

Therapeutic Potential of Exosomes in Tendon and Tendon–Bone Healing: A Systematic Review of Preclinical Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Methods Literature Retrieval
2.2. Data Extraction
2.3. Assessment of Quality of Studies
2.4. Data Analysis
3. Results
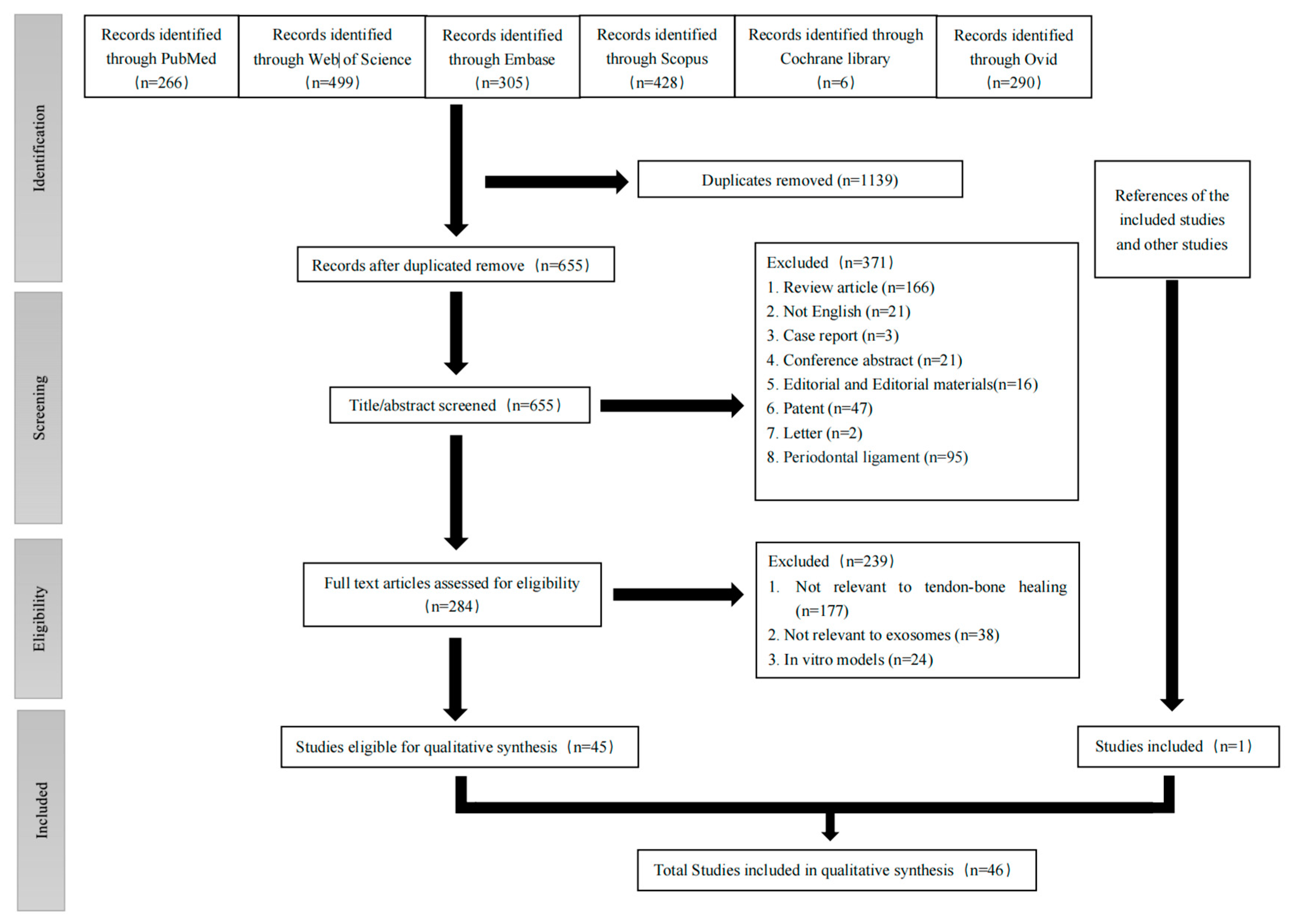
3.1. Search Results
3.2. Assessment of Quality of Studies
3.3. Source of Exosomes
3.4. Isolation and Characterization of Exosomes
3.5. Animal Models
3.6. Group Assignment and Treatment Parameters of Animal Experiments
3.7. Methods of Modification to Improve the Biological Function of Exosomes
3.8. Histological Outcomes
3.9. Biomechanical Outcomes
3.10. Macroscopic Appearance and Morphological Outcomes
3.11. Macrophage Polarization and Regulation of Inflammatory Reaction
3.12. MicroRNAs and Signaling Pathways
3.13. Changes in Gene Expression
3.14. In Vitro Experiment Outcomes (Cell Proliferation, Migration and Differentiation)
4. Discussion
4.1. The Most Suitable Source of Exosomes for Tendon and Tendon–Bone Healing
4.2. Modification of Exosomes or MSCs
4.3. Exosomes Isolation and Administration
4.4. Animal Models
4.5. Mechanism of Exosomes Promoting Tendon Healing
4.6. Mechanism of Exosomes Promoting Tendon–Bone Healing
4.7. Inconsistency and Possible Explanations
4.7.1. Inconsistency between Different Studies
4.7.2. Reasonable Explanations
4.7.3. Future Research Directions
4.8. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviation
References
- Lim, W.L.; Liau, L.L.; Ng, M.H.; Chowdhury, S.R.; Law, J.X. Current Progress in Tendon and Ligament Tissue Engineering. Tissue Eng. Regen. Med. 2019, 16, 549–571. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Rothrauff, B.B.; Tuan, R.S. Tendon and Ligament Regeneration and Repair: Clinical Relevance and Developmental Paradigm. Birth Defects Res. Part C-Embryo Today-Rev. 2013, 99, 203–222. [Google Scholar] [CrossRef]
- Voleti, P.B.; Buckley, M.R.; Soslowsky, L.J. Tendon healing: Repair and regeneration. Annu. Rev. Biomed. Eng. 2012, 14, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Kovacevic, D.; Rodeo, S.A. Biological augmentation of rotator cuff tendon repair. Clin. Orthop. Relat. Res. 2008, 466, 622–633. [Google Scholar] [CrossRef]
- Fang, W.H.; Agrawal, D.K.; Thankam, F.G. “Smart Exosomes”: A Smart Approach for Tendon Regeneration. Tissue Eng. Part B-Rev. 2022, 28, 613–625. [Google Scholar] [CrossRef]
- Chen, Z.; Jin, M.; He, H.; Dong, J.; Li, J.; Nie, J.; Wang, Z.; Xu, J.; Wu, F. Mesenchymal stem cells and macrophages and their interactions in tendon-bone healing. J. Orthop. Translat. 2023, 39, 63–73. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Biology of tendon injury: Healing, modeling and remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar]
