
Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence

 ,
,  ,
,  and
and 
Abstract
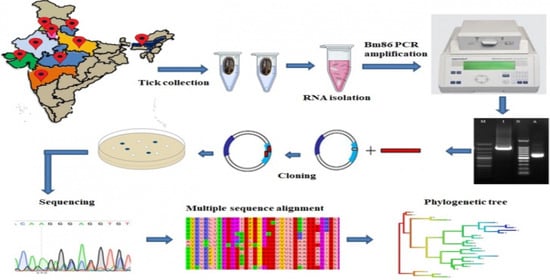
1. Introduction
2. Materials and Methods
2.1. Tick Samples
2.2. Study Area
2.3. RNA Isolation and cDNA Synthesis
2.4. Amplification of Full-Length IVRI-I Bm86 (orf) Gene, Cloning and Sequencing
2.5. Amplification of the Bm86 Conserved Sequence
2.6. Phylogenetic Analysis
2.7. In Silico Prediction of Linear B-Cell Epitopes on IVRI-IBm86 Protein
2.8. Evaluation of Degree of Conservation of Linear B-Cell Epitopes
3. Results
3.1. Sequence Analysis
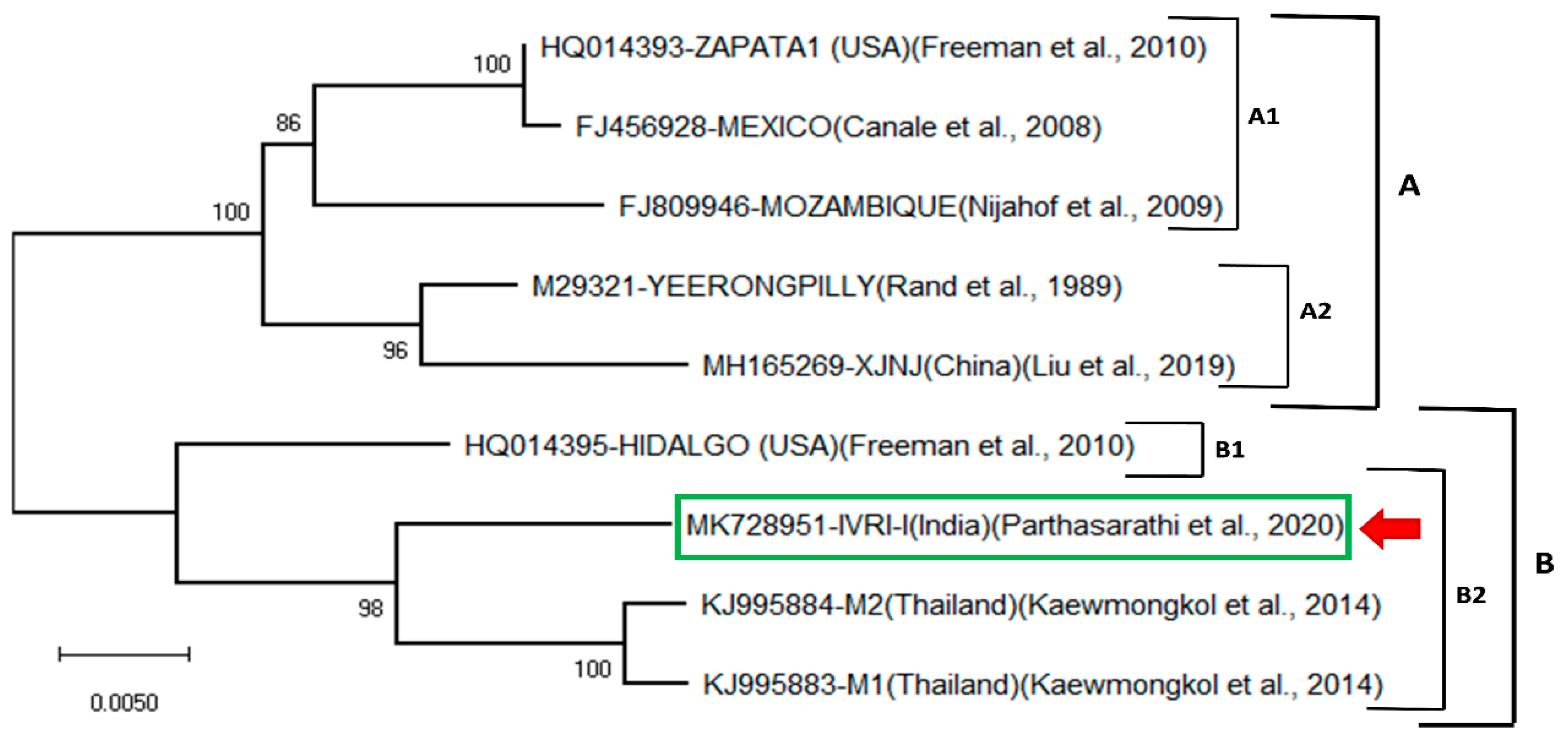
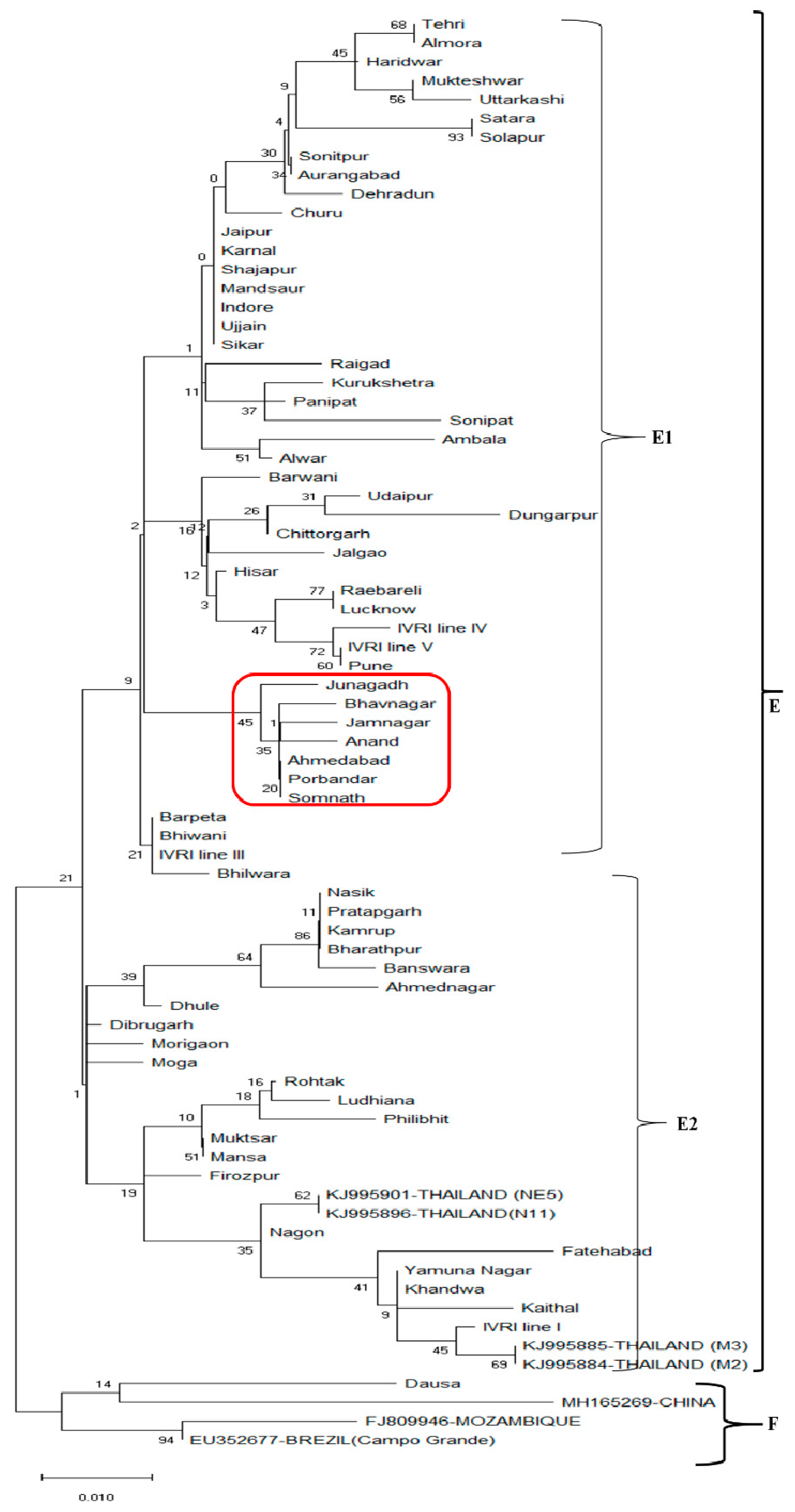
3.2. Phylogenetic Analysis
3.3. In Silico Analysis of IVRI-I Bm86 Protein
- 1.
