol. 2017, 232, 707–716. [Google Scholar] [CrossRef]
- Jin, X.; Shah, S.; Liu, Y.; Zhang, H.; Lees, M.; Fu, Z.; Lippiat, J.D.; Beech, D.J.; Sivaprasadarao, A.; Baldwin, S.A.; et al. Activation of the Cl- channel ANO1 by localized calcium signals in nociceptive sensory neurons requires coupling with the IP3 receptor. Sci. Signal. 2013, 6, ra73. [Google Scholar] [CrossRef]
- Takayama, Y.; Uta, D.; Furue, H.; Tominaga, M. Pain-enhancing mechanism through interaction between TRPV1 and anoctamin 1 in sensory neurons. Proc. Natl. Acad. Sci. USA 2015, 112, 5213–5218. [Google Scholar] [CrossRef]
- Cho, H.; Oh, U. Anoctamin 1 mediates thermal pain as a heat sensor. Curr. Neuropharmacol. 2013, 11, 641–651. [Google Scholar] [CrossRef]
- Liu, B.; Linley, J.E.; Du, X.; Zhang, X.; Ooi, L.; Zhang, H.; Gamper, N. The acute nociceptive signals induced by bradykinin in rat sensory neurons are mediated by inhibition of M-type K+ channels and activation of Ca2+-activated Cl- channels. J. Clin. Investig. 2010, 120, 1240–1252. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, H.; Ke, J.; Wei, Y.; Ji, H.; Qian, Z.; Liu, L.; Tao, J. Inhibition of A-Type K+ Channels by Urotensin-II Induces Sensory Neuronal Hyperexcitability Through the PKCα-ERK Pathway. Endocrinology 2018, 159, 2253–2263. [Google Scholar] [CrossRef]
- Rajagopal, M.; Kathpalia, P.P.; Thomas, S.V.; Pao, A.C. Activation of P2Y1 and P2Y2 receptors induces chloride secretion via calcium-activated chloride channels in kidney inner medullary collecting duct cells. Am. J. Physiology. Ren. Physiol. 2011, 301, F544–553. [Google Scholar] [CrossRef]
- Wang, J.; Haanes, K.A.; Novak, I. Purinergic regulation of CFTR and Ca(2+)-activated Cl(-) channels and K(+) channels in human pancreatic duct epithelium. Am. J. Physiology. Cell Physiol. 2013, 304, C673–684. [Google Scholar] [CrossRef]
- Romero, T.R.L.; Pacheco, D.D.F.; Duarte, I.D.G. Probable involvement of Ca(2+)-activated Cl(-) channels (CaCCs) in the activation of CB1 cannabinoid receptors. Life Sci. 2013, 92, 815–820. [Google Scholar] [CrossRef]
- Pacheco, D.d.F.; Pacheco, C.M.D.F.; Duarte, I.D.G. δ-Opioid receptor agonist SNC80 induces central antinociception mediated by Ca2+ -activated Cl- channels. J. Pharm. Pharmacol. 2012, 64, 1084–1089. [Google Scholar] [CrossRef]
- Cardoso, F.C.; Lewis, R.J. Sodium channels and pain: From toxins to therapies. Br. J. Pharmacol. 2018, 175, 2138–2157. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. The components of membrane conductance in the giant axon of Loligo. J. Physiol. 1952, 116, 473–496. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F. The dual effect of membrane potential on sodium conductance in the giant axon of Loligo. J. Physiol. 1952, 116, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F. Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. J. Physiol. 1952, 116, 449–472. [Google Scholar] [CrossRef] [PubMed]
- Hodgkin, A.L.; Huxley, A.F.; Katz, B. Measurement of current-voltage relations in the membrane of the giant axon of Loligo. J. Physiol. 1952, 116, 424–448. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, B.A.; Merkies, I.S.J.; Gerrits, M.M.; Waxman, S.G.; Hoeijmakers, J.G.J.; Faber, C.G. Painful neuropathies: The emerging role of sodium channelopathies. J. Peripher. Nerv. Syst. JPNS 2014, 19, 53–65. [Google Scholar] [CrossRef]
- Bezanilla, F. How membrane proteins sense voltage. Nat. Rev. Mol. Cell Biol. 2008, 9, 323–332. [Google Scholar] [CrossRef]
- Cardenas, L.M.; Cardenas, C.G.; Scroggs, R.S. 5HT increases excitability of nociceptor-like rat dorsal root ganglion neurons via cAMP-coupled TTX-resistant Na(+) channels. J. Neurophysiol. 2001, 86, 241–248. [Google Scholar] [CrossRef]
- Gold, M.S.; Reichling, D.B.; Shuster, M.J.; Levine, J.D. Hyperalgesic agents increase a tetrodotoxin-resistant Na+ current in nociceptors. Proc. Natl. Acad. Sci. USA 1996, 93, 1108–1112. [Google Scholar] [CrossRef]
