
Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise?




Abstract
1. Introduction
2. Endogenous Synthesis and Cell Transport of l-Carnitine
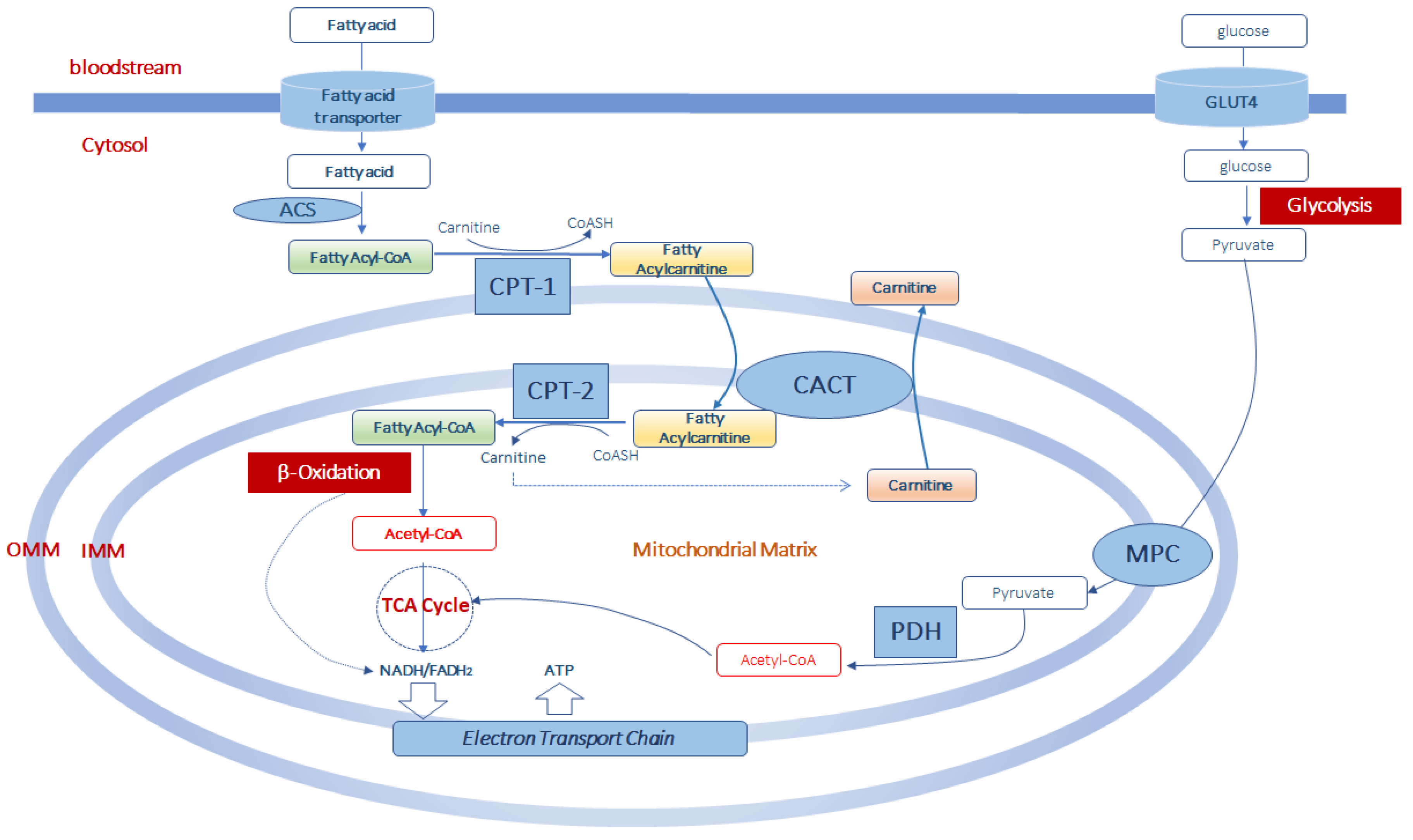
3. Role of Carnitine in Mitochondrial Fatty Acid Transport and β-Oxidation
4. Muscle l-Carnitine Selects Fuels during Exercise
5. How l-Carnitine Can Regulate Fatty Acid Oxidation during Physical Exercise
6. Can Carnitine Supplementation Be Useful in Physical Exercise?
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Siliprandi, N.; Di Lisa, F.; Menabo, R. Clinical use of carnitine. Past, present and future. Adv. Exp. Med. Biol. 1990, 272, 175. [Google Scholar] [PubMed]
- Karlic, H.; Lohninger, A. Supplementation of l-carnitine in athletes: Does it make sense? Nutrition 2004, 20, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Friolet, R.; Hoppeler, H.; Krähenbühl, S. Relationship between the coenzyme A and the carnitine pools in human skeletal muscle at rest and after exhaustive exercise under normoxic and acutely hypoxic conditions. J. Clin. Investig. 1994, 94, 1490–1495. [Google Scholar] [CrossRef]
- Reuter, S.E.; Evans, A.M. Carnitine and acylcarnitines: Pharmacokinetic, pharmacological and clinical aspects. Clin. Pharmacokinet. 2012, 51, 553–572. [Google Scholar] [CrossRef]
- Hiatt, W.R.; Regensteiner, J.G.; Wolfel, E.E.; Ruff, L.; Brass, E.P. Carnitine and acylcarnitine metabolism during exercise in humans. Dependence on skeletal muscle metabolic state. J. Clin. Investig. 1989, 84, 1167–1173. [Google Scholar] [CrossRef]
- Brass, E.P. Pharmacokinetic considerations for the therapeutic use of carnitine in hemodialysis patients. Clin. Ther. 1995, 17, 176–185. [Google Scholar] [CrossRef]
- Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Role of carnitine in disease. Nutr. Metab. 2010, 7, 30. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Brass, E.P. Carnitine and sports medicine: Use or abuse? Ann. N. Y. Acad. Sci. 2004, 1033, 67–78. [Google Scholar] [CrossRef]
- Heinonen, O.J. Carnitine and physical exercise. Sports Med. 1996, 22, 109–132. [Google Scholar] [CrossRef]
- Longo, N.; Frigeni, M.; Pasquali, M. Carnitine transport and fatty acid oxidation. Biochim. Biophys. Acta 2016, 1863, 2422–2435. [Google Scholar] [CrossRef]
- Tanphaichitr, V.; Broquist, H.P. Role of lysine and e-N-trimethyllysine in carnitine biosynthesis. Ii. Studies in the rat. J. Biol. Chem. 1973, 248, 2176–2181. [Google Scholar]
- Vaz, F.M.; Wanders, R.J. Carnitine biosynthesis in mammals. Biochem. J. 2002, 361, 417–429. [Google Scholar] [CrossRef]
- Borum, P.R. Carnitine. Annu. Rev. Nutr. 1983, 3, 233–259. [Google Scholar] [CrossRef]
- Adeva-Andany, M.M.; Calvo-Castro, I.; Fernández-Fernández, C.; Donapetry-García, C.; Pedre-Piñeiro, A.M. Significance of l-carnitine for human health. IUBMB Life 2017, 69, 578–594. [Google Scholar] [CrossRef]
- Hamilton, J.W.; Li, B.U.; Shug, A.L.; Olsen, W.A. Carnitine transport in human intestinal biopsy specimens. Demonstration of an active transport system. Gastroenterology 1986, 91, 10–16. [Google Scholar] [CrossRef]
- Rebouche, C.J. Kinetics, pharmacokinetics, and regulation of l-carnitine and acetyl-l-carnitine metabolism. Ann. N. Y. Acad. Sci. 2004, 1033, 30–41. [Google Scholar] [CrossRef]
- Evans, A.M.; Fornasini, G. Pharmacokinetics of L-carnitine. Clin. Pharmacokinet. 2003, 42, 941–967. [Google Scholar] [CrossRef]
- Rebouche, C. Carnitine: Modern Nutrition in Health and Disease; Shils, M., Shike, M., Ross, A., Eds.; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2006; pp. 537–544. [Google Scholar]
