Effects of Nanotoxicity on Female Reproductivity and Fetal Development in Animal Models


Abstract
:1. Introduction
2. Effects of Nanotoxicity on Female Reproductivity and Development in Murine Models
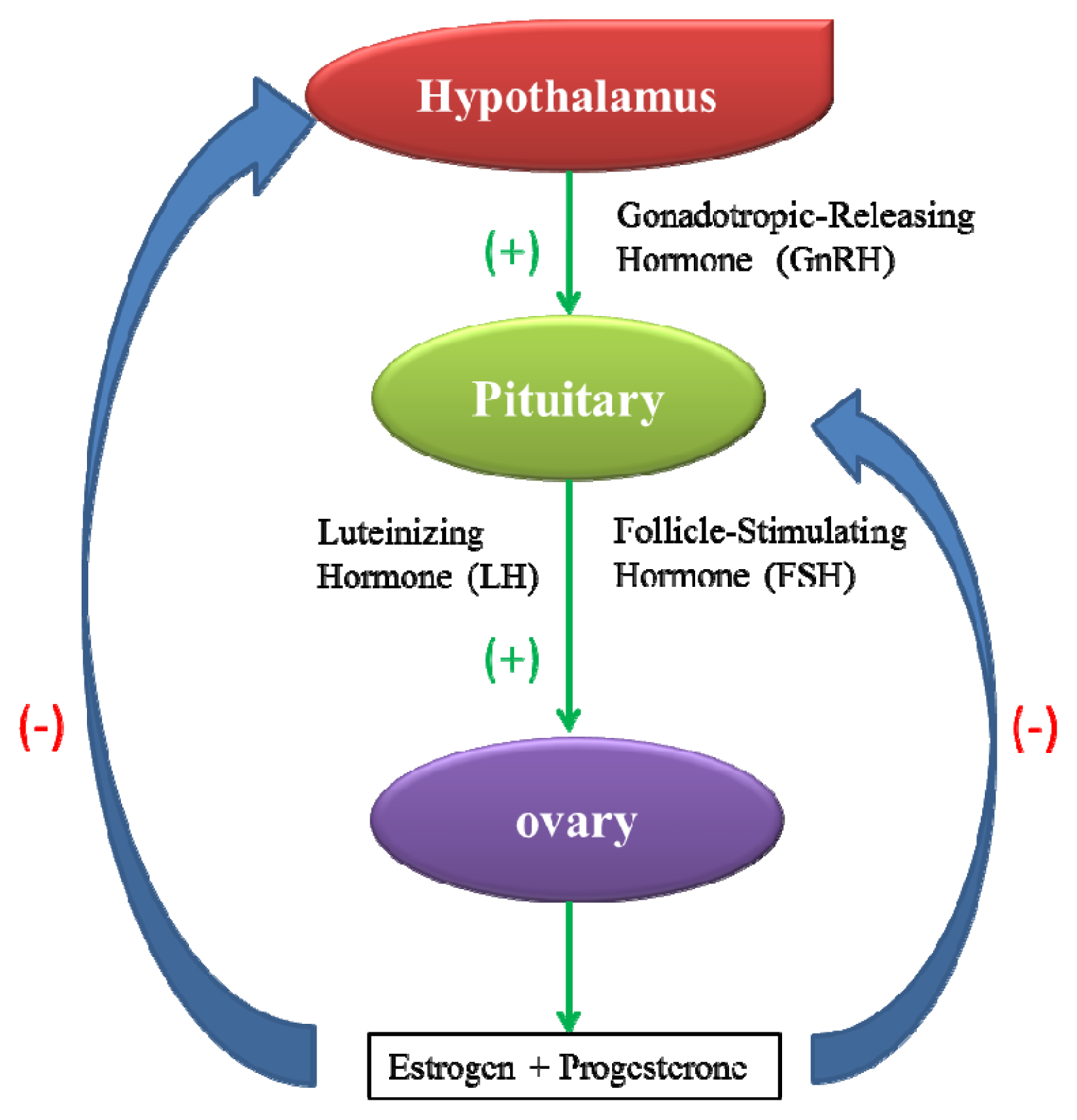
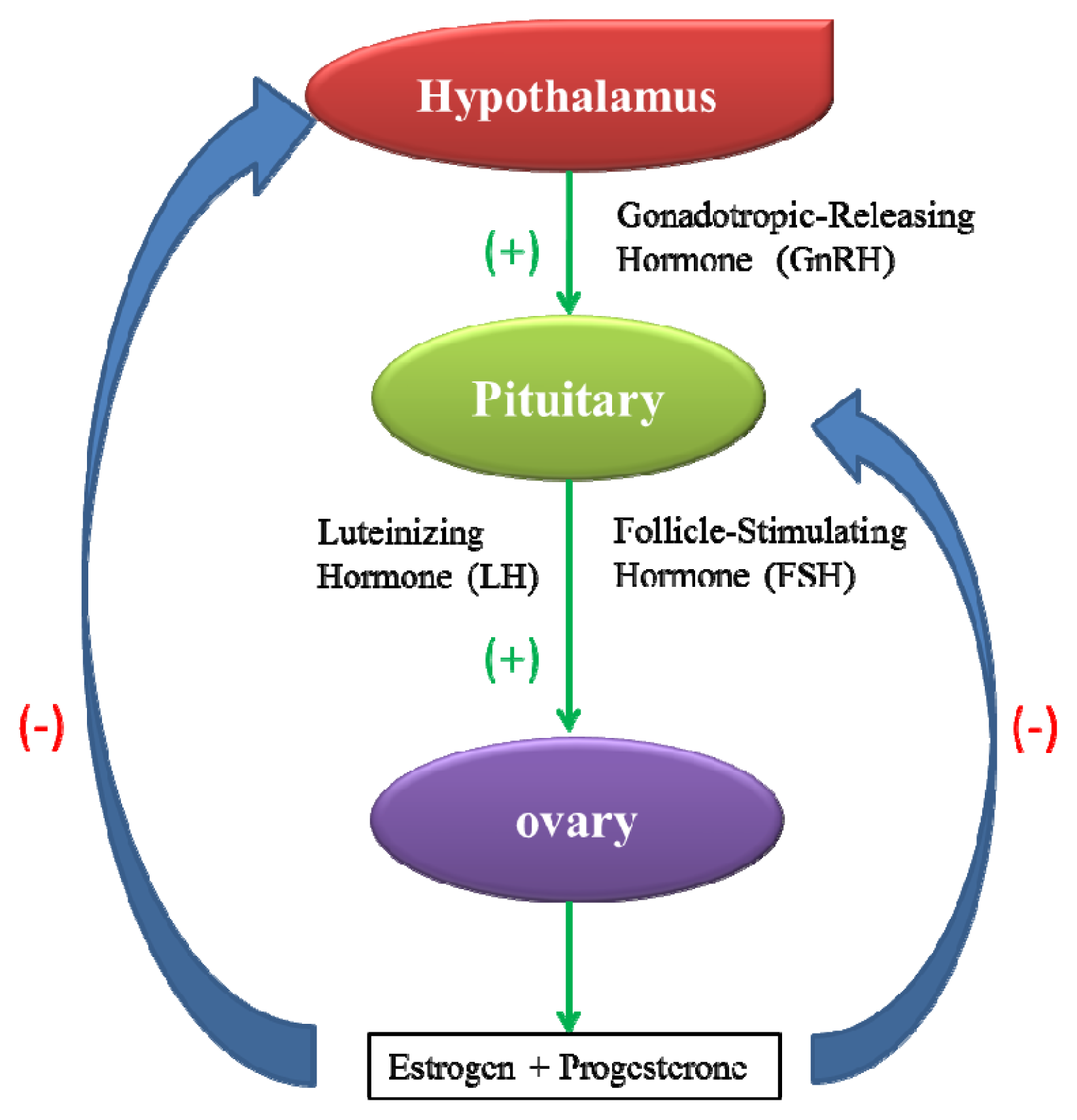
2.1. Toxicity to the Female Reproductive System
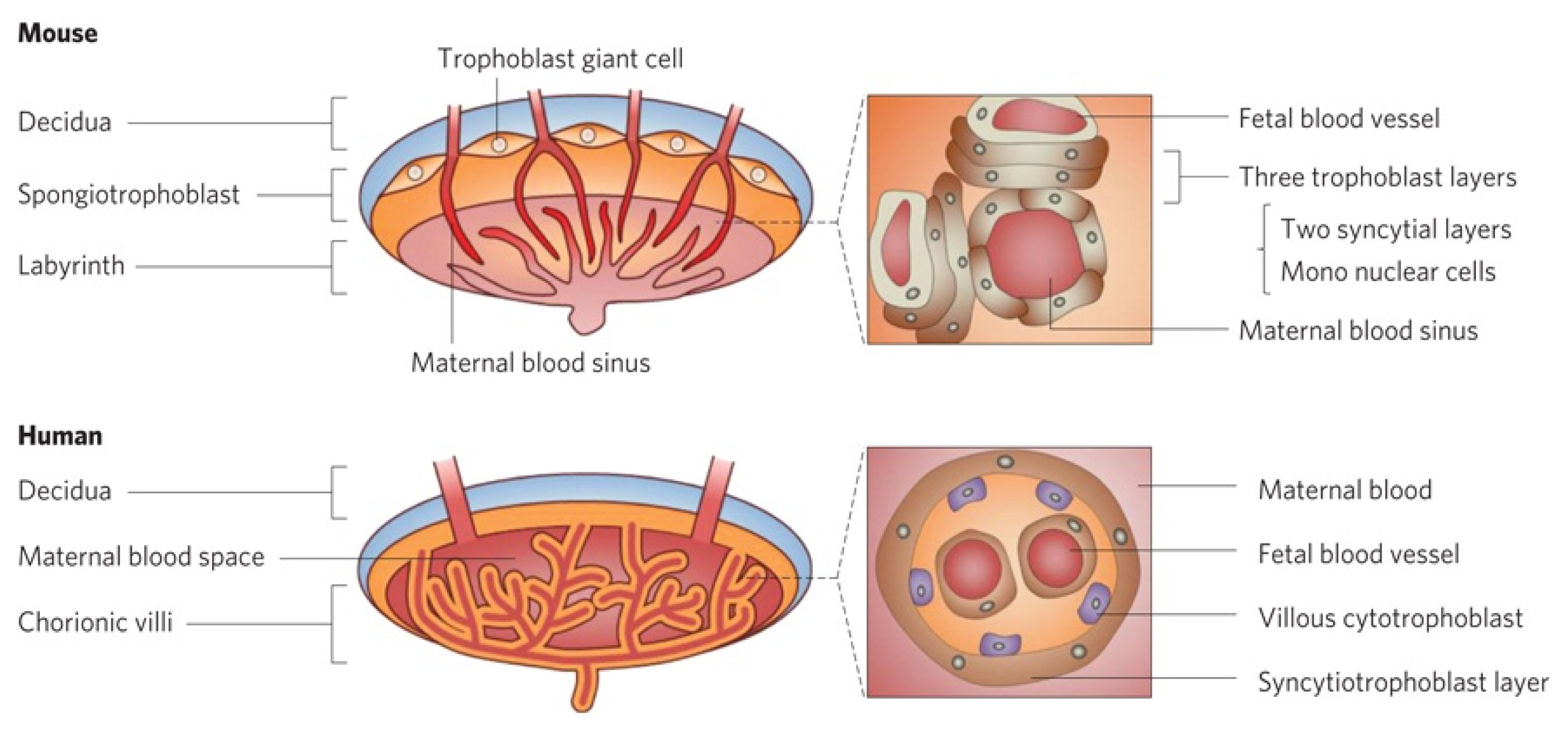
2.2. Transplacental Ability of Nanoparticles
2.3. Effects of Nanoparticles on Fetal Developmental
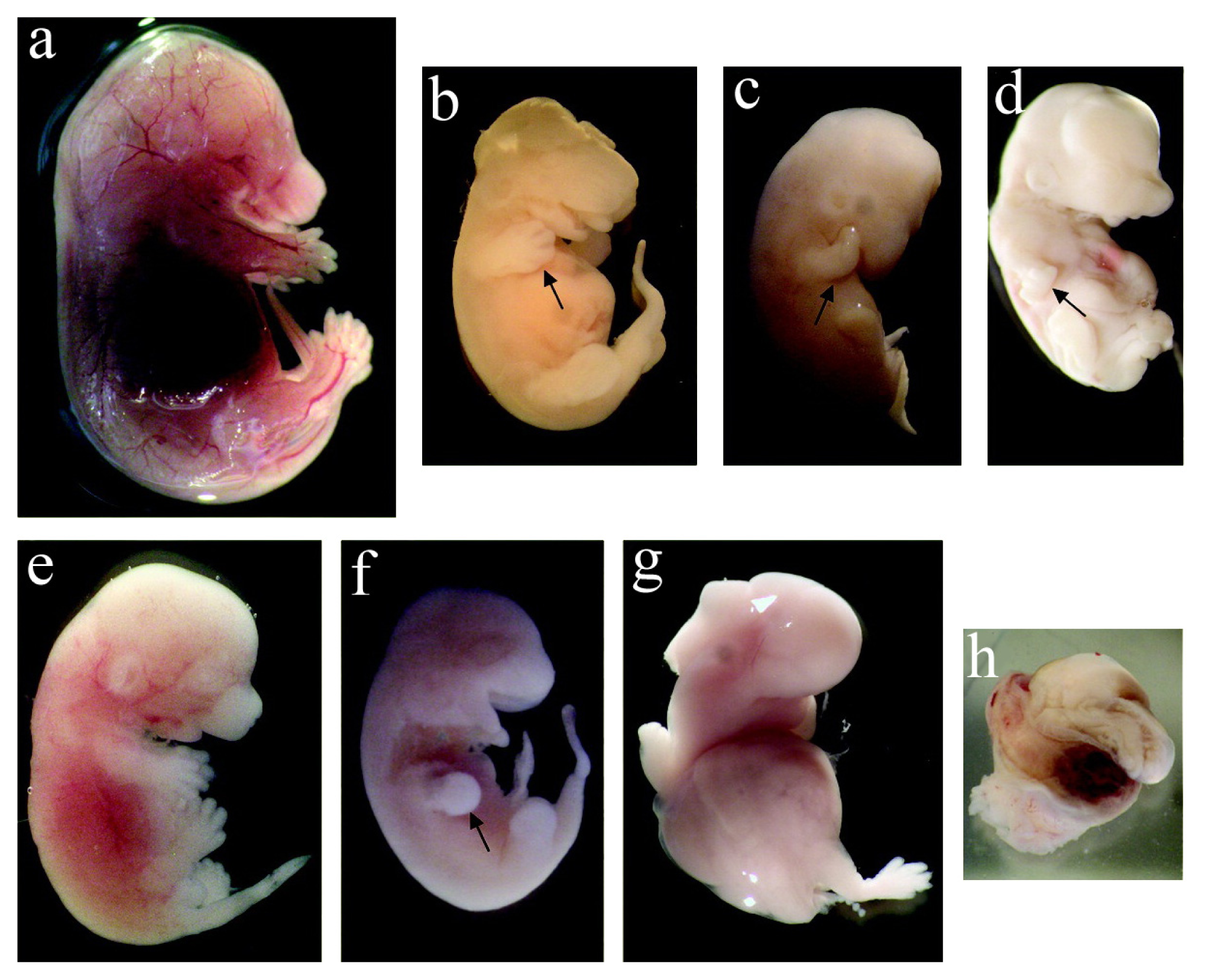
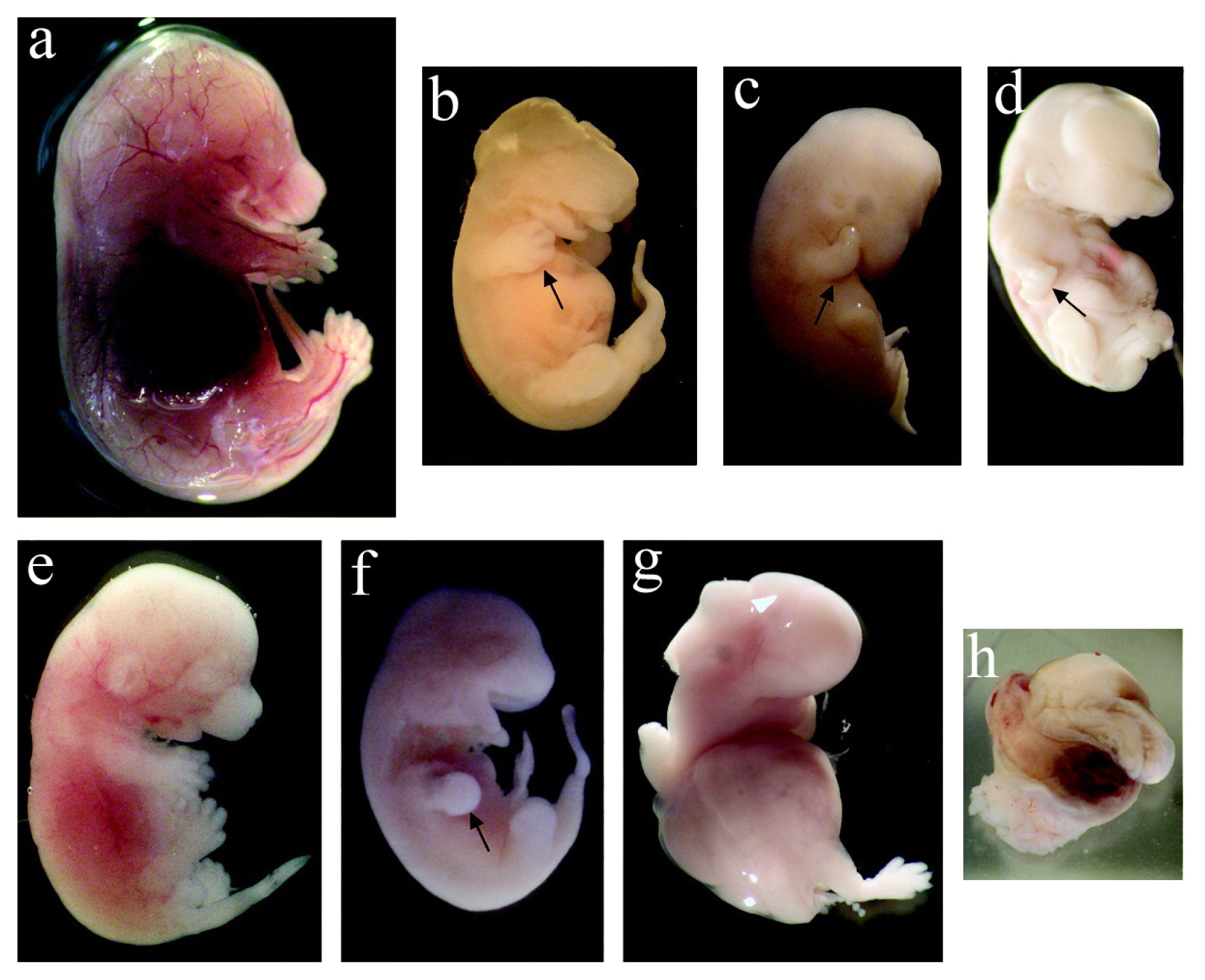
2.3.1. Toxicity on Fetal Organogenesis and Morphology
2.3.2. Toxicity to the Fetal Reproductive Function
2.3.3. Effects on Fetal Neurodevelopment
2.3.4. Mechanistic Studies
2.4. In Vitro Models of Murine Origin
3. Nanotoxicity to Female Reproductivity and Embryonic Development in Zebrafish
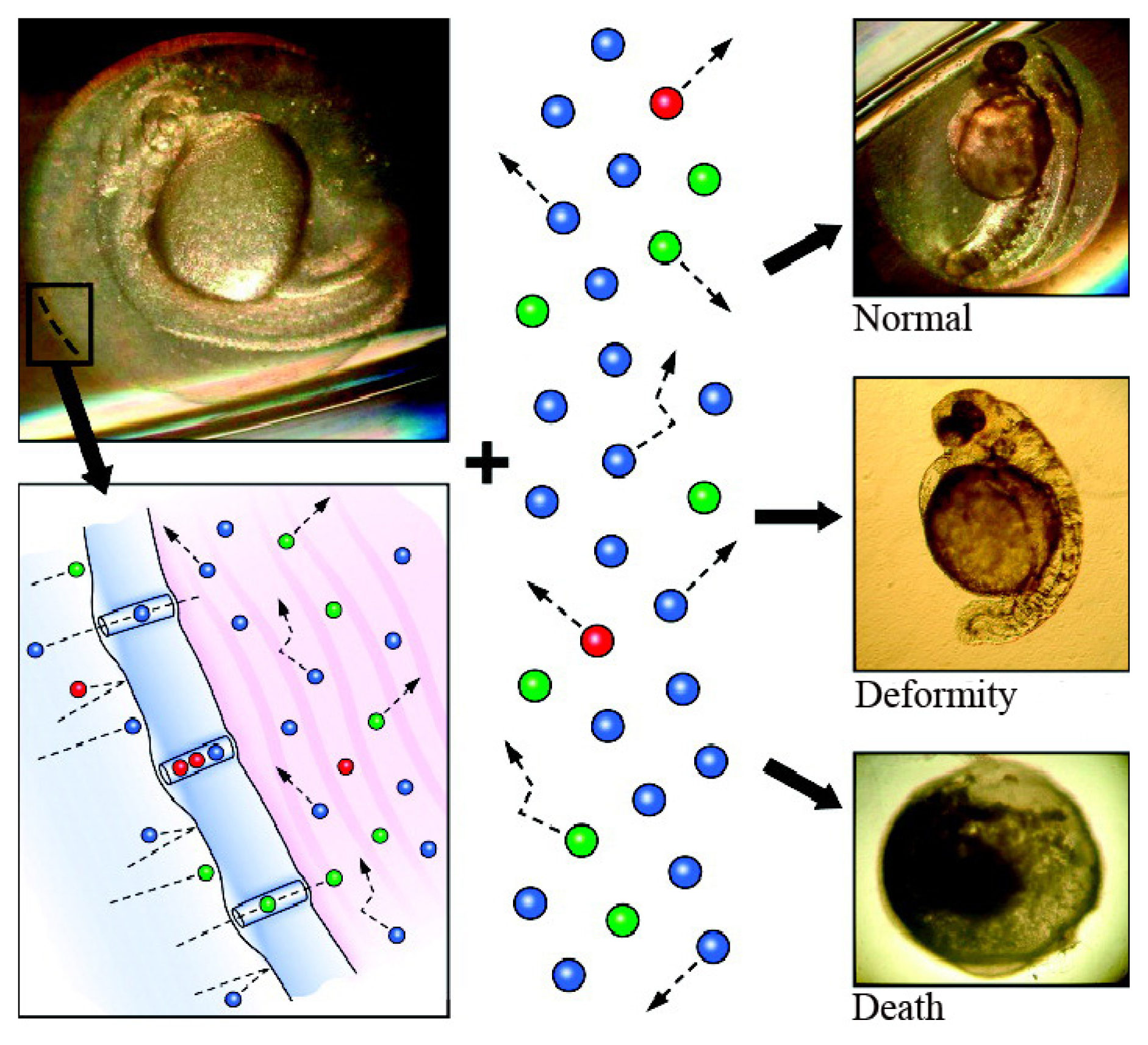
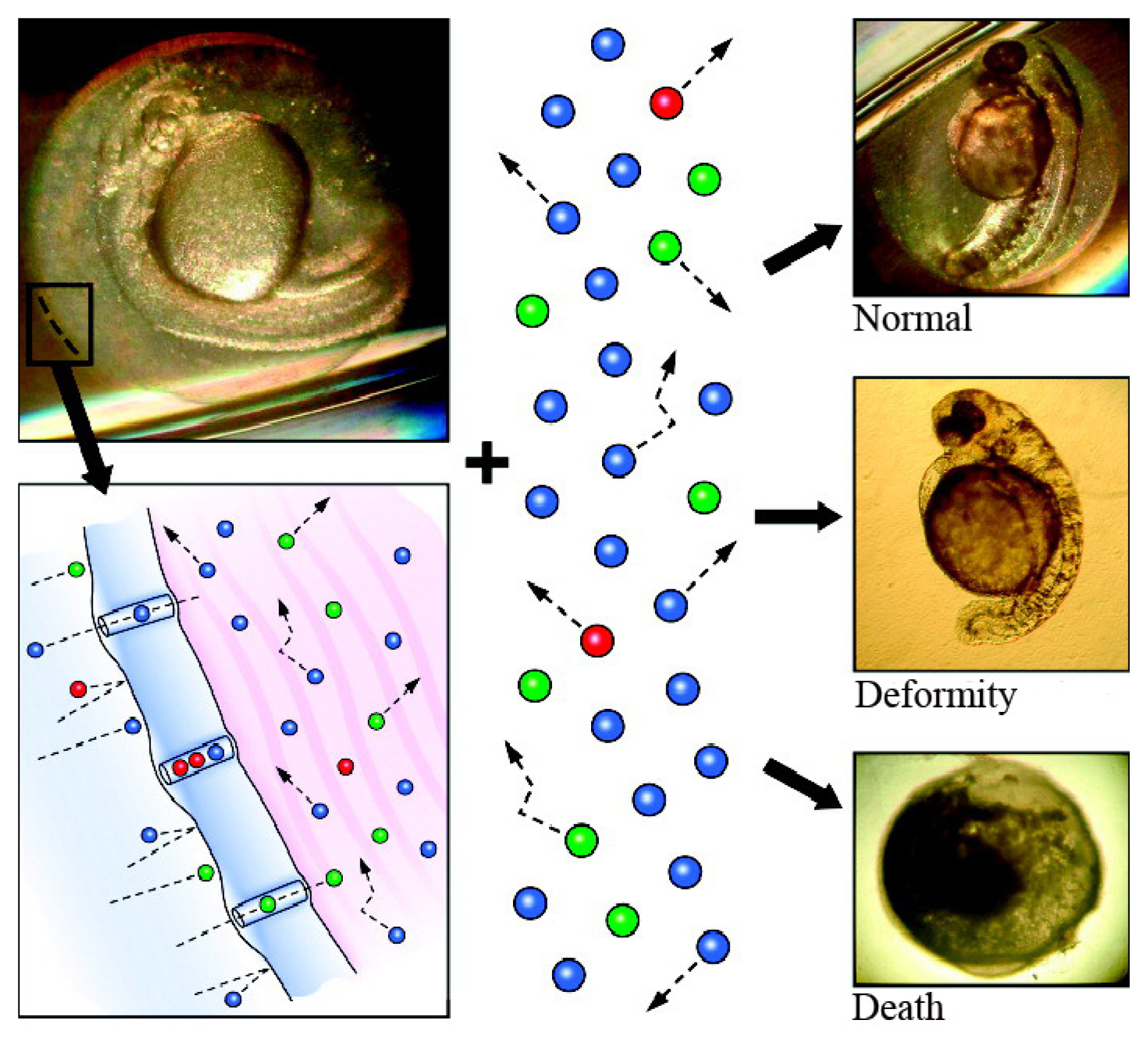
3.1. Translocation of Nanoparticles to the Zebrafish Embryo
3.2. Toxicity of Nanoparticles in Zebrafish Embryos
3.2.1. Metal Nanoparticles
3.2.2. Carbon Nanomaterials
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Zhang, L.; Gu, F.; Chan, J.; Wang, A.; Langer, R.; Farokhzad, O. Nanoparticles in medicine: Therapeutic applications and developments. Clin. Pharmacol. Ther 2007, 83, 761–769. [Google Scholar]
- Lee, J.H.; Huh, Y.M.; Jun, Y.; Seo, J.; Jang, J.; Song, H.T.; Kim, S.; Cho, E.J.; Yoon, H.G.; Suh, J.S. Artificially engineered magnetic nanoparticles for ultra-sensitive molecular imaging. Nat. Med 2006, 13, 95–99. [Google Scholar]
- Das, S.K.; Das, A.R.; Guha, A.K. Gold nanoparticles: Microbial synthesis and application in water hygiene management. Langmuir 2009, 25, 8192–8199. [Google Scholar]
- An inventory of nanotechnology-based consumer products currently on the market. Available online: http://www.nanotechproject.org/inventories/consumer/ (accessed on 23 April 2013).
- Chou, C.C.; Hsiao, H.Y.; Hong, Q.S.; Chen, C.H.; Peng, Y.W.; Chen, H.W.; Yang, P.C. Single-walled carbon nanotubes can induce pulmonary injury in mouse model. Nano Lett 2008, 8, 437–445. [Google Scholar]
- Derfus, A.M.; Chan, W.C.W.; Bhatia, S.N. Probing the cytotoxicity of semiconductor quantum dots. Nano Lett 2004, 4, 11–18. [Google Scholar]
- Bartneck, M.; Ritz, T.; Keul, H.A.; Wambach, M.; Bornemann, J.; Gbureck, U.; Ehling, J.; Lammers, T.; Heymann, F.; Gassler, N. Peptide-functionalized gold nanorods increase liver injury in hepatitis. ACS Nano 2012, 6, 8767–8777. [Google Scholar]
- Schipper, M.L.; Nakayama-Ratchford, N.; Davis, C.R.; Kam, N.W.S.; Chu, P.; Liu, Z.; Sun, X.; Dai, H.; Gambhir, S.S. A pilot toxicology study of single-walled carbon nanotubes in a small sample of mice. Nature Nanotech 2008, 3, 216–221. [Google Scholar]
- Wu, J.; Wang, C.; Sun, J.; Xue, Y. Neurotoxicity of silica nanoparticles: Brain localization and dopaminergic neurons damage pathways. ACS nano 2011, 5, 4476–4489. [Google Scholar]
- Lin, P.; Chen, J.W.; Chang, L.W.; Wu, J.P.; Redding, L.; Chang, H.; Yeh, T.K.; Yang, C.S.; Tsai, M.H.; Wang, H.J. Computational and ultrastructural toxicology of a nanoparticle, Quantum Dot 705, in mice. Environ. Sci. Technol 2008, 42, 6264–6270. [Google Scholar]
- Bai, Y.; Zhang, Y.; Zhang, J.; Mu, Q.; Zhang, W.; Butch, E.R.; Snyder, S.E.; Yan, B. Repeated administrations of carbon nanotubes in male mice cause reversible testis damage without affecting fertility. Nature Nanotech 2010, 5, 683–689. [Google Scholar]
- Song, Y.; Li, X.; Du, X. Exposure to nanoparticles is related to pleural effusion, pulmonary fibrosis and granuloma. Eur. Respir. J 2009, 34, 559–567. [Google Scholar]
