
Cuticular Structures in Micropterous Crickets (Orthoptera, Gryllidae, Petaloptilini, Gryllomorphini)



Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. SEM (Scanning Electron Microscopy)
2.3. Lifestyles
2.4. Troglomorphism
2.5. Statistical Analysis
3. Results
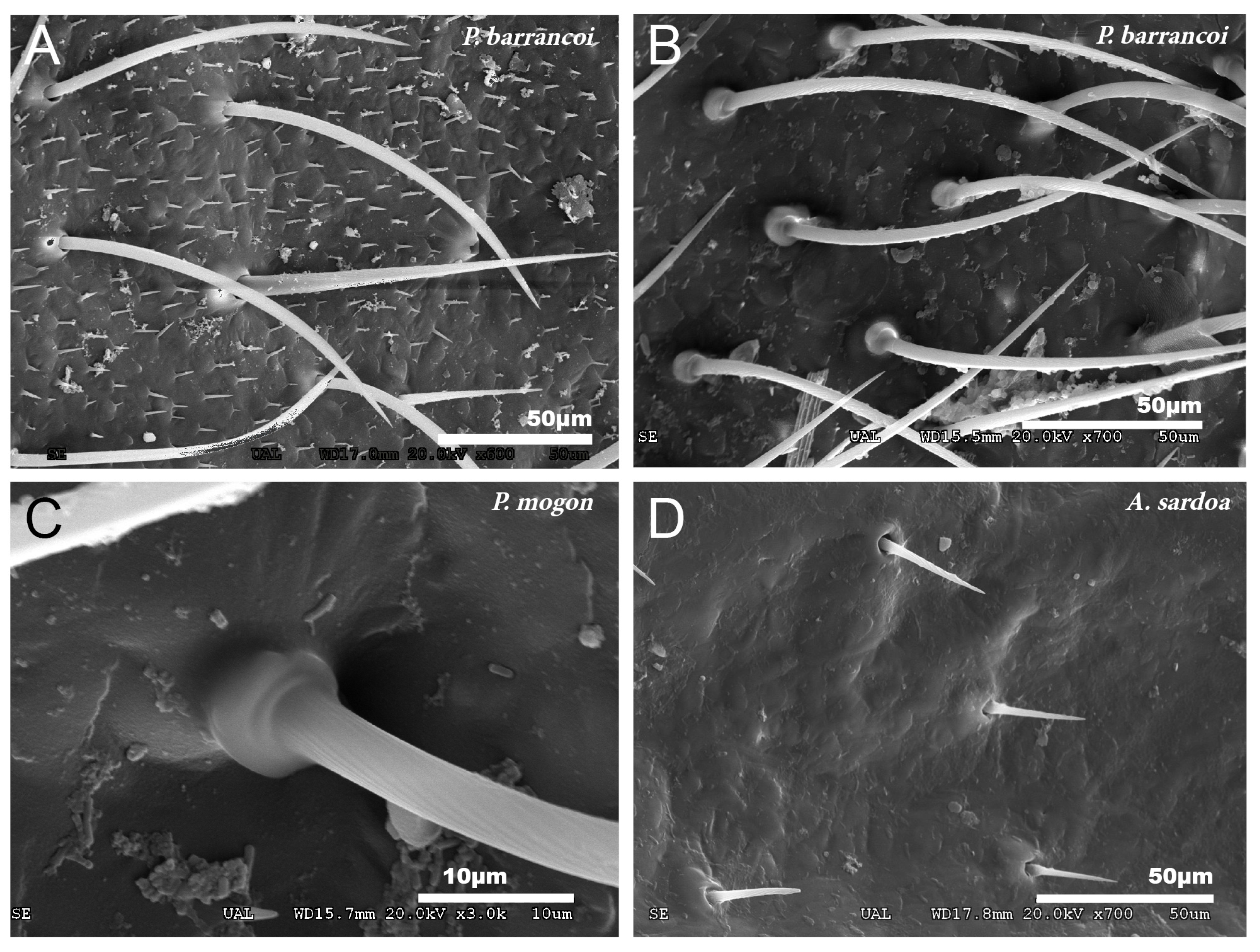
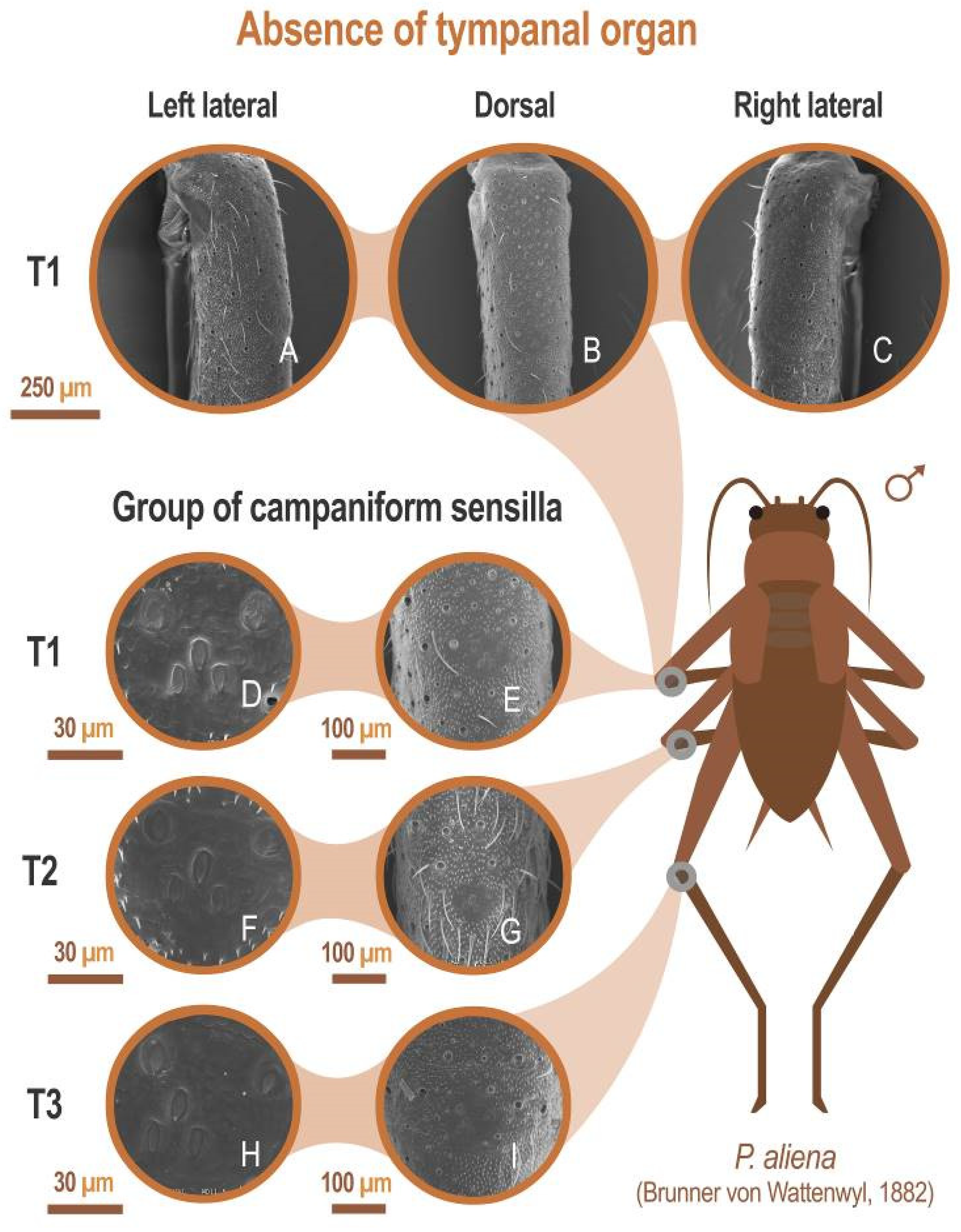
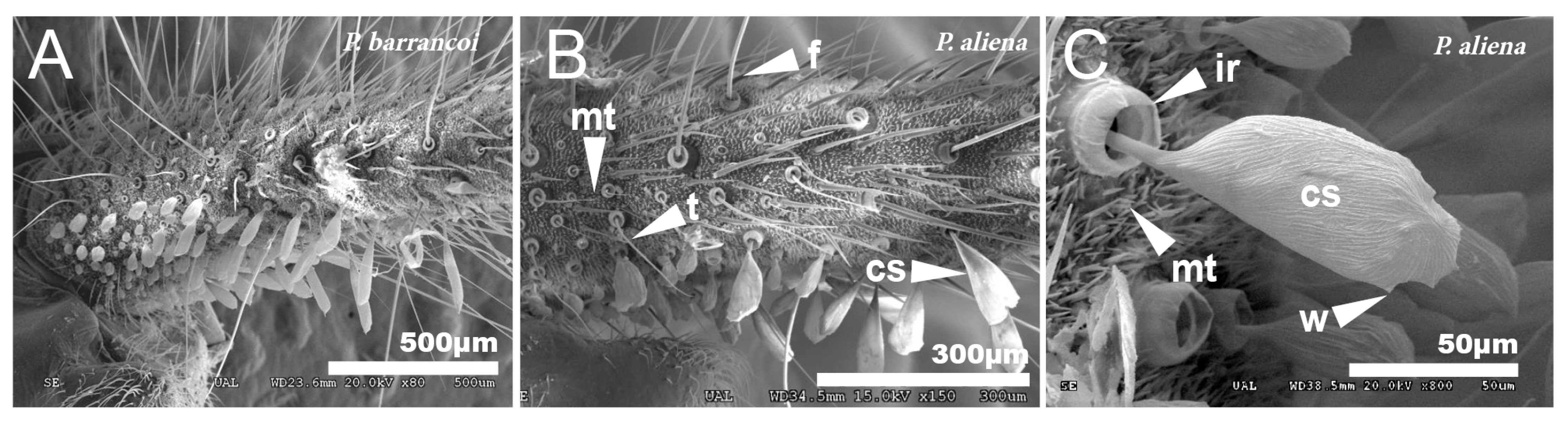
3.1. Study of Cuticular Structure by SEM
3.1.1. Tegmina
3.1.2. Legs
3.1.3. Dorsal Glands
3.1.4. Cerci
3.2. Lifestyles
3.3. Troglomorphic Adaptations in the Genus Petaloptila
3.3.1. Cephalic Lengthening and Ocular Reduction
3.3.2. Depigmentation
3.3.3. Elongation of Appendages
4. Discussion
4.1. Cuticular Structures
4.2. Troglomorphic Adaptations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, H.; Amédégnato, C.; Cigliano, M.M.; Desutter-Grandcolas, L.; Heads, S.W.; Huang, Y.; Otte, D.; Whiting, M.F. 300 million years of diversification: Elucidating the patterns of orthopteran evolution based on comprehensive taxon and gene sampling. Cladistics 2015, 31, 621–651. [Google Scholar] [CrossRef]
- Chang, H.; Qiu, Z.; Yuan, H.; Wang, X.; Li, X.; Sun, H.; Guo, X.; Lu, Y.; Feng, X.; Majid, M.; et al. Evolutionary rates of and selective constraints on the mitochondrial genomes of Orthoptera insects with different wing types. Mol. Phylogenet. Evol. 2020, 145, 106734. [Google Scholar] [CrossRef]
- Cigliano, M.M.; Braun, H.; Eades, D.C.; Otte, D. Orthoptera Species File. Version 5.0/5.0. Available online: http://Orthoptera.SpeciesFile.org (accessed on 19 June 2021).