- Mohammed, A.; Majid, A.; Ayman, W.E. Carnitine inborn errors of metabolism. Molecules 2019, 24, 3251. [Google Scholar]
- Tamai, I. Pharmacological and pathophysiological roles of carnitine/organic cation transporters (OCTNs: Scarnitine22A4, Scarnitine22A5 and Slc22a21). Biopharm. Drug Dispos. 2013, 34, 29–44. [Google Scholar] [CrossRef]
- Lamhonwah, A.M.; Ackerley, C.A.; Tilups, A.; Edwards, V.D.; Wanders, R.J.; Tein, I. OCTN3 is a mammalian peroxisomal membrane carnitine transporter. Biochem. Biophys. Res. Commun. 2005, 338, 1966–1972. [Google Scholar] [CrossRef]
- Stanley, C.A.; DeLeeuw, S.; Coates, P.M.; Vianey-Liaud, C.; Divry, P.; Bonnefont, J.P.; Saudubray, J.M.; Haymond, M.; Trefz, F.K.; Breningstall, G.N.; et al. Chronic cardiomyopathy and weakness or acute coma in children with a defect in carnitine uptake. Ann. Neurol. 1991, 30, 709–716. [Google Scholar] [CrossRef]
- Ingoglia, F.; Visigalli, R.; Rotoli, B.M.; Barilli, A.; Riccardi, B.; Puccini, P.; Dall’Asta, V. Functional activity of l-carnitine transporters in human airway epithelial cells. Biochim. Biophys. Acta 2016, 1858, 210–219. [Google Scholar] [CrossRef]
- Tein, I.; Bukovac, S.W.; Xie, Z.W. Characterization of the human plasmalemmal carnitine transporter in cultured skin fibroblasts. Arch. Biochem. Biophys. 1996, 329, 145–155. [Google Scholar] [CrossRef]
- Nakanishi, T.; Hatanaka, T.; Huang, W.; Prasad, P.D.; Leibach, F.H.; Ganapathy, M.E.; Ganapathy, V. Na+- and Cl- coupled active transport of carnitine by the amino acid transporter ATB(0,1) from mouse colon expressed in HRPE cells and Xenopus oocytes. J. Physiol. 2001, 532, 297–304. [Google Scholar] [CrossRef]
- Enomoto, A.; Wempe, M.F.; Tsuchida, H.; Shin, H.J.; Cha, S.H.; Anzai, N.; Goto, A.; Sakamoto, A.; Niwa, T.; Kanai, Y.; et al. Molecular identification of a novel carnitine transporter specific to human testis. Insights into the mechanism of carnitine recognition. J. Biol. Chem. 2002, 277, 36262–36271. [Google Scholar] [CrossRef]
- Georges, B.; Le Borgne, F.; Galland, S.; Isoir, M.; Ecosse, D.; Grand-Jean, F.; Demarquoy, J. Carnitine transport into muscular cells. Inhibition of transport and cell growth by mildronate. Biochem. Pharmacol. 2000, 59, 1357–1363. [Google Scholar] [CrossRef]
- El-Hattab, A.W.; Scaglia, F. Disorders of carnitine biosynthesis and transport. Mol. Genet. Metab. 2015, 116, 107–112. [Google Scholar] [CrossRef]
- Jain, S.S.; Luiken, J.J.; Snook, L.A.; Han, X.X.; Holloway, G.P.; Glatz, J.F.; Bonen, A. Fatty acid transport and transporters in muscle are critically regulated by Akt2. FEBS Lett. 2015, 589, 2769–2775. [Google Scholar] [CrossRef]
- Radif, Y.; Ndiaye, H.; Kalantzi, V.; Jacobs, R.; Hall, A.; Minogue, S.; Waugh, M.G. The endogenous subcellular localisations of the long chain fatty acid-activatingenzymes ACSL3 and ACSL4 in sarcoma and breast cancer cells. Mol. Cell. Biochem. 2018, 448, 275–286. [Google Scholar] [CrossRef]
- Jia, Z.; Pei, Z.; Maiguel, D.; Toomer, C.J.; Watkins, P.A. The fatty acid transport protein (FATP) family: Very long chain acyl-CoA synthetases or solute carriers? J. Mol. Neurosci. 2007, 33, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Hoppel, C.L. Carnitine and carnitine palmitoyltransferase in fatty acid oxidation and ketosis. Fed. Proc. 1982, 41, 2853–2857. [Google Scholar] [PubMed]
- Rufer, A.C.; Thoma, R.; Hennig, M. Structural insight into function and regulation of carnitine palmitoyltransferase. Cell. Mol. Life Sci. 2009, 66, 2489–2501. [Google Scholar] [CrossRef] [PubMed]