- England, S.; Bevan, S.; Docherty, R.J. PGE2 modulates the tetrodotoxin-resistant sodium current in neonatal rat dorsal root ganglion neurones via the cyclic AMP-protein kinase A cascade. J. Physiol. 1996, 495, 429–440. [Google Scholar] [CrossRef]
- Natura, G.; von Banchet, G.S.; Schaible, H.G. Calcitonin gene-related peptide enhances TTX-resistant sodium currents in cultured dorsal root ganglion neurons from adult rats. Pain 2005, 116, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Scroggs, R.S. Serotonin upregulates low- and high-threshold tetrodotoxin-resistant sodium channels in the same subpopulation of rat nociceptors. Neuroscience 2010, 165, 1293–1300. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.G.; Del Mar, L.P.; Scroggs, R.S. Two parallel signaling pathways couple 5HT1A receptors to N- and L-type calcium channels in C-like rat dorsal root ganglion cells. J. Neurophysiol. 1997, 77, 3284–3296. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Ikeda, M.; Yoshida, S.; Tanimoto, T.; Takeda, M.; Nasu, M. Prostaglandin E2-induced modification of tetrodotoxin-resistant Na+ currents involves activation of both EP2 and EP4 receptors in neonatal rat nodose ganglion neurones. Br. J. Pharmacol. 2005, 145, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Moraes, E.R.; Kushmerick, C.; Naves, L.A. Characteristics of dorsal root ganglia neurons sensitive to Substance P. Mol. Pain 2014, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Cang, C.L.; Zhang, H.; Zhang, Y.Q.; Zhao, Z.Q. PKCepsilon-dependent potentiation of TTX-resistant Nav1.8 current by neurokinin-1 receptor activation in rat dorsal root ganglion neurons. Mol. Pain 2009, 5, 33. [Google Scholar] [CrossRef]
- Song, J.H.; Shin, Y.K.; Lee, C.S.; Bang, H.; Park, M. Effects of ATP on TTX-sensitive and TTX-resistant sodium currents in rat sensory neurons. Neuroreport 2001, 12, 3659–3662. [Google Scholar] [CrossRef]
- Kayssi, A.; Amadesi, S.; Bautista, F.; Bunnett, N.W.; Vanner, S. Mechanisms of protease-activated receptor 2-evoked hyperexcitability of nociceptive neurons innervating the mouse colon. J. Physiol. 2007, 580, 977–991. [Google Scholar] [CrossRef]
- Gold, M.S.; Levine, J.D. DAMGO inhibits prostaglandin E2-induced potentiation of a TTX-resistant Na+ current in rat sensory neurons in vitro. Neurosci. Lett. 1996, 212, 83–86. [Google Scholar] [CrossRef]
- Catterall, W.A. Voltage-gated calcium channels. Cold Spring Harb. Perspect. Biol. 2011, 3, a003947. [Google Scholar] [CrossRef]
- Acosta, C.G.; López, H.S. delta opioid receptor modulation of several voltage-dependent Ca(2+) currents in rat sensory neurons. J. Neurosci. Off. J. Soc. Neurosci. 1999, 19, 8337–8348. [Google Scholar] [CrossRef]
- Bean, B.P. Neurotransmitter inhibition of neuronal calcium currents by changes in channel voltage dependence. Nature 1989, 340, 153–156. [Google Scholar] [CrossRef]
- Dolphin, A.C. Beta subunits of voltage-gated calcium channels. J. Bioenerg. Biomembr. 2003, 35, 599–620. [Google Scholar] [CrossRef]
- Proft, J.; Weiss, N. G protein regulation of neuronal calcium channels: Back to the future. Mol. Pharmacol. 2015, 87, 890–906. [Google Scholar] [CrossRef]
- Scherrer, G.; Imamachi, N.; Cao, Y.Q.; Contet, C.; Mennicken, F.; O’Donnell, D.; Kieffer, B.L.; Basbaum, A.I. Dissociation of the opioid receptor mechanisms that control mechanical and heat pain. Cell 2009, 137, 1148–1159. [Google Scholar] [CrossRef]
- Snyder, L.M.; Chiang, M.C.; Loeza-Alcocer, E.; Omori, Y.; Hachisuka, J.; Sheahan, T.D.; Gale, J.R.; Adelman, P.C.; Sypek, E.I.; Fulton, S.A.; et al. Kappa Opioid Receptor Distribution and Function in Primary Afferents. Neuron 2018, 99, 1274–1288.e6. [Google Scholar] [CrossRef]
- Mudge, A.W.; Leeman, S.E.; Fischbach, G.D. Enkephalin inhibits release of substance P from sensory neurons in culture and decreases action potential duration. Proc. Natl. Acad. Sci. USA 1979, 76, 526–530. [Google Scholar] [CrossRef]
