
Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii

Abstract
:1. Introduction
2. Results
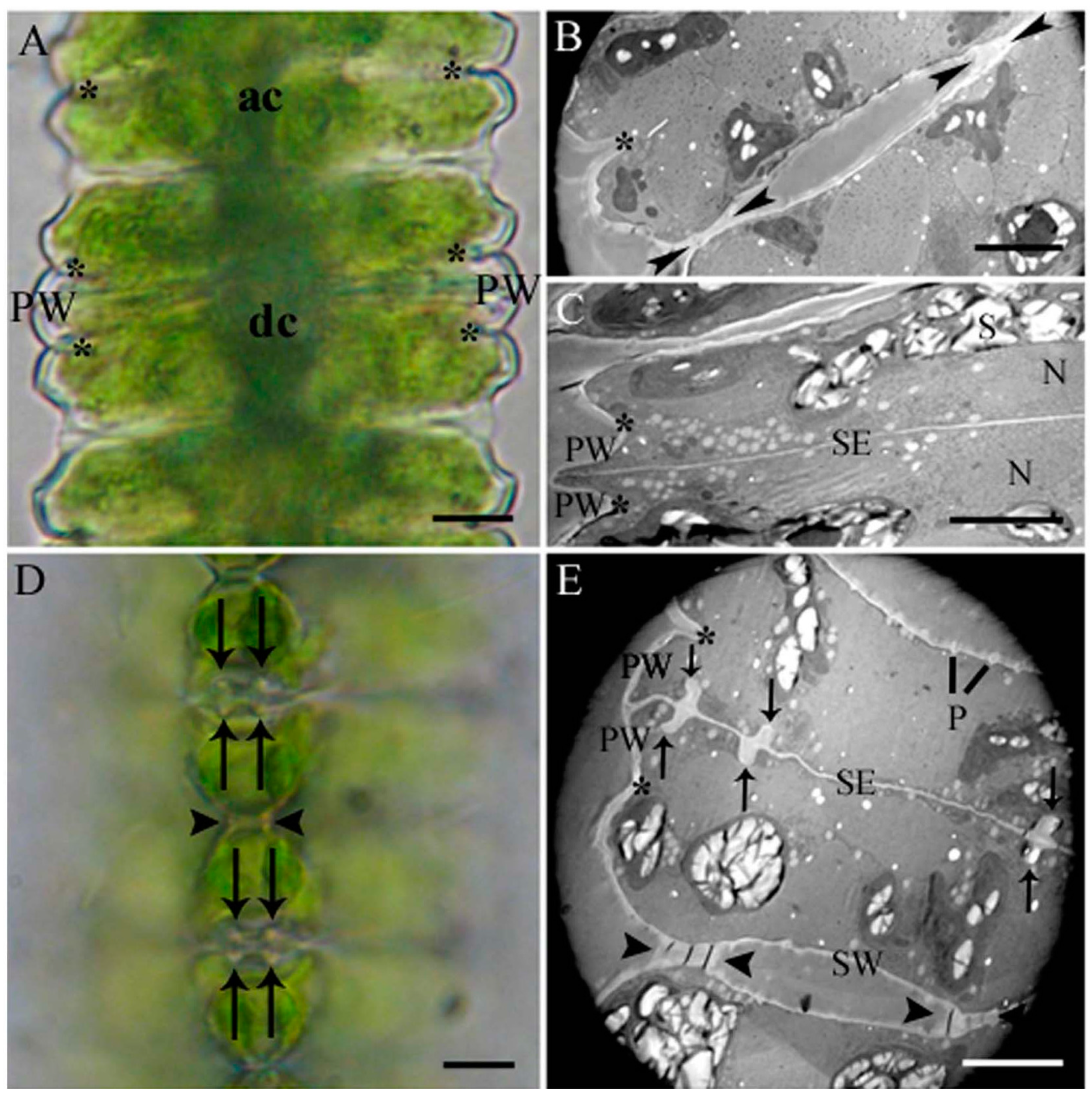
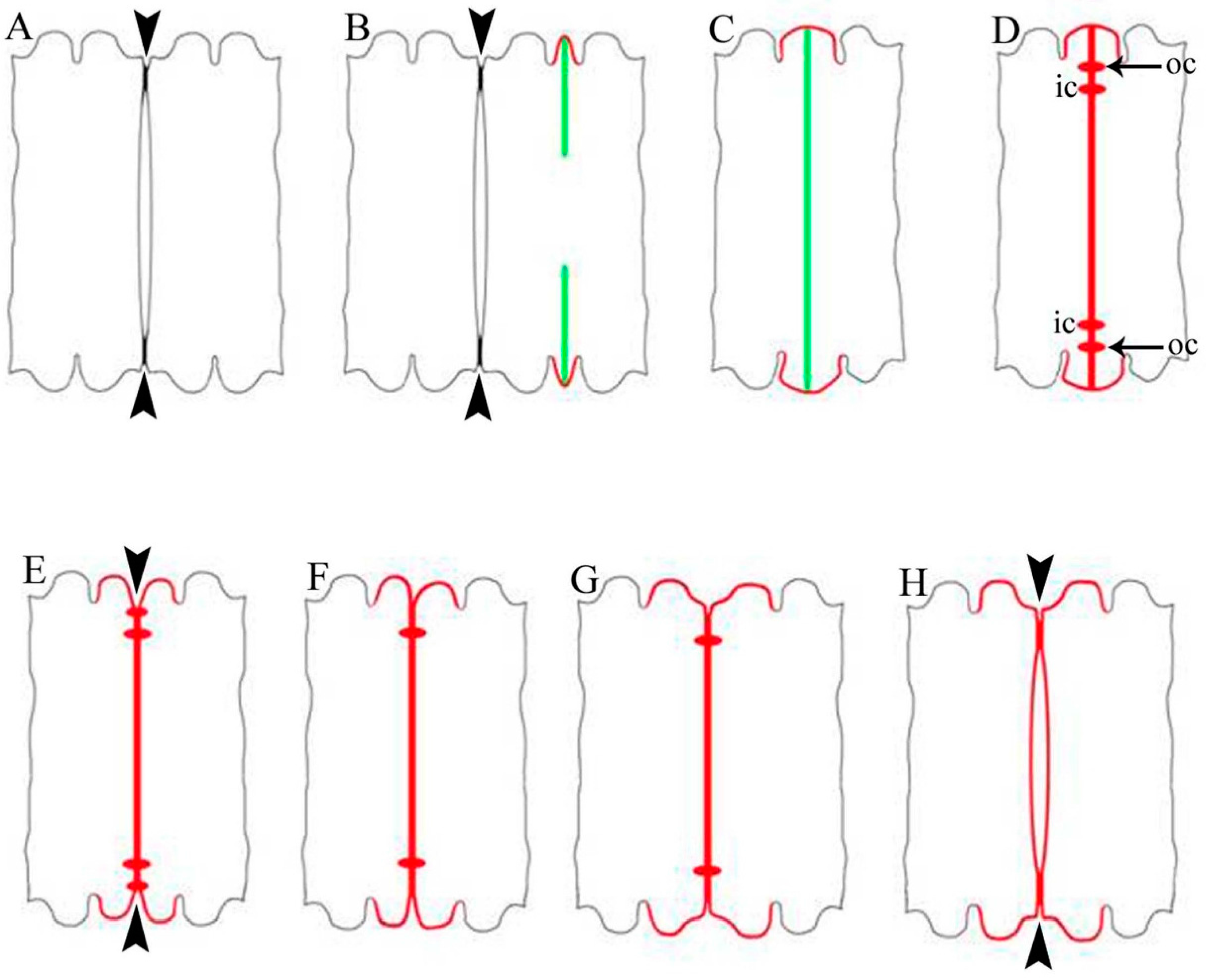
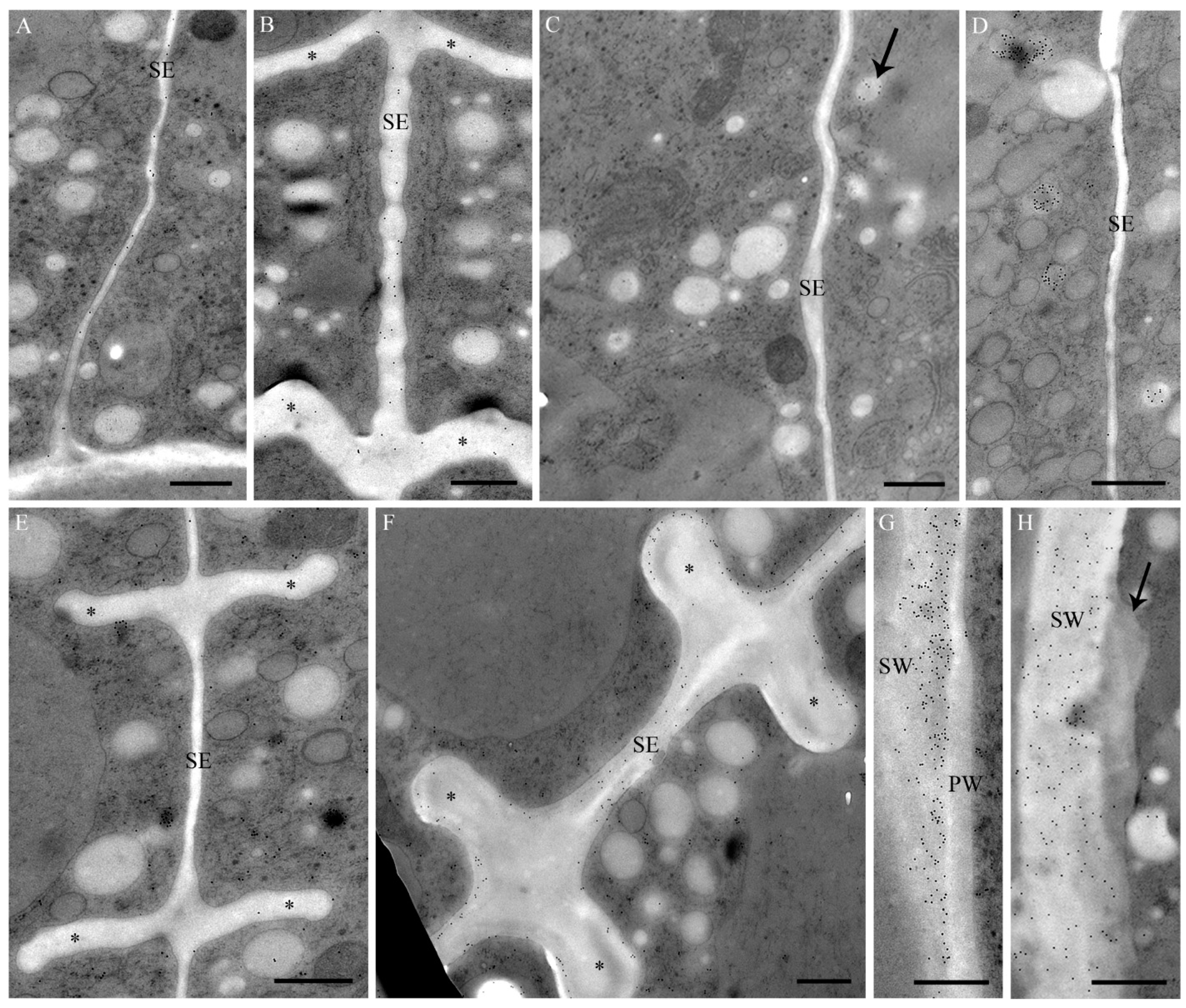
2.1. Cell Development and Cell Wall Composition of Untreated Desmidium Filaments
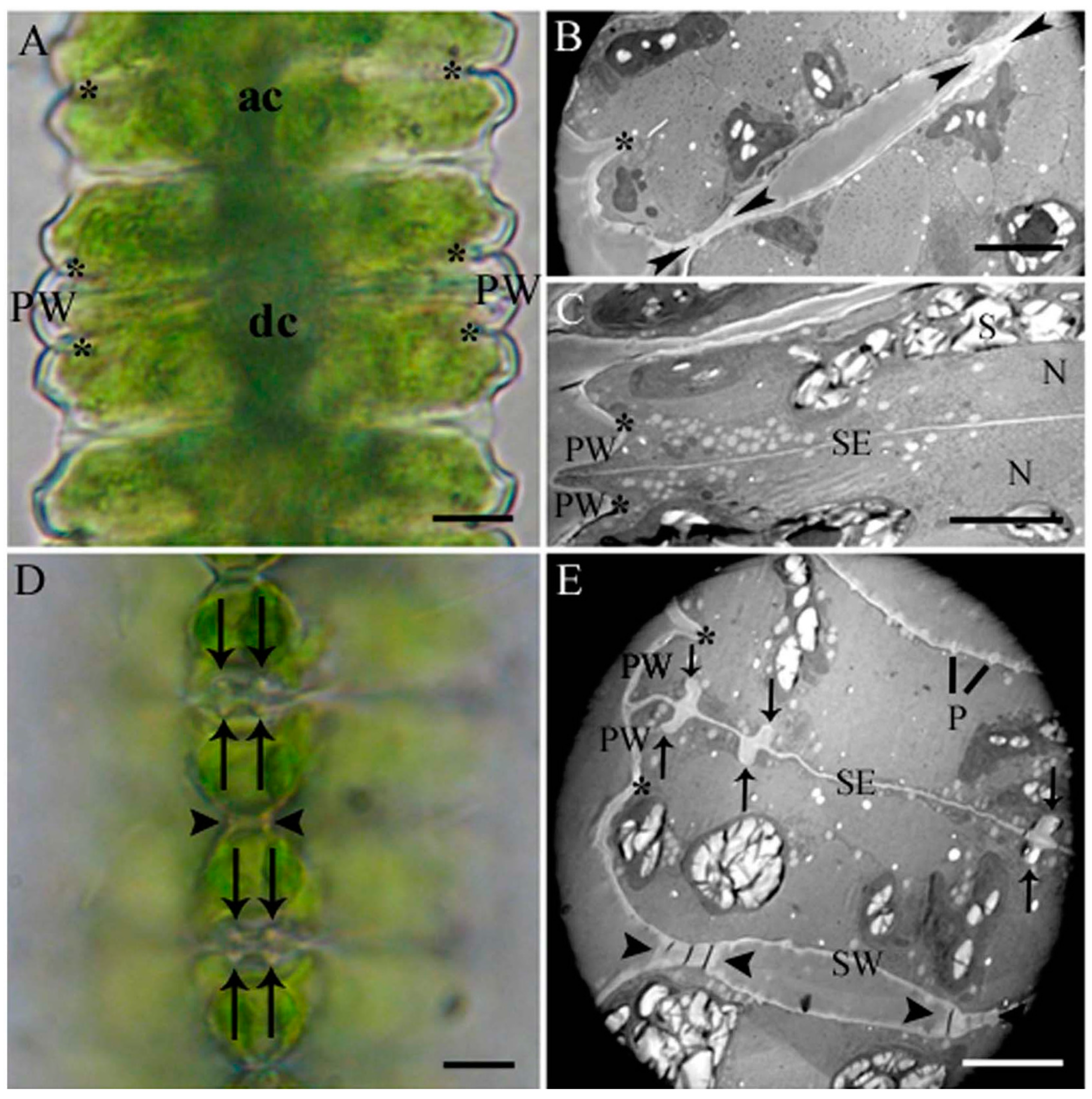
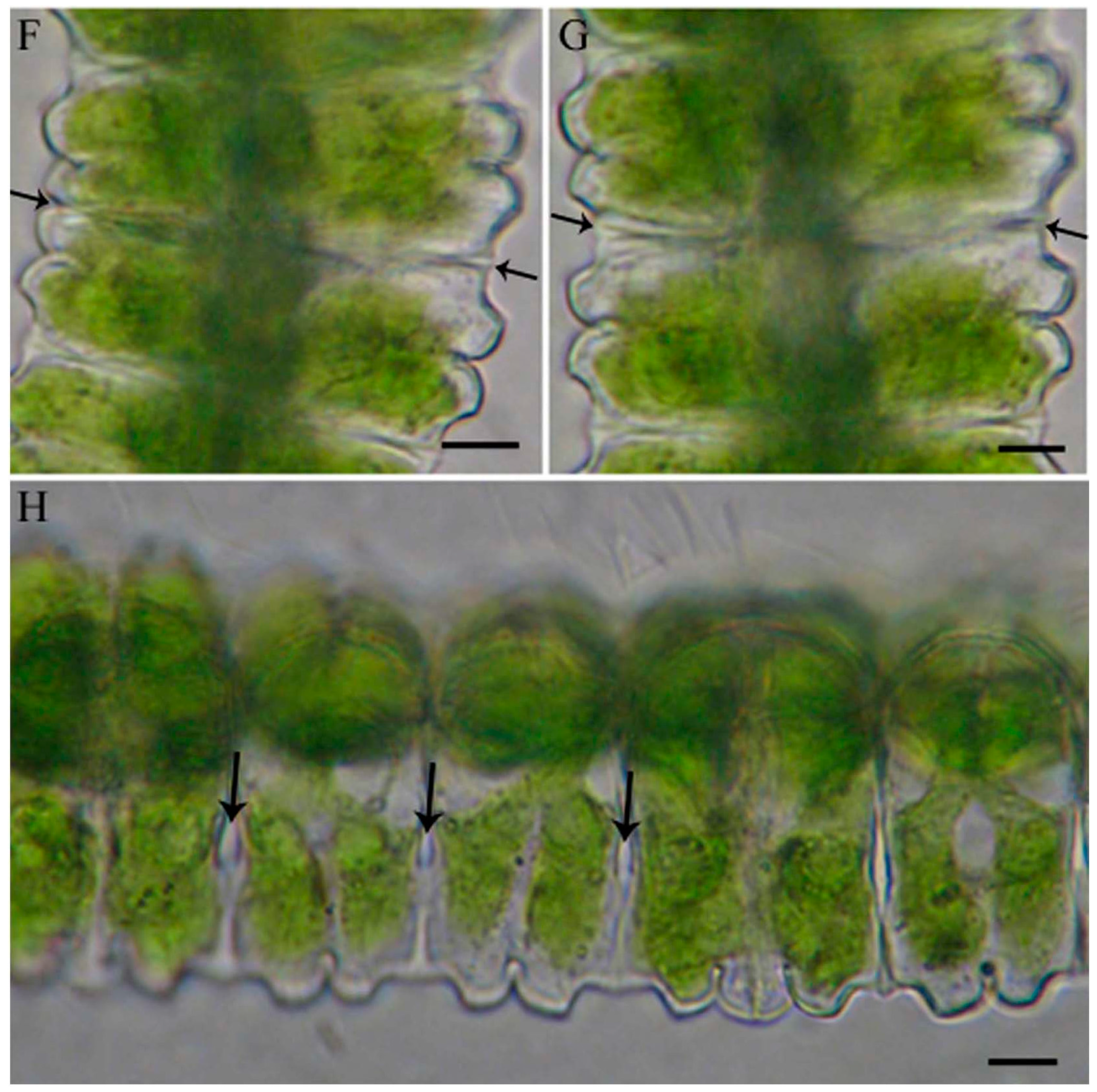
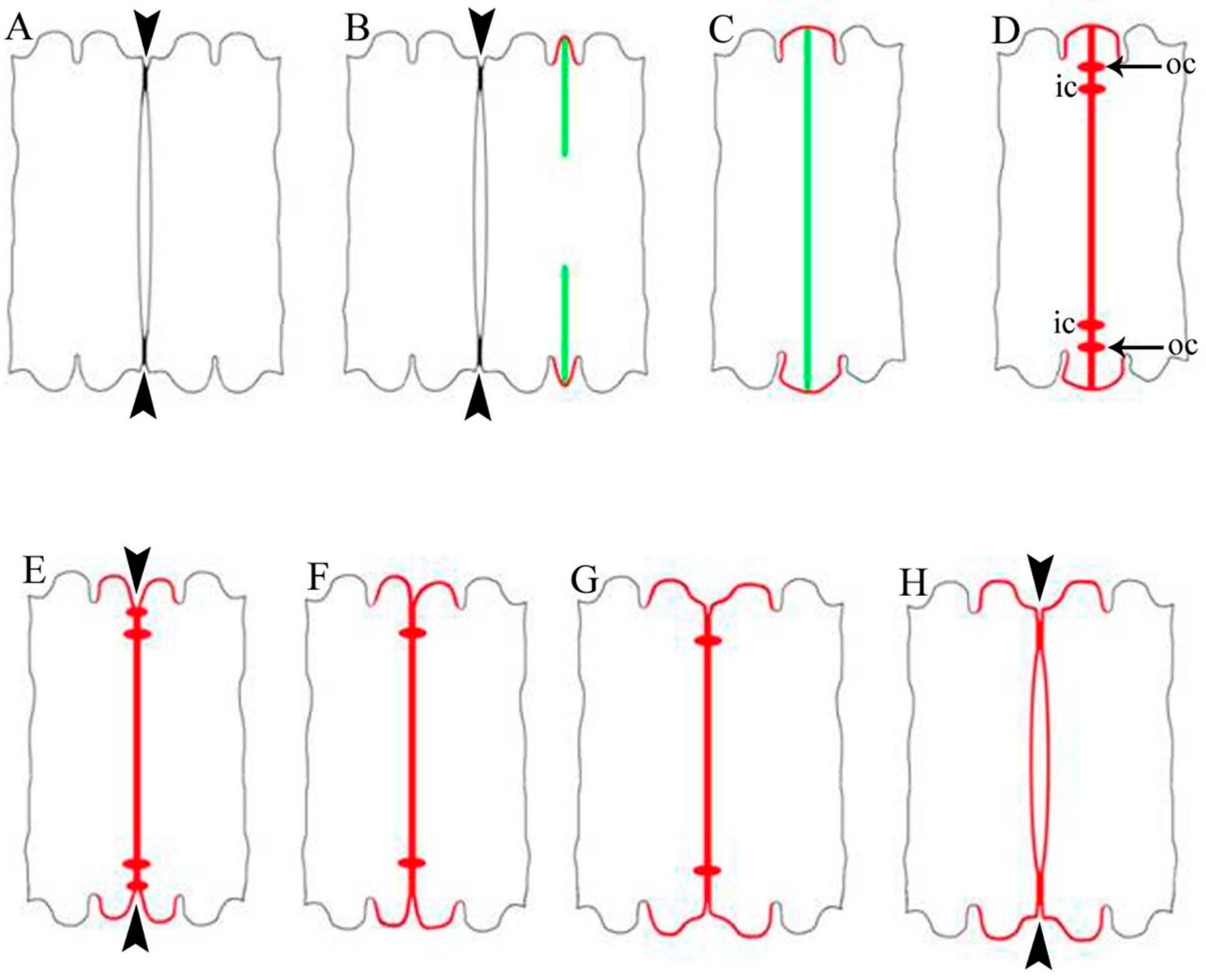
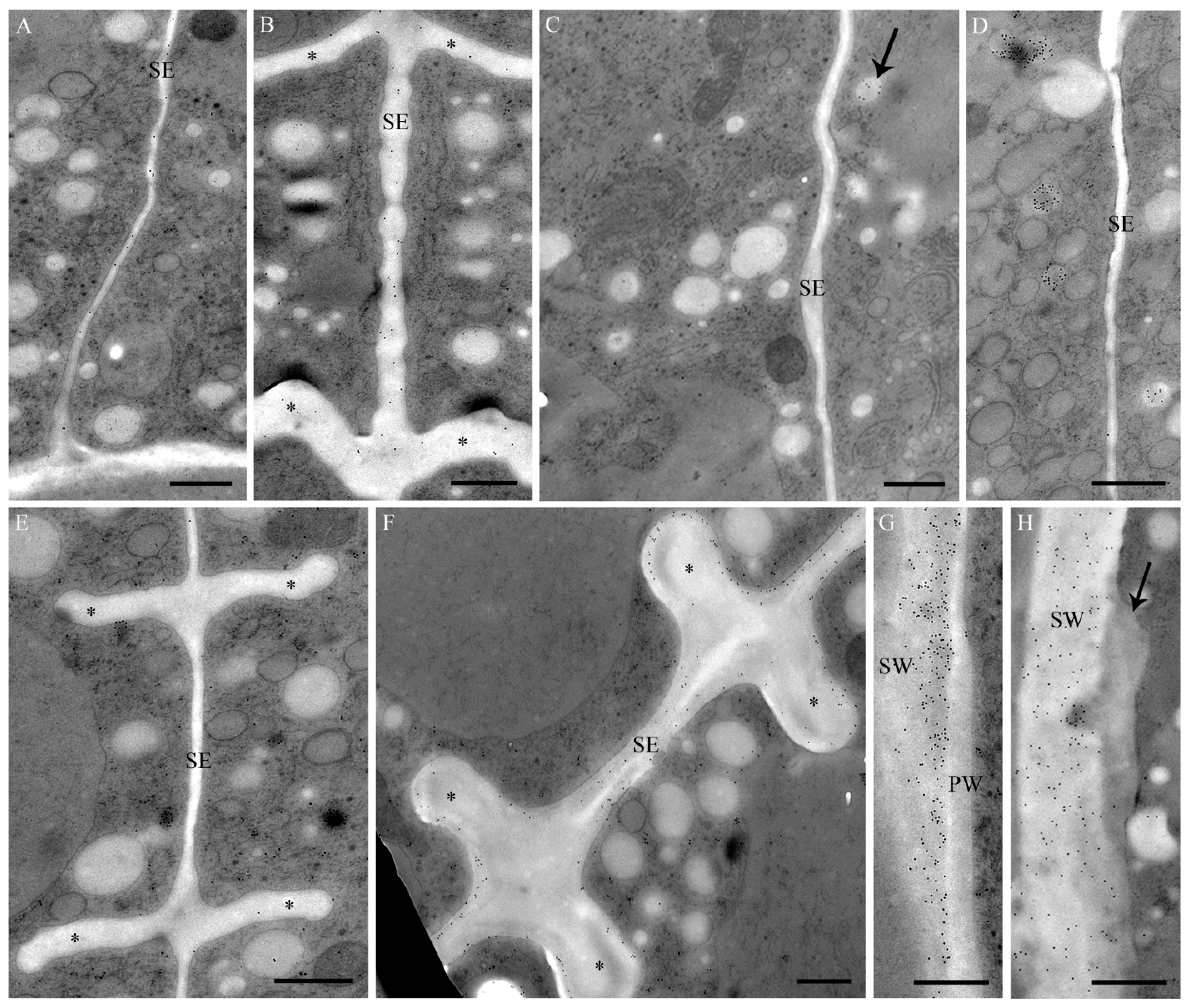
2.2. Heavy Metal Effects on Biomass Production
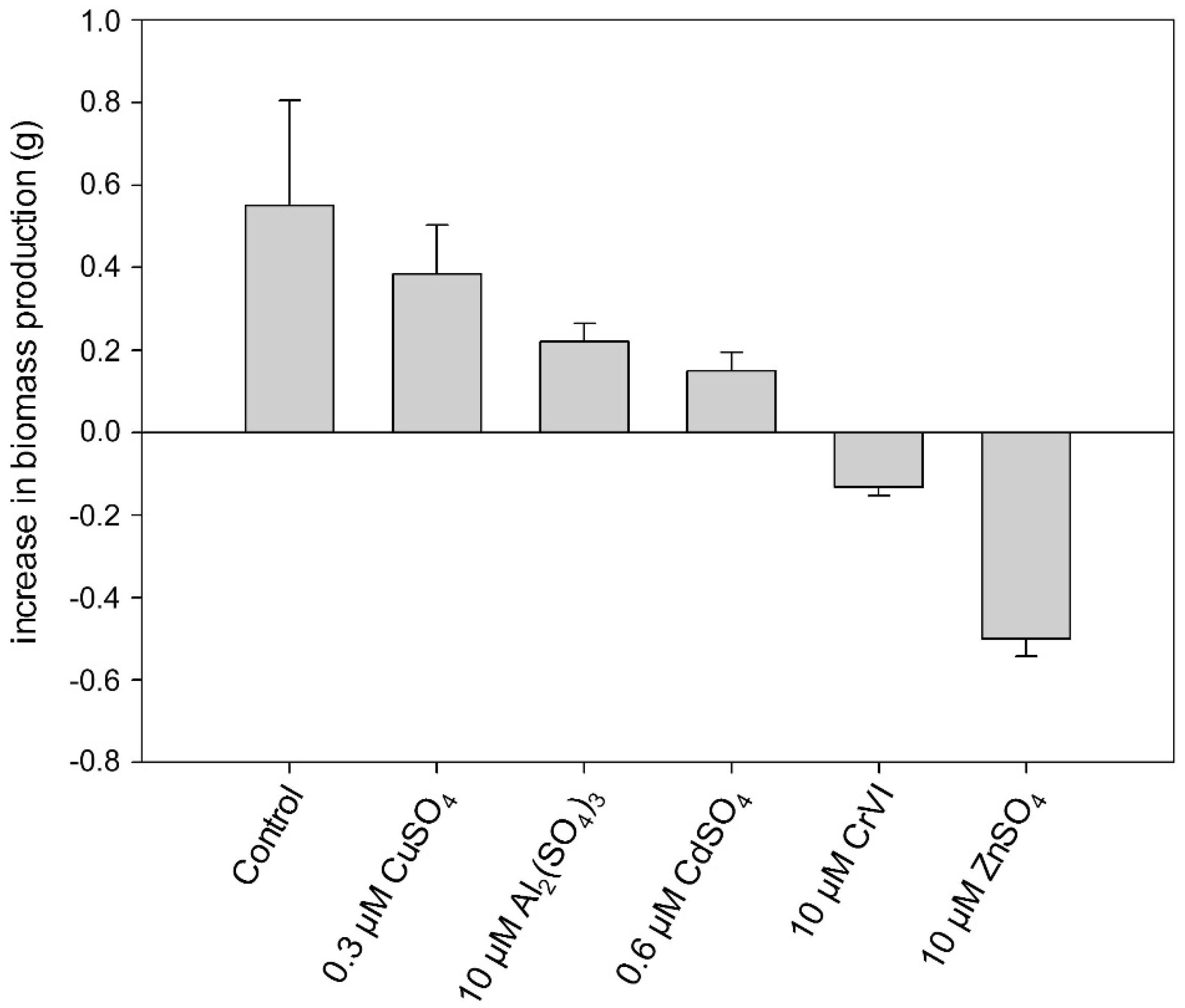
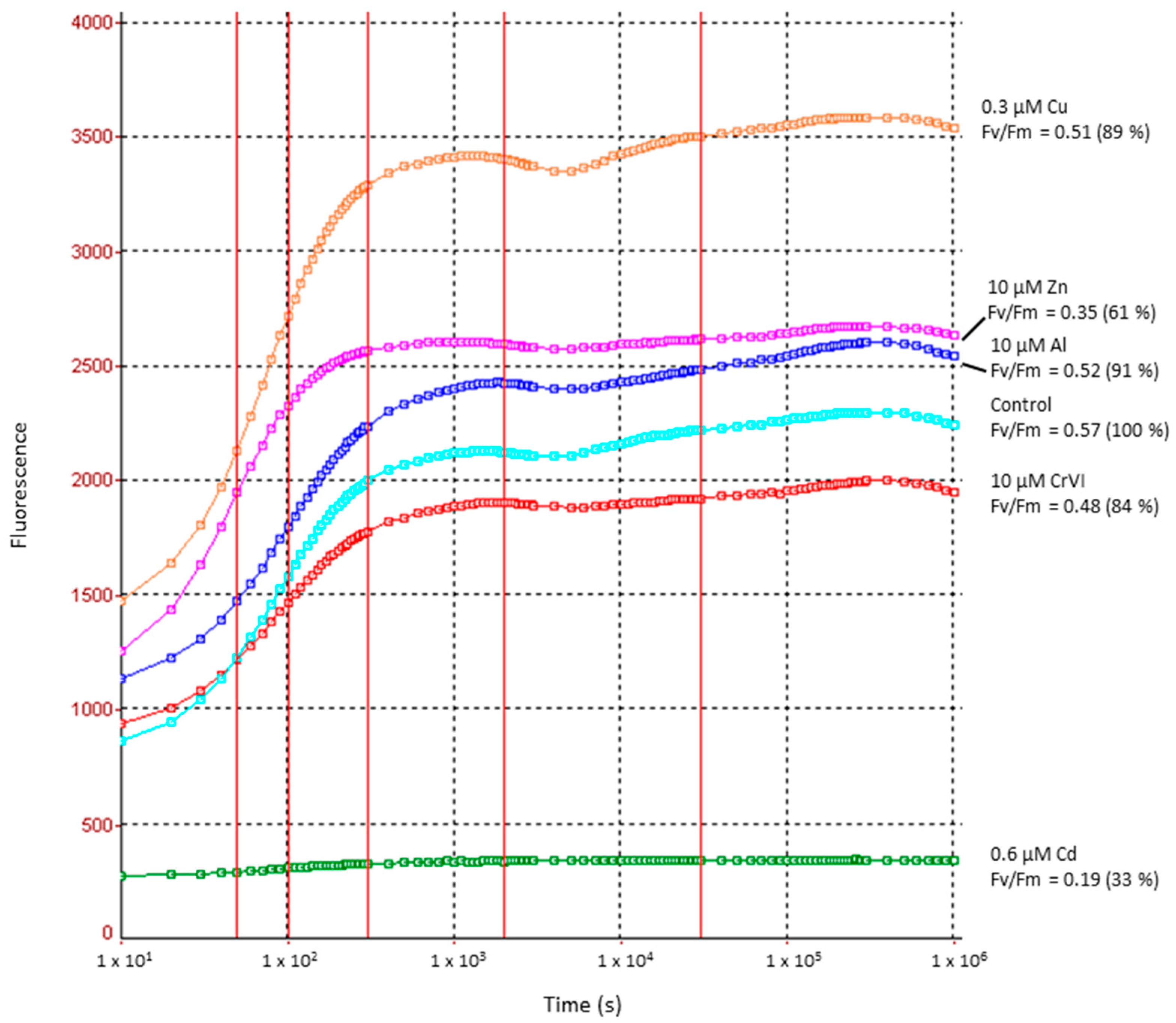
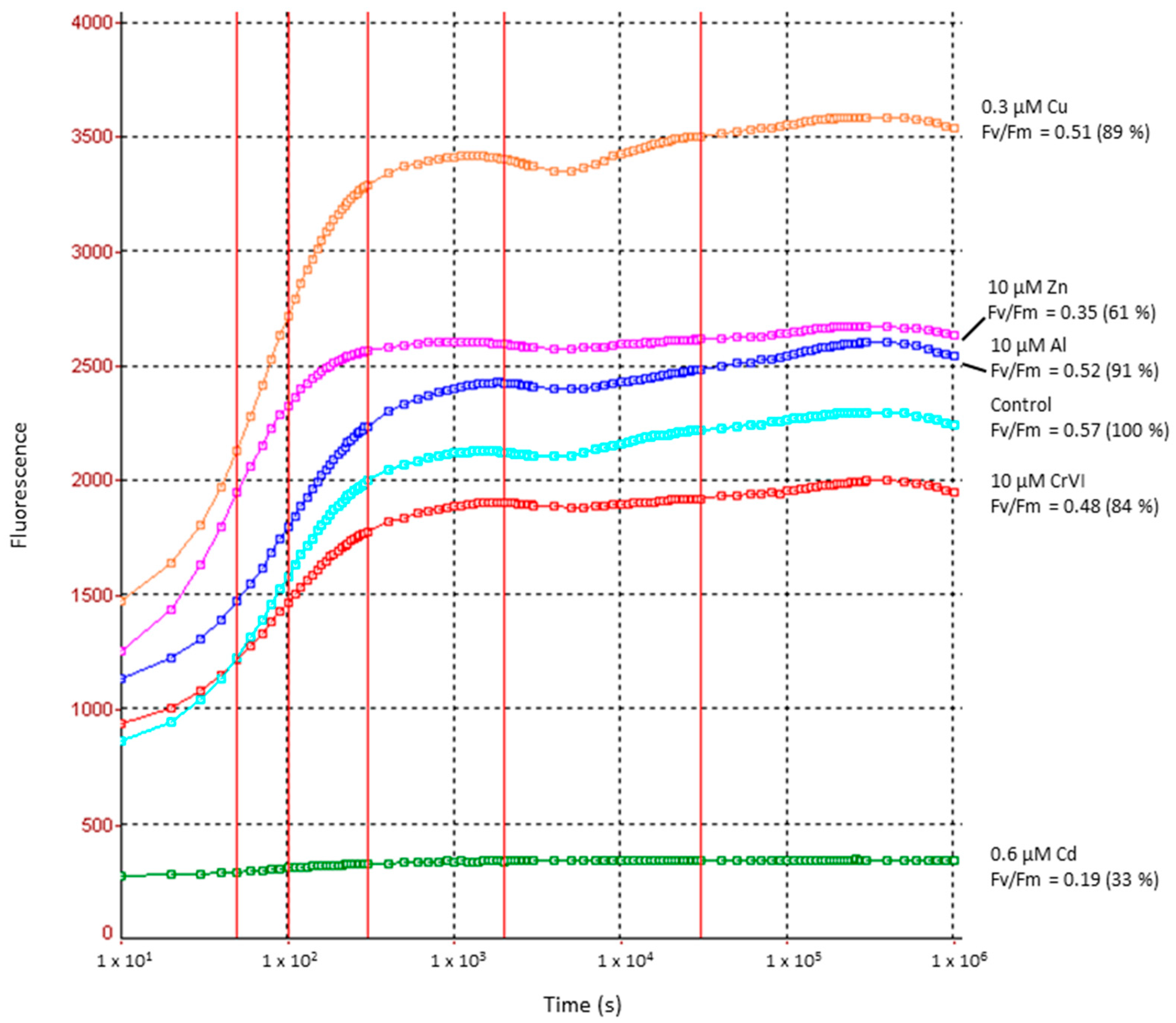
2.3. Heavy Metal Effects on Photosynthetic Electron Transport Efficiency (PSII)
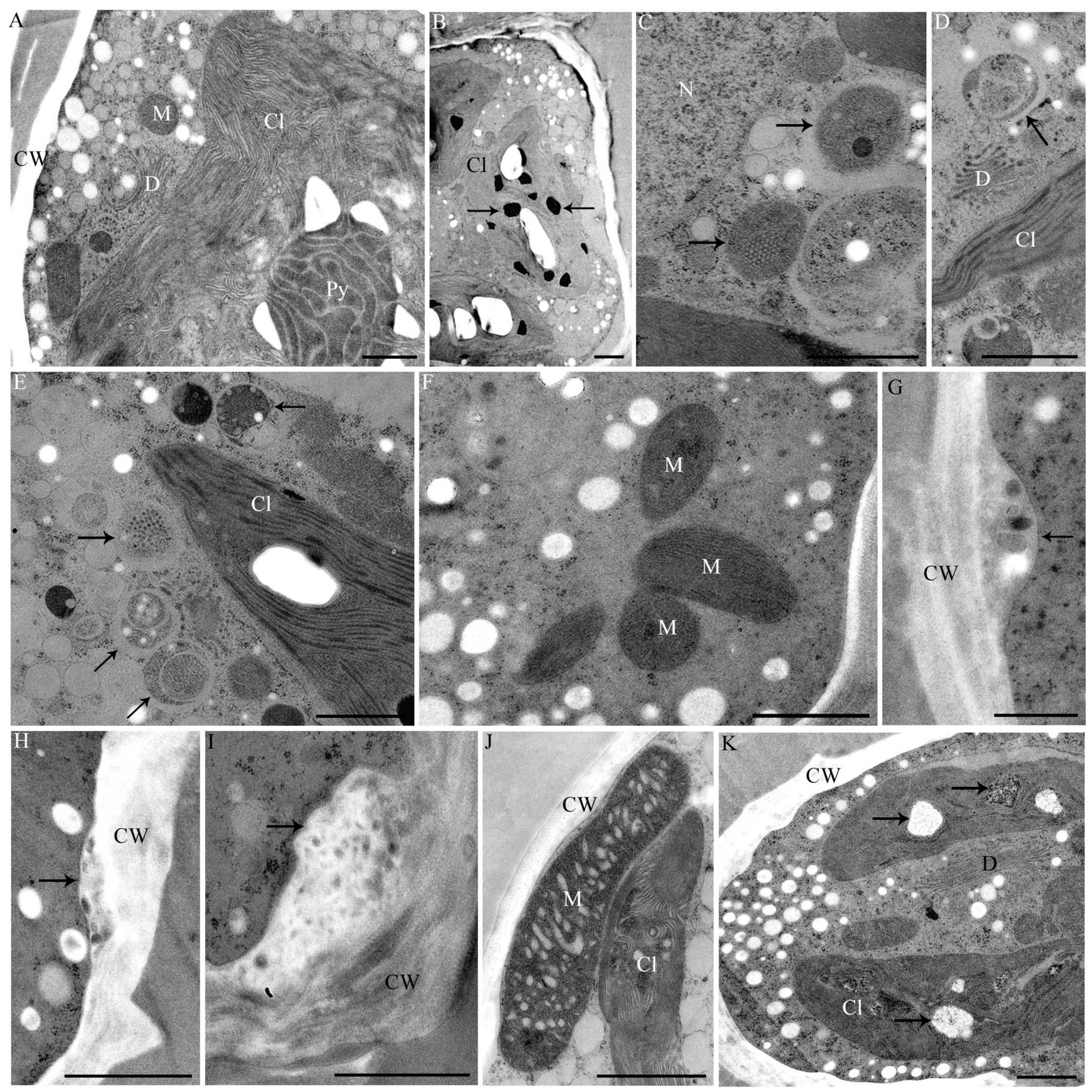
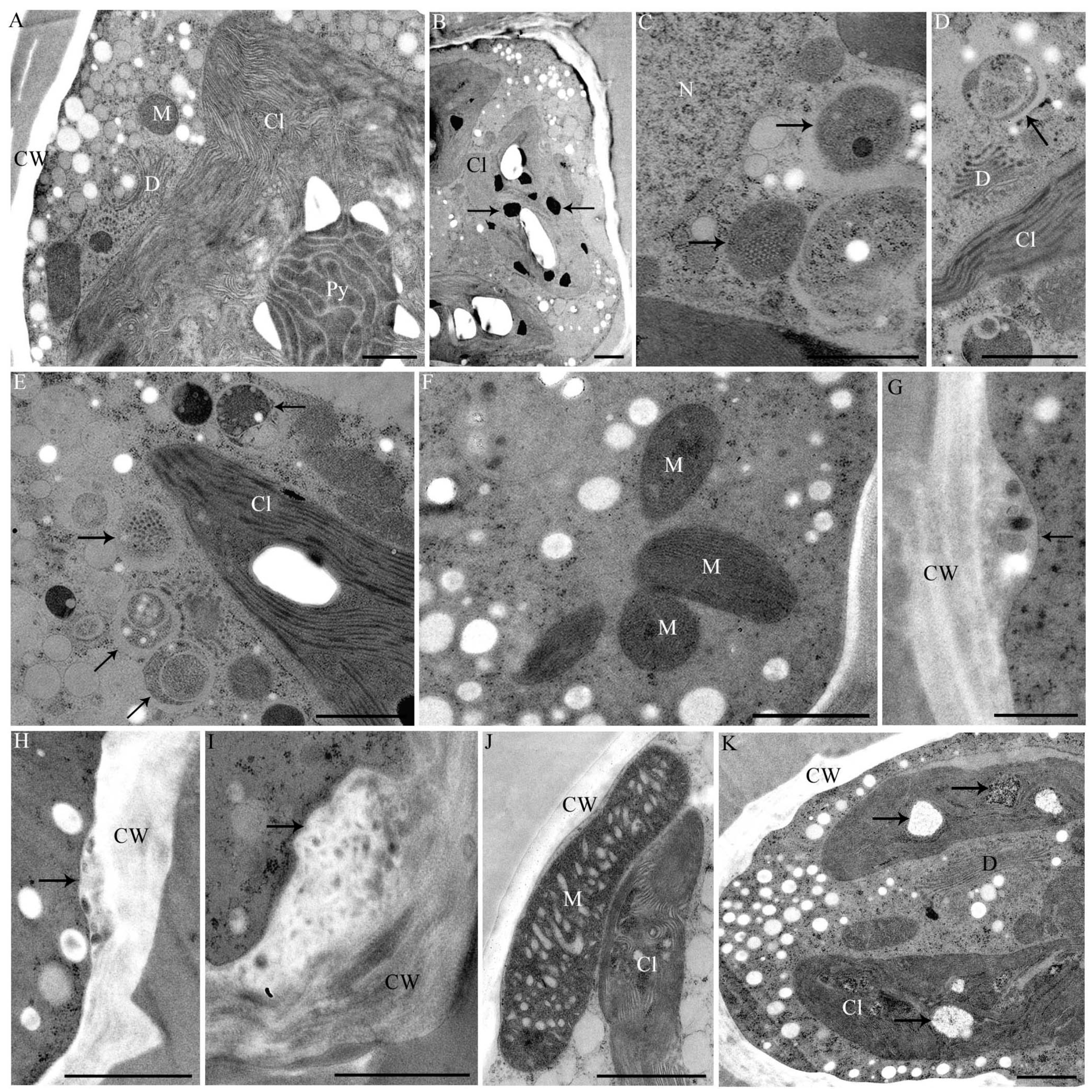
2.4. Changes in Ultrastructure and Organelle Morphology after Heavy Metal Exposure

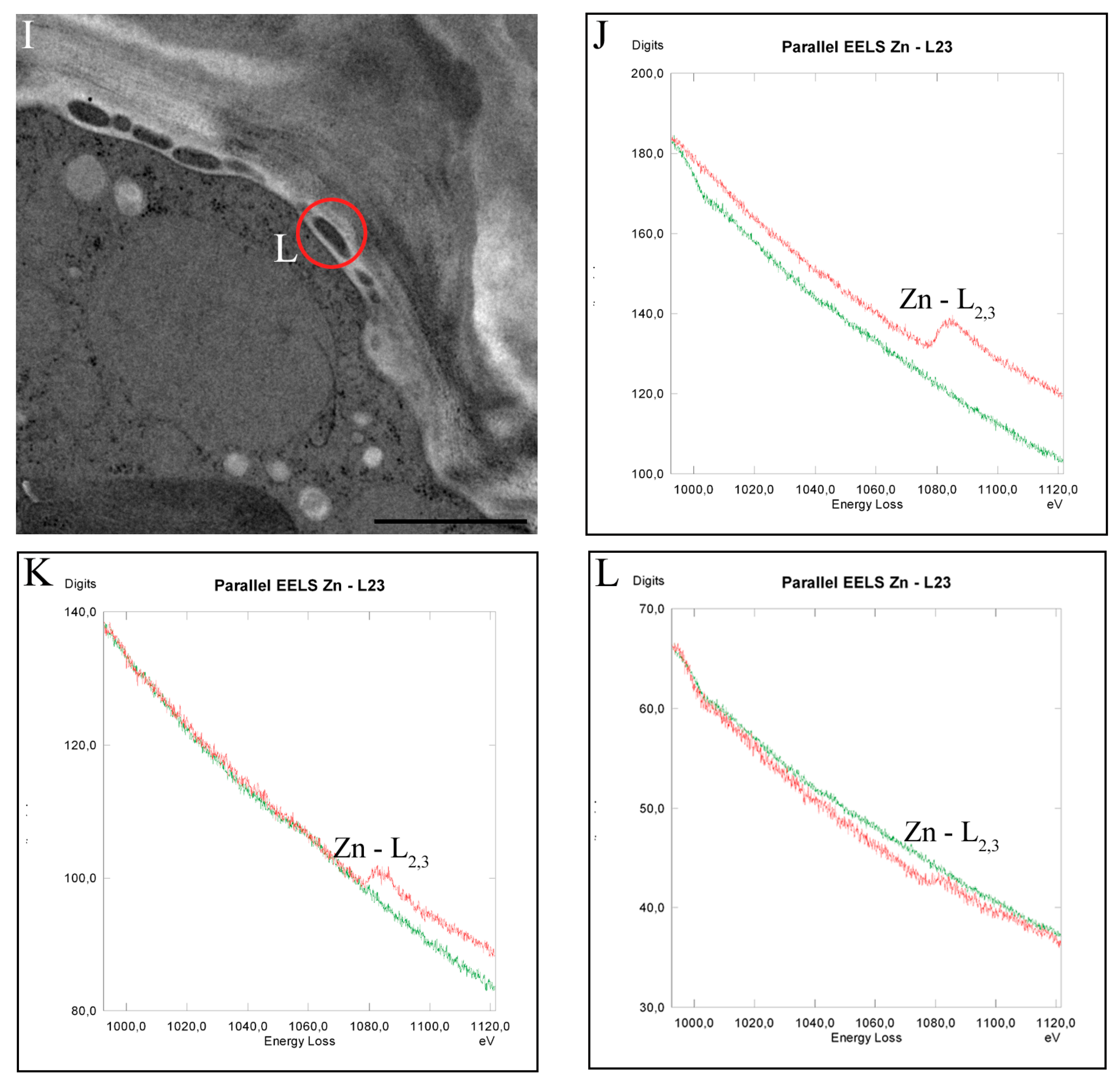
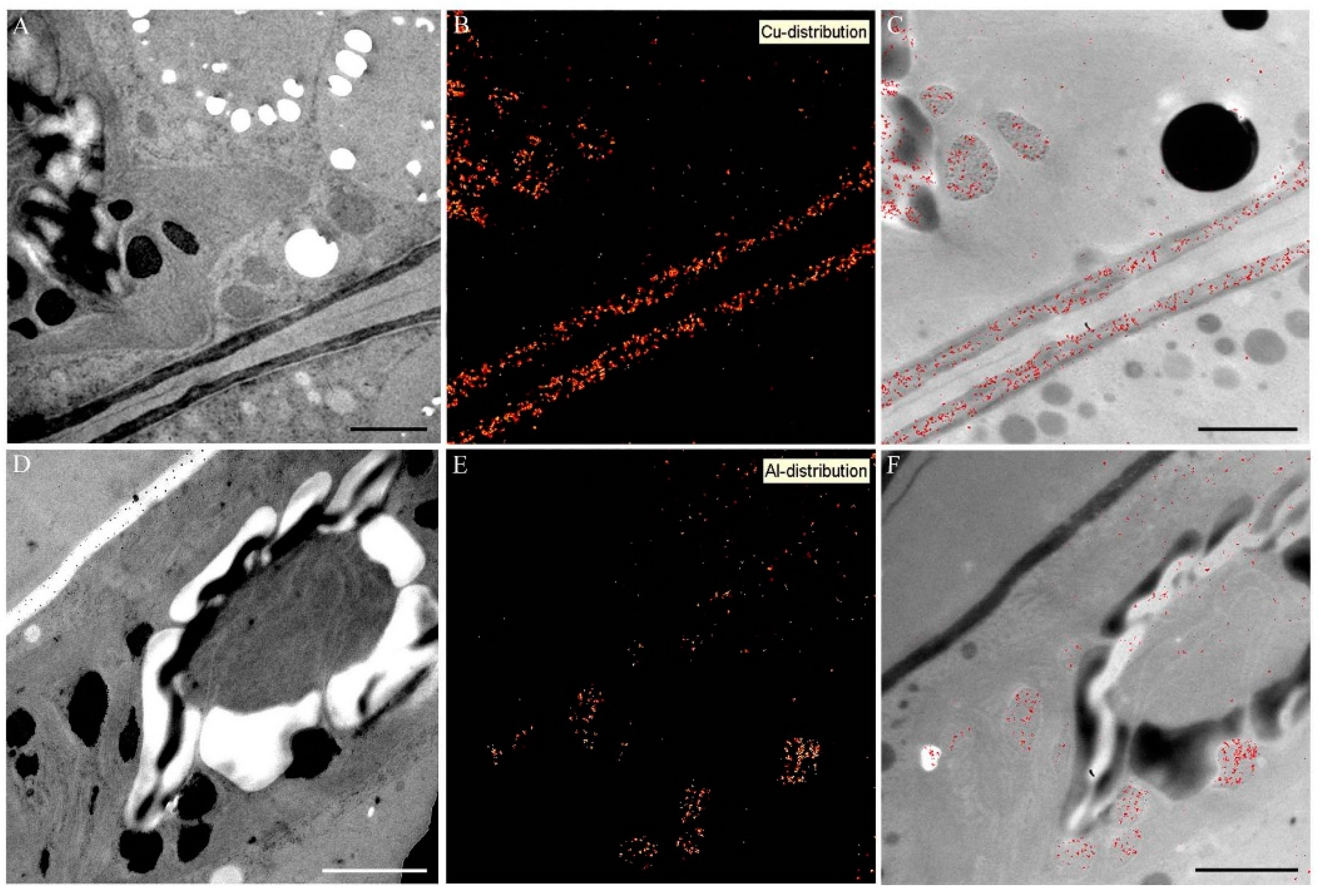
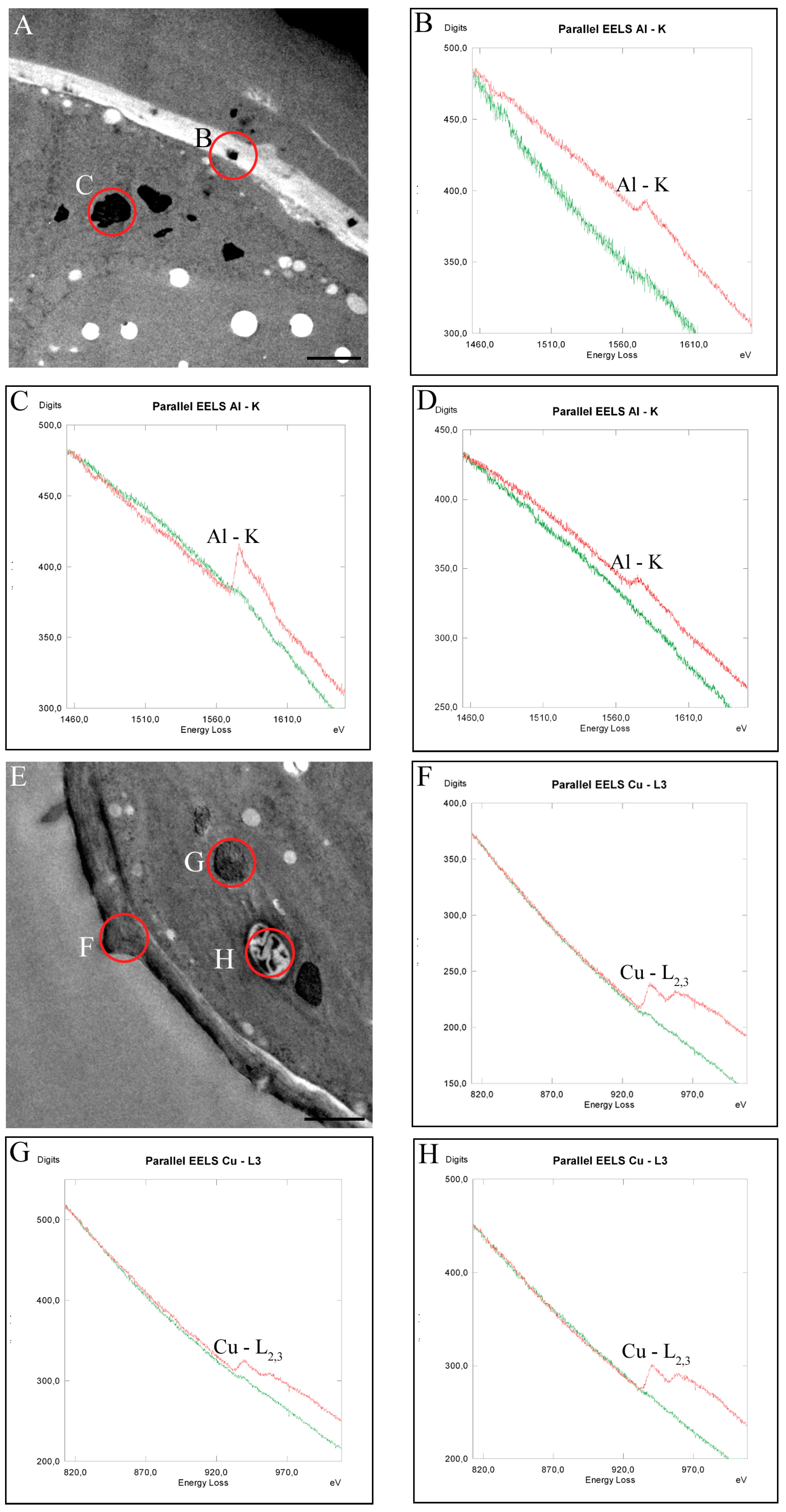
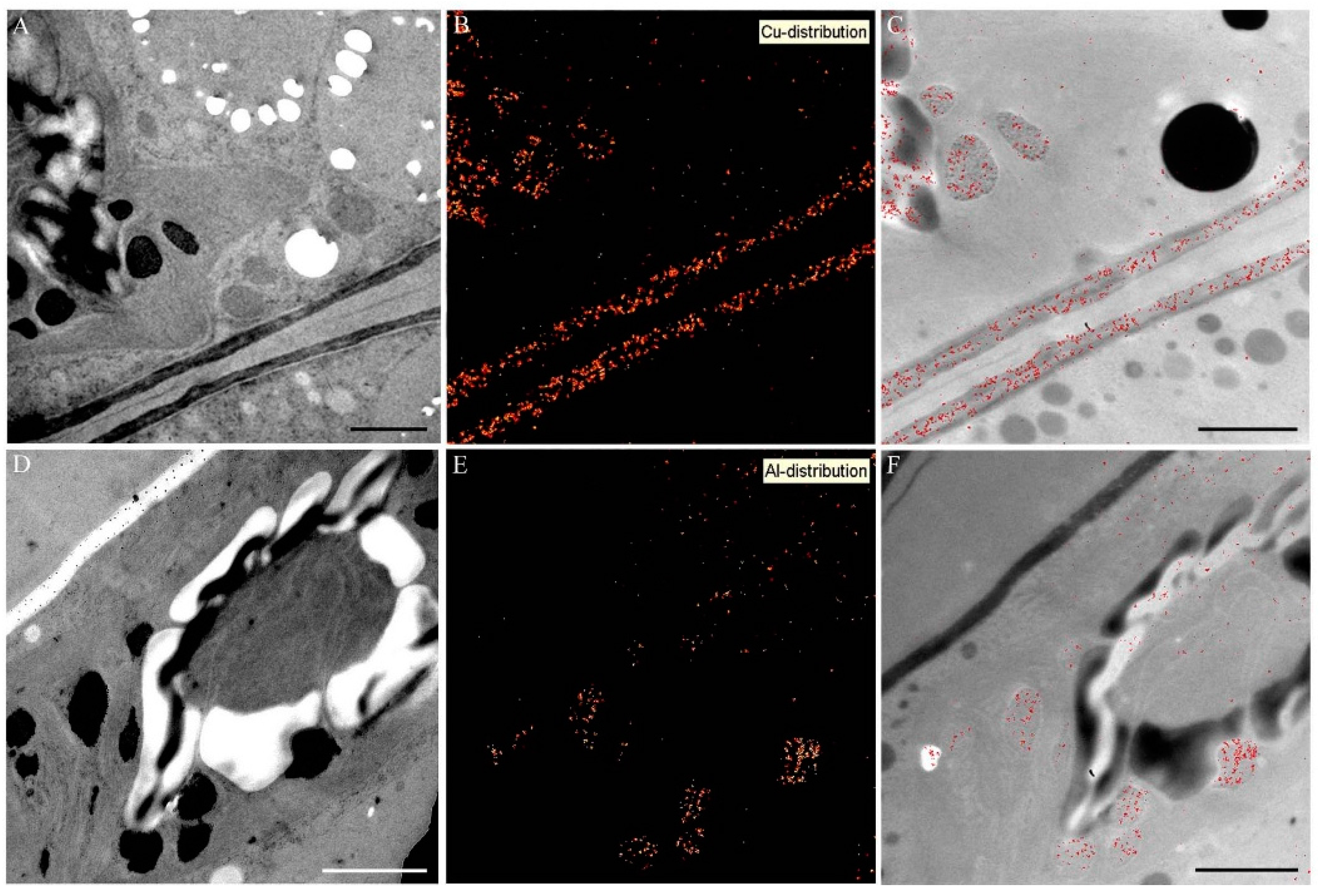
2.5. Intracellular Heavy Metal Localization by Electron Energy Loss Spectroscopy (EELS) and Electron Spectroscopic Imaging (ESI) in TEM
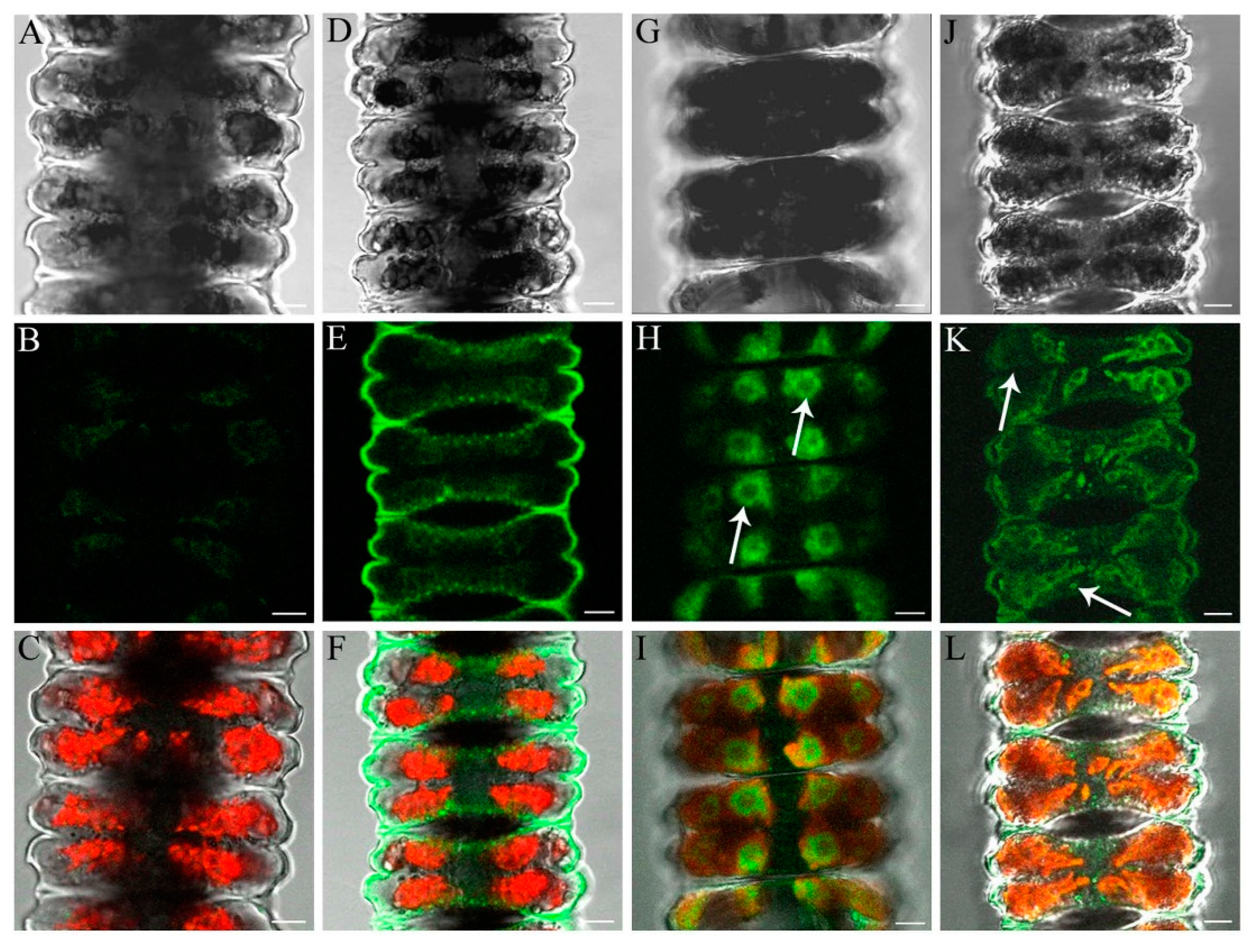
2.6. Determination of Al and Zn Distribution via Morin Fluorescence in CLSM
3. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal | Object | Highest Tolerable Concentration, 21 Days | Cell Division/Biomass Production in %, 21 Days | Ultrastructural Changes | Intracellular Localization Determined by EELS | Photosynthetic Electron Transport (PSII) Fv/Fm in %, 21 Days |
