
p-Aminobenzoate Organic Salts as Potential Plant Growth Regulators for Tomatoes

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion
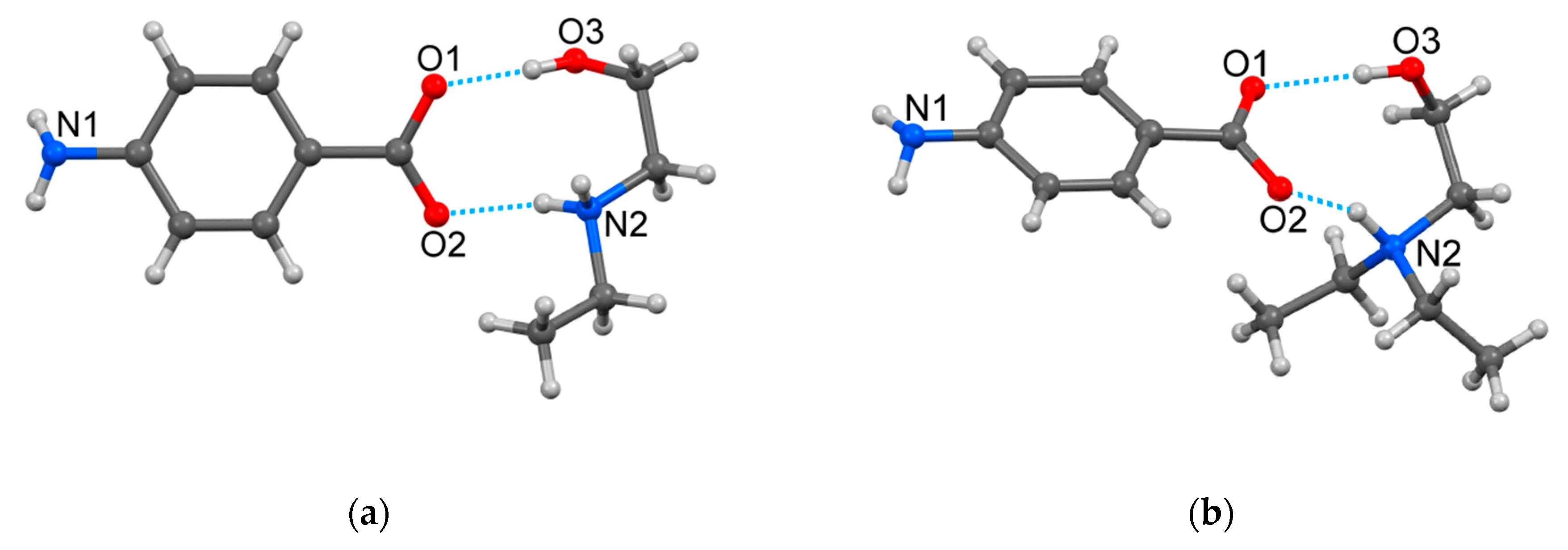
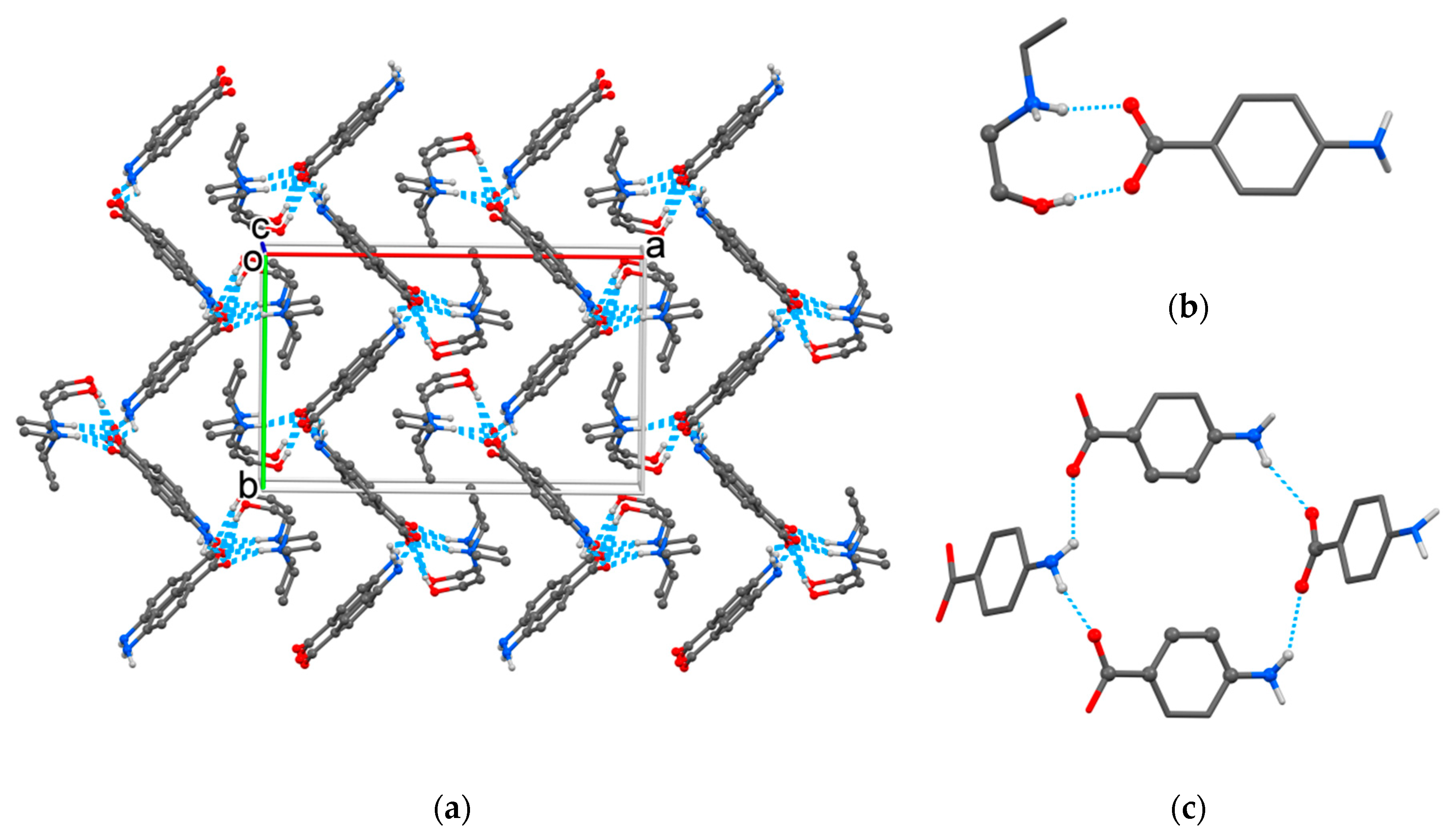
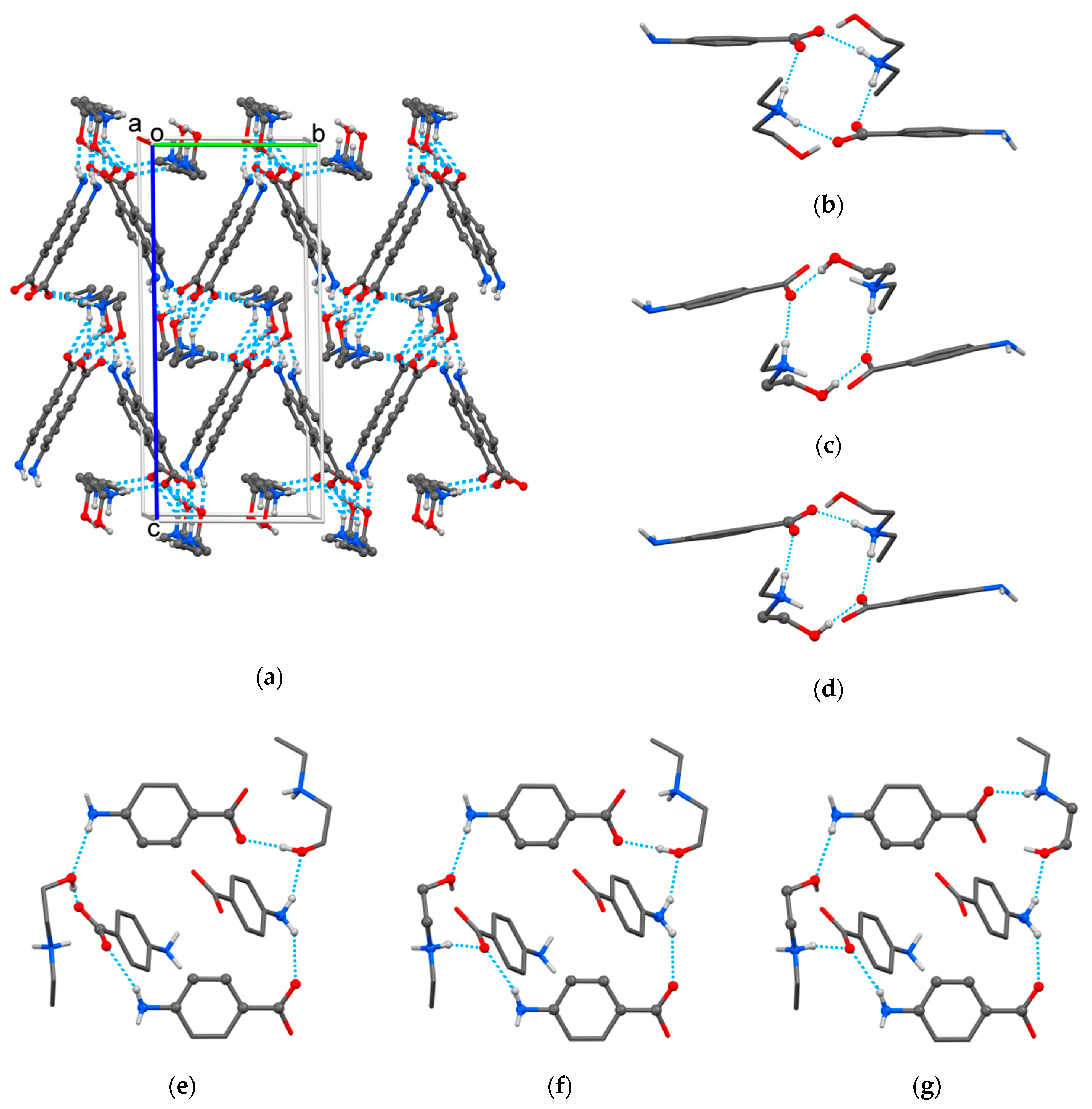
2.1. Crystallographic Study
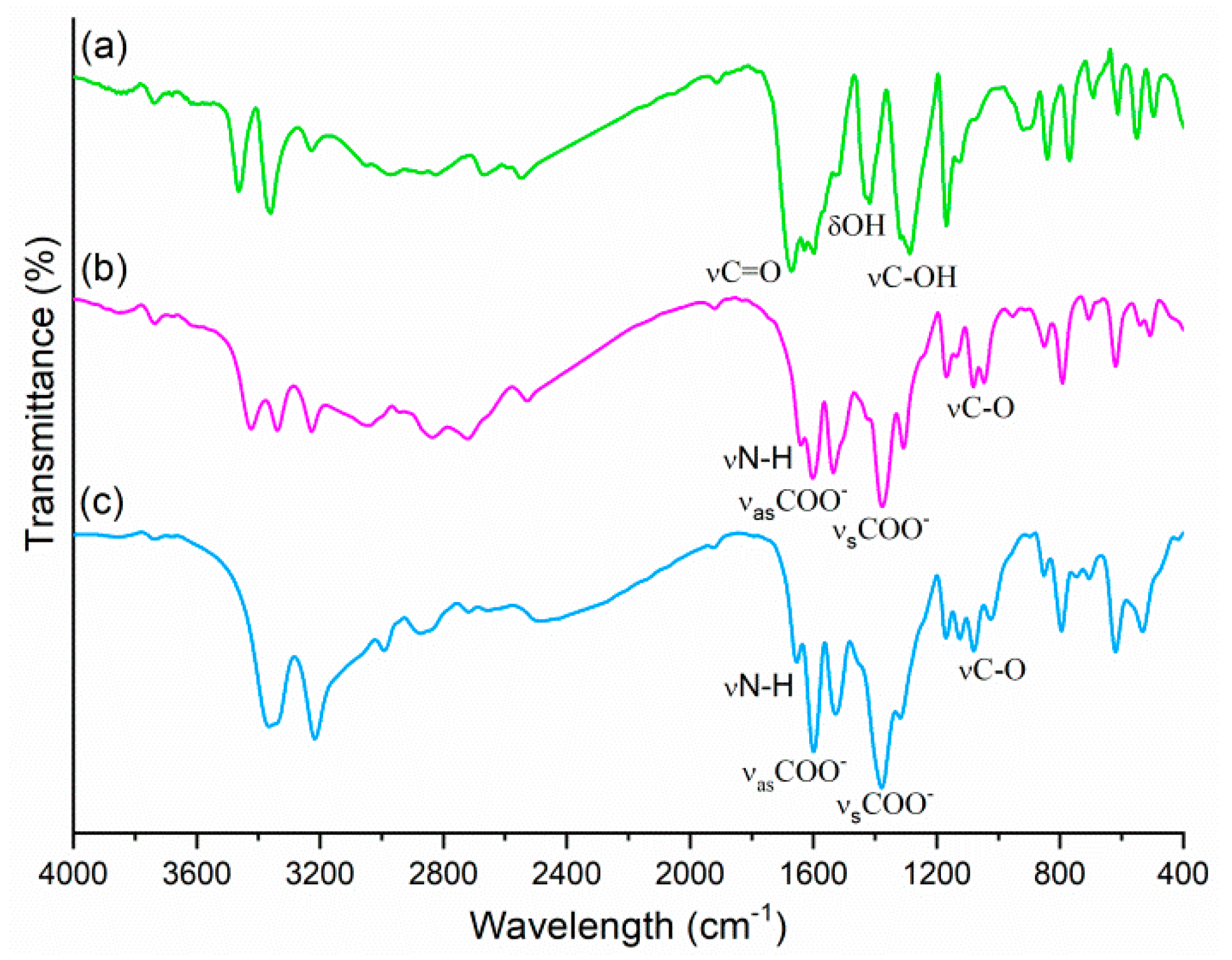
2.2. FTIR Spectroscopy
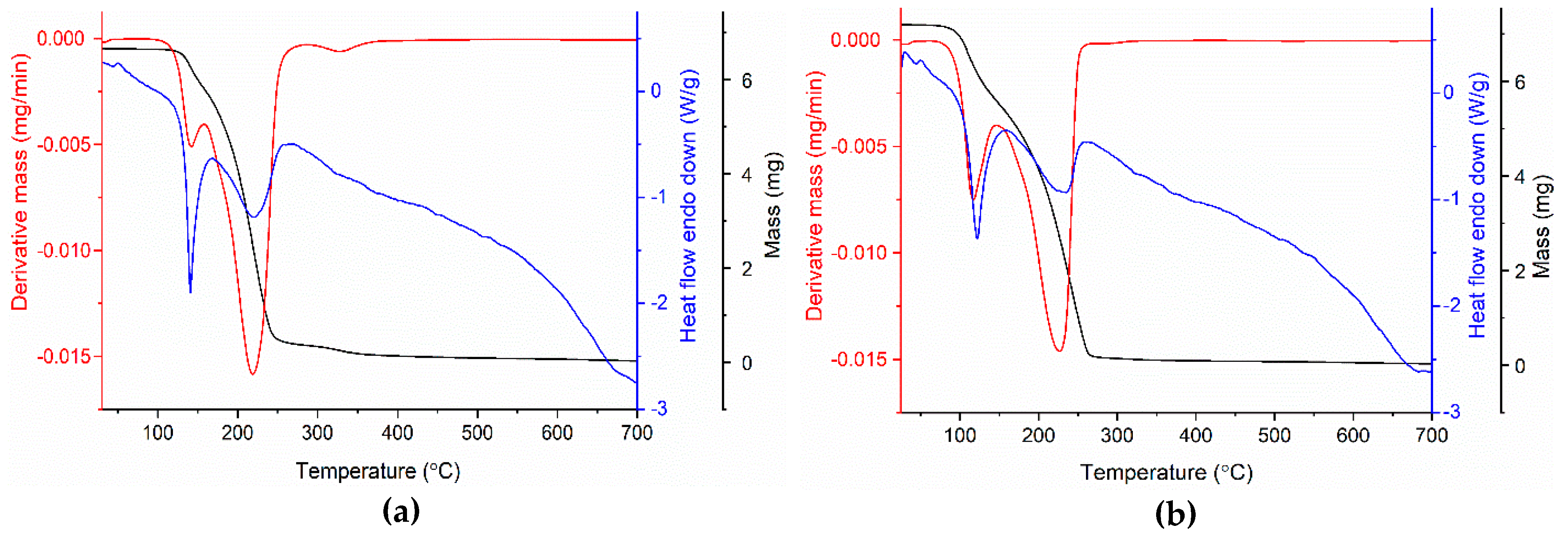
2.3. Thermal Analysis
2.4. Biological Studies
3. Materials and Methods
3.1. Materials and Physical Measurements
3.2. Synthesis Procedure of HEEA-pABA and HDEEA-pABA
3.3. Single Crystal X-ray Diffraction Study
3.4. Biological Assays
3.4.1. Germination Tests (Experiment 1)
3.4.2. Seedling Growth Tests (Experiment 2)
3.4.3. Tests on Tomato Plants (Experiment 3)
3.4.4. Data and Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Song, Y. Insight into the mode of action of 2,4-dichlorophenoxyacetic acid (2,4-D) as an herbicide. J. Integr. Plant Biol. 2014, 56, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Small, C.C.; Degenhardt, D. Plant growth regulators for enhancing revegetation success in reclamation: A review. Ecol. Eng. 2018, 118, 43–51. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G. Plant growth regulators I: Introduction; auxins, their analogues and inhibitors. In Plant Propagation by Tissue Culture, 3rd ed.; George, E.F., Hall, M.A., Klerk, G.J.D., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 175–204. [Google Scholar] [CrossRef]
- Paque, S.; Weijers, D. Q&A: Auxin: The plant molecule that influences almost anything. BMC Biol. 2016, 14, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Ng, P.L.L.; Ferrarese, M.L.L.; Huber, D.A.; Ravagnani, A.L.S.; Ferrarese-Filho, O. Canola (Brassica napus L.) seed germination influenced by cinnamic and benzoic acids and derivatives: Effects on peroxidase. Seed Sci. Technol. 2003, 31, 39–46. [Google Scholar] [CrossRef]
- Crisan, M.E.; Bourosh, P.; Maffei, M.E.; Forni, A.; Pieraccini, S.; Sironi, M.; Chumakov, Y.M. Synthesis, crystal structure and biological activity of 2-hydroxyethylammonium salt of p-aminobenzoic acid. PLoS ONE 2014, 9, e101892. [Google Scholar] [CrossRef]
- Grozav, M.; Neamtiu, I.; Dorosencu, M.; Laichici, M.; Mercea, M. The synthesis of some ammonium salts of benzoic acids with etanolamine, possible plant growth stimulators. Rev. Chim. 2003, 54, 287–288. [Google Scholar]