- Two deletions in IVRI-I epitopes (G177-D224) showed increased antigenicity to Yeerongpilly and Camcord strains;
- 2.
- same VaxiJen scores were observed in four epitopes (T18-D45, D519-K554, C598-T606, T609-K623) of IVRI-I, Yeerongpilly and Camcord strains; the same score may be due to 100% similarity in the epitope sequence;
- 3.
- the substitutions/ mutations can increase (G177-D224, W280-R311) or decrease (D97-G129, K319-K501, H563-Q587) the antigenicity (VaxiJen scores) of IVRI-I epitopes when compared to the same epitopes in Yeerongpilly and Camcord strains (Supplementary Table S5).
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Department of Animal Husbandry & Dairying (DAHD), India. 20th Livestock Census. 2019. Available online: http://164.100.117.97/WriteReadData/userfiles/key%20results.pdf (accessed on 30 May 2020).
- Food and Agriculture Organization (FAO) Statistics. 2019. Available online: https://www.nddb.coop/sites/default/files/statistics/Country%20wise%20MP-ENG-2019.pdf (accessed on 13 March 2020).
- Ghosh, S.; Bansal, G.C.; Gupta, S.C.; Ray, D.; Khan, M.Q.; Irshad, H.; Shahiduzzaman, M.D.; Seitzer, U.; Ahmed, J.S. Status of tick distribution in Bangladesh, India and Pakistan. Parasitol. Res. 2007, 101, 207–216. [Google Scholar] [CrossRef]
- Olwoch, J.M.; Horak, I.G.; Scholtz, C.H.; Van Jaarsveld, A.S. Climate change and the genus Rhipicephalus (Acari: Ixodidae) in Africa. Onderstepoort. J. Vet. Res. 2007, 74, 45–72. [Google Scholar] [CrossRef]
- Barré, N.; Uilenberg, G. Spread of parasites transported with their hosts: Case study of two species of cattle tick. Rev. Sci. Tech. 2010, 29, 149. [Google Scholar] [CrossRef]
- Ghosh, S.; Nagar, G. Problem of ticks and tick-borne diseases in India with special emphasis on progress in tick control research: A review. J.Vector Borne Dis. 2014, 51, 259. [Google Scholar]
- Ticks FAO. Acaricide Resistance, Diagnosis, Management and Prevention. Guideline Resistance Management and Integrated Parasite Control in Ruminants Agriculture Department; FAO: Roma, Italy, 2004. [Google Scholar]
- Minjauw, B.; McLeod, A. Tick-Borne Diseases and Poverty: The Impact of Ticks and Tick-Borne Diseases on the Livelihoods of Small-Scale and Marginal Livestock Owners in India and Eastern and Southern Africa; University of Edinburg: Edinburgh, UK, 2003. [Google Scholar]
- Adehan, S.B.; Biguezoton, A.; Adakal, H.; Assogba, M.N.; Zoungrana, S.; Gbaguidi, A.M.; Tonouhewa, A.; Kandé, S.; Achi, L.; Kagone, H.; et al. Acaricide resistance of Rhipicephalus microplus ticks in Benin. Afr. J. Agric. Res. 2016, 11, 1199–1208. [Google Scholar] [CrossRef]
- Rodríguez-Hidalgo, R.; Pérez-Otáñez, X.; Garcés-Carrera, S.; Vanwambeke, S.O.; Madder, M.; Benítez-Ortiz, W. The current status of resistance to alpha-cypermethrin, ivermectin, and amitraz of the cattle tick (Rhipicephalus microplus) in Ecuador. PLoS ONE 2017, 12, e0174652. [Google Scholar] [CrossRef]
- El-Ashram, S.; Aboelhadid, S.M.; Kamel, A.A.; Mahrous, L.N.; Fahmy, M.M. First Report of Cattle Tick Rhipicephalus (Boophilus) annulatus in Egypt Resistant to Ivermectin. Insects 2019, 10, 404. [Google Scholar] [CrossRef] [PubMed]
