
Production of Polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus Strains Isolated from Salt Fields


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Strain Screenings
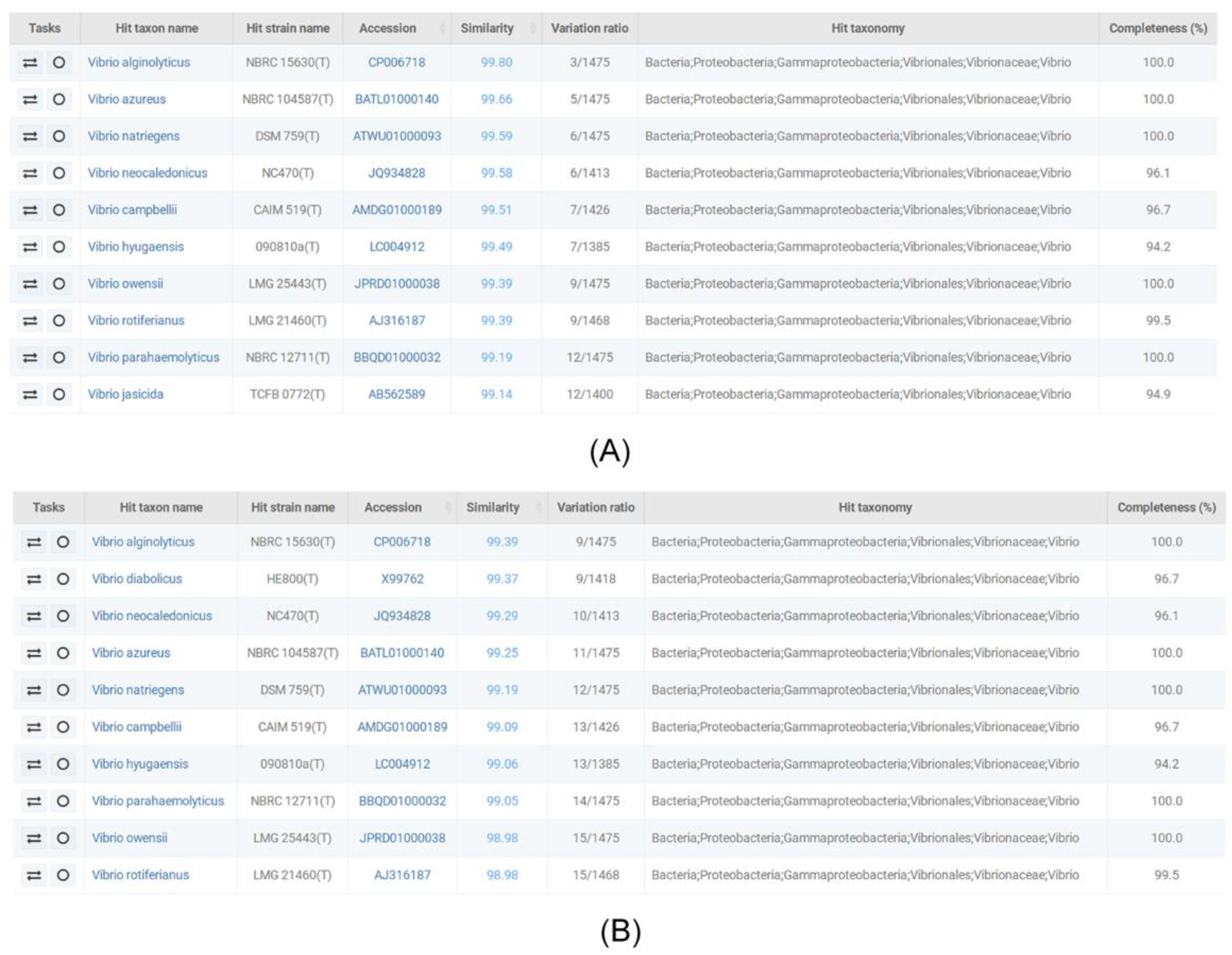
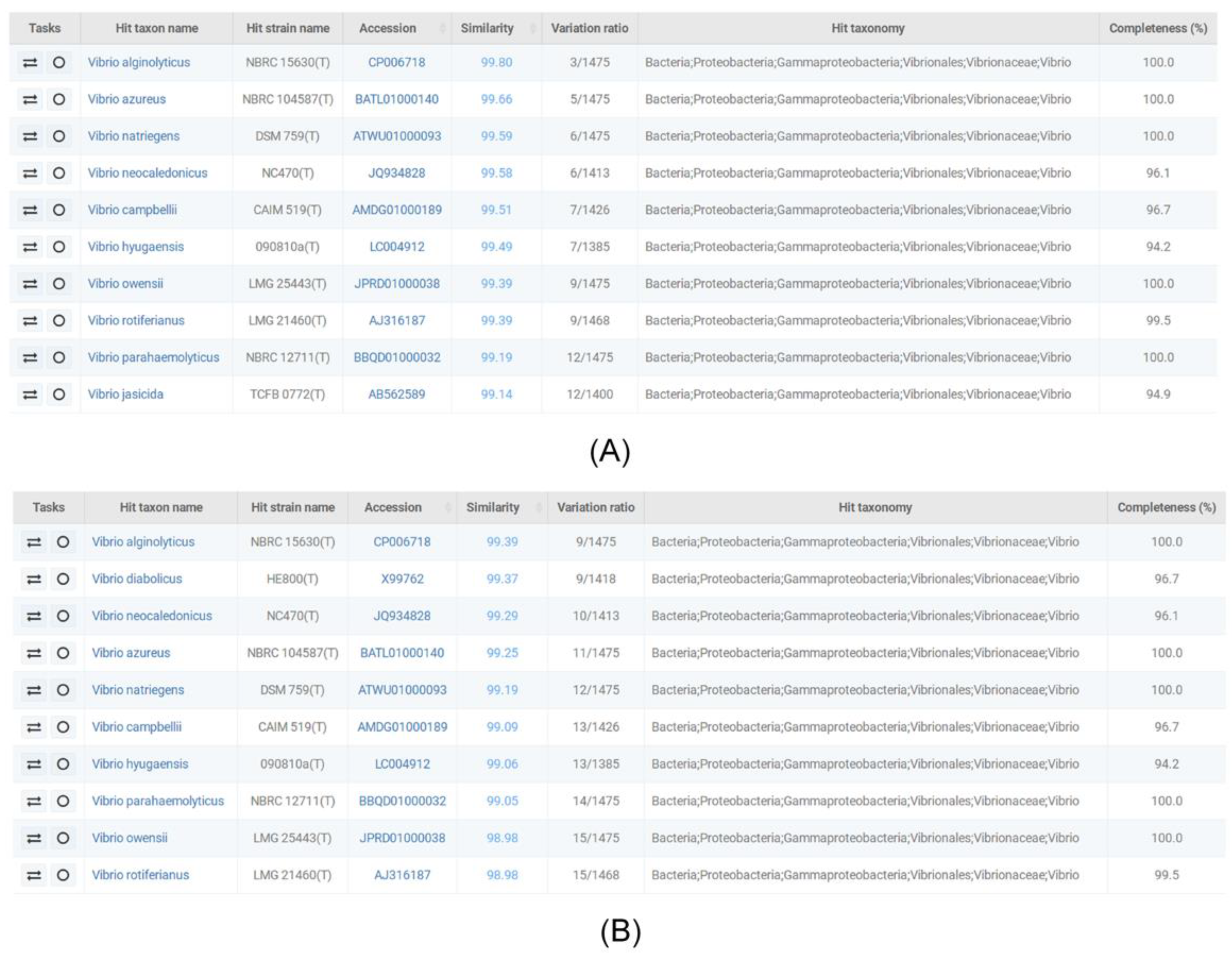
2.2. 16S rDNA Sequencing and Homology Analysis
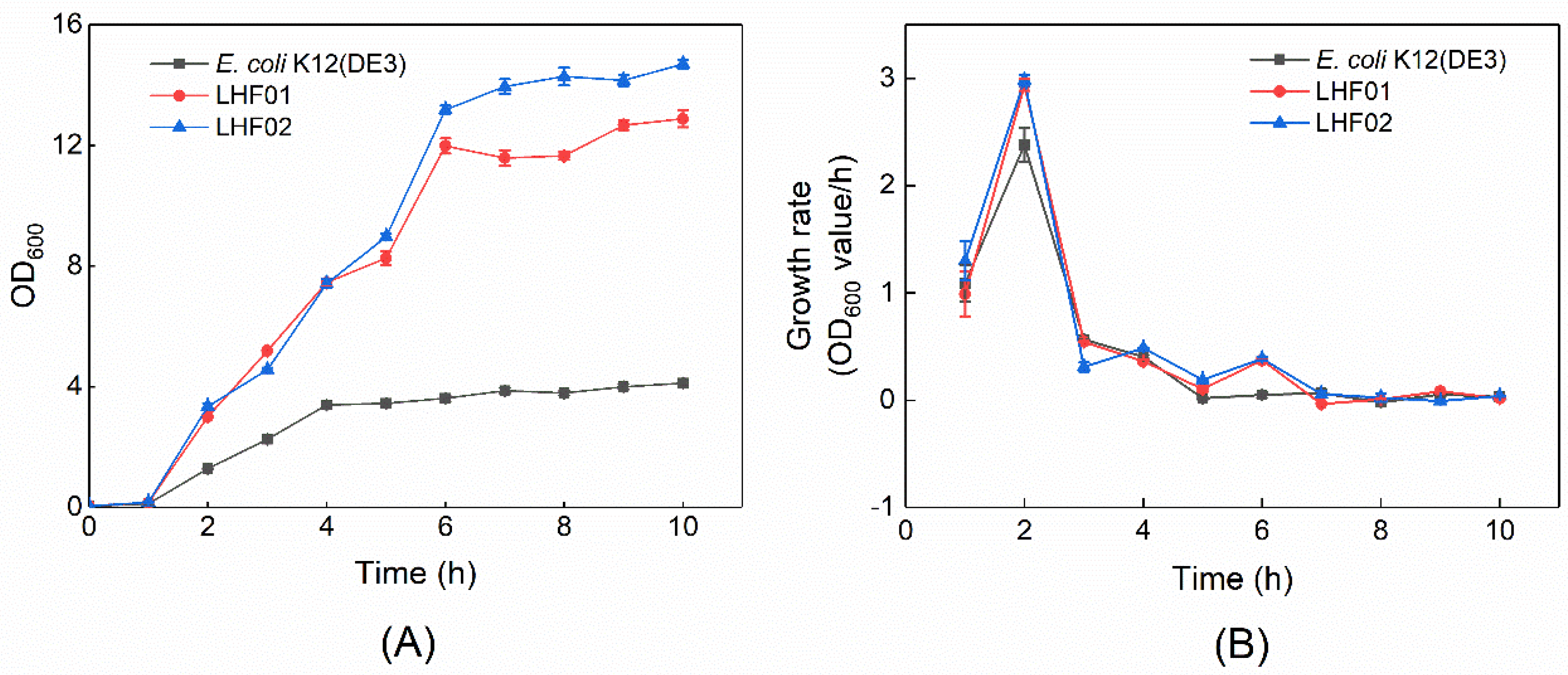
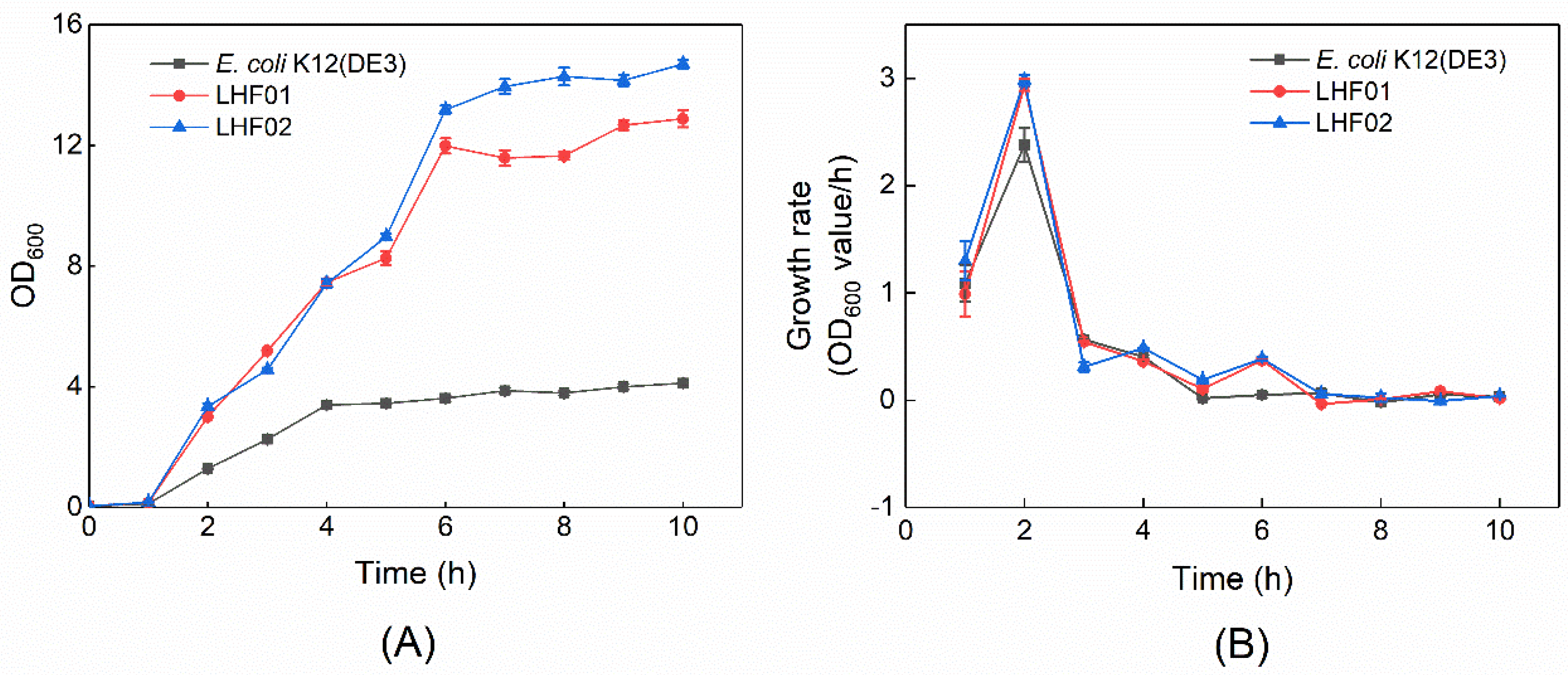
2.3. Cell Growth Rate and Antibiotic Resistance Tests of V. alginolyticus
2.4. Culture Conditions for PHA Production
2.5. Quantification of PHA and Cell Growth
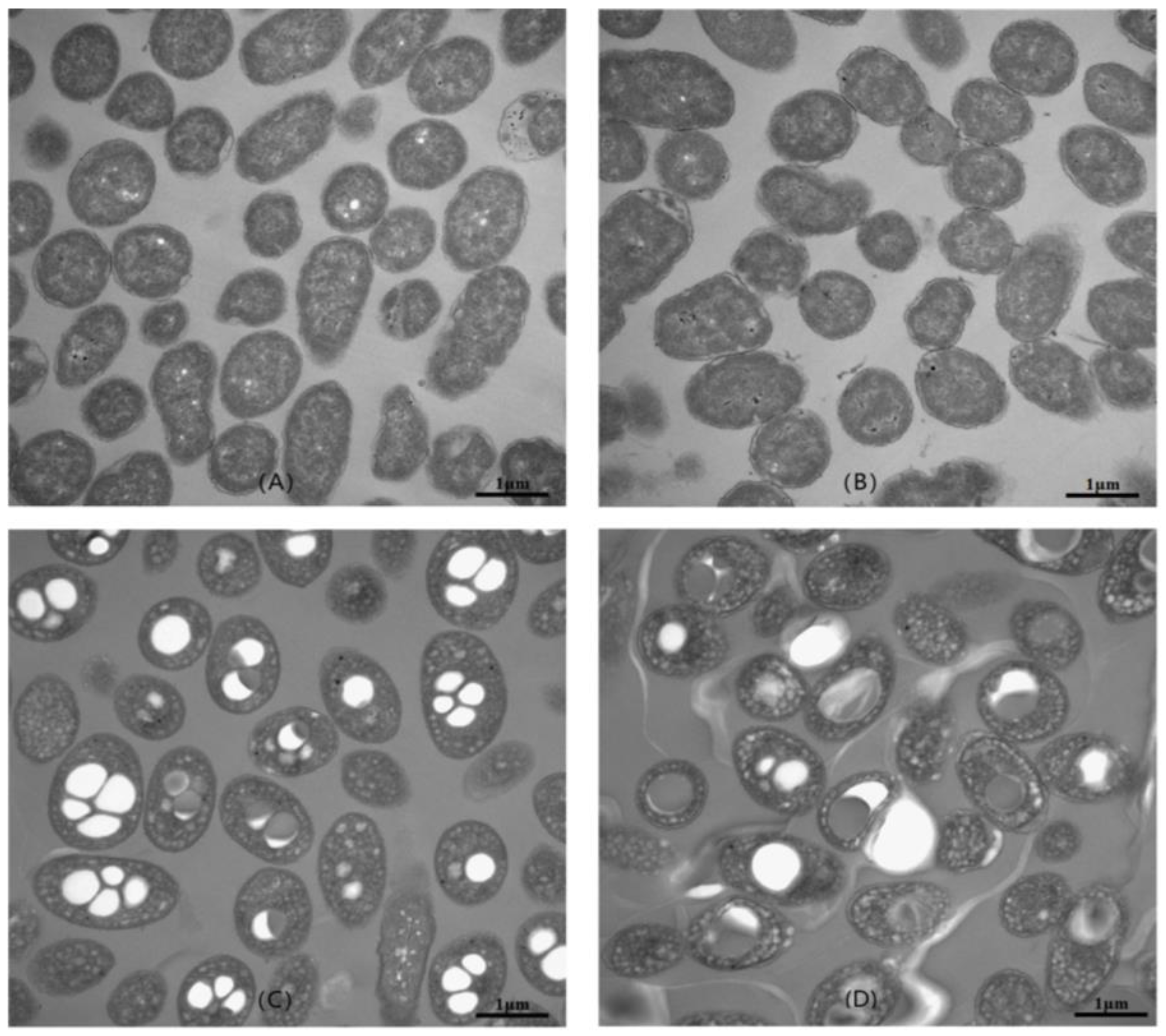
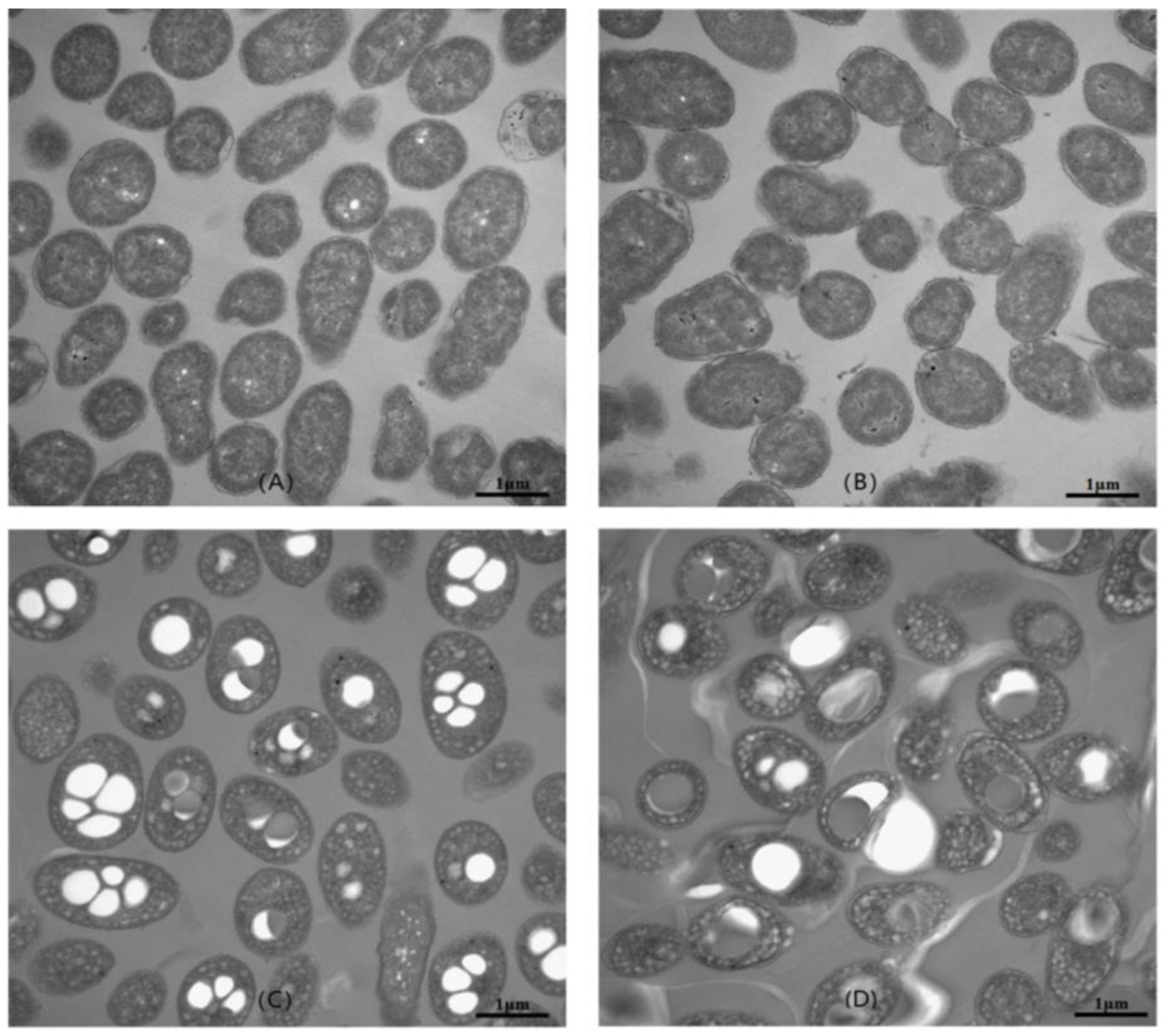
2.6. Transmission Electron Microscopy (TEM) Analysis of Cell Morphology
2.7. PHA Extraction and Molecular Weight Determination
3. Results and Discussion
3.1. Isolation of V. alginolyticus Strains
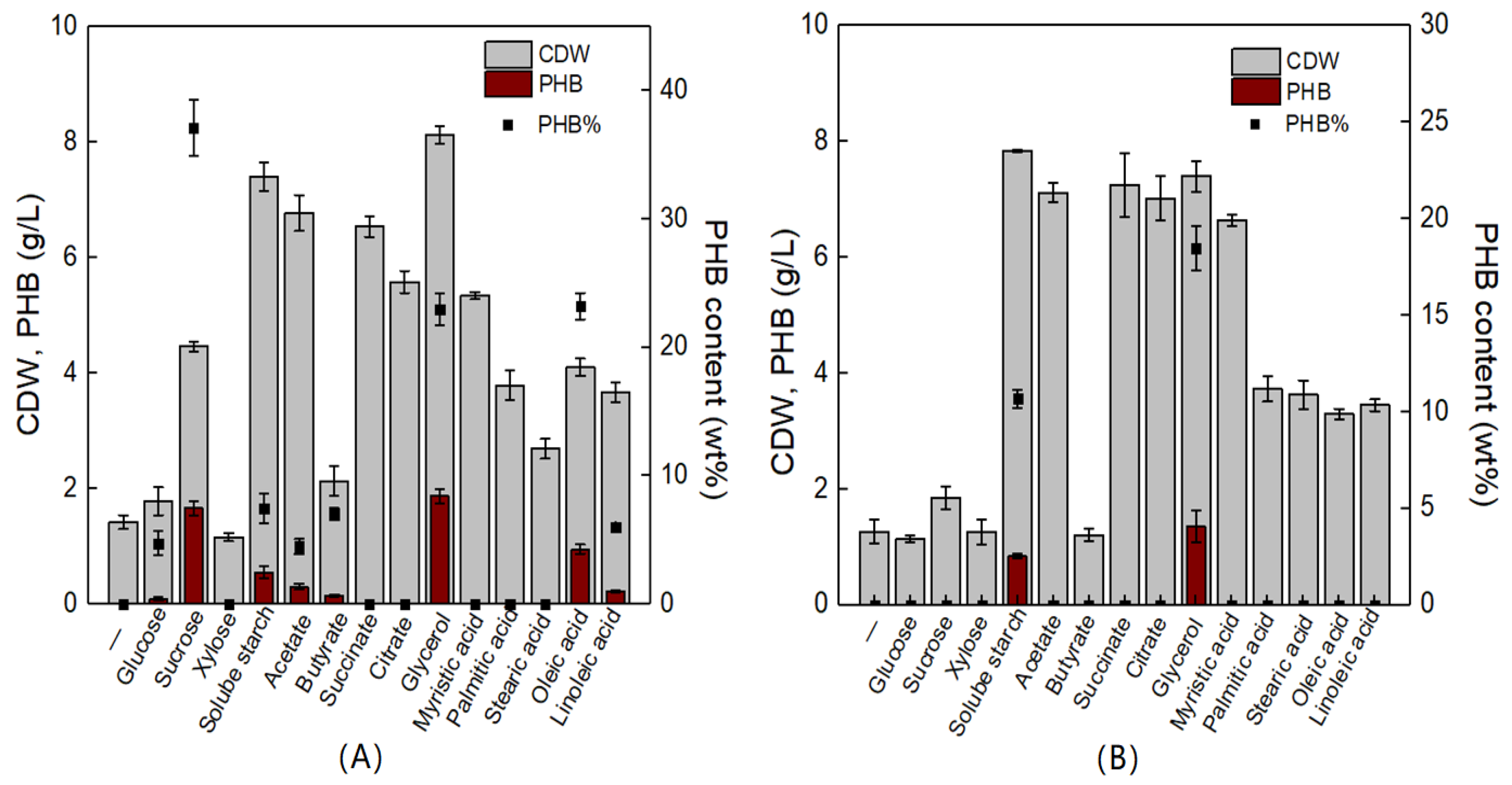
3.2. PHB Production by V. alginolyticus LHF01 and LHF02 on Various Substrates
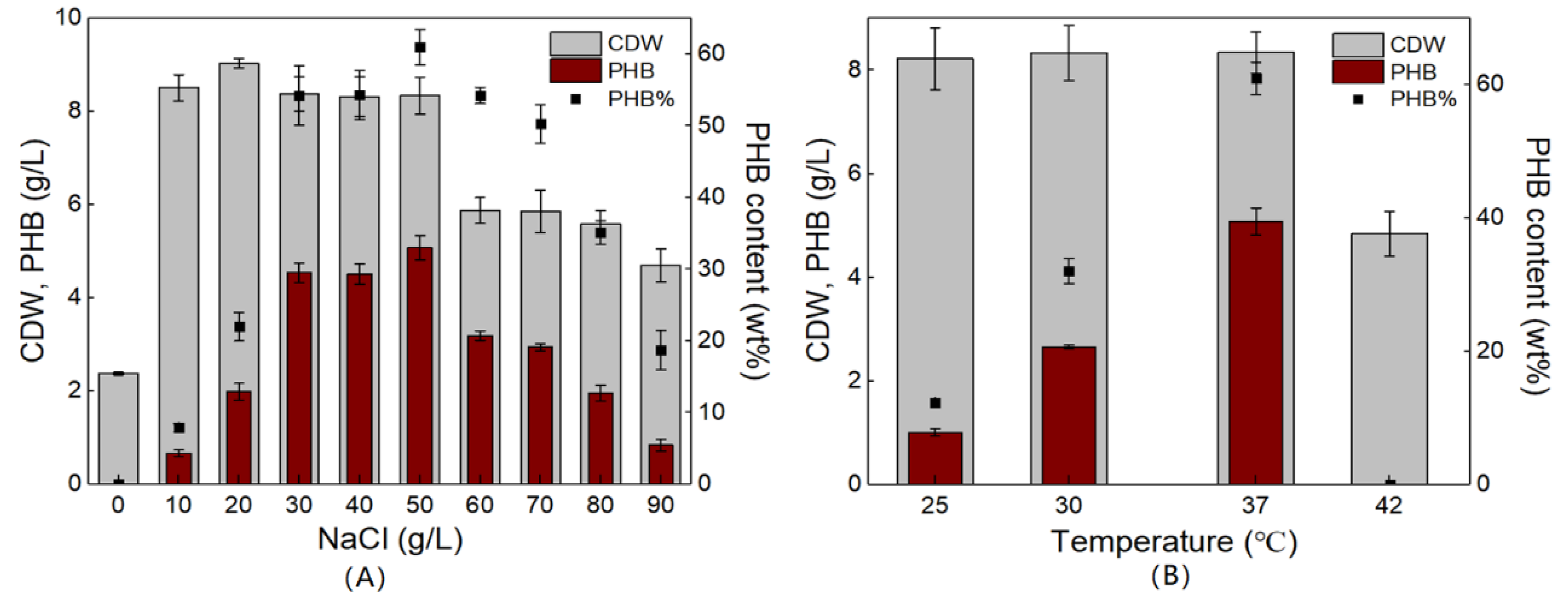
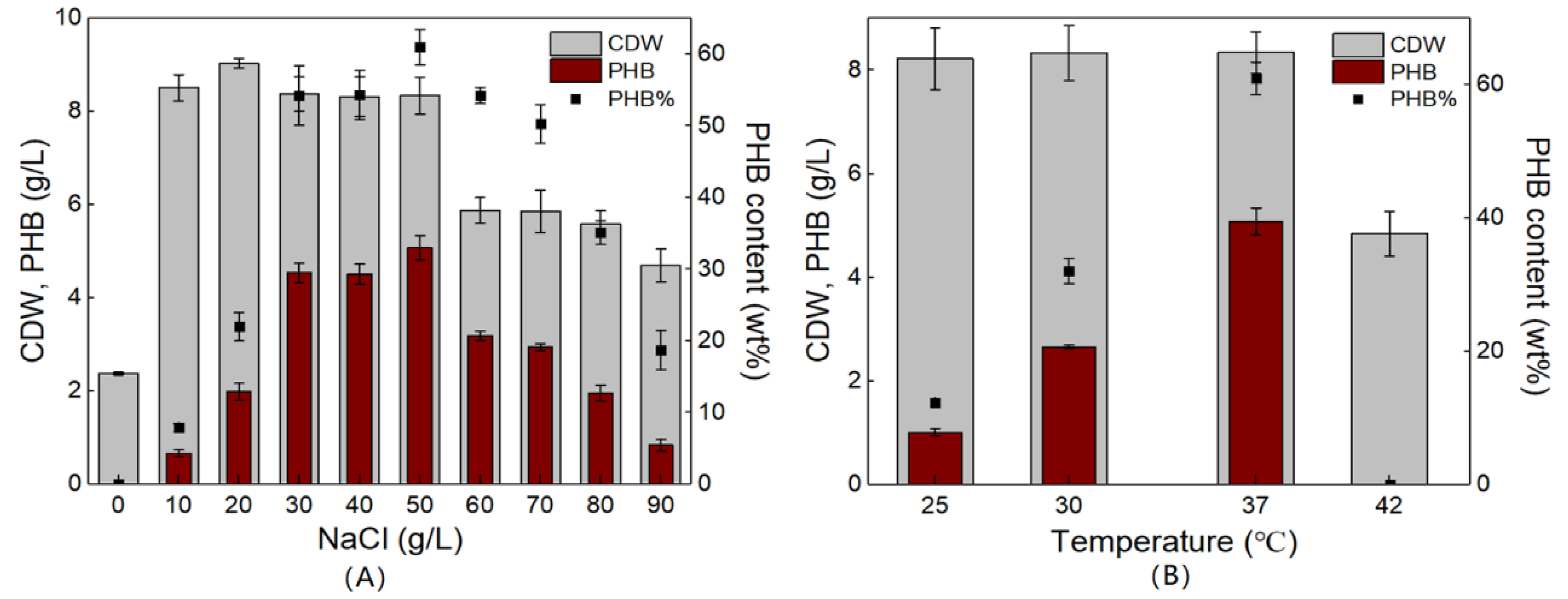
3.3. Optimization of Culture Conditions for PHB Production
3.4. PHBV Production by V. alginolyticus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bedade, D.K.; Edson, C.B.; Gross, R.A. Emergent approaches to efficient and sustainable polyhydroxyalkanoate production. Molecules 2021, 26, 3463. [Google Scholar] [CrossRef]
- Zimmermann, L.; Dombrowski, A.; Volker, C.; Wagner, M. Are bioplastics and plant-based materials safer than conventional plastics? In vitro toxicity and chemical composition. Environ. Int. 2020, 145, 106066. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wilkins, M.R. Recent advances in polyhydroxyalkanoate production: Feedstocks, strains and process developments. Int. J. Biol. Macromol. 2020, 156, 691–703. [Google Scholar] [CrossRef]