- Dempsey, D.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23–33. [Google Scholar] [CrossRef]
- Senaratna, T.; Merritt, D.; Dixon, K.; Bunn, E.; Touchell, D.; Sivasithamparam, K. Benzoic acid may act as the functional group in salicylic acid and derivatives in the induction of multiple stress tolerance in plants. Plant Growth Regul. 2003, 39, 77–81. [Google Scholar] [CrossRef]
- Basset, G.J.C.; Quinlivan, E.P.; Ravanel, S.; Rebeille, F.; Nichols, B.P.; Shinozaki, K.; Seki, M.; Adams-Phillips, L.C.; Giovannoni, J.J.; Gregory, J.F.; et al. Folate synthesis in plants: The p-aminobenzoate branch is initiated by a bifunctional PabA-PabB protein that is targeted to plastids. Proc. Natl. Acad. Sci. USA 2004, 101, 1496–1501. [Google Scholar] [CrossRef]
- Marbois, B.; Xie, L.X.; Choi, S.; Hirano, K.; Hyman, K.; Clarke, C.F. para-Aminobenzoic acid is a precursor in Coenzyme Q6 biosynthesis in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 27827–27838. [Google Scholar] [CrossRef]
- Galbinur, T.; Obolensky, A.; Berenshtein, E.; Vinokur, V.; Chowers, I.; Chevion, M.; Banin, E. Effect of para-aminobenzoic acid on the course of retinal degeneration in the rd10 mouse. J. Ocul. Pharmacol. Ther. 2009, 25, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.M.R.; Xing, D.K.L.; King, T.P. Activity of p-aminobenzoic acid compared with other organic acids against selected bacteria. J. Appl. Bacteriol. 1995, 78, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Vasilieva, S. Para-aminobenzoic acid inhibits a set of SOS functions in Escherichia coli K12. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2001, 496, 89–95. [Google Scholar] [CrossRef]
- Drozd, N.N.; Makarov, V.A.; Miftakhova, N.T.; Kalugin, S.A.; Stroeva, O.G.; Akberova, S.I. Antithrombotic activity of para-aminobenzoic acid. Eksp. Klin. Farmakol. 2000, 63, 40–44. [Google Scholar] [PubMed]
- Andreenko, G.V.; Karabasova, M.A.; Liutova, L.A.; Podorolskaia, L.V.; Serebriakova, T.N.; Sologub, A.A.; Akberova, S.I.; Stroeva, O.G. Effect of para-aminobenzoic acid on the fibrinolytic activity of blood. Dokl. Akad. Nauk 1996, 346, 268–270. [Google Scholar] [PubMed]
- Hanson, K.M.; Gratton, E.; Bardeen, C.J. Sunscreen enhancement of UV-induced reactive oxygen species in the skin. Free Radic. Biol. Med. 2006, 41, 1205–1212. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Kong, X.; Lu, Z.; Xiao, M.; Chen, M.; Zhu, L.; Shen, Y.; Hu, X.; Song, S. para-Aminobenzoic acid (PABA) synthase enhances thermotolerance of mushroom Agaricus bisporus. PLoS ONE 2014, 9, e91298. [Google Scholar] [CrossRef]