- Serviddio, G.; Giudetti, A.M.; Bellanti, F.; Priore, P.; Rollo, T.; Tamborra, R.; Siculella, L.; Vendemiale, G.; Altomare, E.; Gnoni, G.V. Oxidation of hepatic carnitine palmitoyl transferase-I (CPT-I) impairs fatty acid beta-oxidation in rats fed a methionine-choline deficient diet. PLoS ONE 2011, 6, e24084. [Google Scholar] [CrossRef] [PubMed]
- Woldegiorgis, G.; Shi, J.; Zhu, H.; Arvidson, D.N. Functional characterization of mammalian mitochondrial carnitine palmitoyltransferases I and II expressed in the yeast Pichia pastoris. J. Nutr. 2000, 130, 310S–314S. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Wolfgang, M.J. Metabolomic profiling reveals a role for CPT1c in neuronal oxidative metabolism. BMC Biochem. 2012, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Foster, D.W. Malonyl-CoA: The regulator of fatty acid synthesis and oxidation. J. Clin. Investig. 2012, 122, 1958–1959. [Google Scholar] [CrossRef]
- Zeth, K.; Thein, M. Porins in prokaryotes and eukaryotes: Common themes and variations. Biochem. J. 2010, 431, 13–22. [Google Scholar] [CrossRef]
- Indiveri, C.; Iacobazzi, V.; Tonazzi, A.; Giangregorio, N.; Infantino, V.; Convertini, P.; Console, L.; Palmieri, F. The mitochondrial carnitine/acylcarnitine carrier: Function, structure and physiopathology. Mol. Asp. Med. 2011, 32, 223–233. [Google Scholar] [CrossRef]
- Tonazzi, A.; Galluccio, M.; Oppedisano, F.; Indiveri, C. Functional reconstitution into liposomes and characterization of the carnitine transporter from rat liver microsomes. Biochim. Biophys. Acta 2006, 1758, 124–131. [Google Scholar] [CrossRef][Green Version]
- Giudetti, A.M.; Stanca, E.; Siculella, L.; Gnoni, G.V.; Damiano, F. Nutritional and hormonal regulation of citrate and carnitine/acylcarnitine transporters: Two mitochondrial carriers involved in fatty acid metabolism. Int. J. Mol. Sci. 2016, 25, 817. [Google Scholar] [CrossRef] [PubMed]
- Priore, P.; Stanca, E.; Gnoni, G.V.; Siculella, L. Dietary fat types differently modulate the activity and expression of mitochondrial carnitine/acylcarnitine translocase in rat liver. Biochim. Biophys. Acta 2012, 1821, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, A.; Priore, P.; Gnoni, G.V.; Papa, S.; Zanotti, F.; Gnoni, A. 3,5-Diiodo-L-thyronine administration to hypothyroid rats rapidly enhances fatty acid oxidation rate and bioenergetic parameters in liver cells. PLoS ONE 2013, 8, e52328. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, A.; Taurino, F.; Damiano, F.; Siculella, L.; Sardanelli, A.M.; Gnoni, A. Acute administration of 3,5-diiodo-L-thyronine to hypothyroid rats stimulates bioenergetic parameters in liver mitochondria. J. Bioenerg. Biomembr. 2016, 48, 521–529. [Google Scholar] [CrossRef]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–197. [Google Scholar]
- Violante, S.; Ijlst, L.; Te Brinke, H.; Koster, J.; Tavares de Almeida, I.; Wanders, R.J.; Ventura, F.V.; Houten, S.M. Peroxisomes contribute to the acylcarnitine production when the carnitine shuttle is deficient. Biochim. Biophys. Acta 2013, 1831, 1467–1474. [Google Scholar] [CrossRef]
- Schrader, M.; Costello, J.; Godinho, L.F.; Islinger, M. Peroxisome-mitochondria interplay and disease. J. Inherit. Metab. Dis. 2015, 38, 681–702. [Google Scholar] [CrossRef]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Westerblad, H.; Bruton, J.D.; Katz, A. Skeletal muscle: Energy metabolism, fiber types, fatigue and adaptability. Exp. Cell Res. 2010, 316, 3093–3099. [Google Scholar] [CrossRef]