|---|---|---|---|---|---|---|
| control | Desmidium | 0 | 100 | 100 | ||
| Micrasterias | 0 | 100 | 100 | |||
| Al | Desmidium | 10 µM | 40 | dark precipitations in chl and cw | chl, sg, cw | 91 |
| Micrasterias | 20 µM | 28 | d involute and partly disintegrated, increased vacuolization | cw | 97 | |
| Cu | Desmidium | 0.3 µM | 69 | bag-like cwd, dark precipitations and vac in chl | chl, sg, cw | 89 |
| Micrasterias | 0.3 µM | 56 | precipitations in sg and cw, increased vacuolization | sg, cw, mv | 102 | |
| Cd | Desmidium | 0.6 µM | 27 | m with crystalline core autophagosomes | not detectable | 33 |
| Micrasterias | 0.6 µM | 15 | d morphologically severely changed and disintegrated, autophagosomes | not detectable | 26 | |
| CrVI | Desmidium | 10 µM | −23 | bag-like cwd | traces | 84 |
| Micrasterias | 10 µM | 18 | bag-like cwd, increased vacuolization, disturbed chl structure | cw-bags | 41 | |
| Zn | Desmidium | 10 µM | −90 | dark precipitations in chl, bag-like cwd, enlarged and bloated m | chl, cw, cwd | 61 |
| Micrasterias | 30 µM | 0 | dark precipitations in cw, electron dense vac, distrubed chl structure | vac, mv, cwd | 76 |
4. Experimental Section
4.1. Cell Cultures
4.2. Heavy Metal Treatments and Determination of Highest Tolerable Metal Concentration
4.3. Biomass Production