- Song, G.C.; Choi, H.K.; Ryu, C.M. The folate precursor para-aminobenzoic acid elicits induced resistance against Cucumber mosaic virus and Xanthomonas axonopodis. Ann. Bot. 2013, 111, 925–934. [Google Scholar] [CrossRef]
- Groom, C.R.; Bruno, I.J.; Lightfoot, M.P.; Ward, S.C. The Cambridge structural database. Acta Crystallogr. B 2016, 72, 171–179. [Google Scholar] [CrossRef]
- Crisan, M.E.; Gorobet, A.; Siminel, A.V.; Bourosh, P.N.; Croitor, L. A new supramolecular isomer of p-aminobenzoate Zn (II) coordination polymer: Structure and photoluminescent property. Polyhedron 2019, 171, 502–507. [Google Scholar] [CrossRef]
- Chicu, S.A.; Herrmann, K.; Berking, S. An approach to calculate the toxicity of simple organic molecules on the basis of QSAR analysis in Hydractinia echinata (Hydrozoa, Cnidaria). Quant. Struct. Act. Relat. 2000, 19, 227–236. [Google Scholar] [CrossRef]
- Croitor, L.; Petric, M.F.; Szerb, E.I.; Vlase, G.; Bourosh, P.N.; Chumakov, Y.; Crisan, M.E. Role of 4-nitrobenzoic acid polymorphs in the crystallization process of the organic acid-base multicomponent system. CrystEngComm 2019, 21, 6038–6047. [Google Scholar] [CrossRef]
- Crisan, M.; Vlase, G.; Plesu, N.; Petric, M.; Croitor, L.; Kravtsov, V.; Chumakov, Y.; Bouros, P.; Vlase, T. Ethylethanolammonium 4-nitrobenzoate: Synthesis, structural characterization, thermal analysis, non-isothermal kinetic investigations and corrosion inhibitor efficiency. J. Therm. Anal. Calorim. 2018, 134, 343–352. [Google Scholar] [CrossRef]
- Crisan, M.; Halip, L.; Bourosh, P.; Chicu, S.A.; Chumakov, Y. Synthesis, structure and toxicity evaluation of ethanolamine nitro/chloronitrobenzoates: A combined experimental and theoretical study. Chem. Cent. J. 2017, 11, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Chicu, S.A.; Grozav, M.; Kurunczi, L.; Crisan, M. SAR for amine salts of carboxylic acids to Hydractinia echinata. Rev. Chim. 2008, 59, 582–587. [Google Scholar] [CrossRef]
- Crisan, M.; Vlase, G.; Szerb, E.I.; Vlase, T. Thermal and kinetics studies of primary, secondary and tertiary alkanolammonium salts of 4-nitrobenzoic acid. J. Therm. Anal. Calorim. 2018, 132, 1409–1418. [Google Scholar] [CrossRef]
- Cruz-Cabeza, A.J. Acid–base crystalline complexes and the pKa rule. CrystEngComm 2012, 14, 6362–6365. [Google Scholar] [CrossRef]