- Werz, M.A.; Grega, D.S.; MacDonald, R.L. Actions of mu, delta and kappa opioid agonists and antagonists on mouse primary afferent neurons in culture. J. Pharmacol. Exp. Ther. 1987, 243, 258–263. [Google Scholar]
- Werz, M.A.; Macdonald, R.L. Heterogeneous sensitivity of cultured dorsal root ganglion neurones to opioid peptides selective for mu- and delta-opiate receptors. Nature 1982, 299, 730–733. [Google Scholar] [CrossRef][Green Version]
- Werz, M.A.; Macdonald, R.L. Dynorphin reduces calcium-dependent action potential duration by decreasing voltage-dependent calcium conductance. Neurosci. Lett. 1984, 46, 185–190. [Google Scholar] [CrossRef]
- Werz, M.A.; MacDonald, R.L. Opioid peptides selective for mu- and delta-opiate receptors reduce calcium-dependent action potential duration by increasing potassium conductance. Neurosci. Lett. 1983, 42, 173–178. [Google Scholar] [CrossRef]
- Macdonald, R.L.; Werz, M.A. Dynorphin A decreases voltage-dependent calcium conductance of mouse dorsal root ganglion neurones. J. Physiol. 1986, 377, 237–249. [Google Scholar] [CrossRef]
- Andrade, A.; Denome, S.; Jiang, Y.Q.; Marangoudakis, S.; Lipscombe, D. Opioid inhibition of N-type Ca2+ channels and spinal analgesia couple to alternative splicing. Nat. Neurosci. 2010, 13, 1249–1256. [Google Scholar] [CrossRef]
- Gross, R.A.; Macdonald, R.L. Dynorphin A selectively reduces a large transient (N-type) calcium current of mouse dorsal root ganglion neurons in cell culture. Proc. Natl. Acad. Sci. USA 1987, 84, 5469–5473. [Google Scholar] [CrossRef]
- Heinke, B.; Gingl, E.; Sandkühler, J. Multiple targets of μ-opioid receptor-mediated presynaptic inhibition at primary afferent Aδ- and C-fibers. J. Neurosci. Off. J. Soc. Neurosci. 2011, 31, 1313–1322. [Google Scholar] [CrossRef]
- King, A.P.; Hall, K.E.; Macdonald, R.L. kappa- and mu-Opioid inhibition of N-type calcium currents is attenuated by 4beta-phorbol 12-myristate 13-acetate and protein kinase C in rat dorsal root ganglion neurons. J. Pharmacol. Exp. Ther. 1999, 289, 312–320. [Google Scholar]
- Moises, H.C.; Rusin, K.I.; Macdonald, R.L. Mu- and kappa-opioid receptors selectively reduce the same transient components of high-threshold calcium current in rat dorsal root ganglion sensory neurons. J. Neurosci. Off. J. Soc. Neurosci. 1994, 14, 5903–5916. [Google Scholar] [CrossRef]
- Su, X.; Wachtel, R.E.; Gebhart, G.F. Inhibition of calcium currents in rat colon sensory neurons by K- but not mu- or delta-opioids. J. Neurophysiol. 1998, 80, 3112–3119. [Google Scholar] [CrossRef]
- Wiley, J.W.; Moises, H.C.; Gross, R.A.; MacDonald, R.L. Dynorphin A-mediated reduction in multiple calcium currents involves a G(o) alpha-subtype G protein in rat primary afferent neurons. J. Neurophysiol. 1997, 77, 1338–1348. [Google Scholar] [CrossRef][Green Version]
- Dolphin, A.C.; McGuirk, S.M.; Scott, R.H. An investigation into the mechanisms of inhibition of calcium channel currents in cultured sensory neurones of the rat by guanine nucleotide analogues and (-)-baclofen. Br. J. Pharmacol. 1989, 97, 263–273. [Google Scholar] [CrossRef]
- Dolphin, A.C.; Forda, S.R.; Scott, R.H. Calcium-dependent currents in cultured rat dorsal root ganglion neurones are inhibited by an adenosine analogue. J. Physiol. 1986, 373, 47–61. [Google Scholar] [CrossRef]
- Cardenas, C.G.; Del Mar, L.P.; Scroggs, R.S. Variation in serotonergic inhibition of calcium channel currents in four types of rat sensory neurons differentiated by membrane properties. J. Neurophysiol. 1995, 74, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Del Mar, L.P.; Cardenas, C.G.; Scroggs, R.S. Serotonin inhibits high-threshold Ca2+ channel currents in capsaicin-sensitive acutely isolated adult rat DRG neurons. J. Neurophysiol. 1994, 72, 2551–2554. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, K.; Fischbach, G.D. Neurotransmitters decrease the calcium conductance activated by depolarization of embryonic chick sensory neurones. J. Physiol. 1981, 317, 519–535. [Google Scholar] [CrossRef]