- Scribbans, T.D.; Vecsey, S.; Hankinson, P.B.; Foster, W.S.; Gurd, B.J. The Effect of Training Intensity on VO2max in Young Healthy Adults: A Meta-Regression and Meta-Analysis. Int. J. Exerc. Sci. 2016, 9, 230–247. [Google Scholar]
- Askew, E.W. Role of fat metabolism in exercise. Clin. Sports Med. 1984, 3, 605–621. [Google Scholar] [PubMed]
- Joyner, M.J.; Casey, D.P. Regulation of increased blood flow (hyperemia) to muscles during exercise: A hierarchy of competing physiological needs. Physiol. Rev. 2015, 95, 549–601. [Google Scholar] [CrossRef] [PubMed]
- Ranallo, R.F.; Rhodes, E.C. Lipid metabolism during exercise. Sports Med. 1998, 26, 29–42. [Google Scholar] [CrossRef] [PubMed]
- van Loon, L.J.; Greenhaff, P.L.; Constantin-Teodosiu, D.; Saris, W.H.; Wagenmakers, A.J. The effects of increasing exercise intensity on muscle fuel utilisation in humans. J. Physiol. 2001, 536, 295–304. [Google Scholar] [CrossRef]
- Hargreaves, M.; McConell, G.; Proietto, J. Influence of muscle glycogen on glycogenolysis and glucose uptake during exercise in humans. J. Appl. Physiol. 1995, 78, 288–292. [Google Scholar] [CrossRef]
- Hermansen, L.; Hultman, E.; Saltin, B. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 1967, 71, 129–139. [Google Scholar] [CrossRef]
- Ørtenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef]
- Wan, J.J.; Qin, Z.; Wang, P.Y.; Sun, Y.; Liu, X. Muscle fatigue: General understanding and treatment. Exp. Mol. Med. 2017, 49, e384. [Google Scholar] [CrossRef]
- Beneke, R.; Leithauser, R.M.; Ochentel, O. Blood lactate diagnostics in exercise testing and training. Int. J. Sports Physiol. Perform. 2011, 6, 8–24. [Google Scholar] [CrossRef]
- Messonnier, L.; Kristensen, M.; Juel, C.; Denis, C. Importance of pH regulation and lactate/H+ transport capacity for work production during supramaximal exercise in humans. J. Appl. Physiol. 2007, 102, 1936–1944. [Google Scholar] [CrossRef]
- van Hall, G. The physiological regulation of skeletal muscle fatty acid supply and oxidation during moderate-intensity exercise. Sports Med. 2015, 45, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, J.; Kiens, B. Regulation and limitations to fatty acid oxidation during exercise. J. Physiol. 2012, 590, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Purdom, T.; Kravitz, L.; Dokladny, K.; Mermier, C. Understanding the factors that affect maximal fat oxidation. J. Int. Soc. Sports Nutr. 2018, 15, 3. [Google Scholar] [CrossRef] [PubMed]
- Lundsgaard, A.M.; Fritzen, A.M.; Kiens, B. Molecular regulation of fatty acid oxidation in skeletal muscle during aerobic exercise. Trends Endocrinol. Metab. 2018, 29, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Broderick, T.L.; Quinney, H.A.; Lopaschuk, G.D. Carnitine stimulation of glucose oxidation in the fatty acid perfused isolated working rat heart. J. Biol. Chem. 1992, 267, 3758–3763. [Google Scholar] [PubMed]
- Primassin, S.; Ter Veld, F.; Mayatepek, E.; Spiekerkoetter, U. Carnitine supplementation induces acylcarnitine production in tissues of very long-chain acyl-CoA dehydrogenase-deficient mice, without replenishing low free carnitine. Pediatr. Res. 2008, 63, 632–637. [Google Scholar] [CrossRef]
- Bonnet, D.; Martin, D.; Pascale, D.L.; Villain, E.; Jouvet, P.; Rabier, D.; Brivet, m.; Saudubray, J.M. Arrhythmias and conduction defects as presenting symptoms of fatty acidoxidation disorders in children. Circulation 1999, 100, 2248–2253. [Google Scholar] [CrossRef]
- Pooyandjoo, M.; Nouhi, M.; Shab-Bidar, S.; Djafarian, K.; Olyaeemanesh, A. The effect of l-carnitine on weight loss in adults: A systematic review and meta-analysis of randomized controlled trials. Obes. Rev. 2016, 17, 970–976. [Google Scholar] [CrossRef]