- Pavia, D.L.; Lampman, G.M.; Kriz, G.S.; Vyvyan, J.R. Introduction to Spectroscopy, 4th ed.; Brooks/Cole, Cengage Learning: Boston, MA, USA; Belmont, CA, USA, 2009; p. 80. [Google Scholar]
- Liu, X.; Zhang, H.; Zhao, Y.; Feng, Z.; Li, Q.; Yang, H.Q.; Luan, S.; Li, J.; He, Z.H. Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF-mediated ABI3 activation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2013, 110, 15485–15490. [Google Scholar] [CrossRef]
- Ye, N.; Zhu, G.; Liu, Y.; Zhang, A.; Li, Y.; Liu, R.; Shi, L.; Jia, L. Ascorbic acid and reactive oxygen species are involved in the inhibition of seed germination by Abscisic acid in rice seeds. J. Exp. Bot. 2012, 63, 1809–1822. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Plant hormones and seed germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Shuai, H.; Meng, Y.; Luo, X.; Chen, F.; Zhou, W.; Dai, Y.; Qi, Y.; Du, J.; Yang, F.; Liu, J.; et al. Exogenous auxin represses soybean seed germination through decreasing the gibberellin/abscisic acid (GA/ABA) ratio. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tsygankova, V.; Andrusevich, Y.; Shtompel, O.; Myroljubov, O.; Hurenko, A.; Solomyanny, R.; Mrug, G.; Frasinyuk, M.; Shablyn, O.; Brovarets, V. Study of auxin, cytokinetin and gibberellins -like activity of heterocyclic compounds derivatives of pyrimidine, pyridine, pyrazole and isoflavones. Eur. J. Biotechnol. Biosci. 2016, 4, 29–44. [Google Scholar]
- Bethke, P.C.; Libourel, I.G.; Jones, R.L. Nitric oxide reduces seed dormancy in Arabidopsis. J. Exp. Bot. 2006, 57, 517–526. [Google Scholar] [CrossRef]
- Oracz, K.; Karpiński, S. Phytohormones signaling pathways and ROS involvement in seed germination. Front. Plant Sci. 2016, 7, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trends Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Reichheld, J.P.; Foyer, C.H. ROS related redox regulation and signaling in plants. Semin. Cell Dev. Biol. 2017, 80, 3–12. [Google Scholar] [CrossRef]
- Dash, M.; Yordanov, Y.S.; Georgieva, T.; Tschaplinski, T.J.; Yordanova, E.; Busov, V. Poplar PtabZIP1-like enhances lateral root formation and biomass growth under drought stress. Plant J. 2017, 89, 692–705. [Google Scholar] [CrossRef]
- Qin, H.; Huang, R. Auxin controlled by ethylene steers root development. Int. J. Mol. Sci. 2018, 19, 3656. [Google Scholar] [CrossRef]