- Tao, S.C.; Huang, J.Y.; Li, Z.X.; Zhan, S.; Guo, S.C. Small extracellular vesicles with LncRNA H19 “overload”: YAP Regulation as a Tendon Repair Therapeutic Tactic. iScience 2021, 24, 102200. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Li, J.; Wang, X.; Peng, S.; Ning, J.; Qian, Y.; Fan, C. Microrna-21-3p engineered umbilical cord stem cell-derived exosomes inhibit tendon adhesion. J. Inflamm. Res. 2020, 13, 303–316. [Google Scholar] [CrossRef]
- Longo, U.G.; Franceschi, F.; Berton, A.; Maffulli, N.; Droena, V. Conservative treatment and rotator cuff tear progression. Med. Sport Sci. 2012, 57, 90–99. [Google Scholar] [CrossRef]
- Di Benedetto, P.; Di Benedetto, E.; Fiocchi, A.; Beltrame, A.; Causero, A. Causes of Failure of Anterior Cruciate Ligament Reconstruction and Revision Surgical Strategies. Knee Surg. Relat. Res. 2016, 28, 319–324. [Google Scholar] [CrossRef]
- Galanopoulos, I.; Ilias, A.; Karliaftis, K.; Papadopoulos, D.; Ashwood, N. The Impact of Re-tear on the Clinical Outcome after Rotator Cuff Repair Using Open or Arthroscopic Techniques—A Systematic Review. Open Orthop. J. 2017, 11, 95–107. [Google Scholar] [CrossRef]
- Pache, S.; Del Castillo, J.; Moatshe, G.; LaPrade, R.F. Anterior cruciate ligament reconstruction failure and revision surgery: Current concepts. J. Isakos Jt. Disord. Orthop. Sport. Med. 2020, 5, 351–358. [Google Scholar] [CrossRef]
- Kadakia, A.R.; Dekker, R.G., 2nd; Ho, B.S. Acute Achilles Tendon Ruptures: An Update on Treatment. J. Am. Acad. Orthop. Surg. 2017, 25, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.A.; Yablon, C.M.; Henning, P.T.; Kazmers, I.S.; Urquhart, A.; Hallstrom, B.; Bedi, A.; Parameswaran, A. Greater Trochanteric Pain Syndrome: Percutaneous Tendon Fenestration Versus Platelet-Rich Plasma Injection for Treatment of Gluteal Tendinosis. J. Ultrasound Med. 2016, 35, 2413–2420. [Google Scholar] [CrossRef] [PubMed]
- Kane, S.F.; Olewinski, L.H.; Tamminga, K.S. Management of Chronic Tendon Injuries. Am. Fam. Physician 2019, 100, 147–157. [Google Scholar] [PubMed]
- Skjong, C.C.; Meininger, A.K.; Ho, S.S. Tendinopathy treatment: Where is the evidence? Clin. Sport. Med. 2012, 31, 329–350. [Google Scholar] [CrossRef]
- Chimenti, R.L.; Stover, D.W.; Fick, B.S.; Hall, M.M. Percutaneous Ultrasonic Tenotomy Reduces Insertional Achilles Tendinopathy Pain With High Patient Satisfaction and a Low Complication Rate. J. Ultrasound Med. 2019, 38, 1629–1635. [Google Scholar] [CrossRef]
- Fitzpatrick, J.; Bulsara, M.; Zheng, M.H. The Effectiveness of Platelet-Rich Plasma in the Treatment of Tendinopathy: A Meta-analysis of Randomized Controlled Clinical Trials. Am. J. Sport. Med. 2017, 45, 226–233. [Google Scholar] [CrossRef]
- Killian, M.L. Growth and mechanobiology of the tendon-bone enthesis. Semin. Cell Dev. Biol. 2022, 123, 64–73. [Google Scholar] [CrossRef]
- Roffino, S.; Camy, C.; Foucault-Bertaud, A.; Lamy, E.; Pithioux, M.; Chopard, A. Negative impact of disuse and unloading on tendon enthesis structure and function. Life Sci. Space Res. 2021, 29, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Dang, G.P.; Qin, W.; Wan, Q.Q.; Gu, J.T.; Wang, K.Y.; Mu, Z.; Gao, B.; Jiao, K.; Tay, F.R.; Niu, L.N. Regulation and Reconstruction of Cell Phenotype Gradients Along the Tendon-Bone Interface. Adv. Funct. Mater. 2023, 33, 2210275. [Google Scholar] [CrossRef]