- Higa, L.D.; Piña, F.T.; da Silva Rodrigues, V.; Garcia, M.V.; Salas, D.R.; Miller, R.J.; de Leon, A.P.; Barros, J.C.; Andreotti, R. Evidence of acaricide resistance in different life stages of Amblyomma mixtum and Rhipicephalus microplus (Acari: Ixodidae) collected from the same farm in the state of Veracruz. Prev. Vet. Med. 2020, 174, 104837. [Google Scholar] [CrossRef] [PubMed]
- Godara, R.; Katoch, R.; Rafiqi, S.I.; Yadav, A.; Nazim, K.; Sharma, R.; Singh, N.K.; Katoch, M. Synthetic pyrethroid resistance in Rhipicephalus (Boophilus) microplus ticks from north-western Himalayas, India. Trop. Anim. Health Prod. 2019, 51, 1203–1208. [Google Scholar] [CrossRef]
- Shakya, M.; Kumar, S.; Fular, A.; Upadhaya, D.; Sharma, A.K.; Bisht, N.; Nandi, A.; Ghosh, S. Emergence of fipronil resistant Rhipicephalus microplus populations in Indian states. Exp. Appl. Acarol. 2020, 1–2. [Google Scholar] [CrossRef]
- Graf, J.F.; Gogolewski, R.; Leach-Bing, N.; Sabatin, G.A.; Molento, M.B.; Bordin, E.L.; Arantes, G.J. Tick control: An industry point of view. Parasitology 2004, 129 (Suppl. 1), S427. [Google Scholar] [CrossRef]
- Anju, A.; Ravi, S.P.; Bechan, S. Water pollution with special reference to pesticide contamination in India. Water Resour. Prot. 2010. [Google Scholar] [CrossRef]
- Bulut, S.; Akkaya, L.; Gök, V.; Konuk, M. Organochlorine pesticide (OCP) residues in cow’s, buffalo’s, and sheep’s milk from Afyonkarahisar region. Environ. Monit. Assess. 2011, 181, 555–562. [Google Scholar] [CrossRef]
- Bulut, S.; Akkaya, L.; Gök, V.; Konuk, M. Organochlorine pesticide residues in butter and kaymak in Afyonkarahisar, Turkey. Asian. J. Anim. Vet. Adv. 2010, 9, 2797–2801. [Google Scholar] [CrossRef]
- Jayaraj, R.; Megha, P.; Sreedev, P. Organochlorine pesticides, their toxic effects on living organisms and their fate in the environment. Interdiscip. Toxicol. 2016, 9, 90–100. [Google Scholar] [CrossRef]
- de la Fuente, J.; Almazán, C.; Canales, M.; de la Lastra, J.M.; Kocan, K.M.; Willadsen, P. A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Anim. Health Res. Rev. 2007, 8, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Suarez, M.; Rubi, J.; Pérez, D.; Cordova, V.; Salazar, Y.; Vielma, A.; Barrios, F.; Gil, C.A.; Segura, N.; Carrillo, Y.; et al. High impact and effectiveness of Gavac™ vaccine in the national program for control of bovine ticks Rhipicephalus microplus in Venezuela. Livest. Sci. 2016, 187, 48–52. [Google Scholar] [CrossRef]
- de León, A.A.; Mahan, S.; Messenger, M.; Ellis, D.; Varner, K.; Schwartz, A.; Baca, D.; Andreotti, R.; Valle, M.R.; Cruz, R.R.; et al. Public-Private Partnership Enabled Use of Anti-Tick Vaccine for Integrated Cattle Fever Tick Eradication in the USA. In Pests and Vector-Borne Diseases in the Livestock Industry 2018; Wageningen Academic Publishers: TX, USA, 2018; pp. 783–784. [Google Scholar] [CrossRef]
- Perez-Perez, D.; Bechara, G.H.; Machado, R.Z.; Andrade, G.M.; Del Vecchio, R.E.; Pedroso, M.S.; Hernández, M.V.; Farnós, O. Efficacy of the Bm86 antigen against immature instars and adults of the dog tick Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae). Vet. Parasitol. 2010, 167, 321–326. [Google Scholar] [CrossRef]