- Du, Y.; Scheres, B. Lateral root formation and the multiple roles of auxin. J. Exp. Bot. 2018, 69, 155–167. [Google Scholar] [CrossRef]
- Cai, X.T.; Xu, P.; Zhao, P.X.; Liu, R.; Yu, L.H.; Xiang, C.B. Arabidopsis ERF109 mediates cross-talk between jasmonic acid and auxin biosynthesis during lateral root formation. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef]
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Xu, N.; Chen, H.; Wang, G.; Huang, J. OsMADS25 regulates root system development via auxin signalling in rice. Plant J. 2018, 95, 1004–1022. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.; Yang, X.; Zhang, J.; Liu, X.; Zheng, H.; Dong, G.; Nian, J.; Feng, J.; Xia, B.; Qian, Q.; et al. Peptidyl-prolyl isomerization targets rice Aux/IAAs for proteasomal degradation during auxin signalling. Nat. Commun. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Reed, R.C.; Brady, S.R.; Muday, G.K. Inhibition of auxin movement from the shoot into the root inhibits lateral root development in Arabidopsis. Plant Physiol. 1998, 118, 1369–1378. [Google Scholar] [CrossRef] [PubMed]
- Dao, G.H.; Wu, G.X.; Wang, X.X.; Zhuang, L.L.; Zhang, T.Y.; Hu, H.Y. Enhanced growth and fatty acid accumulation of microalgae Scenedesmus sp. LX1 by two types of auxin. Bioresour. Technol. 2018, 247, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, N.; Hou, J.; Xu, L.; Liu, C.; Zhang, J.; Wang, Q.; Zhang, X.; Wu, X. Factors influencing leaf chlorophyll content in natural forests at the biome scale. Front. Ecol. Evol. 2018, 6, 1–10. [Google Scholar] [CrossRef]
- Beinsan, C.; Camen, D.; Sumalan, R.; Babau, M. Study concerning salt stress effect on leaf area dynamics and chlorophyll content in four bean local landraces from Banat area. In Proceedings of the 44th Croatian and 4th International Symposium on Agriculture, Opatija, Croatia, 16–20 February 2009; pp. 416–419. [Google Scholar]
- El-Saeid, H.M.; Abou-Hussein, S.D.; El-Tohamy, W.A. Growth characters, yield and endogenous hormones of cowpea plants in response to IAA application. Res. J. Agric. Biol. Sci. 2010, 6, 27–31. [Google Scholar]
- Finet, C.; Jaillais, Y. Auxology: When auxin meets plant evo-devo. Dev. Biol. 2012, 369, 19–31. [Google Scholar] [CrossRef]
- Khalil, S.; Mandurah, H.M. Growth and metabolic changes of cowpeas plants as affected by water deficiency and indole acetic acid. J. Agron. Crop Sci. 1989, 16, 160–166. [Google Scholar] [CrossRef]