- Resh, V.J.; Cardé, R.T. Encyclopedia of Insects, 2nd ed.; Academic Press: San Diego, CA, USA, 2009; ISBN 978-0123741448. [Google Scholar]
- Aguirre-Segura, A.; Barranco, P. Orden Orthoptera. Ibero Divers. Entomol. 2015, 46, 1–13. [Google Scholar]
- Naskrecki, P. Grasshoppers and their Relatives. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 722–734. ISBN 9780123847201. [Google Scholar]
- Uvarov, B.P. Grasshoppers and Locusts; Cambridge University Press: Cambridge, UK, 1966; Volume 1. [Google Scholar]
- Kevan, D.K.M. Orthoptera. In Synopsis and Classification of Living Organisms; Parker, S.P., Ed.; McGraw-Hill Book Company: New York, NY, USA, 1982; pp. 352–383. [Google Scholar]
- Song, H. Biodiversity of Orthoptera. In Insect Biodiversity: Science and Society; Foottit, R.G., Adler, P.H., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; Volume 2, pp. 245–279. ISBN 9781118945575. [Google Scholar]
- Gangwere, S. A Monograph on Food Selection in Orthoptera. Trans. Am. Entomol. Soc. 1961, 87, 67–230. [Google Scholar]
- Cade, W.H.; Rice, R. Field crickets (Orthoptera: Gryllidae) as prey of the toad Bufo marinus. Can. Entomol. 1980, 112, 335–336. [Google Scholar] [CrossRef]
- Nickle, D.A.; Heymann, E.W. Predation on Orthoptera and other orders in insects by tamarin monkeys, Saguinus mystax mystax and Saguinus fuscicollis nigrifrons (Primates: Callitrichidae), in north-eastern Peru. J. Zool. 1996, 239, 799–819. [Google Scholar] [CrossRef]
- Kok, O.B.; Louw, S.V.D.M. Avian and mammalian predators of Orthoptera in semi-arid regions of South Africa. S. Afr. J. Wildl. Res. 24 Mon. Delayed Open Access 2000, 30, 122–128. [Google Scholar]
- Jiguet, F. Arthropods in diet of Little Bustards Tetrax tetrax during the breeding season in western France. Bird Study 2002, 49, 105–109. [Google Scholar] [CrossRef]
- Rodríguez, C.; Bustamante, J. Patterns of Orthoptera abundance and lesser kestrel conservation in arable landscapes. Biodivers. Conserv. 2008, 17, 1753–1764. [Google Scholar] [CrossRef]
- Gage, S.H.; Mukerji, M.K. Crop losses associated with grasshoppers in relation to economics of crop production. J. Econ. Entomol. 1978, 71, 487–498. [Google Scholar] [CrossRef]
- Gangwere, S.K.; Muralirangan, M.C.; Muralirangan, M. Bionomics of Grasshoppers, Katydids, and Their Kin; CAB International: Wallingford, UK, 1997; ISBN 9780851991412. [Google Scholar]
- Onsager, J.A.; Olfert, O. What tools have potential for grasshopper pest management? In Grasshoppers and Grassland Health: Managing Grasshopper Outbreaks without Risking Environmental Disaste; Lockwood, J., Latchininsky, A., Sergeev, M.G., Eds.; Kluwer Academic Publishers: Dordrecht, NL, USA, 2000; pp. 145–156. ISBN 0792365291. [Google Scholar]
- Kinyuru, J.N.; Kenji, G.M.; Muhoho, S.N.; Ayieko, M.A. Nutritional potential of longhorn grasshopper (Ruspolia differens) consumed in Siaya District, Kenya. J. Agric. Sci. Technol. 2010, 12, 32–46. [Google Scholar]
- Ramos-Elorduy, J.; Pino, J.M.P.; Martínez, V.H. Could Grasshoppers Be a Nutritive Meal? Food Nutr. Sci. 2012, 3, 164–175. [Google Scholar]
- Wang, D.; Zhai, S.W.; Zhang, C.X.; Zhang, Q.; Chen, H. Nutrition value of the Chinese grasshopper Acrida cinerea (Thunberg) for broilers. Anim. Feed Sci. Technol. 2007, 135, 66–74. [Google Scholar] [CrossRef]
- Das, M.; Mandal, S. Assessment of nutritional quality and anti-nutrient composition of two edible grasshoppers (Orthoptera: Acrididae)—A search for new food alternative. Int. J. Med. Pharm. Sci. 2013, 3, 31–48. [Google Scholar]
- Barroso, F.G.; de Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-Bañón, C. The potential of various insect species for use as food for fish. Aquaculture 2014, 422, 193–201. [Google Scholar] [CrossRef]
- Baccetti, B. Notulae orthopterologicae XIV. Descrizione di un nuovo genere cavernicole de Ortotteri scoperto in Sardegna. Ann. Della Fac. Agrar. Dell Univ. Sassari 1960, 7, 3–13. [Google Scholar]