- Hillier, S. Current concepts of the roles of follicle stimulating hormone and luteinizing hormone in folliculogenesis. Hum. Reprod 1994, 9, 188–191. [Google Scholar]
- Warren, M.; Perlroth, N. The effects of intense exercise on the female reproductive system. J. Endocrinol 2001, 170, 3–11. [Google Scholar]
- Armenti, A.E.; Zama, A.M.; Passantino, L.; Uzumcu, M. Developmental methoxychlor exposure affects multiple reproductive parameters and ovarian folliculogenesis and gene expression in adult rats. Toxicol. Appl. Pharm 2008, 233, 286–296. [Google Scholar]
- Anway, M.D.; Cupp, A.S.; Uzumcu, M.; Skinner, M.K. Epigenetic transgenerational actions of endocrine disruptors and male fertility. Science 2005, 308, 1466–1469. [Google Scholar]
- Tsuchiya, T.; Oguri, I.; Yamakoshi, Y.N.; Miyata, N. Novel harmful effects of [60]fullerene on mouse embryos in vitro and in vivo. FEBS Lett 1996, 393, 139–145. [Google Scholar]
- Wang, J.; Zhu, X.; Zhang, X.; Zhao, Z.; Liu, H.; George, R.; Wilson-Rawls, J.; Chang, Y.; Chen, Y. Disruption of zebrafish (Danio rerio) reproduction upon chronic exposure to TiO2 nanoparticles. Chemosphere 2011, 83, 461–467. [Google Scholar]
- Shoults-Wilson, W.A.; Reinsch, B.C.; Tsyusko, O.V.; Bertsch, P.M.; Lowry, G.V.; Unrine, J.M. Effect of silver nanoparticle surface coating on bioaccumulation and reproductive toxicity in earthworms (Eisenia fetida). Nanotoxicology 2011, 5, 432–444. [Google Scholar]
- Dooley, K.; Zon, L.I. Zebrafish: A model system for the study of human disease. Curr. Opin. Genet. Dev 2000, 10, 252–256. [Google Scholar]
- Apter, D. Development of the hypothalamic-pituitary-ovarian axis. Ann. N. Y. Acad. Sci 1997, 816, 9–21. [Google Scholar]
- Mattison, D.; Plowchalk, D.; Meadows, M.; Al-Juburi, A.; Gandy, J.; Malek, A. Reproductive toxicity: Male and female reproductive systems as targets for chemical injury. Med. Clin. N. Am 1990, 74, 391–411. [Google Scholar]
- Gao, G.; Ze, Y.; Li, B.; Zhao, X.; Zhang, T.; Sheng, L.; Hu, R.; Gui, S.; Sang, X.; Sun, Q. Ovarian dysfunction and gene-expressed characteristics of female mice caused by long-term exposure to titanium dioxide nanoparticles. J. Hazard. Mater 2012, 243, 19–27. [Google Scholar]
- Blum, J.L.; Xiong, J.Q.; Hoffman, C.; Zelikoff, J.T. Cadmium associated with inhaled cadmium oxide nanoparticles impacts fetal and neonatal development and growth. Toxicol. Sci 2012, 126, 478–486. [Google Scholar]
- Yamashita, K.; Yoshioka, Y.; Higashisaka, K.; Mimura, K.; Morishita, Y.; Nozaki, M.; Yoshida, T.; Ogura, T.; Nabeshi, H.; Nagano, K. Silica and titanium dioxide nanoparticles cause pregnancy complications in mice. Nature Nanotech 2011, 6, 321–328. [Google Scholar]
- Martino-Andrade, A.J.; Chahoud, I. Reproductive toxicity of phthalate esters. Mol. Nutr. Food Res 2009, 54, 148–157. [Google Scholar]
- Olivero, O.A.; Yuspa, S.H.; Poirier, M.C.; Anderson, L.M.; Jones, A.B.; Wang, C.; Diwan, B.A.; Haines, D.C.; Logsdon, D.; Harbaugh, S.W. Transplacental effects of 3′-azido-2′, 3′-dideoxythymidine (AZT): Tumorigenicity in mice and genotoxicity in mice and monkeys. J. Natl. Cancer Inst 1997, 89, 1602–1608. [Google Scholar]
- Perera, F.P.; Rauh, V.; Tsai, W.Y.; Kinney, P.; Camann, D.; Barr, D.; Bernert, T.; Garfinkel, R.; Tu, Y.H.; Diaz, D. Effects of transplacental exposure to environmental pollutants on birth outcomes in a multiethnic population. Environ. Health Persp 2003, 111, 201–205. [Google Scholar]