- Fielding, R.; Riede, L.; Lugo, J.P.; Bellamine, A. l-Carnitine supplementation in recovery after exercise. Nutrients 2018, 10, 349. [Google Scholar] [CrossRef]
- Varney, J.L.; Fowler, J.W.; Gilbert, W.C.; Coon, C.N. Utilisation of supplemented l-carnitine for fuel efficiency, as an antioxidant, and for muscle recovery in Labrador retrievers. J. Nutr. Sci. 2017, 6. [Google Scholar] [CrossRef]
- Marconi, C.; Sassi, G.; Carpinelli, A.; Cerretelli, P. Effects of l-carnitine loading on the aerobic and anaerobic performance of endurance athletes. Eur. J. Appl. Physiol. Occup. Physiol. 1985, 54, 131–135. [Google Scholar] [CrossRef]
- Drăgan, G.I.; Vasiliu, A.; Georgescu, E.; Dumas, I. Studies concerning chronic and acute effects of l-carnitine on some biological parameters in elite athletes. Physiologie 1987, 24, 23–28. [Google Scholar] [PubMed]
- Drăgan, G.I.; Wagner, W.; Ploeşteanu, E. Studies concerning the ergogenic value of protein supply and l-carnitine in elite junior cyclists. Physiologie 1988, 25, 129–132. [Google Scholar] [PubMed]
- Drăgan, I.G.; Vasiliu, A.; Georgescu, E.; Eremia, N. Studies concerning chronic and acute effects of l-carnitina in elite athletes. Physiologie 1989, 26, 111–129. [Google Scholar] [PubMed]
- Vecchiet, L.; Di Lisa, F.; Pieralisi, G.; Ripari, P.; Menabò, R.; Giamberardino, M.A.; Siliprandi, N. Influence of l-carnitine administration on maximal physical exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1990, 61, 486–490. [Google Scholar] [CrossRef] [PubMed]
- Huertas, R.; Campos, Y.; Díaz, E.; Esteban, J.; Vechietti, L.; Montanari, G.; D’Iddio, S.; Corsi, M.; Arenas, J. Respiratory chain enzymes in muscle of endurance athletes: Effect of L-carnitine. Biochem. Biophys. Res. Commun. 1992, 188, 102–107. [Google Scholar] [CrossRef]
- Vukovich, M.D.; Costill, D.L.; Fink, W.J. Carnitine supplementation: Effect on muscle carnitine and glycogen content during exercise. Med. Sci. Sports Exerc. 1994, 26, 1122–1129. [Google Scholar] [CrossRef][Green Version]
- Greig, C.; Finch, K.M.; Jones, D.A.; Cooper, M.; Sargeant, A.J.; Forte, C.A. The effect of oral supplementation with l-carnitine on maximum and submaximum exercise capacity. Eur. J. Appl. Physiol. Occup. Physiol. 1987, 56, 457–460. [Google Scholar] [CrossRef]
- Siliprandi, N.; Di Lisa, F.; Pieralisi, G.; Ripari, P.; Maccari, F.; Menabo, R.; Giamberardino, M.A.; Vecchiet, L. Metabolic changes induced by maximal exercise in human subjects following l-carnitine administration. Biochim. Biophys. Acta 1990, 1034, 17–21. [Google Scholar] [CrossRef]
- Colombani, P.; Wenk, C.; Kunz, I.; Krähenbühl, S.; Kuhnt, M.; Arnold, M.; Frey-Rindova, P.; Frey, W.; Langhans, W. Effects of l-carnitine supplementation on physical performance and energy metabolism of endurance-trained athletes: A double-blind crossover field study. Eur. J. Appl. Physiol. Occup. Physiol. 1996, 73, 434–439. [Google Scholar] [CrossRef]
- Trappe, S.W.; Costill, D.L.; Goodpaster, B.; Vukovich, M.D.; Fink, W.J. The effects of l-carnitine supplementation on performance during interval swimming. Int. J. Sports Med. 1994, 15, 181–185. [Google Scholar] [CrossRef] [PubMed]
- Wächter, S.; Vogt, M.; Kreis, R.; Boesch, C.; Bigler, P.; Hoppeler, H.; Krähenbühl, S. Long-term administration of l-carnitine to humans: Effect on skeletal muscle carnitine content and physical performance. Clin. Chim. Acta 2002, 318, 51–61. [Google Scholar] [CrossRef]