- Gorochov, A.V. Faunistic and systematic notes on Gryllidae (Orthoptera) of the Crimea. Vestn. Zool. 1984, 1, 12–20. [Google Scholar]
- Gorochov, A.V.; Ünal, M. Review of Turkish crickets from the subfamily Gryllomorphinae (Orthoptera: Gryllidae). Zoosyst. Ross. 2012, 21, 28–59. [Google Scholar] [CrossRef]
- Gorochov, A.V. Systematic position of the Hymenoptila Chopard and notes on the geographic history of Gryllomorphinae (Orthoptera: Gryllidae). Zoosyst. Ross. 2016, 25, 216–225. [Google Scholar] [CrossRef]
- Gorochov, A.V. A study of the genus Gryllomorpha Fieber, 1853 (Orthoptera: Gryllidae: Gryllomorphinae). Zoosyst. Ross. 2009, 18, 25–47. [Google Scholar] [CrossRef]
- Barranco, P. Estudio del subgénero Zapetaloptila Gorochov & Llorente, 2001 y descripción de cuatro nuevas especies (Petaloptila Pantel, 1890, Orthoptera, Gryllidae). Graellsia 2004, 60, 81–93. [Google Scholar]
- Barranco, P. Una nueva especie de Petaloptila de la provincia de Málaga (España) (Orthoptera, Gryllidae). Bol. Asoc. Esp. Entomol. 2010, 34, 207–217. [Google Scholar]
- Barranco, P. Estudio de una población natural del grillo cavernícola Petaloptila barrancoi Gorochov & Llorente 2001 (Orthoptera, Gryllidae). Bol. Asoc. Esp. Entomol. 2012, 36, 401–416. [Google Scholar]
- Grimaldi, D.; Engel, M.S.; Engel, M.S. Evolution of the Insects; Cambridge University Press: New York, NY, USA, 2005; ISBN 9780521821490. [Google Scholar]
- Boldyrev, B.T. Einige Episoden aus dem Geschlechtsleben von Discoptila fragosoi Bol. (Orthoptera, Gryllidae). Rev. Russe Entomol. 1928, 22, 137–147. [Google Scholar]
- Lopes-Andrade, C.; Sperber, C.F. Evaluation of the presence of glandular structures in preserved crickets (Orthoptera, Grylloidea, Phalangopsidae) using a comparative scanning electron microscopy technique. J. Orthoptera Res. 2001, 10, 343–345. [Google Scholar] [CrossRef]
- Prado, R. Reproductive behaviour of Eidmanacris corumbatai Garcia (Orthoptera: Phalangopsidae). Neotrop. Entomol. 2006, 35, 452–457. [Google Scholar] [CrossRef]
- Römer, H. Acoustic communication. In Insect Behavior: From Mechanisms to Ecological and Evolutionary Consequences; Córdoba-Aguilar, A., González-Tokman, D., González-Santoyo, I., Eds.; Oxford University Press: Oxford, UK, 2018; pp. 174–188. [Google Scholar]
- Riede, K. Acoustic monitoring of Orthoptera and its potential for conservation. J. Insect Conserv. 1998, 2, 217–223. [Google Scholar] [CrossRef]
- Ragge, D.R.; Reynolds, W.J. The Songs of the Grasshoppers and Crickets of Western Europe; Harley Books: London, UK, 1998; ISBN 9780946589494. [Google Scholar]
- Morris, G.K. Calling display and mating behaviour of Copiphora rhinoceros Pictet (Orthoptera: Tettigoniidae). Anim. Behav. 1980, 28, 42–51. [Google Scholar] [CrossRef]
- Field, L.H. Structure and evolution of stridulatory mechanisms in New Zealand wetas (Orthoptera: Stenopelmatidae). Int. J. Insect Morphol. Embryol. 1993, 22, 163–183. [Google Scholar] [CrossRef]
- López, H.; García, M.D.; Clemente, E.; Presa, J.J.; Oromí, P. Sound production mechanism in pamphagid grasshoppers (Orthoptera). J. Zool. 2008, 275, 1–8. [Google Scholar] [CrossRef]
- Massa, B. The role of the Krauss’s organ in sound production in Pamphagidae (Caelifera: Orthoptera). Ital. J. Zool. 2012, 79, 441–449. [Google Scholar] [CrossRef]
- Stritih, N.; Čokl, A. Mating behaviour and vibratory signalling in non-hearing cave crickets reflect primitive communication of Ensifera. PLoS ONE 2012, 7, e47646. [Google Scholar] [CrossRef]
- Lakes-Harlan, R.; Bailey, W.J.; Schikorski, T. The auditory system of an atympanate bushcricket Phasmodes ranatriformes (Westwood) (Tettigoniidae: Orthoptera). J. Exp. Biol. 1991, 158, 307–324. [Google Scholar] [CrossRef]