- Friese, N.; Gierschner, M.B.; Schadzek, P.; Roger, Y.; Hoffmann, A. Regeneration of Damaged Tendon-Bone Junctions (Entheses)-TAK1 as a Potential Node Factor. Int. J. Mol. Sci. 2020, 21, 5177. [Google Scholar] [CrossRef] [PubMed]
- Parmar, K. Tendon and ligament: Basic science, injury and repair. Orthop. Trauma 2018, 32, 241–244. [Google Scholar] [CrossRef]
- Qian, S.J.; Wang, Z.; Zheng, Z.F.; Ran, J.S.; Zhu, J.F.; Chen, W.S. A Collagen and Silk Scaffold for Improved Healing of the Tendon and Bone Interface in a Rabbit Model. Med. Sci. Monit. 2019, 25, 269–278. [Google Scholar] [CrossRef]
- Yao, S.Y.; Yung, P.S.H.; Lui, P.P.Y. Tackling the Challenges of Graft Healing After Anterior Cruciate Ligament Reconstruction-Thinking From the Endpoint. Front. Bioeng. Biotechnol. 2021, 9, 1192. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Afif, I.Y.; Maula, M.I.; Winarni, T.I.; Tauviqirrahman, M.; Akbar, I.; Basri, H.; van der Heide, E.; Jamari, J. Tresca Stress Simulation of Metal-on-Metal Total Hip Arthroplasty during Normal Walking Activity. Materials 2021, 14, 7554. [Google Scholar] [CrossRef]
- Ammarullah, M.I.; Hartono, R.; Supriyono, T.; Santoso, G.; Sugiharto, S.; Permana, M.S. Polycrystalline Diamond as a Potential Material for the Hard-on-Hard Bearing of Total Hip Prosthesis: Von Mises Stress Analysis. Biomedicines 2023, 11, 951. [Google Scholar] [CrossRef]
- Vukomanović, M.; Gazvoda, L.; Kurtjak, M.; Maček-Kržmanc, M.; Spreitzer, M.; Tang, Q.; Wu, J.; Ye, H.; Chen, X.; Mattera, M.; et al. Filler-Enhanced Piezoelectricity of Poly-L-Lactide and Its Use as a Functional Ultrasound-Activated Biomaterial. Small 2023, e2301981. [Google Scholar] [CrossRef]
- Chen, C.-H.; Chen, S.-H.; Chen, S.-H.; Chuang, A.D.-C.; Darshan, T.G.; Chen, J.-P. Hyaluronic acid/platelet rich plasma-infused core-shell nanofiber membrane to prevent postoperative tendon adhesion and promote tendon healing. Int. J. Biol. Macromol. 2023, 231, 123312. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, X.; Ng, K.W.; Rao, Y.; Wang, C.Y.; Gharaibeh, B.; Lin, S.E.; Abrams, G.; Safran, M.; Cheung, E.; et al. Growth and differentiation factor-7 immobilized, mechanically strong quadrol-hexamethylene diisocyanate-methacrylic anhydride polyurethane polymer for tendon repair and regeneration. Acta Biomater. 2022, 154, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Chen, J.; Yin, Z.; Chen, X.; Liu, H.; Heng, B.C.; Chen, W.; Ouyang, H.W. Allogenous tendon stem/progenitor cells in silk scaffold for functional shoulder repair. Cell Transpl. 2012, 21, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Darshan, T.G.; Chen, C.-H.; Kuo, C.-Y.; Shalumon, K.T.; Chien, Y.-M.; Kao, H.-H.; Chen, J.-P. Development of high resilience spiral wound suture-embedded gelatin/PCL/heparin nanofiber membrane scaffolds for tendon tissue engineering. Int. J. Biol. Macromol. 2022, 221, 314–333. [Google Scholar] [CrossRef]