- Semmler-Behnke, M.; Fertsch, S.; Schmid, G.; Wenk, A.; Kreyling, W.G. Uptake of 1.4 nm versus 18 nm gold nanoparticles in secondary target organs is size dependent in control and pregnant rats after intratracheal or intravenous application. Proceedings of EuroNanoForum 2007, Nanotechnology in Industrial Applications, Düsseldorf, Germany, 19–21 June 2007.
- Takeda, K.; Suzuki, K.; Ishihara, A.; Kubo-Irie, M.; Fujimoto, R.; Tabata, M.; Oshio, S.; Nihei, Y.; Ihara, T.; Sugamata, M. Nanoparticles transferred from pregnant mice to their offspring can damage the genital and cranial nerve systems. J. Health Sci 2009, 55, 95–102. [Google Scholar]
- Refuerzo, J.S.; Godin, B.; Bishop, K.; Srinivasan, S.; Shah, S.K.; Amra, S.; Ramin, S.M.; Ferrari, M. Size of the nanovectors determines the transplacental passage in pregnancy: Study in rats. Am. J. Obstet. Gynecol 2011, 204, 546, e5–546.e9.. [Google Scholar]
- Chu, M.; Wu, Q.; Yang, H.; Yuan, R.; Hou, S.; Yang, Y.; Zou, Y.; Xu, S.; Xu, K.; Ji, A.; et al. Transfer of quantum dots from pregnant mice to pups across the placental barrier. Small 2010, 6, 670–678. [Google Scholar]
- Sumner, S.C.J.; Fennell, T.R.; Snyder, R.W.; Taylor, G.F.; Lewin, A.H. Distribution of carbon-14 labeled C60 ([14C] C60) in the pregnant and in the lactating dam and the effect of C60 exposure on the biochemical profile of urine. J. Appl. Toxicol 2010, 30, 354–360. [Google Scholar]
- Tian, F.; Razansky, D.; Estrada, G.G.; Semmler-Behnke, M.; Beyerle, A.; Kreyling, W.; Ntziachristos, V.; Stoeger, T. Surface modification and size dependence in particle translocation during early embryonic development. Inhal. Toxicol 2009, 21, 92–96. [Google Scholar]
- Morgan, K. Development of a preliminary framework for informing the risk analysis and risk management of nanoparticles. Risk Anal 2005, 25, 1621–1635. [Google Scholar]
- Zhang, L.; Fischer, W.; Pippel, E.; Hause, G.; Brandsch, M.; Knez, M. Receptor-mediated cellular uptake of nanoparticles: A switchable delivery system. Small 2011, 7, 1538–1541. [Google Scholar]
- Wick, P.; Malek, A.; Manser, P.; Meili, D.; Maeder-Althaus, X.; Diener, L.; Diener, P.-A.; Zisch, A.; Krug, H.F.; von Mandach, U. Barrier capacity of human placenta for nanosized materials. Environ. Health Persp 2009, 118, 432–436. [Google Scholar]
- Fujimoto, A.; Tsukue, N.; Watanabe, M.; Sugawara, I.; Yanagisawa, R.; Takano, H.; Yoshida, S.; Takeda, K. Diesel exhaust affects immunological action in the placentas of mice. Environ. Toxicol 2005, 20, 431–440. [Google Scholar]
- Umezawa, M.; Kudo, S.; Yanagita, S.; Shinkai, Y.; Niki, R.; Oyabu, T.; Takeda, K.; Ihara, T.; Sugamata, M. Maternal exposure to carbon black nanoparticle increases collagen type VIII expression in the kidney of offspring. J. Toxicol. Sci 2011, 36, 461–468. [Google Scholar]
- Jackson, P.; Halappanavar, S.; Hougaard, K.S.; Williams, A.; Madsen, A.M.; Lamson, J.S.; Andersen, O.; Yauk, C.; Wallin, H.; Vogel, U. Maternal inhalation of surface-coated nanosized titanium dioxide (UV-Titan) in C57BL/6 mice: Effects in prenatally exposed offspring on hepatic DNA damage and gene expression. Nanotoxicology 2012. [Google Scholar] [CrossRef]
- Philbrook, N.A.; Walker, V.K.; Afrooz, A.R.M.N.; Saleh, N.B.; Winn, L.M. Investigating the effects of functionalized carbon nanotubes on reproduction and development in Drosophila melanogaster and CD-1 mice. Reprod. Toxicol 2011, 32, 442–448. [Google Scholar]
- Hougaard, K.S.; Jackson, P.; Jensen, K.A.; Sloth, J.J.; Löschner, K.; Larsen, E.H.; Birkedal, R.K.; Vibenholt, A.; Boisen, A.-M.Z.; Wallin, H.; et al. Effects of prenatal exposure to surface-coated nanosized titanium dioxide (UV-Titan). A study in mice. Part. Fiber Toxicol. 2010, 7. [Google Scholar] [CrossRef]