- Eibl, E. Morphology of the sense organs in the proximal parts of the tibiae of Gryllus campestris L. and Gryllus bimaculatus deGeer (Insecta, Ensifera). Zoomorphologie 1978, 89, 185–205. [Google Scholar] [CrossRef]
- Strauß, J.; Lomas, K.; Laurence, H.F. The complex tibial organ of the New Zealand ground weta: Sensory adaptations for vibrational signal detection. Sci. Rep. 2017, 7, 2031. [Google Scholar] [CrossRef]
- Deharveng, L.; Bedos, A. Diversity of Terrestrial Invertebrates in Subterranean Habitats. In Cave Ecology. Ecological Studies (Analysis and Synthesis); Moldovan, O., Kováč, Ľ., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 235. [Google Scholar]
- Hubbell, T.H.; Norton, R.M. The systematics and biology of the cave-crickets of the North American tribe Hadenoecini (Orthoptera: Saltatoria: Ensifera: Rhaphidophoridae: Dolichopodinae). Misc. Pub. Mus. Zool. Univ. Mich. 1978, 156, 1–124. [Google Scholar]
- Peck, S.B. The cave fauna of Alabama. Part II: The insects. NSS Bull. 1995, 57, 1–19. [Google Scholar]
- Lavoie, K.H.; Helf, K.L.; Poulson, T.L. The biology and ecology of North American cave crickets. J. Cave Karst Stud. 2007, 69, 114–134. [Google Scholar]
- Barranco, P.; Amate, J. Distribución y estructura de edades del grillo cavernícola Petaloptila barrancoi Gorochov & Llorente, 2001. (Orthoptera, Gryllidae). Espeleotemas 2008, 6, 50–57. [Google Scholar]
- Racovitza, E.G. Essay on Biospeological Problems; Casa Cărţii de Ştiinţă: Cluj-Napoca, Romania, 1907; pp. 127–183. [Google Scholar]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Howarth, F.G.; Moldovan, O.T. The Ecological Classification of Cave Animals and Their Adaptations. In Cave Ecology. Ecological Studies (Analysis and Synthesis); Moldovan, O., Kováč, Ľ., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; Volume 235. [Google Scholar]
- Ruffo, S. Le attuali conoscenze sulla fauna cavernicola della Regione Pugliese. Mem. Biogeogr. Adriat. 1957, 3, 1–143. [Google Scholar]
- Desutter-Grandcolas, L. The cricket fauna of Chiapanecan caves (Mexico): Systematics, phylogeny and the evolution of troglobitic life (Orthoptera, Grylloidea, Phalangopsidae, Luzarinae). Int. J. Speleol. 1993, 22, 1. [Google Scholar] [CrossRef]
- Desutter-Grandcolas, L. Studies in cave life evolution: A rationale for future theoretical developments using phylogenetic inference. J. Zool. Syst. Evol. Res. 1997, 35, 23–32. [Google Scholar] [CrossRef]
- Gould, S.J.; Vrba, E.S. Exaptation—A missing term in the science of form. Palaeobiology 1982, 8, 4–15. [Google Scholar] [CrossRef]
- Christiansen, K. Proposition pour la classification des animaux cavernicoles. Spelunca 1962, 2, 76–78. [Google Scholar]
- Christiansen, K.A. Morphological adaptations. In Encyclopedia of Caves, 2nd ed.; White, W.B., Culver, D.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 517–528. [Google Scholar]
- Desutter-Grandcolas, L. Test phylogénétique de l’adaptation á la vie troglobie chez des grillons (Insecta, Orthoptera, Grylloidea). Comptes Rendus De L’académie Des Sci. Série 3 Sci. De La Vie 1994, 317, 907–912. [Google Scholar]
- Desutter-Grandcolas, L. Are troglobitic taxa troglobiomorphic? A test using phylogenetic inference. Int. J. Speleol. 1997, 26, 1. [Google Scholar] [CrossRef]
- Ortuño, V.M.; Gilgado, J.D.; Jiménez-Valverde, A.; Sendra, A.; Pérez-Suárez, G.; Herrero-Borgoñón, J.J. The “alluvial mesovoid shallow substratum”, a new subterranean habitat. PLoS ONE 2013, 8, e76311. [Google Scholar] [CrossRef] [PubMed]
- Olmo-Vidal, J.M.; Hernando, C. A new cave cricket of the genus Petaloptila from the Pyrenean mountains (N. Iberian Peninsula) (Orthoptera: Gryllidae). J. Orthoptera Res. 2000, 9, 135–138. [Google Scholar] [CrossRef]
- Barranco, P. Nuevas citas de grillos para la Península Ibèrica (Orthoptera, Gryllidae). Bol. Asoc. Esp. Entomol. 2012, 36, 215–222. [Google Scholar]