4.4. Light Microscopy
4.5. Determination of Photosynthetic Electron Transport Efficiency (PSII)
4.6. TEM Analysis, EELS and ESI Measurements and Immuno TEM
4.7. Detection of Al and Zn by Morin Fluorescence in CLSM
4.8. Comparison of Data between Desmidium and Micrasterias
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Alloway, B.J.; Ayres, D.C. Schadstoffe in der Umwelt: Chemische Grundlagen zur Beurteilung von Luft-, Wasser-und Bodenverschmutzungen. Spektrum; Akademischer Verlag: Heidelberg, Germany, 1996; pp. 23–83. [Google Scholar]
- Kovacik, J.; Klejdus, B.; Hedbavny, J.; Backor, M. Effect of copper and salicylic acid on phenolic metabolites and free amino acids in Scenedesmus quadricauda (Chlorophyceae). Plant Sci. 2010, 178, 307–311. [Google Scholar] [CrossRef]
- Andosch, A.; Affenzeller, M.J.; Lütz, C.; Lütz-Meindl, U. A freshwater green alga under cadmium stress: Ameliorating calcium effects on ultrastructure and photosynthesis in the unicellular model Micrasterias. J. Plant Physiol. 2012, 169, 1489–1500. [Google Scholar] [CrossRef] [PubMed]
- Volland, S.; Lütz, C.; Michalke, B.; Lütz-Meindl, U. Intracellular chromium localization and cell physiological response in the unicellular alga Micrasterias. Aquat. Toxicol. 2012, 109, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Volland, S.; Bayer, E.; Baumgartner, V.; Andosch, A.; Lütz, C.; Sima, E.; Lütz-Meindl, U. Rescue of heavy metal effects on cell physiology of the algal model system Micrasterias by divalent ions. J. Plant Physiol. 2014, 171, 154–163. [Google Scholar] [PubMed]