- Meereboer, K.W.; Misra, M.; Mohanty, A.K. Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites. Green Chem. 2020, 22, 5519–5558. [Google Scholar] [CrossRef]
- Javaid, H.; Nawaz, A.; Riaz, N.; Mukhtar, H.; Ul-Haq, I.; Shah, K.A.; Khan, H.; Naqvi, S.M.; Shakoor, S.; Rasool, A.; et al. Biosynthesis of polyhydroxyalkanoates (PHAs) by the valorization of biomass and synthetic waste. Molecules 2020, 25, 5539. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-L.; Song, H.-S.; Choi, T.-R.; Lee, S.M.; Park, S.L.; Lee, H.S.; Kim, H.-J.; Bhatia, S.K.; Gurav, R.; Park, K.; et al. Revealing of sugar utilization systems in Halomonas sp. YLGW01 and application for poly(3-hydroxybutyrate) production with low-cost medium and easy recovery. Int. J. Biol. Macromol. 2021, 167, 151–159. [Google Scholar] [CrossRef]
- Fu, X.Z.; Tan, D.; Aibaidula, G.; Wu, Q.; Chen, J.C.; Chen, G.Q. Development of Halomonas TD01 as a host for open production of chemicals. Metab. Eng. 2014, 23, 78–91. [Google Scholar] [CrossRef]
- Shang, L.; Yim, S.C.; Park, H.G.; Chang, H.N. Sequential feeding of glucose and valerate in a fed-batch culture of Ralstonia eutropha for production of poly(hydroxybutyrate-co-hydroxyvalerate) with high 3-hydroxyvalerate fraction. Biotechnol. Prog. 2004, 20, 140–144. [Google Scholar] [CrossRef]