- Holz, G.G.; Shefner, S.A.; Anderson, E.G. Serotonin decreases the duration of action potentials recorded from tetraethylammonium-treated bullfrog dorsal root ganglion cells. J. Neurosci. Off. J. Soc. Neurosci. 1986, 6, 620–626. [Google Scholar] [CrossRef]
- Gerevich, Z.; Borvendeg, S.J.; Schröder, W.; Franke, H.; Wirkner, K.; Nörenberg, W.; Fürst, S.; Gillen, C.; Illes, P. Inhibition of N-type voltage-activated calcium channels in rat dorsal root ganglion neurons by P2Y receptors is a possible mechanism of ADP-induced analgesia. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.A.; Coutts, A.A.; McFarlane, S.M.; Anavi-Goffer, S.; Irving, A.J.; Pertwee, R.G.; MacEwan, D.J.; Scott, R.H. Actions of cannabinoid receptor ligands on rat cultured sensory neurones: Implications for antinociception. Neuropharmacology 2001, 40, 221–232. [Google Scholar] [CrossRef]
- Bleakman, D.; Colmers, W.F.; Fournier, A.; Miller, R.J. Neuropeptide Y inhibits Ca2+ influx into cultured dorsal root ganglion neurones of the rat via a Y2 receptor. Br. J. Pharmacol. 1991, 103, 1781–1789. [Google Scholar] [CrossRef] [PubMed]
- Ewald, D.A.; Matthies, H.J.; Perney, T.M.; Walker, M.W.; Miller, R.J. The effect of down regulation of protein kinase C on the inhibitory modulation of dorsal root ganglion neuron Ca2+ currents by neuropeptide Y. J. Neurosci. Off. J. Soc. Neurosci. 1988, 8, 2447–2451. [Google Scholar] [CrossRef]
- Walker, M.W.; Ewald, D.A.; Perney, T.M.; Miller, R.J. Neuropeptide Y modulates neurotransmitter release and Ca2+ currents in rat sensory neurons. J. Neurosci. Off. J. Soc. Neurosci. 1988, 8, 2438–2446. [Google Scholar] [CrossRef]
- Gorham, L.; Just, S.; Doods, H. Somatostatin 4 receptor activation modulates G-protein coupled inward rectifying potassium channels and voltage stimulated calcium signals in dorsal root ganglion neurons. Eur. J. Pharmacol. 2014, 736, 101–106. [Google Scholar] [CrossRef]
- Abdulla, F.A.; Smith, P.A. Ectopic alpha2-adrenoceptors couple to N-type Ca2+ channels in axotomized rat sensory neurons. J. Neurosci. Off. J. Soc. Neurosci. 1997, 17, 1633–1641. [Google Scholar] [CrossRef]
- Gamper, N.; Reznikov, V.; Yamada, Y.; Yang, J.; Shapiro, M.S. Phosphatidylinositol [correction] 4,5-bisphosphate signals underlie receptor-specific Gq/11-mediated modulation of N-type Ca2+ channels. J. Neurosci. Off. J. Soc. Neurosci. 2004, 24, 10980–10992. [Google Scholar] [CrossRef] [PubMed]
- Lechner, S.G.; Hussl, S.; Schicker, K.W.; Drobny, H.; Boehm, S. Presynaptic inhibition via a phospholipase C- and phosphatidylinositol bisphosphate-dependent regulation of neuronal Ca2+ channels. Mol. Pharmacol. 2005, 68, 1387–1396. [Google Scholar] [CrossRef]
- Wu, L.; Bauer, C.S.; Zhen, X.G.; Xie, C.; Yang, J. Dual regulation of voltage-gated calcium channels by PtdIns(4,5)P2. Nature 2002, 419, 947–952. [Google Scholar] [CrossRef]
- Li, Z.; He, S.Q.; Tseng, P.Y.; Xu, Q.; Tiwari, V.; Yang, F.; Shu, B.; Zhang, T.; Tang, Z.; Raja, S.N.; et al. The inhibition of high-voltage-activated calcium current by activation of MrgC11 involves phospholipase C-dependent mechanisms. Neuroscience 2015, 300, 393–403. [Google Scholar] [CrossRef][Green Version]
- Huang, D.; Huang, S.; Peers, C.; Du, X.; Zhang, H.; Gamper, N. GABAB receptors inhibit low-voltage activated and high-voltage activated Ca(2+) channels in sensory neurons via distinct mechanisms. Biochem. Biophys. Res. Commun. 2015, 465, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, F.; Flockerzi, V.; Kahl, S.; Wegener, J.W. L-type CaV1.2 calcium channels: From in vitro findings to in vivo function. Physiol. Rev. 2014, 94, 303–326. [Google Scholar] [CrossRef]
- Gutman, G.A.; Chandy, K.G.; Adelman, J.P.; Aiyar, J.; Bayliss, D.A.; Clapham, D.E.; Covarriubias, M.; Desir, G.V.; Furuichi, K.; Ganetzky, B.; et al. International Union of Pharmacology. XLI. Compendium of voltage-gated ion channels: Potassium channels. Pharmacol. Rev. 2003, 55, 583–586. [Google Scholar] [CrossRef]