- Eusébio, R.P.; Enghoff, H.; Solodovnikov, A.; Michelsen, A.; Barranco, P.; Salgado, J.M.; Sendra, A.; Reboleira, A.S.P.S. Temporal and spatial dynamics of arthropod groups in terrestrial subsurface habitats in central Portugal. Zoology 2021, 147, 125931. [Google Scholar] [CrossRef] [PubMed]
- Desutter-Grandcolas, L. Comparative morphology of cercal structures in true crickets and their allies (Orthoptera, Ensifera). Zoomorphology 1998, 118, 235–243. [Google Scholar] [CrossRef]
- Gorochov, A.V.; Llorente, V. Estudio taxonómico preliminar de los Grylloidea de España (Insecta, Orthoptera). Graellsia 2001, 57, 95–139. [Google Scholar] [CrossRef]
- Braud, Y.; Roesti, C.; Sardet, E. Petaloptila aliena (Brunner von Wattenwyl, 1882), nouvelle espèce pour la France (Orthoptera, Gryllidae). Matér. Orthoptériques Entomocénotiques 2008, 13, 65–68. [Google Scholar]
- Kevan, K.M.; Hsiung, C.C.A. Summary of the orthopteroid insect known from Lanzarote, Canary Island, with description of a new cricket species of the genus Hymenoptila Chopard, 1943 (Grylloptera; Grylloidea; Gryllidae). Vieraea Folia Sci. Biol. Canar. 1992, 21, 77–100. [Google Scholar]
- Pérez, A.J.; Morales, E.; Oromí, P.; López, H. Fauna de artrópodos de Montaña Clara (Islas Canarias) II: Hexápodos (no coleópteros). Vieraea 2003, 31, 237–251. [Google Scholar]
- Cadeddu, G. Acroneuroptila puddui. Nuova specie di Grillomorfino cavernicolo della Sardegna. Boll. Soc. Sarda Sc. Nat. 1970, 4, 3–14. [Google Scholar]
- Massa, B.; Fontana, P.; Buzzetti, F.M.; Kleukers, R.; Odè, B. Orthoptera. Fauna d’Italia, XLVIII; Edizioni Calderini: Bologna, Italy, 2012; 563p. [Google Scholar]
- Desutter-Grandcolas, L. Functional forewing morphology and stridulation in crickets (Orthoptera, Grylloidea). J. Zool. 1995, 236, 243–252. [Google Scholar] [CrossRef]
- Quennedey, A.; Drugmand, D.; Deligne, J. Morphology and ultrastructure of paired prototergal glands in the adult rove beetle Philonthus varians (Coleoptera, Staphylinidae). Arthropod Struct. Dev. 2002, 31, 173–183. [Google Scholar] [CrossRef]
- Sun, M.; Liang, A.; Watson, G.S.; Watson, J.A.; Zheng, Y.; Jiang, L. Compound Microstructures and Wax Layer of Beetle Elytral Surfaces and Their Influence on Wetting Properties. PLoS ONE 2012, 7, e46710. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liua, C.; Bhushanb, B. A review of beetle hindwings: Structure, mechanical properties, mechanism and bioinspiration. J. Mech. Behav. Biomed. Mater. 2019, 94, 63–73. [Google Scholar] [CrossRef]
- Mays, D.L. Mating behaviour if Nemobiine crickets Hygronemobius, Nemobius, and Pteronemobius (Orthoptera: Gryllidae). Fla. Entomol. 1971, 54, 113–126. [Google Scholar] [CrossRef]
- Fedorka, K.M.; Mousseau, T. Tibial spur feeding in ground crickets: Larger males contribute larger gifts (Orthoptera: Gryllidae). Fla. Entomol. 2002, 85, 317–323. [Google Scholar] [CrossRef]
- Alexander, R.D.; Otte, D. Cannibalism during copulation in the brown bush cricket, Hapithus agitator (Gryllidae). Fla. Entomol. 1967, 50, 79–87. [Google Scholar] [CrossRef]
- Ono, T.; Ando, C.; Kondo, Y. Effect of metanotal secretion ingestion on oviposition in a tree cricket, Truljalia hibinonis (Orthoptera: Gryllidae). Entomol. Sci. 2004, 7, 9–13. [Google Scholar] [CrossRef]
- Vahed, K. The function of nuptial feeding in insects: A review of empirical studies. Biol. Rev. 1998, 73, 43–78. [Google Scholar] [CrossRef]
- Prokop, P.; Maxwell, M.R. Interactions between multiple forms of nuptial feeding in the wood cricket Nemobius sylvestris (Bosc.): Dual spermatophores and male forewings. Ethology 2008, 114, 1173–1182. [Google Scholar] [CrossRef]
- Barranco, P.; Gilgado, T.; Ortuño, V. A new mute species of the genus Nemobius Serville (Orthoptera, Gryllidae, Nemobiinae) discovered in colluvial, stony debris in the Iberian Peninsula: A biological, phenological and biometric study). Zootaxa 2013, 3691, 201–219. [Google Scholar] [CrossRef]