- Carfagna, S.; Lanza, N.; Salbitani, G.; Basile, A.; Sorbo, S.; Vona, V. Physiological and morphological responses of lead or cadmium exposed Chlorella sorokiniana 211–8K (Chlorophyceae). SpringerPlus 2013, 2, 147. [Google Scholar] [CrossRef] [PubMed]
- Di Toppi, L.S.; Gabbrielli, R. Response to cadmium in higher plants. Environ. Exp. Bot. 1999, 41, 105–130. [Google Scholar]
- Pinto, E.; Sigaud-Kutner, T.C.S.; Leitao, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy metal-induced oxidative stress in algae. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Shanker, A.K.; Cervantes, C.; Loza-Tavera, H.; Avudainayagam, S. Chromium toxicity in plants. Environ. Int. 2005, 31, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K.J. The relationship between metal toxicity and cellular redox imbalance. Trends Plant Sci. 2009, 14, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Kottke, I. Subcellular localization of chromium and nickel in root cells of Allium cepa by EELS and ESI. Cell Biol. Toxicol. 2003, 19, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Volland, S.; Andosch, A.; Milla, M.; Stöger, B.; Lütz, C.; Lütz-Meindl, U. Intracellular metal compartmentalization in the green algal model system Micrasterias denticulata (Streptophyta) measured by transmission electron microscopy-coupled electron energy loss spectroscopy. J. Phycol. 2011, 47, 565–579. [Google Scholar] [CrossRef]