- Lui, P.P.; Wong, O.T.; Lee, Y.W. Application of tendon-derived stem cell sheet for the promotion of graft healing in anterior cruciate ligament reconstruction. Am. J. Sport. Med. 2014, 42, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Sato, Y.; Kobayashi, T.; Suzuki, K.; Kimura, A.; Soma, T.; Ito, E.; Kikuchi, T.; Kobayashi, S.; Harato, K.; et al. Adipose-Derived Stem Cell Sheets Improve Early Biomechanical Graft Strength in Rabbits After Anterior Cruciate Ligament Reconstruction. Am. J. Sport. Med. 2021, 49, 3508–3518. [Google Scholar] [CrossRef] [PubMed]
- Herberts, C.A.; Kwa, M.S.; Hermsen, H.P. Risk factors in the development of stem cell therapy. J. Transl. Med. 2011, 9, 29. [Google Scholar] [CrossRef] [PubMed]
- Marks, P.W.; Witten, C.M.; Califf, R.M. Clarifying Stem-Cell Therapy’s Benefits and Risks. N. Engl. J. Med. 2017, 376, 1007–1009. [Google Scholar] [CrossRef]
- Rosland, G.V.; Svendsen, A.; Torsvik, A.; Sobala, E.; McCormack, E.; Immervoll, H.; Mysliwietz, J.; Tonn, J.C.; Goldbrunner, R.; Lonning, P.E.; et al. Long-term Cultures of Bone Marrow-Derived Human Mesenchymal Stem Cells Frequently Undergo Spontaneous Malignant Transformation (This article contains errors due to a cross contamination of the cell lines we used. Cancer Res. 2009, 69, 5331–5339. [Google Scholar] [CrossRef]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef]
- Shao, H.; Im, H.; Castro, C.M.; Breakefield, X.; Weissleder, R.; Lee, H. New Technologies for Analysis of Extracellular Vesicles. Chem. Rev. 2018, 118, 1917–1950. [Google Scholar] [CrossRef]
- Conde-Vancells, J.; Rodriguez-Suarez, E.; Embade, N.; Gil, D.; Matthiesen, R.; Valle, M.; Elortza, F.; Lu, S.C.; Mato, J.M.; Falcon-Perez, J.M. Characterization and comprehensive proteome profiling of exosomes secreted by hepatocytes. J. Proteome Res. 2008, 7, 5157–5166. [Google Scholar] [CrossRef]
- Santiago-Dieppa, D.R.; Steinberg, J.; Gonda, D.; Cheung, V.J.; Carter, B.S.; Chen, C.C. Extracellular vesicles as a platform for ‘liquid biopsy’ in glioblastoma patients. Expert. Rev. Mol. Diagn. 2014, 14, 819–825. [Google Scholar] [CrossRef] [PubMed]
- Dorayappan, K.D.P.; Wallbillich, J.J.; Cohn, D.E.; Selvendiran, K. The biological significance and clinical applications of exosomes in ovarian cancer. Gynecol. Oncol. 2016, 142, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Mallin, K.; Graves, A.J.; Chang, S.S.; Penson, D.F.; Resnick, M.J.; Barocas, D.A. Recent Changes in Prostate Cancer Screening Practices and Epidemiology. J. Urol. 2017, 198, 1230–1240. [Google Scholar] [CrossRef] [PubMed]
- Kapogiannis, D.; Boxer, A.; Schwartz, J.B.; Abner, E.L.; Biragyn, A.; Masharani, U.; Frassetto, L.; Petersen, R.C.; Miller, B.L.; Goetzl, E.J. Dysfunctionally phosphorylated type 1 insulin receptor substrate in neural-derived blood exosomes of preclinical Alzheimer’s disease. Faseb J. 2015, 29, 589–596. [Google Scholar] [CrossRef]