- Rodríguez-Valle, M.; Taoufik, A.; Valdés, M.; Montero, C.; Hassan, I.; Hassan, S.M.; Jongejan, F.; de la Fuente, J. Efficacy of Rhipicephalus (Boophilus) microplus Bm86 against Hyalomma dromedarii and Amblyomma cajennense tick infestations in camels and cattle. Vaccine 2012, 30, 3453–3458. [Google Scholar] [CrossRef]
- Hüe, T.; Petermann, J.; Bonnefon, R.; Mermoud, I.; Rantoen, D.; Vuocolo, T. Experimental efficacy of a vaccine against Rhipicephalus australis. Exp. Appl. Acarol. 2017, 73, 245–256. [Google Scholar] [CrossRef]
- Valle, M.R.; Mèndez, L.; Valdez, M.; Redondo, M.; Espinosa, C.M.; Vargas, M.; Cruz, R.L.; Barrios, H.P.; Seoane, G.; Ramirez, E.S.; et al. Integrated control of Boophilus microplus ticks in Cuba based on vaccination with the anti-tick vaccine Gavac TM. Exp. Appl. Acarol. 2004, 34, 375–382. [Google Scholar] [CrossRef]
- García-García, J.C.; Gonzalez, I.L.; González, D.M.; Valdés, M.; Méndez, L.; Lamberti, J.; D’Agostino, B.; Citroni, D.; Fragoso, H.; Ortiz, M.; et al. Sequence variations in the Boophilus microplusBm86 locus and implications for immunoprotection in cattle vaccinated with this antigen. Exp. Appl. Acarol. 1999, 23, 883–895. [Google Scholar] [CrossRef]
- Freeman, J.M.; Davey, R.B.; Kappmeyer, L.S.; Kammlah, D.M.; Olafson, P.U. Bm86 midgut protein sequence variation in South Texas cattle fever ticks. Parasit. Vectors 2010, 3, 101. [Google Scholar] [CrossRef] [PubMed]
- Kaewmongkol, S.; Kaewmongkol, G.; Inthong, N.; Lakkitjaroen, N.; Sirinarumitr, T.; Berry, C.M.; Jonsson, N.N.; Stich, R.W.; Jittapalapong, S. Variation among Bm86 sequences in Rhipicephalus (Boophilus) microplus ticks collected from cattle across Thailand. Exp. Appl. Acarol. 2015, 66, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R. Ticks of Domestic Animals in Africa: A Guide to Identification of Species; Bioscience Reports: Edinburgh, Scotland, 2003. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Anbarasi, P.; Latha, B.R.; Sreekumar, C.; Senthuran, S. Partial sequencing of Bm86 gene for studying the phylogeny of an Indian isolate of Rhipicephalus (Boophilus) microplus tick. J. Parasit. Dis. 2014, 38, 260–264. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Lin, J.; Han, H.; Dong, H.; Zhao, Q.; Zhu, S.; Huang, B. Identification and characterization of Eimeria tenella apical membrane antigen-1 (AMA1). PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Blake, D.P.; Billington, K.J.; Copestake, S.L.; Oakes, R.D.; Quail, M.A.; Wan, K.L.; Shirley, M.W.; Smith, A.L. Genetic mapping identifies novel highly protective antigens for an apicomplexan parasite. PLoS Pathog 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Eisen, D.P.; Saul, A.; Fryauff, D.J.; Reeder, J.C.; Coppel, R.L. Alterations in Plasmodium falciparum genotypes during sequential infections suggest the presence of strain specific immunity. Am. J. Trop. Med. Hyg. 2002, 67, 8–16. [Google Scholar] [CrossRef]
- Healer, J.; Murphy, V.; Hodder, A.N.; Masciantonio, R.; Gemmill, A.W.; Anders, R.F.; Cowman, A.F.; Batchelor, A. Allelic polymorphisms in apical membrane antigen-1 are responsible for evasion of antibody-mediated inhibition in Plasmodium falciparum. Mol. Microbiol. 2004, 52, 159–168. [Google Scholar] [CrossRef]