- Tsantoulas, C.; McMahon, S.B. Opening paths to novel analgesics: The role of potassium channels in chronic pain. Trends Neurosci. 2014, 37, 146–158. [Google Scholar] [CrossRef]
- Swartz, K.J. Towards a structural view of gating in potassium channels. Nat. Reviews. Neurosci. 2004, 5, 905–916. [Google Scholar] [CrossRef]
- Kuang, Q.; Purhonen, P.; Hebert, H. Structure of potassium channels. Cell. Mol. Life Sci. CMLS 2015, 72, 3677–3693. [Google Scholar] [CrossRef]
- Brown, D.A.; Passmore, G.M. Neural KCNQ (Kv7) channels. Br. J. Pharmacol. 2009, 156, 1185–1195. [Google Scholar] [CrossRef]
- Du, X.; Gao, H.; Jaffe, D.; Zhang, H.; Gamper, N. M-type K+ channels in peripheral nociceptive pathways. Br. J. Pharmacol. 2018, 175, 2158–2172. [Google Scholar] [CrossRef]
- Howard, R.J.; Clark, K.A.; Holton, J.M.; Minor, D.L. Structural insight into KCNQ (Kv7) channel assembly and channelopathy. Neuron 2007, 53, 663–675. [Google Scholar] [CrossRef]
- Delmas, P.; Brown, D.A. Pathways modulating neural KCNQ/M (Kv7) potassium channels. Nat. Rev. Neurosci. 2005, 6, 850–862. [Google Scholar] [CrossRef]
- Du, X.; Hao, H.; Gigout, S.; Huang, D.; Yang, Y.; Li, L.; Wang, C.; Sundt, D.; Jaffe, D.B.; Zhang, H.; Gamper, N. Control of somatic membrane potential in nociceptive neurons and its implications for peripheral nociceptive transmission. Pain 2014, 155, 2306–2322. [Google Scholar] [CrossRef]
- Rivera-Arconada, I.; Roza, C.; Lopez-Garcia, J.A. Enhancing m currents: A way out for neuropathic pain? Front. Mol. Neurosci. 2009, 2, 10. [Google Scholar] [CrossRef]
- Passmore, G.M.; Selyanko, A.A.; Mistry, M.; Al-Qatari, M.; Marsh, S.J.; Matthews, E.A.; Dickenson, A.H.; Brown, T.A.; Burbidge, S.A.; Main, M.; et al. KCNQ/M currents in sensory neurons: Significance for pain therapy. J. Neurosci. Off. J. Soc. Neurosci. 2003, 23, 7227–7236. [Google Scholar] [CrossRef]
- Zheng, Y.; Xu, H.; Zhan, L.; Zhou, X.; Chen, X.; Gao, Z. Activation of peripheral KCNQ channels relieves gout pain. Pain 2015, 156, 1025–1035. [Google Scholar] [CrossRef]
- Ray, S.; Salzer, I.; Kronschläger, M.T.; Boehm, S. The paracetamol metabolite N-acetylp-benzoquinone imine reduces excitability in first- and second-order neurons of the pain pathway through actions on KV7 channels. Pain 2019, 160, 954–964. [Google Scholar] [CrossRef]
- Zheng, Q.; Fang, D.; Liu, M.; Cai, J.; Wan, Y.; Han, J.S.; Xing, G.G. Suppression of KCNQ/M (Kv7) potassium channels in dorsal root ganglion neurons contributes to the development of bone cancer pain in a rat model. Pain 2013, 154, 434–448. [Google Scholar] [CrossRef]
- Yu, T.; Li, L.; Liu, H.; Li, H.; Liu, Z.; Li, Z. KCNQ2/3/5 channels in dorsal root ganglion neurons can be therapeutic targets of neuropathic pain in diabetic rats. Mol. Pain 2018, 14, 1744806918793229. [Google Scholar] [CrossRef]
- Brown, B.S.; Yu, S.P. Modulation and genetic identification of the M channel. Prog. Biophys. Mol. Biol. 2000, 73, 135–166. [Google Scholar] [CrossRef]
- Ishimatsu, M. Substance P produces an inward current by suppressing voltage-dependent and -independent K+ currents in bullfrog primary afferent neurons. Neurosci. Res. 1994, 19, 9–20. [Google Scholar] [CrossRef]
- Akasu, T.; Ishimatsu, M.; Yamada, K. Tachykinins cause inward current through NK1 receptors in bullfrog sensory neurons. Brain Res. 1996, 713, 160–167. [Google Scholar] [CrossRef]
- Lin, C.C.J.; Chen, W.N.; Chen, C.J.; Lin, Y.W.; Zimmer, A.; Chen, C.C. An antinociceptive role for substance P in acid-induced chronic muscle pain. Proc. Natl. Acad. Sci. USA 2012, 109, E76–E83. [Google Scholar] [CrossRef]
- Neves, S.R.; Ram, P.T.; Iyengar, R. G protein pathways. Science 2002, 296, 1636–1639. [Google Scholar] [CrossRef]