- Orer, G.E.; Guzel, N.A. The effects of acute l-carnitine supplementation on endurance performance of athletes. J. Strength Cond. Res. 2014, 28, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Broad, E.M.; Maughan, R.J.; Galloway, S.D. Carbohydrate, protein, and fat metabolism during exercise after oral carnitine supplementation in humans. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Broad, E.M.; Maughan, R.J.; Galloway, S.D.R. Effects of exercise intensity and altered substrate availability on cardiovascular and metabolic responses to exercise after oral carnitine supplementation in athletes. Int. J. Sport Nutr. Exerc. Metab. 2011, 21, 385–397. [Google Scholar] [CrossRef][Green Version]
- Oyono-Enguelle, S.; Freund, H.; Ott, C.; Gartner, M.; Heitz, A.; Marbach, J.; Maccari, F.; Frey, A.; Bigot, H.; Bach, A.C. Prolonged submaximal exercise and l-carnitine in humans. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 58, 53–61. [Google Scholar] [CrossRef]
- Soop, M.; Björkman, O.; Cederblad, G.; Hagenfeldt, L.; Wahren, J. Influence of carnitine supplementation on muscle substrate and carnitine metabolism during exercise. J. Appl. Physiol. 1988, 64, 2394–2399. [Google Scholar] [CrossRef]
- Wyss, V.; Ganzit, G.P.; Rienzi, A. Effects of l-carnitine administration on VO2max and the aerobic-anaerobic threshold in normoxia and acute hypoxia. Eur. J. Appl Physiol. Occup. Physiol. 1990, 60, 1–6. [Google Scholar] [CrossRef]
- Decombaz, J.; Deriaz, O.; Acheson, K.; Gmuender, B.; Jequier, E. Effect of l-carnitine on submaximal exercise metabolism after depletion of muscle glycogen. Med. Sci. Sports Exerc. 1993, 25, 733–740. [Google Scholar] [CrossRef]
- Brass, E.P.; Hoppel, C.L.; Hiatt, W.R. Effect of intravenous l-carnitine on carnitine homeostasis and fuel metabolism during exercise in humans. Clin. Pharmacol. Ther. 1994, 55, 681–692. [Google Scholar] [CrossRef]
- Broad, E.M.; Maughan, R.J.; Galloway, S.D. Effects of four weeks l-carnitine L-tartrate ingestion on substrate utilization during prolonged exercise. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Gorostiaga, E.M.; Maurer, C.A.; Eclache, J.P. Decrease in respiratory quotient during exercise following l-carnitine supplementation. Int. J. Sports Med. 1989, 10, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Novakova, K.; Kummer, O.; Bouitbir, J.; Stoffel, S.D.; Hoerler-Koerner, U.; Bodmer, M.; Roberts, P.; Urwyler, A.; Ehrsam, R.; Krähenbühl, S. Effect of l-carnitine supplementation on the body carnitine pool, skeletal muscle energy metabolism and physical performance in male vegetarians. Eur. J. Nutr. 2016, 55, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Harper, P.; Elwin, C.E.; Cederblad, G. Pharmacokinetics of intravenous and oral bolus doses of l-carnitine in healthy subjects. Eur. J. Clin. Pharmacol. 1988, 35, 555–562. [Google Scholar] [CrossRef]
- Stephens, F.B.; Constantin-Teodosiu, D.; Laithwaite, D.; Simpson, E.J.; Greenhaff, P.L. Insulin stimulates l-carnitine accumulation in human skeletal muscle. FASEB J. 2006, 20, 377–379. [Google Scholar] [CrossRef]
- Dubelaar, M.L.; Lucas, C.M.; Hulsmann, W.C. The effect of l-carnitine on force development of the latissimus dorsi muscle in dogs. J. Card. Surg. 1991, 6, 270–275. [Google Scholar] [CrossRef]
- Hulsmann, W.C.; Dubelaar, M.L. Carnitine requirement of vascular endothelial and smooth muscle cells in imminent ischemia. Mol. Cell. Biochem. 1992, 116, 125–129. [Google Scholar] [CrossRef]