- Yoshida, S.; Hiyoshi, K.; Oshio, S.; Takano, H.; Takeda, K.; Ichinose, T. Effects of fetal exposure to carbon nanoparticles on reproductive function in male offspring. Fertil. Steril 2010, 93, 1695–1699. [Google Scholar]
- Hemmingsen, J.G.; Hougaard, K.S.; Talsness, C.; Wellejus, A.; Loft, S.; Wallin, H.; Møller, P. Prenatal exposure to diesel exhaust particles and effect on the male reproductive system in mice. Toxicology 2009, 264, 61–68. [Google Scholar]
- Li, C.M.; Taneda, S.; Taya, K.; Watanabe, G.; Li, X.; Fujitani, Y.; Nakajima, T.; Suzuki, A.K. Effects of in utero exposure to nanoparticle-rich diesel exhaust on testicular function in immature male rats. Toxicol. Lett 2009, 185, 1–8. [Google Scholar]
- Takahashi, Y.; Mizuo, K.; Shinkai, Y.; Oshio, S.; Takeda, K. Prenatal exposure to titanium dioxide nanoparticles increases dopamine levels in the prefrontal cortex and neostriatum of mice. J. Toxicol. Sci 2010, 35, 749–756. [Google Scholar]
- Shimizu, M.; Tainaka, H.; Oba, T.; Mizuo, K.; Umezawa, M.; Takeda, K. Maternal exposure to nanoparticulate titanium dioxide during the prenatal period alters gene expression related to brain development in the mouse. Part. Fiber Toxicol. 2009, 6. [Google Scholar] [CrossRef]
- Yokota, S.; Mizuo, K.; Moriya, N.; Oshio, S.; Sugawara, I.; Takeda, K. Effect of prenatal exposure to diesel exhaust on dopaminergic system in mice. Neurosci. Lett 2009, 449, 38–41. [Google Scholar]
- Sugamata, M.; Ihara, T.; Takano, H.; Oshio, S.; Takeda, K. Maternal diesel exhaust exposure damages newborn murine brains. J. Health Sci 2006, 52, 82–84. [Google Scholar]
- Hougaard, K.S.; Jensen, K.A.; Nordly, P.; Taxvig, C.; Vogel, U.; Saber, A.T.; Wallin, H. Effects of prenatal exposure to diesel exhaust particles on postnatal development, behavior, genotoxicity and inflammation in mice. Part. Fibre Toxicol. 2008, 5. [Google Scholar] [CrossRef]
- Jackson, P.; Vogel, U.; Wallin, H.; Hougaard, K.S. Prenatal exposure to carbon black (Printex 90): Effects on sexual development and neurofunction. Basic Clin. Pharmacol 2011, 109, 434–437. [Google Scholar]
- Pietroiusti, A.; Massimiani, M.; Fenoglio, I.; Colonna, M.; Valentini, F.; Palleschi, G.; Camaioni, A.; Magrini, A.; Siracusa, G.; Bergamaschi, A. Low doses of pristine and oxidized single wall carbon nanotubes affect mammalian embryonic development. ACS Nano 2011, 5, 4624–4633. [Google Scholar]
- Lim, J.-H.; Kim, S.-H.; Shin, I.-S.; Park, N.-H.; Moon, C.; Kang, S.-S.; Kim, S.-H.; Park, S.-C.; Kim, J.-C. Maternal exposure to multi-wall carbon nanotubes does not induce embryo-fetal developmental toxicity in rats. Birth Defects Res. B 2011, 92, 69–76. [Google Scholar]
- Park, E.J.; Kim, H.; Kim, Y.; Park, K. Effects of platinum nanoparticles on the postnatal development of mouse pups by maternal exposure. Evn. Heal.Toxicol 2010, 25, 279–286. [Google Scholar]
- Tsukue, N.; Tsubone, H.; Suzuki, A.K. Diesel exhaust affects the abnormal delivery in pregnant mice and the growth of their young. Inhal. Toxicol 2002, 14, 635–651. [Google Scholar]
- Ono, N.; Oshio, S.; Niwata, Y.; Yoshida, S.; Tsukue, N.; Sugawara, I.; Takano, H.; Takeda, K. Prenatal exposure to diesel exhaust impairs mouse spermatogenesis. Inhal. Toxicol 2007, 19, 275–281. [Google Scholar]
- Kubo-Irie, M.; Oshio, S.; Niwata, Y.; Ishihara, A.; Sugawara, I.; Takeda, K. Pre-and postnatal exposure to low-dose diesel exhaust impairs murine spermatogenesis. Inhal. Toxicol 2011, 23, 805–813. [Google Scholar]