- Suh, B.C.; Hille, B. Recovery from muscarinic modulation of M current channels requires phosphatidylinositol 4,5-bisphosphate synthesis. Neuron 2002, 35, 507–520. [Google Scholar] [CrossRef]
- Higashida, H.; Brown, D.A. Two polyphosphatidylinositide metabolites control two K+ currents in a neuronal cell. Nature 1986, 323, 333–335. [Google Scholar] [CrossRef]
- Selyanko, A.A.; Brown, D.A. Intracellular calcium directly inhibits potassium M channels in excised membrane patches from rat sympathetic neurons. Neuron 1996, 16, 151–162. [Google Scholar] [CrossRef]
- Gamper, N.; Shapiro, M.S. Calmodulin mediates Ca2+-dependent modulation of M-type K+ channels. J. Gen. Physiol. 2003, 122, 17–31. [Google Scholar] [CrossRef]
- Brown, D.A.; Hughes, S.A.; Marsh, S.J.; Tinker, A. Regulation of M(Kv7.2/7.3) channels in neurons by PIP(2) and products of PIP(2) hydrolysis: Significance for receptor-mediated inhibition. J. Physiol. 2007, 582, 917–925. [Google Scholar] [CrossRef]
- Soh, U.J.; Dores, M.R.; Chen, B.; Trejo, J. Signal transduction by protease-activated receptors. Br. J. Pharmacol. 2010, 160, 191–203. [Google Scholar] [CrossRef]
- Linley, J.E.; Rose, K.; Patil, M.; Robertson, B.; Akopian, A.N.; Gamper, N. Inhibition of M current in sensory neurons by exogenous proteases: A signaling pathway mediating inflammatory nociception. J. Neurosci. Off. J. Soc. Neurosci. 2008, 28, 11240–11249. [Google Scholar] [CrossRef]
- von Kügelgen, I. Pharmacological profiles of cloned mammalian P2Y-receptor subtypes. Pharmacol. Ther. 2006, 110, 415–432. [Google Scholar] [CrossRef]
- Dong, X.; Han, S.; Zylka, M.J.; Simon, M.I.; Anderson, D.J. A diverse family of GPCRs expressed in specific subsets of nociceptive sensory neurons. Cell 2001, 106, 619–632. [Google Scholar] [CrossRef]
- Choi, S.S.; Lahn, B.T. Adaptive evolution of MRG, a neuron-specific gene family implicated in nociception. Genome Res. 2003, 13, 2252–2259. [Google Scholar] [CrossRef]
- Zhang, L.; Taylor, N.; Xie, Y.; Ford, R.; Johnson, J.; Paulsen, J.E.; Bates, B. Cloning and expression of MRG receptors in macaque, mouse, and human. Brain Res. Mol. Brain Res. 2005, 133, 187–197. [Google Scholar] [CrossRef]
- Zylka, M.J.; Rice, F.L.; Anderson, D.J. Topographically distinct epidermal nociceptive circuits revealed by axonal tracers targeted to Mrgprd. Neuron 2005, 45, 17–25. [Google Scholar] [CrossRef]
- Crozier, R.A.; Ajit, S.K.; Kaftan, E.J.; Pausch, M.H. MrgD activation inhibits KCNQ/M-currents and contributes to enhanced neuronal excitability. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 4492–4496. [Google Scholar] [CrossRef]
- Baldwin, T.J.; Tsaur, M.L.; Lopez, G.A.; Jan, Y.N.; Jan, L.Y. Characterization of a mammalian cDNA for an inactivating voltage-sensitive K+ channel. Neuron 1991, 7, 471–483. [Google Scholar] [CrossRef]
- Shibata, R.; Nakahira, K.; Shibasaki, K.; Wakazono, Y.; Imoto, K.; Ikenaka, K. A-type K+ current mediated by the Kv4 channel regulates the generation of action potential in developing cerebellar granule cells. J. Neurosci. Off. J. Soc. Neurosci. 2000, 20, 4145–4155. [Google Scholar] [CrossRef]
- Vydyanathan, A.; Wu, Z.Z.; Chen, S.R.; Pan, H.L. A-type voltage-gated K+ currents influence firing properties of isolectin B4-positive but not isolectin B4-negative primary sensory neurons. J. Neurophysiol. 2005, 93, 3401–3409. [Google Scholar] [CrossRef]
- Chien, L.Y.; Cheng, J.K.; Chu, D.; Cheng, C.F.; Tsaur, M.L. Reduced expression of A-type potassium channels in primary sensory neurons induces mechanical hypersensitivity. J. Neurosci. Off. J. Soc. Neurosci. 2007, 27, 9855–9865. [Google Scholar] [CrossRef]
- Liu, P.W.; Blair, N.T.; Bean, B.P. Action Potential Broadening in Capsaicin-Sensitive DRG Neurons from Frequency-Dependent Reduction of Kv3 Current. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 9705–9714. [Google Scholar] [CrossRef]
- Zemel, B.M.; Ritter, D.M.; Covarrubias, M.; Muqeem, T. A-Type KV Channels in Dorsal Root Ganglion Neurons: Diversity, Function, and Dysfunction. Front. Mol. Neurosci. 2018, 11, 253. [Google Scholar] [CrossRef]