- Drew, D.R.; Hodder, A.N.; Wilson, D.W.; Foley, M.; Mueller, I.; Siba, P.M.; Dent, A.E.; Cowman, A.F.; Beeson, J.G. Defining the antigenic diversity of Plasmodium falciparum apical membrane antigen 1 and the requirements for a multi-allele vaccine against malaria. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Hodder, A.N.; Crewther, P.E.; Anders, R.F. Specificity of the protective antibody response to apical membrane antigen 1. Infect. Immun. 2001, 69, 3286–3294. [Google Scholar] [CrossRef]
- Terheggen, U.; Drew, D.R.; Hodder, A.N.; Cross, N.J.; Mugyenyi, C.K.; Barry, A.E.; Anders, R.F.; Dutta, S.; Osier, F.H.; Elliott, S.R.; et al. Limited antigenic diversity of Plasmodium falciparum apical membrane antigen 1 supports the development of effective multi-allele vaccines. BMC Med. 2014, 12, 183. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.L.; Tomley, F.M.; Blake, D.P. Are Eimeria genetically diverse, and does it matter? Trends Parasitol. 2017, 33, 231–241. [Google Scholar] [CrossRef]
- Ghoshal, S.; Gajendra, P.; Kanjilal, S.D.; Mitra, M.; Sengupta, S. Diversity analysis of MSP1 identifies conserved epitope organization in block 2 amidst high sequence variability in Indian Plasmodium falciparum isolates. Malaria J. 2018, 17, 447. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, S.P.; Bhatt, R.; Singh, V. Genetic profiling of the Plasmodium falciparum parasite population in uncomplicated malaria from India. Malaria J. 2019, 18, 385. [Google Scholar] [CrossRef]
- Sossai, S.; Peconick, A.P.; Sales-Junior, P.A.; Marcelino, F.C.; Vargas, M.I.; Neves, E.S.; Patarroyo, J.H. Polymorphism of the Bm86 gene in South American strains of the cattle tick Boophilus microplus. Exp. Appl. Acarol. 2005, 37, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Murugan, K.; Ray, D.D.; Ghosh, S. Efficacy of rBm86 against Rhipicephalus (Boophilus) microplus (IVRI-I line) and Hyalomma anatolicum anatolicum (IVRI-II line) infestations on bovine calves. Parasitol. Res. 2012, 111, 629–635. [Google Scholar] [CrossRef] [PubMed]
- Doytchinova, I.A.; Flower, D.R. VaxiJen: A server for prediction of protective antigens, tumour antigens and subunit vaccines. BMC Bioinform. 2007, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Flower, D.R.; Macdonald, I.K.; Ramakrishnan, K.; Davies, M.N.; Doytchinova, I.A. Computer aided selection of candidate vaccine antigens. Immun. Res. 2010, 6, S1. [Google Scholar] [CrossRef] [PubMed]
- Fact Sheet: RTS, S Malaria Vaccine Candidate (Mosquirix™). Available online: https://www.malariavaccine.org/files/content/page/files/RTSS%20vaccine%20candidate%20Factsheet_FINAL.pdf/ (accessed on 1 September 2020).
- Roy, S.; Bhandari, V.; Dandasena, D.; Murthy, S.; Sharma, P. Genetic Profiling Reveals High Allelic Diversity, Heterozygosity and Antigenic Diversity in the Clinical Isolates of the Theileria annulata From India. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Kumar, B.; Manjunathachar, H.V.; Nagar, G.; Ravikumar, G.; de la Fuente, J.; Saravanan, B.C.; Ghosh, S. Functional characterization of candidate antigens of Hyalomma anatolicum and evaluation of its cross-protective efficacy against Rhipicephalus microplus. Vaccine 2017, 35, 5682–5692. [Google Scholar] [CrossRef] [PubMed]