- Volland, S.; Schaumloffel, D.; Dobritzsch, D.; Krauss, G.J.; Lütz-Meindl, U. Identification of phytochelatins in the cadmium-stressed conjugating green alga Micrasterias denticulata. Chemosphere 2013, 91, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.D.; McCourt, R.M.; Delwiche, C.F. Patterns of cell division in the filamentous Desmidiaceae, close green algal relatives of land plants. Am. J. Bot. 2008, 95, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Url, W. Resistenz von Desmidiaceen gegen Schwermetallsalze. Sitzungsber. Österr. Akad. Wiss. Mathem. Naturw. 1955, 164, 207–230. [Google Scholar]
- Taylor, G.J.; McDonald-Stephens, J.L.; Hunter, D.B.; Bertsch, P.M.; Elmore, D.; Rengel, Z.; Reid, R.J. Direct measurement of aluminum uptake and distribution in single cells of Chara corallina. Plant Physiol. 2000, 123, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, R.; del Hoyo, A.; García-Breijo, F.; Reig-Armiñana, J.; del Campo, E.M.; Guéra, A.; Barreno, E.; Casano, L.M. Different strategies to achieve Pb-tolerance by the two Trebouxia algae coexisting in the lichen Ramalina farinacea. J. Plant Physiol. 2012, 169, 1797–1806. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jiang, Y.; Chen, C.-S.; Creeley, D.; Schwehr, K.A.; Quigg, A.; Chin, W.-C.; Santschi, P.H. Ameliorating effects of extracellular polymeric substances excreted by Thalassiosira pseudonana on algal toxicity of CdSe quantum dots. Aquat. Toxicol. 2013, 126, 214–223. [Google Scholar] [CrossRef] [PubMed]