- Watanabe, N. Decreased number of sperms and Sertoli cells in mature rats exposed to diesel exhaust as fetuses. Toxicol. Lett 2005, 155, 51–58. [Google Scholar]
- Freire, C.; Ramos, R.; Puertas, R.; Lopez-Espinosa, M.J.; Julvez, J.; Aguilera, I.; Cruz, F.; Fernandez, M.F.; Sunyer, J.; Olea, N. Association of traffic-related air pollution with cognitive development in children. J. Epidemiol. Comm. H 2010, 64, 223–228. [Google Scholar]
- Sugamata, M.; Ihara, T.; Umezawa, M.; Takeda, K. P-999-Maternal exposure to nanoparticles enhances the risk of mental neurological disorders in offspring. Eur. Psychiat 2012, 27, 1. [Google Scholar]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-κB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol 2006, 72, 1493–1505. [Google Scholar]
- Jomova, K.; Jenisova, Z.; Feszterova, M.; Baros, S.; Liska, J.; Hudecova, D.; Rhodes, C.; Valko, M. Arsenic: Toxicity, oxidative stress and human disease. J. Appl. Toxicol 2011, 31, 95–107. [Google Scholar]
- Peden, D.B. The role of oxidative stress and innate immunity in O3 and endotoxin-induced human allergic airway disease. Immunol. Rev 2011, 242, 91–105. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1999; Volume 3, pp. 1–543. [Google Scholar]
- Schoonbroodt, S.; Piette, J. Oxidative stress interference with the nuclear factor-kappa B activation pathways. Biochem. Pharmacol 2000, 60, 1075–1083. [Google Scholar]
- Wang, X.; Martindale, J.L.; Liu, Y.; Holbrook, N.J. The cellular response to oxidative stress: Influences of mitogen-activated protein kinase signalling pathways on cell survival. Biochem. J 1998, 333, 291–300. [Google Scholar]
- Ornoy, A. Embryonic oxidative stress as a mechanism of teratogenesis with special emphasis on diabetic embryopathy. Reprod. Toxicol 2007, 24, 31–41. [Google Scholar]
- Wells, P.G.; Winn, L.M. Biochemical toxicology of chemical teratogenesis. Crit. Rev. Biochem. Mol 1996, 31, 1–40. [Google Scholar]
- Wells, P.G.; Bhuller, Y.; Chen, C.S.; Jeng, W.; Kasapinovic, S.; Kennedy, J.C.; Kim, P.M.; Laposa, R.R.; McCallum, G.P.; Nicol, C.J. Molecular and biochemical mechanisms in teratogenesis involving reactive oxygen species. Toxicol. Appl. Pharm 2005, 207, 354–366. [Google Scholar]
- Li, Y.; Liu, Y.; Fu, Y.; Wei, T.; Le Guyader, L.; Gao, G.; Liu, R.S.; Chang, Y.Z.; Chen, C. The triggering of apoptosis in macrophages by pristine graphene through the MAPK and TGF-beta signaling pathways. Biomaterials 2011, 33, 402–411. [Google Scholar]
- Meyer, U.; Feldon, J.; Fatemi, S.H. In-vivo rodent models for the experimental investigation of prenatal immune activation effects in neurodevelopmental brain disorders. Neurosci. Biobehav. Rev 2009, 33, 1061–1079. [Google Scholar]
- Tkachenko, A.G.; Xie, H.; Coleman, D.; Glomm, W.; Ryan, J.; Anderson, M.F.; Franzen, S.; Feldheim, D.L. Multifunctional gold nanoparticle-peptide complexes for nuclear targeting. J. Am. Chem. Soc 2003, 125, 4700–4701. [Google Scholar]
- Zhu, L.; Chang, D.W.; Dai, L.; Hong, Y. DNA damage induced by multiwalled carbon nanotubes in mouse embryonic stem cells. Nano Lett 2007, 7, 3592–3597. [Google Scholar]
- Li, P.-W.; Kuo, T.-H.; Chang, J.-H.; Yeh, J.-M.; Chan, W.-H. Induction of cytotoxicity and apoptosis in mouse blastocysts by silver nanoparticles. Toxicol. Lett 2010, 197, 82–87. [Google Scholar]
- Chan, W.; Shiao, N. Cytotoxic effect of CdSe quantum dots on mouse embryonic development. Acta Pharmacol. Sin 2008, 29, 259–266. [Google Scholar]
- Stelzer, R.; Hutz, R.J. Gold nanoparticles enter rat ovarian granulosa cells and subcellular organelles, and alter in vitro estrogen accumulation. J. Reprod. Develop 2009, 55, 685–690. [Google Scholar]