- El-Mergawi, R.A. Sensitivity of faba bean cultivars to low glyphosate doses and the efficiency of IAA as indicator to glyphosate effects. Egypt J. Hort. 2003, 30, 197–214. [Google Scholar]
- Gaspar, T.H.; Kevers, C.; Faivre-Rampant, O.; Crèvecour, M.; Penel, C.L.; Greppin, H.; Dommes, J. Changing concepts in plant hormone action. Vitro Cell. Dev. Biol. Plant 2003, 39, 85–106. [Google Scholar] [CrossRef]
- Alarcón, M.V.; Salguero, J.; Lloret, P.G. Auxin modulated initiation of lateral roots is linked to pericycle cell length in maize. Front. Plant Sci. 2019, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Jia, B.; Liu, H.; Kan, X.; Zhang, Y.; Zhou, R.; Li, Z.; Yang, L.; Deng, D.; Yin, Z. Genetic mapping of the leaf number above the primary ear and its relationship with plant height and flowering time in maize. Front. Plant Sci. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Shaver, G.R. Mineral nutrition and leaf longevity in Ledum palustre: The role of individual nutrients and the timing of leaf mortality. Oecologia 1983, 56, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Mauro, P.R.; Agnello, M.; Distefano, M.; Sabatino, L.; san Bautista, P.A.; Leonardi, C.; Giuffrida, F. Chlorophyll fluorescence, photosynthesis and growth of tomato plants as affected by long-term oxygen root zone deprivation and grafting. Agronomy 2020, 10, 137. [Google Scholar] [CrossRef]
- Sheldrick, G.M. Crystal structure refinement with SHELXL. Acta Cryst. 2015, C71, 3–8. [Google Scholar] [CrossRef]
- Macrae, C.F.; Edgington, P.R.; McCabe, P.; Pidcock, E.; Shields, G.P.; Taylor, R.; Towler, M.; Streek, J. Mercury: Visualization and analysis of crystal structures. J. Appl. Crystallogr. 2006, 39, 453–457. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | HEEA-pABA | HDEEA-pABA |
|---|---|---|
| Empirical formula | C11H18N2O3 | C13H22N2O3 |
| Fw | 226.27 | 254.32 |
| T (K) | 293(2) | 293(2) |
| Crystal system | Monoclinic | Orthorhombic |
| Space group | P21/c | Pna21 |
| a (Å) | 8.3465(4) | 17.9492(17) |
| b (Å) | 8.1802(5) | 11.1812(9) |
| c (Å) | 18.8676(9) | 7.1445(6) |
| β (°) | 102.684(5) | 90 |
| V (Å3) | 1256.76(11) | 1433.9(2) |
| Z | 4 | 4 |
| ρcalcd (g cm−3) | 1.196 | 1.178 |
| μ (mm−1) | 0.087 | 0.084 |
| Crystal size (mm) | 0.60 × 0.25 × 0.05 | 0.400 × 0.200 × 0.040 |
| 2Θ range (°) | 3.332 to 24.990 | 3.569 to 24.996 |
| Reflections collected/unique | 4344/2199 (Rint = 0.0243) | 3177/1870 (Rint = 0.0417) |