- Ritter, D.M.; Ho, C.; O’Leary, M.E.; Covarrubias, M. Modulation of Kv3.4 channel N-type inactivation by protein kinase C shapes the action potential in dorsal root ganglion neurons. J. Physiol. 2012, 590, 145–161. [Google Scholar] [CrossRef]
- Ritter, D.M.; Zemel, B.M.; Hala, T.J.; O’Leary, M.E.; Lepore, A.C.; Covarrubias, M. Dysregulation of Kv3.4 channels in dorsal root ganglia following spinal cord injury. J. Neurosci. Off. J. Soc. Neurosci. 2015, 35, 1260–1273. [Google Scholar] [CrossRef]
- Zemel, B.M.; Muqeem, T.; Brown, E.V.; Goulão, M.; Urban, M.W.; Tymanskyj, S.R.; Lepore, A.C.; Covarrubias, M. Calcineurin Dysregulation Underlies Spinal Cord Injury-Induced K+ Channel Dysfunction in DRG Neurons. J. Neurosci. Off. J. Soc. Neurosci. 2017, 37, 8256–8272. [Google Scholar] [CrossRef]
- Zhang, Y.; Jiang, D.; Zhang, Y.; Jiang, X.; Wang, F.; Tao, J. Neuromedin U type 1 receptor stimulation of A-type K+ current requires the βγ subunits of Go protein, protein kinase A, and extracellular signal-regulated kinase 1/2 (ERK1/2) in sensory neurons. J. Biol. Chem. 2012, 287, 18562–18572. [Google Scholar] [CrossRef]
- Saetrum Opgaard, O.; Nothacker, H.; Ehlert, F.J.; Krause, D.N. Human urotensin II mediates vasoconstriction via an increase in inositol phosphates. Eur. J. Pharmacol. 2000, 406, 265–271. [Google Scholar] [CrossRef]
- Maguire, J.J.; Davenport, A.P. Is urotensin-II the new endothelin? Br. J. Pharmacol. 2002, 137, 579–588. [Google Scholar] [CrossRef]
- Zhang, M.; Gao, C.X.; Wang, Y.P.; Ma, K.T.; Li, L.; Yin, J.W.; Dai, Z.G.; Wang, S.; Si, J.Q. The association between the expression of PAR2 and TMEM16A and neuropathic pain. Mol. Med. Rep. 2018, 17, 3744–3750. [Google Scholar] [CrossRef]
- Li, N.; Lu, Z.Y.; Yu, L.H.; Burnstock, G.; Deng, X.M.; Ma, B. Inhibition of G protein-coupled P2Y2 receptor induced analgesia in a rat model of trigeminal neuropathic pain. Mol. Pain 2014, 10, 21. [Google Scholar] [CrossRef]
- Jacobson, K.A.; Ivanov, A.A.; de Castro, S.; Harden, T.K.; Ko, H. Development of selective agonists and antagonists of P2Y receptors. Purinergic Signal. 2009, 5, 75–89. [Google Scholar] [CrossRef]
- Glaaser, I.W.; Slesinger, P.A. Structural Insights into GIRK Channel Function. Int. Rev. Neurobiol. 2015, 123, 117–160. [Google Scholar]
- Nagi, K.; Pineyro, G. Kir3 channel signaling complexes: Focus on opioid receptor signaling. Front. Cell. Neurosci. 2014, 8, 186. [Google Scholar] [CrossRef]
- Peleg, S.; Varon, D.; Ivanina, T.; Dessauer, C.W.; Dascal, N. G(alpha)(i) controls the gating of the G protein-activated K(+) channel, GIRK. Neuron 2002, 33, 87–99. [Google Scholar] [CrossRef]
- Huang, C.L.; Feng, S.; Hilgemann, D.W. Direct activation of inward rectifier potassium channels by PIP2 and its stabilization by Gbetagamma. Nature 1998, 391, 803–806. [Google Scholar] [CrossRef]
- Sui, J.L.; Petit-Jacques, J.; Logothetis, D.E. Activation of the atrial KACh channel by the betagamma subunits of G proteins or intracellular Na+ ions depends on the presence of phosphatidylinositol phosphates. Proc. Natl. Acad. Sci. USA 1998, 95, 1307–1312. [Google Scholar] [CrossRef]
- Khodorova, A.; Navarro, B.; Jouaville, L.S.; Murphy, J.E.; Rice, F.L.; Mazurkiewicz, J.E.; Long-Woodward, D.; Stoffel, M.; Strichartz, G.R.; Yukhananov, R.; et al. Endothelin-B receptor activation triggers an endogenous analgesic cascade at sites of peripheral injury. Nat. Med. 2003, 9, 1055–1061. [Google Scholar] [CrossRef]
- Gao, X.F.; Zhang, H.L.; You, Z.D.; Lu, C.L.; He, C. G protein-coupled inwardly rectifying potassium channels in dorsal root ganglion neurons. Acta Pharmacol. Sin. 2007, 28, 185–190. [Google Scholar] [CrossRef]
- Stötzner, P.; Spahn, V.; Celik, M.Ö.; Labuz, D.; Machelska, H. Mu-Opioid Receptor Agonist Induces Kir3 Currents in Mouse Peripheral Sensory Neurons - Effects of Nerve Injury. Front. Pharmacol. 2018, 9, 1478. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GPCR Ligand | Involved GPCR | Pathway | Effect on TRPV1 | Reference |