- Volek, J.S.; Judelson, D.A.; Silvestre, R.; Yamamoto, L.M.; Spiering, B.A.; Hatfield, D.L.; Vingren, J.L.; Quann, E.E.; Anderson, J.M.; Maresh, C.M.; et al. Effects of carnitine supplementation on flow-mediated dilation and vascular inflammatory responses to a high-fat meal in healthy young adults. Am. J. Cardiol. 2008, 102, 1413–1417. [Google Scholar] [CrossRef]
- Stephens, F.B.; Constantin-Teodosiu, D.; Laithwaite, D.; Simpson, E.J.; Greenhaff, P.L. An acute increase in skeletal muscle carnitine content alters fuel metabolism in resting human skeletal muscle. J. Clin. Endocrinol. Metab. 2006, 91, 5013–5018. [Google Scholar] [CrossRef]
- Steenge, G.R.; Simpson, E.J.; Greenhaff, P.L. Protein- and carbohydrate-induced augmentation of whole body creatine retention in humans. J. Appl. Physiol. 2000, 89, 1165–1171. [Google Scholar] [CrossRef]
- Koeth, R.A.; Lam-Galvez, B.R.; Kirsop, J.; Wang, Z.; Levison, B.S.; Gu, X.; Copeland, M.F.; Bartlett, D.; Cody, D.B.; Dai, H.J.; et al. l-Carnitine in omnivorous diets induces an atherogenic gut microbial pathway in humans. J. Clin. Investig. 2019, 129, 373–387. [Google Scholar] [CrossRef]
- Wu, W.K.; Chen, C.C.; Liu, P.Y.; Panyod, S.; Liao, B.Y.; Chen, P.C.; Kao, H.L.; Kuo, H.C.; Kuo, C.H.; Chiu, T.H.T.; et al. Identification of TMAO-producer phenotype and host-diet-gut dysbiosis by carnitine challenge test in human and germ-free mice. Gut 2019, 68, 1439–1449. [Google Scholar] [CrossRef]
- Randrianarisoa, E.; Lehn-Stefan, A.; Wang, X.; Hoene, M.; Peter, A.; Heinzmann, S.S.; Zhao, X.; Königsrainer, I.; Königsrainer, A.; Balletshofer, B.; et al. Relationship of serum Trimethylamine N-Oxide (TMAO) levels with early atherosclerosis in humans. Sci. Rep. 2016, 6, 26745. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.H.; Hazen, S.L. Microbiome, Trimethylamine N-Oxide (TMAO), and cardiometabolic disease. Transl. Res. 2017, 179, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chang, M.; Guo, Y.; Zhang, L.; Xue, C.; Yanagita, T.; Zhang, T.; Wang, Y. Trimethylamine-N-oxide (TMAO)-induced atherosclerosis is associated with bile acid metabolism. Lipids Health Dis. 2018, 17, 286. [Google Scholar] [CrossRef] [PubMed]
- Arsenian, M.A. Carnitine and its derivatives in cardiovascular disease. Prog. Cardiovasc. Dis. 1997, 40, 265–286. [Google Scholar] [CrossRef]
- Gao, X.; Tian, Y.; Randell, E.; Zhou, H.; Sun, G. Unfavorable associations between serum trimethylamine N-oxide and l-carnitine levels with components of metabolic syndrome in the Newfoundland population. Front. Endocrinol. 2019, 10, 168. [Google Scholar] [CrossRef]


{kind=link}
| Study | Population | Daily Carnitine Dose and Treatment Duration | l-Carnitine Effects |
|---|---|---|---|
| Marconi et al., 1985 [72] | 6 competitive walkers | 4g orally, 2 wks | Slight but significant increase in VO2max. |
| Greig et al., 1987 [79] | 9 untrained subjects | 2 g orally, 2 wks | Very little benefit to exercise performance. |
| Dragan et al., 1987 [73] | 40 top athletes | 3 g orally, 3 wks | Increase in VO2max. |
| Dragan et al., 1988 [75] | 7 junior athletes | 1 g orally, 6 wks and 2 g, 10 d | Supplemented group obtained higher performances. |
| Oyono-Enguelle et al., 1988 [87] | 10 exercising subjects | 2 g orally, 4 wks | No distinct increase of the relative participation of endogenous lipids in the fuel supply. |
| Soop et al., 1988 [88] | 7 moderately trained males | 5 g orally, 5 d | Carnitine does not influence muscle substrate utilization either at rest or during prolonged exercise. |
| Dragan et al., 1989 [74] | 110 top athletes | 1 g intravenously (single dose) 3 g orally, 3 wks | Single dose: beneficial effects on physical output, lipid metabolism and muscular function (contraction). 3 weeks treatment: beneficial effects on the lipid metabolism and VO2max. |