- García-Ríos, V.; Freile-Pelegrín, Y.; Robledo, D.; Mendoza-Cózatl, D.; Moreno-Sánchez, R.; Gold-Bouchot, G. Cell wall composition affects Cd2+ accumulation and intracellular thiol peptides in marine red algae. Aquat. Toxicol. 2007, 81, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Eder, M.; Tenhaken, R.; Driouich, A.; Lütz-Meindl, U. Occurrence and characterization of arabinogalactan-like proteins and hemicelluloses in Micrasterias (Streptophyta). J. Phycol. 2008, 44, 1221–1234. [Google Scholar] [CrossRef]
- Andrade, L.R.; Leal, R.N.; Noseda, M.; Duarte, M.E.R.; Pereira, M.S.; Mourão, P.A.S.; Farina, M.; Amado Filho, G.M. Brown algae overproduce cell wall polysaccharides as a protection mechanism against the heavy metal toxicity. Mar. Pollut. Bull. 2010, 60, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Salgado, L.T.; Andrade, L.R.; Filho, G.M. A. Localization of specific monosaccharides in cells of the brown alga Padina gymnospora and the relation to heavy-metal accumulation. Protoplasma 2005, 225, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Gerrath, J.F. Notes on desmid ultrastructure. Nova Hedwig. 1973, 42, 103–113. [Google Scholar]
- Krupp, J.M.; Lang, N.J. Cell division and filament formation in the desmid Bambusina brebissonii (Chlorophyta). J. Phycol. 1985, 21, 16–25. [Google Scholar] [CrossRef]