| Reflections with (I > 2 δ(I)) | 1576 | 1059 |
| Parameters | 148 | 167 |
| R1, wR2 (I > 2 δ(I)) | 0.0528, 0.1382 | 0.0609, 0.1259 |
| R1, wR2 (all data) | 0.0774, 0.1564 | 0.1147, 0.1484 |
| GOF [c] | 1.000 | 1.000 |
| Largest diff. peak/hole (e Å−3) | 0.332/−0.306 | 0.178/−0.151 |
| D‒H∙∙∙A | d(D–H) | d(H∙∙∙A) | d(D∙∙∙A) | ∠(DHA) | Symmetry Transformations for Acceptor |
|---|---|---|---|---|---|
| HEEA-pABA | |||||
| O(3)‒H(1)∙∙∙O(1) | 0.82 | 1.83 | 2.650(2) | 178 | x, y, z |
| N(1)‒H(1)∙∙∙O(3) | 0.87 | 2.27 | 3.077(3) | 155 | −x + 1, y + 1/2, −z + 1/2 |
| N(1)‒H(2)∙∙∙O(2) | 0.87 | 2.34 | 3.010(3) | 135 | −x, y + 1/2, −z + 1/2 |
| N(2)‒H(1)∙∙∙O(1) | 0.89 | 1.89 | 2.775(2) | 174 | −x + 1, −y + 1, −z + 1 |
| N(2)‒H(2)∙∙∙O(2) | 0.89 | 1.92 | 2.787(2) | 165 | x, y, z |
| C(8)‒H(8)∙∙∙O(3) | 0.97 | 2.62 | 3.508(3) | 152 | −x + 1, −y, −z + 1 |
| HDEEA-pABA | |||||
| O(3)‒H(3)∙∙∙O(1) | 0.82 | 1.87 | 2.676(5) | 170 | x, y, z |
| N(1)‒H(1)∙∙∙O(2) | 0.87 | 2.09 | 2.908(8) | 158 | −x + 3/2, y + 1/2, z + 1/2 |
| N(1)‒H(2)∙∙∙O(1) | 0.87 | 2.29 | 2.858(9) | 123 | −x + 3/2, y + 1/2, z−1/2 |
| N(2)‒H(2)∙∙∙O(2) | 0.98 | 1.78 | 2.701(6) | 156 | x, y, z |
| C(8)‒H(8)∙∙∙O(3) | 0.97 | 2.54 | 3.501(9) | 172 | −x + 1, −y + 1, z−1/2 |
| C(13)‒H(8)∙∙∙O(2) | 0.96 | 2.60 | 3.285(8) | 129 | x, y, z |
| Moment | After 7 Days | After 10 Days | ||||
|---|---|---|---|---|---|---|
| Treatment | 0.1 mM | 0.5 mM | 1 mM | 0.1 mM | 0.5 mM | 1 mM |
| Control (MES) | 91.11 a | 91.11 a | 91.11 a | 100 a | 100 a | 100 a |
| Indole-3-acetic acid (IAA) | x 8.89 c | x 0 d | x 0 c | x 75.56 b | y 0 b | y 0 b |
| HEEA-pABA | x 46.67 b | y 26.67 c | y 17.78 b | x 88.89 ab | x 93.33 a | x 91.11 a |
| HDEEA-pABA | x 40.00 b | x 44.44 b | y 20.00 b | x 97.78 a | x 93.33 a | x 88.89 a |
| Treatment. LSD5% = 17.68 (a,b,c,d); Concentration. LSD5% = 11.21 (x,y) | Treatment. LSD5% = 16.64 (a,b); Concentration. LSD5% = 17.11 (x,y) | |||||
| Character/ Treatment | Height (cm) | Chlorophyll (SPAD) | Primary Root Length (cm) | No of Secondary Roots | Maximum Length of Secondary Roots (cm) |
|---|---|---|---|---|---|
| Control (MES) | 5.50±0.50 b | 31.20 ± 1.49 c | 11.10 ± 0.37 ab | 25.00 ± 2.99 c | 9.27 ± 0.32 a |
| IAA | 3.20 ± 0.18 c | 32.25 ± 0.25 c | 8.65 ± 0.55 c | 104.50 ± 18.42 a | 3.50 ± 0.55 b |
| HEEA-pABA | 5.25 ± 0.75 b | 38.20 ± 0.60 a | 11.50 ± 0.50 a | 46.50 ± 8.47 b | 8.28 ± 0.43 a |