- Liu, H.; Kumar, V.; Jia, L.; Sarsaiya, S.; Kumar, D.; Juneja, A.; Zhang, Z.; Sindhu, R.; Binod, P.; Bhatia, S.K.; et al. Biopolymer poly-hydroxyalkanoates (PHA) production from apple industrial waste residues: A review. Chemosphere 2021, 284, 131427. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Otari, S.V.; Jeon, J.-M.; Gurav, R.; Choi, Y.-K.; Bhatia, R.K.; Pugazhendhi, A.; Kumar, V.; Banu, J.R.; Yoon, J.-J.; et al. Biowaste-to-bioplastic (polyhydroxyalkanoates): Conversion technologies, strategies, challenges, and perspective. Bioresour. Technol. 2021, 19, 326. [Google Scholar]
- Mitra, R.; Xu, T.; Xiang, H.; Han, J. Current developments on polyhydroxyalkanoates synthesis by using halophiles as a promising cell factory. Microb. Cell Factories 2020, 19, 86. [Google Scholar] [CrossRef]
- Thuoc, D.V.; My, D.N.; Loan, T.T.; Sudesh, K. Utilization of waste fish oil and glycerol as carbon sources for polyhydroxyalkanoate production by Salinivibrio sp. M318. Int. J. Biol. Macromol. 2019, 141, 885–892. [Google Scholar] [CrossRef]
- Tan, D.; Wu, Q.; Chen, J.C.; Chen, G.Q. Engineering Halomonas TD01 for the low-cost production of polyhydroxyalkanoates. Metab. Eng. 2014, 26, 34–47. [Google Scholar] [CrossRef]
- Zheng, Y.; Chen, J.C.; Ma, Y.M.; Chen, G.Q. Engineering biosynthesis of polyhydroxyalkanoates (PHA) for diversity and cost reduction. Metab. Eng. 2019, 58, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Shen, R.; Ning, Z.Y.; Lan, Y.X.; Chen, J.C.; Chen, G.Q. Manipulation of polyhydroxyalkanoate granular sizes in Halomonas bluephagenesis. Metab. Eng. 2019, 54, 117–126. [Google Scholar] [CrossRef]