- Krupp, J.M.; Lang, N.J. Cell division and the role of the primary wall in the filamentous desmid Onychonema laeve (Chlorophyta). J. Phycol. 1985, 21, 316–322. [Google Scholar] [CrossRef]
- Eder, M.; Lütz-Meindl, U. Pectin-like carbohydrates in the green alga Micrasterias characterized by cytochemical analysis and energy filtering TEM. J. Microsc. 2008, 231, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Schlösser, U.G. Sammlungen von Algenkulturen. Ber. Deutsch. Bot. Ges. 1982, 95, 181–276. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Govindjee. Polyphasic chlorophyll-α fluorescence transient in plants and Cyanobacteria. Photochem. Photobiol. 1995, 61, 32–42. [Google Scholar]
- Lütz, C.; Seidlitz, H.K.; Meindl, U. Physiological and structural changes in the chloroplast of the green alga Micrasterias denticulata induced by UV-B simulation. Plant Ecol. 1997, 128, 54–64. [Google Scholar] [CrossRef]
- Affenzeller, M.J.; Darehshouri, A.; Andosch, A.; Lütz, C.; Lütz-Meindl, U. Salt stress-induced cell death in the unicellular green alga Micrasterias denticulata. J. Exp. Bot. 2009, 60, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Meindl, U.; Lancelle, S.; Hepler, P. Vesicle production and fusion during lobe formation in Micrasterias visualized by high-pressure freeze fixation. Protoplasma 1992, 170, 104–114. [Google Scholar] [CrossRef]