|---|---|---|---|---|
| Bradykinin | B | G-DAG-PKC | increased current | [2,41] |
| Serotonin | 5-HT | G-DAG-PKC | increased current | [42,43,44] |
| 5-HT | G-AC-PKA | increased current | [43] | |
| 5-HT | G-AC-PKA | increased current | [42] | |
| UTP | P2Y | G-DAG-PKC | increased current | [45,46,47] |
| BAM 8-22 | MRGPRX1 | G-DAG-PKC-PIP | increased current | [48] |
| Proteases | PAR2 | G-PKC | increased current | [49,50] |
| PAR1 | G-PKC | increased current | [51] | |
| PAR4 | G-PKC | increased current | [51] | |
| PGE | EP | G-PKC | increased current | [52] |
| EP | G-PKA | increased current | [52,53,54] | |
| PGI | IP | G-PKC | increased current | [52] |
| IP | G-PKA | increased current | [52] | |
| Endothelin-1 | ET | G -PKC | increased current | [55,56] |
| Substance P | NK | G -PKC | increased current | [57] |
| NK | G-PKC | increased current | [58] | |
| H | TDAG8 (GPR65) | G-PKA | increased current | [59] |
| Morphine | MOP | G-reduced AC | decreased current | [60,61] |
| Endocannabinoids | CB | G-reduced AC | decreased current | [62] |
| CB | G-reduced AC | decreased current | [63] |
| Endogenous Ligand | GPCR | ASIC | CaCC | Ca | GIRK | KP | K1.4 K3.4 K4 | K7 | Na | TRPA1 TRPM3 TRPM8 TRPV1 | Piezo |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Adenosine | A | Ca | |||||||||
| A | TRPA1 | ||||||||||
| Alanine | MrgD | K7 | |||||||||
| BAM 8-22 | MRGPRX1 | TRPV1 | |||||||||
| Bradykinin | B | TRPA1 | |||||||||
| B | CaCC | K7 | TRPA1 | Piezo2 | |||||||
| TRPM8 | |||||||||||
| TRPV1 | |||||||||||
| CGRP | CGRP-R | Na | |||||||||
| Endo- | CB | ASIC | CaCC | TRPV1 | |||||||
| cannabinoids | CB | TRPV1 | |||||||||
| CB | Ca | ||||||||||
| Endothelin 1 | ET | TRPV1 | |||||||||
| GABA | GABA | Ca | |||||||||
| H | TDAG8/ | TRPV1 | |||||||||
| (GPR65) | |||||||||||
| Histamine | H | TRPA1 | |||||||||
| Neuromedin U | NMUR1 | K1.4 | |||||||||
| K3.4 | |||||||||||
| K4 | |||||||||||
| Neuropeptide Y | Y | Ca | |||||||||
| Noradrenaline | Ca | ||||||||||
| Ca | |||||||||||
| Nucleotides | P2Y | CaCC | Ca | K7 | |||||||
| P2Y | ASIC | CaCC | K1.4 | K7 | Piezo2 | ||||||
| K3.4 | |||||||||||
| K4 | |||||||||||
| P2Y | KP | ||||||||||
| Opioids | MOP | ASIC | Ca | GIRK | KP | Na | TRPV1 | ||||
| DOP | CaCC | Ca | |||||||||
| KOP | Ca | ||||||||||
| Prostaglandins | EP | TRPV1 | |||||||||
| EP | Na | TRPV1 | |||||||||
| IP | TRPV1 | ||||||||||
| FP | KP | ||||||||||
| Proteases | PAR1 | TRPV1 | |||||||||
| PAR2 | ASIC | CaCC | TRPV1 | ||||||||
| PAR4 | TRPV1 | ||||||||||
| Serotonin | 5-HT | ASIC | CaCC | TRPV1 | |||||||
| 5-HT | KP | Na | TRPV1 | ||||||||
| 5-HT | TRPV1 | ||||||||||
| 5-HT | Ca | ||||||||||
| Somatostatin | SST | Ca | |||||||||
| Substance P/ | NK | K7 | Na | TRPV1 | |||||||
| neurokinin A | NK | TRPV1 | |||||||||
| Urotensin | UTR | K1.4 | |||||||||
| K3.4 | |||||||||||
| K4 | |||||||||||
| ? | MrgC | Ca |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salzer, I.; Ray, S.; Schicker, K.; Boehm, S. Nociceptor Signalling through ion Channel Regulation via GPCRs. Int. J. Mol. Sci. 2019, 20, 2488. https://doi.org/10.3390/ijms20102488
Salzer I, Ray S, Schicker K, Boehm S. Nociceptor Signalling through ion Channel Regulation via GPCRs. International Journal of Molecular Sciences. 2019; 20(10):2488. https://doi.org/10.3390/ijms20102488
Chicago/Turabian StyleSalzer, Isabella, Sutirtha Ray, Klaus Schicker, and Stefan Boehm. 2019. "Nociceptor Signalling through ion Channel Regulation via GPCRs" International Journal of Molecular Sciences 20, no. 10: 2488. https://doi.org/10.3390/ijms20102488
APA StyleSalzer, I., Ray, S., Schicker, K., & Boehm, S. (2019). Nociceptor Signalling through ion Channel Regulation via GPCRs. International Journal of Molecular Sciences, 20(10), 2488. https://doi.org/10.3390/ijms20102488