- Otte, D. Relation between hearing and flying in crickets. Entomol. News 1990, 101, 29–34. [Google Scholar]
- Otte, D. Evolution of cricket songs. J. Orthoptera Res. 1992, 1, 25–49. [Google Scholar] [CrossRef]
- Field, L.H.; Matheson, T. Chordotonal organs of insects. Adv. Insect physiol. 1998, 27, 1–228. [Google Scholar]
- Desutter-Grandcolas, L.; Blanchet, E.; Robillard, T.; Magal, C.; Vannier, F.; Dangles, O. Evolution of the cercal sensory system in a tropical cricket clade (Orthoptera: Grylloidea: Eneopterinae): A phylogenetic approach. Biol. J. Linn. Soc. 2010, 99, 614–631. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Gilgado, J.D.; Sendra, A.; Pérez-Suárez, G.; Herrero-Borgoñón, J.J.; Ortuño, V.M. Exceptional invertebrate diversity in a scree slope in Eastern Spain. J. Insect Conserv. 2015, 19, 713–728. [Google Scholar] [CrossRef]
- Leroy, Y. Gryllides et Gryllacrides cavernicoles. Ann. Spéléol. 1967, 22, 659–722. [Google Scholar]
- Jugovic, J.; Šumer, N. Differences in troglomorphism and sexual dimorphism in two sympatric subtroglophile crickets of genus Troglophilus (Insecta: Orthoptera). Pol. J. Ecol. 2021, 69, 35–50. [Google Scholar]
- De Campos, L.D.; Souza-Dias, P.G.B.D.; Desutter-Grandcolas, L.; Shigueo Nihei, S. Colonization of different biomes drove the diversification of the Neotropical Eidmanacris crickets (Insecta: Orthoptera: Grylloidea: Phalangopsidae). PLoS ONE 2021, 16, e0245325. [Google Scholar] [CrossRef]
- Yang, J.; Dong, H.; He, M.; Gao, J. Mitochondrial genome characterization of Gryllodes sigillatus (Orthoptera: Gryllidae) and its phylogenetic implications. Mitochondrial DNA Part B 2021, 6, 1056–1058. [Google Scholar] [CrossRef]
- Desuter-Grandcolas, L. A phylogenetic analysis of the evolution of the stridulatory apparatus in true crickets (Orthoptera, Grylloidea). Cladistics 1997, 13, 101–108. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | SEM | Biometry |
|---|---|---|
| Gryllomorphini Saussure, 1877 | ||
| Gryllomorpha Fieber, 1853 | ||
| Hymenoptila Chopard, 1943 | ||
| Gryllomorpha (Hymenoptila) lanzarotensis Kevan versus Hsiung, 1992 | + | − |
| Petaloptilini Baccetti, 1959 | ||
| Acroneuroptila Baccetti, 1959 | ||
| Acroneuroptila puddui Cadeddu, 1970 | + | − |
| Acroneuroptila sardoa Baccetti, 1959 | + | − |
| Petaloptila Pantel, 1890 | ||
| Petaloptila Pantel, 1890 | ||
| Petaloptila (Petaloptila) aliena (Brunner von Wattenwyl, 1882) | + | + |
| Petaloptila (Petaloptila) fermini Gorochov & Llorente, 2001 | + | + |
| Petaloptila (Petaloptila) isabelae Gorochov & Llorente, 2001 | − | + |
| Petaloptila (Petaloptila) pallescens Bolívar, 1927 | − | + |
| Petaloptila (Petaloptila) pyrenaea Olmo-Vidal & Hernando, 2000 | − | + |
| Zapetaloptila Gorochov & Llorente, 2001 | ||
| Petaloptila (Zapetaloptila) baenai Barranco, 2004 | + | + |
| Petaloptila (Zapetaloptila) barrancoi Gorochov & Llorente, 2001 | + | + |
| Petaloptila (Zapetaloptila) bolivari (Cazurro, 1888) | − | + |
| Petaloptila (Zapetaloptila) malaciatana Barranco, 2010 | − | + |
| Petaloptila (Zapetaloptila) mogon Barranco, 2004 | + | + |
| Petaloptila (Zapetaloptila) venosa Gorochov & Llorente, 2001 | − | + |
| Species | Tegmina Glands | Protibial Tympani | Group of Campaniform Sensilla | Dorsal Glands |
|---|---|---|---|---|
| G. lanzarotensis | − | − | + | − |
| A. puddui | + | − | + | − |
| A. sardoa | + | − | + | − |
| P. (P.) aliena | + | − | + | + |
| P. (P.) fermini | + | − | + | + |
| P. (Z.) baenai | + | − | + | + |
| P. (Z.) barrancoi | + | − | + | + |
| P. (Z.) mogon | + | − | + | + |
| Species | Epigean | Hypogean | Distribution | ||
|---|---|---|---|---|---|
| MSS | Trogloxene | Troglophile | |||
| G. lanzarotensis | [70] | [71] | Canary Islands | ||