- Lütz-Meindl, U.; Aichinger, N. Use of energy-filtering transmission electron microscopy for routine ultrastrutural analysis of high-pressure-frozen or chemically fixed plant cells. Protoplasma 2004, 223, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Lütz-Meindl, U.; Lütz, C. Analysis of element accumulation in cell wall attached and intracellular particles of snow algae by EELS and ESI. Micron 2006, 37, 452–458. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andosch, A.; Höftberger, M.; Lütz, C.; Lütz-Meindl, U. Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii. Int. J. Mol. Sci. 2015, 16, 10389-10410. https://doi.org/10.3390/ijms160510389
Andosch A, Höftberger M, Lütz C, Lütz-Meindl U. Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii. International Journal of Molecular Sciences. 2015; 16(5):10389-10410. https://doi.org/10.3390/ijms160510389
Chicago/Turabian StyleAndosch, Ancuela, Margit Höftberger, Cornelius Lütz, and Ursula Lütz-Meindl. 2015. "Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii" International Journal of Molecular Sciences 16, no. 5: 10389-10410. https://doi.org/10.3390/ijms160510389
APA StyleAndosch, A., Höftberger, M., Lütz, C., & Lütz-Meindl, U. (2015). Subcellular Sequestration and Impact of Heavy Metals on the Ultrastructure and Physiology of the Multicellular Freshwater Alga Desmidium swartzii. International Journal of Molecular Sciences, 16(5), 10389-10410. https://doi.org/10.3390/ijms160510389