- Shakya, M.; Kumar, B.; Nagar, G.; de la Fuente, J.; Ghosh, S. Subolesin: A candidate vaccine antigen for the control of cattle tick infestations in Indian situation. Vaccine 2014, 32, 3488–3494. [Google Scholar] [CrossRef] [PubMed]
- Manjunathachar, H.V.; Kumar, B.; Saravanan, B.C.; Choudhary, S.; Mohanty, A.K.; Nagar, G.; Chigure, G.; Ravi Kumar, G.V.; de la Fuente, J.; Ghosh, S. Identification and characterization of vaccine candidates against Hyalomma anatolicum—Vector of Crimean-Congo haemorrhagic fever virus. Transbound. Emerg. Dis. 2019, 66, 422–434. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | B | C | D | E | F | G | H | I | J | K | L | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | ID | 100 | 93.2 | 94.4 | 90.1 | 96.7 | 97.8 | 93 | 93.3 | 28.6 | 93.5 | 90.7 |
| B | 100 | ID | 93.2 | 94.4 | 90.1 | 96.7 | 97.8 | 93 | 93.3 | 28.6 | 93.5 | 90.7 |
| C | 93.2 | 93.2 | ID | 97 | 86.9 | 93.6 | 92.4 | 96.3 | 96.6 | 28.3 | 89.3 | 87.5 |
| D | 94.4 | 94.4 | 97 | ID | 88.7 | 95.5 | 93.5 | 94.6 | 94.9 | 29.3 | 91.5 | 89 |
| E | 90.1 | 90.1 | 86.9 | 88.7 | ID | 92.9 | 89.5 | 86.4 | 86.7 | 27 | 93.5 | 99 |
| F | 96.7 | 96.7 | 93.6 | 95.5 | 92.9 | ID | 95.8 | 93.2 | 93.5 | 28.6 | 93.8 | 93.5 |
| G | 97.8 | 97.8 | 92.4 | 93.5 | 89.5 | 95.8 | ID | 91.8 | 92.1 | 27.6 | 92.6 | 89.8 |
| H | 93 | 93 | 96.3 | 94.6 | 86.4 | 93.2 | 91.8 | ID | 98.9 | 27.5 | 88.9 | 87 |
| I | 93.3 | 93.3 | 96.6 | 94.9 | 86.7 | 93.5 | 92.1 | 98.9 | ID | 27.5 | 89.2 | 87.3 |
| J | 28.6 | 28.6 | 28.3 | 29.3 | 27 | 28.6 | 27.6 | 27.5 | 27.5 | ID | 26.1 | 27.3 |
| K | 93.5 | 93.5 | 89.3 | 91.5 | 93.5 | 93.8 | 92.6 | 88.9 | 89.2 | 26.1 | ID | 93 |
| L | 90.7 | 90.7 | 87.5 | 89 | 99 | 93.5 | 89.8 | 87 | 87.3 | 27.3 | 93 | ID |
| A | B | C | D | E | F | G | H | I | J | K | L | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | ID | 100 | 95 | 95.6 | 91.3 | 98.1 | 98.6 | 95.5 | 95.5 | 28.9 | 94.8 | 91.7 |
| B | 100 | ID | 95 | 95.6 | 91.3 | 98.1 | 98.6 | 95.5 | 95.5 | 28.9 | 94.8 | 91.7 |
| C | 95 | 95 | ID | 97.3 | 88.4 | 95.1 | 94.8 | 97.5 | 97.5 | 28.6 | 91.9 | 88.7 |
| D | 95.6 | 95.6 | 97.3 | ID | 89.6 | 96.3 | 95.6 | 96.4 | 96.4 | 29.2 | 92.7 | 89.7 |
| E | 91.3 | 91.3 | 88.4 | 89.6 | ID | 93.1 | 90.6 | 88.7 | 88.7 | 27.5 | 94.5 | 99.4 |
| F | 98.1 | 98.1 | 95.1 | 96.3 | 93.1 | ID | 97.2 | 95.5 | 95.5 | 28.9 | 95 | 93.4 |
| G | 98.6 | 98.6 | 94.8 | 95.6 | 90.6 | 97.2 | ID | 94.9 | 94.9 | 28.3 | 94 | 90.8 |
| H | 95.5 | 95.5 | 97.5 | 96.4 | 88.7 | 95.5 | 94.9 | ID | 99.5 | 28.3 | 92.2 | 89 |
| I | 95.5 | 95.5 | 97.5 | 96.4 | 88.7 | 95.5 | 94.9 | 99.5 | ID | 28.3 | 92.2 | 89 |
| J | 28.9 | 28.9 | 28.6 | 29.2 | 27.5 | 28.9 | 28.3 | 28.3 | 28.3 | ID | 26.4 | 27.5 |
| K | 94.8 | 94.8 | 91.9 | 92.7 | 94.5 | 95 | 94 | 92.2 | 92.2 | 26.4 | ID | 94.2 |
| L | 91.7 | 91.7 | 88.7 | 89.7 | 99.4 | 93.4 | 90.8 | 89 | 89 | 27.5 | 94.2 | ID |
| B- Cell Epitope Sequence | Length | IEDB Server Score | VaxiJen Score |
|---|---|---|---|
| 18TAESSICSDFGNEFCRNAECEVVPGAE45 | 28 | 0.52 | 0.795 (Probable ANTIGEN) |
| 24DNMYFNAAEKQCEYKDTCKTRECS77 | 24 | 0.54 | Probable NON-ANTIGEN |