- Amatruda, J.F.; Shepard, J.L.; Stern, H.M.; Zon, L.I. Zebrafish as a cancer model system. Cancer Cell 2002, 1, 229–231. [Google Scholar]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a model vertebrate for investigating chemical toxicity. Toxicol. Sci 2005, 86, 6–19. [Google Scholar]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dynam 1995, 203, 253–310. [Google Scholar]
- Fadool, J.M.; Dowling, J.E. Zebrafish: A model system for the study of eye genetics. Prog. Retin. Eye Res 2008, 27, 89–110. [Google Scholar]
- Zon, L.I.; Peterson, R.T. In vivo drug discovery in the zebrafish. Nat. Rev. Drug Discov 2005, 4, 35–44. [Google Scholar]
- Selman, K.; Wallace, R.A.; Sarka, A.; Qi, X. Stages of oocyte development in the zebrafish, Brachydanio rerio. J. Morphol 1993, 218, 203–224. [Google Scholar]
- Bonsignorio, D.; Perego, L.; Giacco, L.D.; Cotelli, F. Structure and macromolecular composition of the zebrafish egg chorion. Zygote 1996, 4, 101–108. [Google Scholar]
- Lee, K.J.; Nallathamby, P.D.; Browning, L.M.; Osgood, C.J.; Xu, X.H.N. In vivo imaging of transport and biocompatibility of single silver nanoparticles in early development of zebrafish embryos. ACS Nano 2007, 1, 133–143. [Google Scholar]
- Nallathamby, P.D.; Lee, K.J.; Xu, X.H.N. Design of stable and uniform single nanoparticle photonics for in vivo dynamics imaging of nanoenvironments of zebrafish embryonic fluids. ACS Nano 2008, 2, 1371–1380. [Google Scholar]
- Cheng, J.; Flahaut, E.; Cheng, S.H. Effect of carbon nanotubes on developing zebrafish (Danio rerio) embryos. Environ. Toxicol. Chem 2007, 26, 708–716. [Google Scholar]
- Fent, K.; Weisbrod, C.J.; Wirth-Heller, A.; Pieles, U. Assessment of uptake and toxicity of fluorescent silica nanoparticles in zebrafish (Danio rerio) early life stages. Aquat. Toxicol 2010, 100, 218–228. [Google Scholar]
- Asharani, P.V.; lianwu, Y.; Gong, Z.; Valiyaveettil, S. Comparison of the toxicity of silver, gold and platinum nanoparticles in developing zebrafish embryos. Nanotoxicology 2011, 5, 43–54. [Google Scholar]
- George, S.; Lin, S.; Ji, Z.; Thomas, C.R.; Li, L.J.; Mecklenburg, M.; Meng, H.; Wang, X.; Zhang, H.; Xia, T. Surface defects on plate-shaped silver nanoparticles contribute to its hazard potential in a fish gill cell line and zebrafish embryos. ACS Nano 2012, 6, 3745–3759. [Google Scholar]
- Truong, L.; Tilton, S.C.; Zaikova, T.; Richman, E.; Waters, K.M.; Hutchison, J.E.; Tanguay, R.L. Surface functionalities of gold nanoparticles impact embryonic gene expression responses. Nanotoxicology 2013, 7, 192–201. [Google Scholar]
- Bar-Ilan, O.; Albrecht, R.M.; Fako, V.E.; Furgeson, D.Y. Toxicity assessments of multisized gold and silver nanoparticles in zebrafish embryos. Small 2009, 5, 1897–1910. [Google Scholar]
- Browning, L.M.; Lee, K.J.; Huang, T.; Nallathamby, P.D.; Lowman, J.E.; Xu, X.H.N. Random walk of single gold nanoparticles in zebrafish embryos leading to stochastic toxic effects on embryonic developments. Nanoscale 2009, 1, 138–152. [Google Scholar]
- Jovanović, B.; Ji, T.; Palić, D. Gene expression of zebrafish embryos exposed to titanium dioxide nanoparticles and hydroxylated fullerenes. Ecotoxicol. Environ. Saf 2011, 74, 1518–1525. [Google Scholar]
- Zhu, X.; Zhu, L.; Duan, Z.; Qi, R.; Li, Y.; Lang, Y. Comparative toxicity of several metal oxide nanoparticle aqueous suspensions to Zebrafish (Danio rerio) early developmental stage. J. Environ. Sci. Health A 2008, 43, 278–284. [Google Scholar]