- Jiang, X.R.; Yao, Z.H.; Chen, G.Q. Controlling cell volume for efficient PHB production by Halomonas. Metab. Eng. 2017, 44, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, S.; Yamadaa, M.; Matsumoto, K.; Tajima, K.; Satoh, Y.; Munekata, M.; Ohno, K.; Kohda, K.; Shimamura, T.; Kambe, H.; et al. A microbial factory for lactate-based polyesters using a lactate-polymerizing enzyme. Proc. Natl. Acad. Sci. USA 2008, 105, 17323–17327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.Y.; Park, S.J.; Kim, W.J.; Yang, J.E.; Lee, H.; Shin, J.; Lee, S.Y. One-step fermentative production of poly(lactate-co-glycolate) from carbohydrates in Escherichia coli. Nat. Biotechnol. 2016, 34, 435–440. [Google Scholar] [CrossRef]
- Yang, J.E.; Park, S.J.; Kim, W.J.; Kim, H.J.; Kim, B.J.; Lee, H.; Shin, J.; Lee, S.Y. One-step fermentative production of aromatic polyesters from glucose by metabolically engineered Escherichia coli strains. Nat. Commun. 2018, 9, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukjancenko, O.; Ussery, D. Vibrio chromosome-specific families. Front. Microbiol. 2014, 5, 73. [Google Scholar] [CrossRef]
- Sun, W.; Cao, J.G.; Teng, K.; Meighen, E.A. Biosynthesis of poly-3-hydroxybutyrate in the luminescent bacterium, Vibrio harveyi, and regulation by the lux autoinducer, N-(3-hydroxybutanoyl) homoserine lactone. J. Biol. Chem. 1994, 269, 20785–20790. [Google Scholar] [CrossRef]
- Mohandas, S.P.; Balan, L.; Lekshmi, N.; Cubelio, S.S.; Philip, R.; Bright, S.I.S. Production and characterization of polyhydroxybutyrate from Vibrio harveyi MCCB 284 utilizing glycerol as carbon source. J. Appl. Microbiol. 2017, 122, 698–707. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.C.; Chen, C.C.; Choi, M.H.; Kung, S.S.; Wei, Y.H. Production of poly-β-hydroxybutyrate (PHB) by Vibrio spp. isolated from marine environment. J. Biotechnol. 