| Gorostiaga et al., 1989 [93] | 10 trained athletes | 2 g orally, 4wks | Increased lipid utilization by muscle during exercise. |
| Siliprandi et al., 1990 [80] | 10 moderately trained males | 2 g orally 1 dose 1 h before exam | Stimulation of PDH activity, and decrease in plasma lactate and pyruvate. |
| Vecchiet et al., 1990 [76] | 10 moderately trained males | 2 g orally, before high-intensity exercise | Increased VO2max. |
| Wyss et al., 1990 [89] | 7 healthy males | 3 g orally, 7 d | Lower rate of carbohydrate transformation during hypoxia. |
| Huertas et al., 1992 [77] | 14 athletes | 2 g orally, 4 wks | Increase in respiratory-chain enzyme activities in the muscle. |
| Decombaz et al., 1993 [90] | 9 healthy males | 3 g orally, 7 d | No influence of l-carnitine on muscle metabolism. |
| Trappe et al., 1994 [82] | 20 male swimmers | 2 g orally, 7 d | No differences in performance times were observed between trials or between groups. |
| Brass et al., 1994 [91] | 14 healthy males | 185 µmol/kg intravenously | l-carnitine administration has no significant effect on fuel metabolism during exercise in humans. |
| Vukovich et al., 1994 [78] | 8 healthy males | 6 g orally, 7–14 d | No differences in VO2max and respiratory exchange ratio. |
| Colombani et al., 1996 [81] | 7 male subjects | 2 g orally, 2 h before the start of marathon and after 20 km run | l-carnitine does not affect the metabolism and the physical performance of the endurance-trained athletes during the run and did not alter their recovery. |
| Wachter, et al., 2002 [83] | 8 healthy males | 4 g orally, 3 months | l-carnitine supplementation is not associated with a significant increase in physical performance. |
| Broad et al., 2005 [92] | 15 trained males | 3 g orally, 4 wks | No effect on substrate utilization or endurance performance. |
| Broad et al., 2008 [85] | 20 active male athletes | 2 g orally, 2 wks | No effect on fat, carbohydrate, or protein contribution to metabolism during prolonged moderate-intensity cycling exercise. |
| Broad et al., 2011 [86] | 15 athletes | 3 g orally, 15 d | l-carnitine induces changes in substrate utilization in metabolically active tissues but it does not affect whole-body substrate utilization during short-duration exercise. |
| Orer et al., 2014 [84] | 26 athletes | 12 received 3 g orally 14 received 4 g orally | Both 3 g and 4 g of l-carnitine taken before physical exercise prolonged exhaustion. |
| Novakova et al., 2016 [94] | 16 vegetarians and 8 omnivores | 2 g orally, 12 wks | l-carnitine supplementation does not affect muscle function and energy metabolism in vegetarian. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gnoni, A.; Longo, S.; Gnoni, G.V.; Giudetti, A.M. Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise? Molecules 2020, 25, 182. https://doi.org/10.3390/molecules25010182
Gnoni A, Longo S, Gnoni GV, Giudetti AM. Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise? Molecules. 2020; 25(1):182. https://doi.org/10.3390/molecules25010182
Chicago/Turabian StyleGnoni, Antonio, Serena Longo, Gabriele V. Gnoni, and Anna M. Giudetti. 2020. "Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise?" Molecules 25, no. 1: 182. https://doi.org/10.3390/molecules25010182
APA StyleGnoni, A., Longo, S., Gnoni, G. V., & Giudetti, A. M. (2020). Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise? Molecules, 25(1), 182. https://doi.org/10.3390/molecules25010182