- Bar-Ilan, O.; Louis, K.M.; Yang, S.P.; Pedersen, J.A.; Hamers, R.J.; Peterson, R.E.; Heideman, W. Titanium dioxide nanoparticles produce phototoxicity in the developing zebrafish. Nanotoxicology 2012, 6, 670–679. [Google Scholar]
- Truong, L.; Moody, I.S.; Stankus, D.P.; Nason, J.A.; Lonergan, M.C.; Tanguay, R.L. Differential stability of lead sulfide nanoparticles influences biological responses in embryonic zebrafish. Arch. Toxicol 2010, 85, 787–798. [Google Scholar]
- Lei, Y.; Xiao, Q.; Huang, S.; Xu, W.; Zhang, Z.; He, Z.; Liu, Y.; Deng, F. Impact of CdSe/ZnS quantum dots on the development of zebrafish embryos. J. Nanopart. Res 2011, 13, 6895–6906. [Google Scholar]
- Zhang, W.; Sun, X.; Chen, L.; Lin, K.F.; Dong, Q.X.; Huang, C.J.; Fu, R.B.; Zhu, J. Toxicological effect of joint cadmium selenium quantum dots and copper ion exposure on zebrafish. Environ. Toxicol. Chem 2012, 31, 2117–2123. [Google Scholar]
- Zhang, W.; Lin, K.; Miao, Y.; Dong, Q.; Huang, C.; Wang, H.; Guo, M.; Cui, X. Toxicity assessment of zebrafish following exposure to CdTe QDs. J. Hazard. Mater. 2012, 213–214, 413–420. [Google Scholar]
- Zhang, W.; Lin, K.; Sun, X.; Dong, Q.; Huang, C.; Wang, H.; Guo, M.; Cui, X. Toxicological effect of MPA–CdSe QDs exposure on zebrafish embryo and larvae. Chemosphere 2012, 89, 52–59. [Google Scholar]
- King-Heiden, T.C.; Wiecinski, P.N.; Mangham, A.N.; Metz, K.M.; Nesbit, D.; Pedersen, J.A.; Hamers, R.J.; Heideman, W.; Peterson, R.E. Quantum dot nanotoxicity assessment using the zebrafish embryo. Environ. Sci. Technol 2009, 43, 1605–1611. [Google Scholar]
- Asharani, P.; Serina, N.; Nurmawati, M.; Wu, Y.; Gong, Z.; Valiyaveettil, S. Impact of multi-walled carbon nanotubes on aquatic species. J. Nanosci. Nanotechnol 2008, 8, 3603–3609. [Google Scholar]
- Cheng, J.; Cheng, S.H. Influence of carbon nanotube length on toxicity to zebrafish embryos. Int. J. Nanomed 2012, 7, 3731–3739. [Google Scholar]
- Usenko, C.Y.; Harper, S.L.; Tanguay, R.L. In vivo evaluation of carbon fullerene toxicity using embryonic zebrafish. Carbon 2007, 45, 1891–1898. [Google Scholar]
- Yamakoshi, Y.; Umezawa, N.; Ryu, A.; Arakane, K.; Miyata, N.; Goda, Y.; Masumizu, T.; Nagano, T. Active oxygen species generated from photoexcited fullerene (C60) as potential medicines: O2 −.versus 1O2. J. Am. Chem. Soc 2003, 125, 12803–12809. [Google Scholar]
- Usenko, C.Y.; Harper, S.L.; Tanguay, R.L. Fullerene C60 exposure elicits an oxidative stress response in embryonic zebrafish. Toxicol. Appl. Pharm 2008, 229, 44–55. [Google Scholar]
- Zhu, R.R.; Wang, S.L.; Chao, J.; Shi, D.L.; Zhang, R.; Sun, X.Y.; Yao, S.D. Bio-effects of Nano-TiO2 on DNA and cellular ultrastructure with different polymorph and size. Mater. Sci. Eng. C 2009, 29, 691–696. [Google Scholar]
- Gorth, D.J.; Rand, D.M.; Webster, T.J. Silver nanoparticle toxicity in Drosophila: Size does matter. Int. J. Nanomed 2011, 6, 343–350. [Google Scholar]
- Ispas, C.; Andreescu, D.; Patel, A.; Goia, D.V.; Andreescu, S.; Wallace, K.N. Toxicity and developmental defects of different sizes and shape nickel nanoparticles in zebrafish. Environ. Sci. Technol 2009, 43, 6349–6356. [Google Scholar]
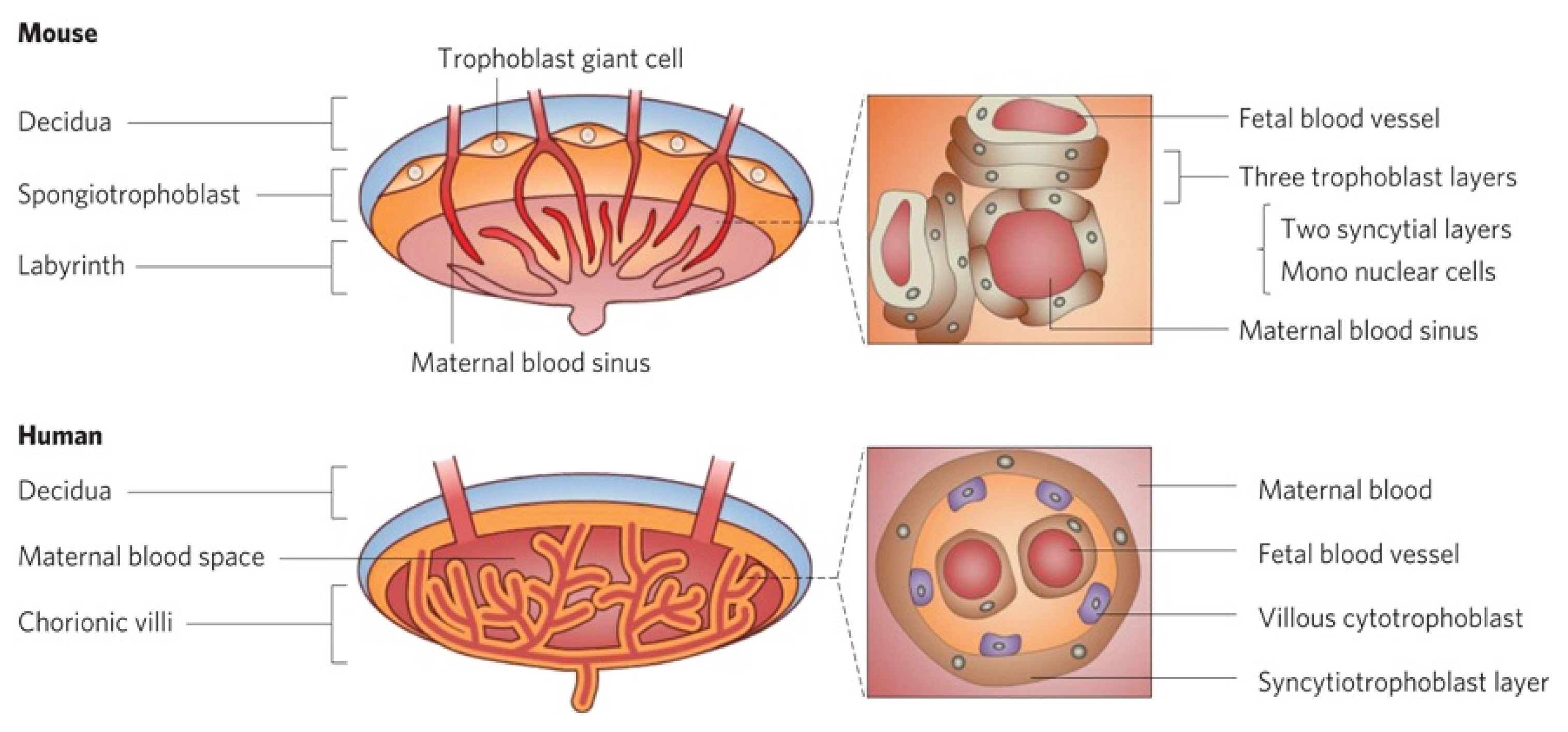
- Carter, A. Animal models of human placentation–a review. Placenta 2007, 28, S41–S47. [Google Scholar]
- Malassine, A.; Frendo, J.L.; Evain-Brion, D. A comparison of placental development and endocrine functions between the human and mouse model. Hum. Reprod. Update 2003, 9, 531–539. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticles/characteristics | Mouse strain | Exposure | Dose | Duration | Toxicity in offspring | Ref. |