2007, 132, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Sasidharan, R.S.; Bhat, S.G.; Chandrasekaran, M. Biocompatible polyhydroxybutyrate (PHB) production by marine Vibrio azureus BTKB33 under submerged fermentation. Ann. Microbiol. 2014, 65, 455–465. [Google Scholar] [CrossRef]
- Boyandin, A.; Kalacheva, G.S.; Medvedeva, S.; Rodicheva, E.; Volova, T.G. Luminous bacteria as producers of polyhydroxyalkanoates. Macromol. Symp. 2008, 269, 17–22. [Google Scholar] [CrossRef]
- Hong, J.W.; Song, H.S.; Moon, Y.M.; Hong, Y.G.; Bhatia, S.K.; Jung, H.R.; Choi, T.R.; Yang, S.Y.; Park, H.Y.; Choi, Y.K.; et al. Polyhydroxybutyrate production in halophilic marine bacteria Vibrio proteolyticus isolated from the Korean peninsula. Bioprocess Biosyst. Eng. 2019, 42, 603–610. [Google Scholar] [CrossRef]
- de Vogel, F.A.; Schlundt, C.; Stote, R.E.; Ratto, J.A.; Amaral-Zettler, L.A. Comparative genomics of marine bacteria from a historically defined plastic biodegradation consortium with the capacity to biodegrade polyhydroxyalkanoates. Microorganisms 2021, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Eagon, R.G. Pseudomonas natriegens, a marine bacterium with a generation time of less than 10 min. J. Bacteriol. 1962, 83, 736–737. [Google Scholar]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Balcazar, J.L.; Gallo-Bueno, A.; Planas, M.; Pintado, J. Isolation of Vibrio alginolyticus and Vibrio splendidus from captive-bred seahorses with disease symptoms. Antonie Leeuwenhoek 2010, 97, 207–210. [Google Scholar] [CrossRef]
- Tao, G.B.; Tan, B.W.; Li, Z.J. Production of polyhydroxyalkanoates by a moderately halophilic bacterium of Salinivibrio sp. TGB10. Int. J. Biol. Macromol. 2021, 186, 574–579. [Google Scholar] [CrossRef]