| A. puddui | [72] | Sardinia | |||
| A. sardoa | [73] | [73] | Sardinia | ||
| P. P. aliena | [68] | [65] | [65] | Spain, France | |
| P. P. fermini | [68] | [65] | Spain, Portugal | ||
| P. P. isabelae | [68] | [65] | Spain | ||
| P. P. pallescens | [68] | Spain | |||
| P. P. pyrenaea | [68] | [64,66] | * | Spain | |
| P. Z. baenai | * | * | [29] | Spain | |
| P. Z. barrancoi | * | * | [68] | Spain | |
| P. Z. bolivari | [63] | [68] | Spain | ||
| P. Z. malacitana | [30] | Spain | |||
| P. Z. mogon | [29] | Spain | |||
| P. Z. venosa | [68] | Spain | |||
| Species | Sex | n | PF/LP | PT/LP | HH/HE | HE/WE | LP/HH | Coloration |
|---|---|---|---|---|---|---|---|---|
| P. P. aliena | ♂ | 10 | 3.74 ± 0.18 a | 2.63 ± 0.22 a | 2.55 ± 0.19 a | 1.42 ± 0.15 ab | 0.79 ± 0.05 a | Dark |
| P. P. fermini | ♂ | 3 | 3.62 ± 0.40 a | 2.87 ± 0.21 a | 3.16 ± 0.21 a | 1.31 ± 0.05 ab | 0.73 ± 0.07 a | Dark |
| P. P. isabelae | ♂ | 3 | 3.53 ± 0.07 a | 2.82 ± 0.10 a | 3.17 ± 0.03 a | 1.33 ± 0.02 ab | 0.76 ± 0.01 a | Dark |
| P. P. pallescens | ♂ | 1 | 3.88 a | 2.92 a | 3.00 a | 1.29 ab | 0.76 a | Dark |
| P. P. aliena | ♀ | 10 | 3.71 ± 0.46 a | 2.49 ± 0.36 a | 2.66 ± 0.09 a | 1.35 ± 0.06 a | 0.81 ± 0.13 a | Dark |
| P. P. fermini | ♀ | 2 | 3.77 ± 0.07 a | 2.91 ± 0.31 a | 3.11 ± 0.23 a | 1.44 ± 0.09 a | 0.70 ± 0.01 a | Dark |
| P. P. isabelae | ♀ | 3 | 3.53 ± 0.23 a | 2.69 ± 0.11 a | 3.18 ± 0.03 a | 1.33 ± 0.08 a | 0.78 ± 0.03 a | Dark |
| P. P. pyrenaea | ♀ | 4 | 3.61 ± 0.25 a | 2.70 ± 0.11 a | 3.28 ± 0.07 a | 1.28 ± 0.07 a | 0.74 ± 0.06 a | Dark |
| P. Z. baenai | ♂ | 10 | 4.27 ± 0.11 b | 3.54 ± 0.13 b | 4.25 ± 0. 15 b | 1.54 ± 0.08 ab | 0.63 ± 0.01 b | Clear |
| P. Z. barrancoi | ♂ | 10 | 4.93 ± 0.59 b | 4.24 ± 0.47 b | 3.72 ± 0.24 b | 1.40 ± 0.07 ab | 0.64 ± 0.05 b | Clear |
| P. Z. bolivari | ♂ | 19 | 4.21 ± 0.18 b | 3.51 ± 0.23 b | 3.32 ± 0.16 b | 1.35 ± 0.10 ab | 0.70 ± 0.03 b | Clear |
| P. Z. malacitana | ♂ | 10 | 4.41 ± 0.03 b | 3.88 ± 0.29 b | 3.62 ± 0.22 b | 1.52 ± 0.17 ab | 0.69 ± 0.04 b | Clear |
| P. Z. mogon | ♂ | 10 | 4.41 ± 0.31 b | 3.65 ± 0.30 b | 3.68 ± 0.27 b | 1.46 ± 0.17 ab | 0.65 ± 0.07 b | Clear |
| P. Z. venosa | ♂ | 3 | 4.58 ± 0.68 b | 3.95 ± 0.61 b | 3.21 ± 0.35 b | 1.36 ± 0.02 ab | 0.67 ± 0.01 b | Clear |
| P. Z. baenai | ♀ | 10 | 4.15 ± 0.13 b | 3.45 ± 0.15 b | 4.31 ± 0.20 b | 1.50 ± 0.13 b | 0.67 ± 0.03 b | Clear |
| P. Z. barrancoi | ♀ | 10 | 4.61 ± 0.52 b | 3.88 ± 0.39 b | 3.65 ± 0.34 b | 1.53 ± 0.19 b | 0.69 ± 0.07 b | Clear |
| P. Z. bolivari | ♀ | 19 | 4.14 ± 0.17 b | 3.38 ± 0.20 b | 3.61 ± 0.30 b | 1.38 ± 0.17 b | 0.71 ± 0.05 b | Clear |
| P. Z. malacitana | ♀ | 10 | 4.24 ± 0.18 b | 3.59 ± 0.26 b | 3.68 ± 0.24 b | 1.43 ± 0.14 b | 0.70 ± 0.05 b | Clear |
| P. Z. mogon | ♀ | 10 | 4.45 ± 0.46 b | 3.70 ± 0.39 b | 3.75 ± 0.28 b | 1.63 ± 0.19 b | 0.66 ± 0.08 b | Clear |
| P. Z. venosa | ♀ | 8 | 4.01 ± 0.17 b | 3.46 ± 0.42 b | 3.50 ± 0.02 b | 1.49 ± 0.13 b | 0.71 ± 0.08 b | Clear |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barranco, P.; Molina-Pardo, J.L. Cuticular Structures in Micropterous Crickets (Orthoptera, Gryllidae, Petaloptilini, Gryllomorphini). Insects 2021, 12, 708. https://doi.org/10.3390/insects12080708
Barranco P, Molina-Pardo JL. Cuticular Structures in Micropterous Crickets (Orthoptera, Gryllidae, Petaloptilini, Gryllomorphini). Insects. 2021; 12(8):708. https://doi.org/10.3390/insects12080708
Chicago/Turabian StyleBarranco, Pablo, and José Luis Molina-Pardo. 2021. "Cuticular Structures in Micropterous Crickets (Orthoptera, Gryllidae, Petaloptilini, Gryllomorphini)" Insects 12, no. 8: 708. https://doi.org/10.3390/insects12080708
APA StyleBarranco, P., & Molina-Pardo, J. L. (2021). Cuticular Structures in Micropterous Crickets (Orthoptera, Gryllidae, Petaloptilini, Gryllomorphini). Insects, 12(8), 708. https://doi.org/10.3390/insects12080708