|---|---|---|---|---|---|---|
| SWCNT, 1–2 nm in diameter, 5–30 μm in length | CD-1 | Oral gavage | 5, 10 or 100 mg/kg | GD 9 | Skeletal abnormalities and external defects | [41] |
| TiO2, rutile, 21 nm, coated with polyalcohol | C57BL/6BomTac | Inhalation | 42 mg/m3 | GD 8–18, 1 h/day | Avoidance of the central zone in the open field test; enhanced prepulse inhibition in female offspring | [42] |
| TiO2, anatase, 25–70 nm, surface area of 20–25 m2/g | Slc:ICR | Subcutaneous injection | 100 μg/mouse | 3, 7, 10 and 14 days post-coitus | Decreased daily sperm production and sperm motility; disorganised and disrupted seminiferous tubules; apoptosis in the olfactory bulb | [30] |
| Carbon nanoparticles, 14 nm | ICR | Intratracheal injection | 200 μg/mouse | GD 7 and14 | Decreased daily sperm production | [43] |
| Diesel exhaust | C57BL/6BomTac | Inhalation | 20 mg/m3, 1 × 106 particles/cm3 | GD 7–19, 1 h/day | Decreased daily sperm production | [44] |
| Nanoparticle-rich diesel exhaust, filtered diesel exhaust | F344/DuCrlCrli | Inhalation | Nanoparticle-rich DE: 168.84 μg/m3, 1.36 × 106 particles/cm3; filtered-DE: 3.1 μg/m3, 2.66 particles/cm3 | GD 1–19 | Decreased seminal vesicle and prostate organ index; decreased testosterone, progesterone, corticosterone and FSH levels; altered steroidogenic acute regulatory protein, 17β-hydroxysteroid dehydrogenase and follicle-stimulating hormone receptor mRNA | [45] |
| TiO2, anatase, 25–70 nm, surface area of 20–25 m2/g | ICR | Subcutaneous injection | 0.1 mL, 1 mg/mL | GD 6, 9, 12, 15 and 18 | Increased DA and metabolites in the prefrontal cortex and neostriatum | [46] |
| TiO2, anatase, 25–70 nm, surface area of 20–25 m2/g | ICR | Subcutaneous injection | 100 μL, 1 mg/mL | GD 6, 9, 12 and 15 | Altered gene expression associated with apoptosis, oxidative stress and neurotransmitters in the brain | [47] |
| Diesel exhaust | ICR | Inhalation | 1.0 mg/m3 | GD2–17, 8 h/day, 5 days per week | Reduced locomotion; decreased DA turnover in the striatum and nucleus accumbens | [48] |
| Diesel exhaust | ICR | Inhalation | 0.3, 1 and 3.0 particles/m3 | 2 to 16 days post-coitus | Apoptosis in brain tissue | [49] |
| Diesel exhaust, 240 nm | C57BL/6 BomTac | Inhalation | 19 mg/m3, 1 × 106 particles/cm3 | GD 9–19, 1 h/day | Increased activity in female DE offspring | [50] |
| Carbon black, average zeta potential of 140 nm, hydrodynamic size of 50–60 nm | C57BL/6BomTac | Intratracheal instillation | 11, 54 and 268 μg/animal | GD 7, 10, 15 and 18 | Altered habituation pattern in female offspring | [51] |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sun, J.; Zhang, Q.; Wang, Z.; Yan, B. Effects of Nanotoxicity on Female Reproductivity and Fetal Development in Animal Models. Int. J. Mol. Sci. 2013, 14, 9319-9337. https://doi.org/10.3390/ijms14059319
Sun J, Zhang Q, Wang Z, Yan B. Effects of Nanotoxicity on Female Reproductivity and Fetal Development in Animal Models. International Journal of Molecular Sciences. 2013; 14(5):9319-9337. https://doi.org/10.3390/ijms14059319
Chicago/Turabian StyleSun, Jianling, Qiu Zhang, Zhiping Wang, and Bing Yan. 2013. "Effects of Nanotoxicity on Female Reproductivity and Fetal Development in Animal Models" International Journal of Molecular Sciences 14, no. 5: 9319-9337. https://doi.org/10.3390/ijms14059319
APA StyleSun, J., Zhang, Q., Wang, Z., & Yan, B. (2013). Effects of Nanotoxicity on Female Reproductivity and Fetal Development in Animal Models. International Journal of Molecular Sciences, 14(5), 9319-9337. https://doi.org/10.3390/ijms14059319