| HDEEA-pABA | 6.25 ± 0.95 a | 36.15 ± 0.25 b | 10.50 ± 0.50 b | 51.00 ± 1.00 b | 8.39 ± 0.37 a |
| LSD5% | 0.61 | 1.51 | 0.59 | 15.93 | 1.28 |
| Development Stage/ Treatments | Beginning of Flowering | Flowering | Emergence of First Fruiting Floor |
|---|---|---|---|
| Control (MES) | z 42.50 ± 3.07 b | z 66.75 ± 5.02 b | yz 83.75 ± 7.53 b |
| IAA | z 49.00 ± 0.58 a | z 77.00 ± 2.89 a | yz 96.67 ± 3.67 a |
| HEEA-pABA | z 43.00 ± 4.51 b | z 64.67 ± 4.67 b | yz 85.67 ± 7.85 b |
| HDEEA-pABA | x 47.00 ± 1.00 a | zx 79.67 ± 1.20 a | yz 104.33 ± 3.39 a |
| Development Stage/ Treatments | Beginning of Flowering | Flowering | Emergence of First Fruiting Floor |
|---|---|---|---|
| Control (MES) | y 6.50 ± 0.50 a | x 10.50 ± 0.87 b | x 12.75 ± 0.48 b |
| IAA | y 7.33 ± 0.67 a | x 12.33 ± 0.67 a | x 13.67 ± 0.88 ab |
| HEEA-pABA | y 7.33 ± 0.33 a | x 10.67 ± 1.45 b | x 13.33 ± 0.88 ab |
| HDEEA-pABA | y 6.67 ± 0.88 a | x 12.00 ± 1.00 ab | x 14.33 ± 0.67 a |
| Development Stage/ Treatments | Beginning of Flowering | Flowering | Emergence of First Fruiting Floor |
|---|---|---|---|
| Control (MES) | u 39.13 ± 1.43 b | z 47.00 ± 2.14 b | yz 48.75 ± 0.70 b |
| IAA | y 39.33 ± 1.53 b | x 51.50 ± 1.45 a | x 50.73 ± 0.69 a |
| HEEA-pABA | z 37.07 ± 1.74 c | y 47.43 ± 2.38 b | y 49.30 ± 1.46 b |
| HDEEA-pABA | y 41.43 ± 0.69 a | x 50.37 ± 0.03 a | x 51.17 ± 0.23 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sumalan, R.-L.; Croitor, L.; Petric, M.; Radulov, I.; Bourosh, P.; Sumalan, R.-M.; Crisan, M. p-Aminobenzoate Organic Salts as Potential Plant Growth Regulators for Tomatoes. Molecules 2020, 25, 1635. https://doi.org/10.3390/molecules25071635
Sumalan R-L, Croitor L, Petric M, Radulov I, Bourosh P, Sumalan R-M, Crisan M. p-Aminobenzoate Organic Salts as Potential Plant Growth Regulators for Tomatoes. Molecules. 2020; 25(7):1635. https://doi.org/10.3390/molecules25071635
Chicago/Turabian StyleSumalan, Radu-Liviu, Lilia Croitor, Mihaela Petric, Isidora Radulov, Paulina Bourosh, Renata-Maria Sumalan, and Manuela Crisan. 2020. "p-Aminobenzoate Organic Salts as Potential Plant Growth Regulators for Tomatoes" Molecules 25, no. 7: 1635. https://doi.org/10.3390/molecules25071635
APA StyleSumalan, R.-L., Croitor, L., Petric, M., Radulov, I., Bourosh, P., Sumalan, R.-M., & Crisan, M. (2020). p-Aminobenzoate Organic Salts as Potential Plant Growth Regulators for Tomatoes. Molecules, 25(7), 1635. https://doi.org/10.3390/molecules25071635