| 83QSNP86 | 4 | 0.5 | Probable NON-ANTIGEN |
| 97DTLTLQCNIKDDYATDCRNSGGTAKLRTDGVIG129 | 33 | 0.58 | 1.3361 (Probable ANTIGEN) |
| 136EWGAMNKTTRN146 | 11 | 0.52 | Probable NON-ANTIGEN |
| 152CLRPDLTCKDLCEKNLLQRDSR173 | 22 | 0.55 | Probable NON-ANTIGEN |
| 175C175 | 1 | 1.0 | Probable NON-ANTIGEN |
| 177GWNSPKCSAPADSYCSPGSPKGPDGQCKDACKTKEAG FVCKHGCRSTD224 | 48 | 0.54 | 0.9020 (Probable ANTIGEN) |
| 235FTVAEDGITCKSIPYTGGCTVEQKQTCR262 | 28 | 0.54 | Probable NON-ANTIGEN |
| 280WNQHLVGDKCIGDCVENKCHGEFTDCGVYMNR311 | 32 | 0.536 | 1.3679 (Probable ANTIGEN) |
| 319KSRKPGPNVNINECLLNEYYYTVSFTPNISLDSDHCDWYED RVLEAIRTSIGKEVFKVEILNCTQDIKARLIAEKPLSKHVL RKLQACEHPIGEWCMMYPKLLIKKNSATEIEEENLCDSLLK NQEAAYKGQNKCVKVDNLFWFQCADGYTTTYEMTRGRLR RSVCKAGVSCNENEQLECADK501 | 183 | 0.57 | 0.6052 (Probable ANTIGEN) |
| 507YEN509 | 3 | 0.50 | Probable NON-ANTIGEN |
| 511K511 | 1 | 1.0 | Probable NON-ANTIGEN |
| 519DTKPGEIGCIERTTCNPKEIQECQDKKLECVYKNHK554 | 36 | 0.56 | 0.8751 (Probable ANTIGEN) |
| 563HECSREPAKDSCSEEDNGKCQSSGQ587 | 25 | 0.55 | 1.4514 (Probable ANTIGEN) |
| 598CKEKSEATT606 | 9 | 0.510 | 1.4151 (Probable ANTIGEN) |
| 609TTTTKAKDKDPDPGK623 | 15 | 0.58 | 0.7273 (Probable ANTIGEN) |
| Predicted IVRI-I Bm86 B-cell Epitopes | Yeerongpilly | Thailand M1 | Thailand M2 | Thailand S1 | USA (Hidalgo) | USA (Zapta 1) | USA (Starr 2) | China |
|---|---|---|---|---|---|---|---|---|
| Bm86 | 93% | 96.4% | 96.1% | 96.3% | 97.0% | 93.5% | 93.3% | 92.3% |
| Bm86(T18-D45) | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% |
| Bm86(D97-G129) | 84.8% | 96.9% | 96.9% | 93.9% | 84.8% | 81.8% | 81.8% | 78.7% |
| Bm86(G177-D224) | 82.0% | 84% | 84% | 84% | 97.9% | 84% | 84% | 82% |
| Bm86(W280-R311) | 81.2% | 93.7% | 90.6% | 93.7% | 93.7% | 81.2% | 81.2% | 81.2% |
| Bm86(K319-K501) | 97.8% | 97.2% | 96.7% | 97.8% | 98.9% | 97.2% | 96.7% | 98.3% |
| Bm86(D519-K554) | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 100% |
| Bm86(H563-Q587) | 100% | 100% | 100% | 100% | 100% | 96% | 100% | 100% |
| Bm86(C598-K606) | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 88.8% |
| Bm86(T609-K623) | 100% | 100% | 100% | 100% | 100% | 100% | 100% | 86.6% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parthasarathi, B.C.; Kumar, B.; Nagar, G.; Manjunathachar, H.V.; de la Fuente, J.; Ghosh, S. Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence. Vaccines 2021, 9, 194. https://doi.org/10.3390/vaccines9030194
Parthasarathi BC, Kumar B, Nagar G, Manjunathachar HV, de la Fuente J, Ghosh S. Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence. Vaccines. 2021; 9(3):194. https://doi.org/10.3390/vaccines9030194
Chicago/Turabian StyleParthasarathi, Balasamudram Chandrasekhar, Binod Kumar, Gaurav Nagar, Haranahally Vasanthachar Manjunathachar, José de la Fuente, and Srikant Ghosh. 2021. "Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence" Vaccines 9, no. 3: 194. https://doi.org/10.3390/vaccines9030194
APA StyleParthasarathi, B. C., Kumar, B., Nagar, G., Manjunathachar, H. V., de la Fuente, J., & Ghosh, S. (2021). Analysis of Genetic Diversity in Indian Isolates of Rhipicephalus microplus Based on Bm86 Gene Sequence. Vaccines, 9(3), 194. https://doi.org/10.3390/vaccines9030194