- Ilham, M.; Nakanomori, S.; Kihara, T.; Hokamura, A.; Matsusaki, H.; Tsuge, T.; Mizuno, K. Characterization of polyhydroxyalkanoate synthases from Halomonas sp. O-1 and Halomonas elongata DSM2581: Site-directed mutagenesis and recombinant expression. Polym. Degrad. Stabil. 2014, 109, 416–423. [Google Scholar] [CrossRef]
- Tsuge, T. Fundamental factors determining the molecular weight of polyhydroxyalkanoate during biosynthesis. Polym. J. 2016, 48, 1051–1057. [Google Scholar] [CrossRef]
- Sim, S.J.; Snell, K.D.; Hogan, S.A.; Stubbe, J.; Rha, C.; Sinskey, A.J. PHA synthase activity controls the molecular weight and polydispersity of polyhydroxybutyrate in vivo. Nat. Biotechnol. 1997, 15, 63–67. [Google Scholar] [CrossRef]
- Meereboer, K.W.; Pal, A.K.; Cisneros-Lopez, E.O.; Misra, M.; Mohanty, A.K. The effect of natural fillers on the marine biodegradation behaviour of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) (PHBV). Sci. Rep. 2021, 11, 911. [Google Scholar] [CrossRef] [PubMed]
- Griffitt, K.J.; Grimes, D.J. Abundance and distribution of Vibrio cholerae, V. parahaemolyticus, and V. vulnificus following a major freshwater intrusion into the Mississippi sound. Microb. Ecol. 2013, 65, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Numata, K.; Doi, Y. Biosynthesis of polyhydroxyalkanaotes by a novel facultatively anaerobic Vibrio sp. under marine conditions. Mar. Biotechnol. 2012, 14, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, R.Y.U.; Castilho, N.A.S.; Silva, M.; Miotto, M.C.; Lima, A.O.S. Prospecting for marine bacteria for polyhydroxyalkanoate production on low-cost bubstrates. Bioengineering 2017, 4, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoff, J.; Daniel, B.; Stukenberg, D.; Thuronyi, B.W.; Waldminghaus, T.; Fritz, G. Vibrio natriegens: An ultrafast-growing marine bacterium as emerging synthetic biology chassis. Environ. Microbiol. 2020, 22, 4394–4408. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics (μg/mL) | 10 | 25 | 50 | 80 | 100 | 150 | |
|---|---|---|---|---|---|---|---|
| V. alginolyticus LHF01 | Chloramphenicol | - | - | - | - | - | - |
| Ampicillin | ++ | ++ | ++ | ++ | ++ | ++ | |
| Kanamycin | ++ | ++ | ++ | + | + | - | |
| Spectinomycin | ++ | + | + | - | - | - | |
| V. alginolyticus LHF02 | Chloramphenicol | + | - | - | - | - | - |
| Ampicillin | ++ | ++ | ++ | ++ | ++ | ++ | |
| Kanamycin | ++ | ++ | + | + | - | - | |
| Spectinomycin | ++ | + | + | - | - | - |
| Strain | Carbon Source | Mw (×103 Da) | Mn (×103 Da) | Mw/Mn |
|---|---|---|---|---|
| V. alginolyticus LHF01 | glycerol | 201 | 90 | 2.2 |
| V. alginolyticus LHF02 | glycerol | 1380 | 323 | 4.3 |
| Carbon Source | CDW (g/L) | PHA Content (wt%) | PHA (g/L) | 3HB (mol%) | 3HV (mol%) |
|---|---|---|---|---|---|
| Gly + PA(2) * | 8.47 ± 0.32 | 13.48 ± 0.35 | 1.14 ± 0.02 | 100 | ND |
| Gly + PA(4) | 8.55 ± 0.43 | 16.61 ± 0.49 | 1.44 ± 0.16 | 75.58 ± 0.25 | 24.42 ± 0.25 |
| Gly + PA(6) | 1.83 ± 0.06 | ND | ND | ND | ND |
| Microorganism | Carbon Source | PHAs | CDW (g/L) | PHA Content (wt%) | PHA Titer (g/L) | Reference |
|---|---|---|---|---|---|---|
| V. harveyi | Glycerol | PHB | (OD660) 4.2 | 2.6 | — | [21] |
| V. harveyiMCCB 284 | Glycerol | PHB | 3.2 | 72 | 2.3 | [22] |
| Vibrio sp. M11 | Glycerol | PHB | (OD600) 12.2 | 30.2 | — | [23] |
| V. azureus BTKB33 | Glucose | PHB | 1.12 | 42.69 | 0.48 | [24] |
| V. fischeri 1231 | Glycerol | PHB | — | 0.4 | 0.003 | [25] |
| V. proteolyticus | Fructose | PHB | 4.94 | 54.7 | 2.7 | [26] |
| V. proteolyticus | Fructose+ Propionate | P(3HB-co-15.8% 3HV) | — | — | — | [26] |
| Vibrio sp. KN01 | Soybean oil | PHB | 2.4 | 8 | 0.19 | [37] |
| Vibrio sp. KN01 | Soybean oil | P(3HB-co-14%3HP-co-3%5HV) | 0.4 | 40 | 0.16 | [37] |
| V. alginolyticus LHF01 | Glycerol | PHB | 8.34 | 60.97 | 5.08 | This study |
| V. alginolyticus LHF01 | Glycerol+ Propionate | P(3HB-co-24.42% 3HV) | 8.55 | 16.11 | 1.44 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-F.; Wang, M.-R.; Tian, L.-Y.; Li, Z.-J. Production of Polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus Strains Isolated from Salt Fields. Molecules 2021, 26, 6283. https://doi.org/10.3390/molecules26206283
Li H-F, Wang M-R, Tian L-Y, Li Z-J. Production of Polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus Strains Isolated from Salt Fields. Molecules. 2021; 26(20):6283. https://doi.org/10.3390/molecules26206283
Chicago/Turabian StyleLi, Hong-Fei, Meng-Ru Wang, Lin-Yue Tian, and Zheng-Jun Li. 2021. "Production of Polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus Strains Isolated from Salt Fields" Molecules 26, no. 20: 6283. https://doi.org/10.3390/molecules26206283
APA StyleLi, H.-F., Wang, M.-R., Tian, L.-Y., & Li, Z.-J. (2021). Production of Polyhydroxyalkanoates (PHAs) by Vibrio alginolyticus Strains Isolated from Salt Fields. Molecules, 26(20), 6283. https://doi.org/10.3390/molecules26206283