- Zhou, H.; Pisitkun, T.; Aponte, A.; Yuen, P.S.; Hoffert, J.D.; Yasuda, H.; Hu, X.; Chawla, L.; Shen, R.F.; Knepper, M.A.; et al. Exosomal Fetuin-A identified by proteomics: A novel urinary biomarker for detecting acute kidney injury. Kidney Int. 2006, 70, 1847–1857. [Google Scholar] [CrossRef]
- Zhu, Z.; Gao, R.; Ye, T.; Feng, K.; Zhang, J.; Chen, Y.; Xie, Z.; Wang, Y. The Therapeutic Effect of iMSC-Derived Small Extracellular Vesicles on Tendinopathy Related Pain Through Alleviating Inflammation: An in vivo and in vitro Study. J. Inflamm. Res. 2022, 15, 1421–1436. [Google Scholar] [CrossRef]
- Hayashi, Y.; Yimiti, D.; Sanada, Y.; Ding, C.; Omoto, T.; Ogura, T.; Nakasa, T.; Ishikawa, M.; Hiemori, K.; Tateno, H.; et al. The therapeutic capacity of bone marrow MSC-derived extracellular vesicles in Achilles tendon healing is passage-dependent and indicated by specific glycans. FEBS Lett. 2022, 596, 1047–1058. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, H.; Cui, Q.; Han, P.; Yang, S.; Shi, M.; Zhang, T.; Zhang, Z.; Li, Z. Tendon stem cell-derived exosomes regulate inflammation and promote the high-quality healing of injured tendon. Stem Cell Res. Ther. 2020, 11, 402. [Google Scholar] [CrossRef]
- Shen, H.; Yoneda, S.; Abu-Amer, Y.; Guilak, F.; Gelberman, R. Stem cell-derived extracellular vesicles attenuate the early inflammatory response after tendon injury and repair. J. Orthop. Res. 2019, 38, 117–127. [Google Scholar] [CrossRef]
- Gao, R.; Ye, T.; Zhu, Z.; Li, Q.; Zhang, J.; Yuan, J.; Zhao, B.; Xie, Z.; Wang, Y. Small extracellular vesicles from iPSC-derived mesenchymal stem cells ameliorate tendinopathy pain by inhibiting mast cell activation. Nanomedicine 2022, 17, 513–529. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Wang, Y.; Wang, Z.; Thoreson, A.R.; Jacobson, D.S.; Amadio, P.C.; Behfar, A.; Moran, S.L.; Zhao, C. A novel engineered purified exosome product patch for tendon healing: An explant in an ex vivo model. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2020, in press. [CrossRef] [PubMed]
- Liu, A.; Wang, Q.; Zhao, Z.; Wu, R.; Wang, M.; Li, J.; Sun, K.; Sun, Z.; Lv, Z.; Xu, J.; et al. Nitric Oxide Nanomotor Driving Exosomes-Loaded Microneedles for Achilles Tendinopathy Healing. ACS Nano 2021, 15, 13339–13350. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.R.; Garcia, S.; Liu, M.; Chi, H.; Kim, H.T.; Raffai, R.L.; Liu, X.; Feeley, B.T. Muscle-Derived Beige Adipose Precursors Secrete Promyogenic Exosomes That Treat Rotator Cuff Muscle Degeneration in Mice and Are Identified in Humans by Single-Cell RNA Sequencing. Am. J. Sport. Med. 2022, 50, 2247–2257. [Google Scholar] [CrossRef]
- Yao, Z.; Li, J.; Xiong, H.; Cui, H.; Ning, J.; Wang, S.; Ouyang, X.; Qian, Y.; Fan, C. MicroRNA engineered umbilical cord stem cell-derived exosomes direct tendon regeneration by mTOR signaling. J. Nanobiotechnol. 2021, 19, 169. [Google Scholar] [CrossRef]
- Cui, H.; He, Y.; Chen, S.; Zhang, D.; Yu, Y.; Fan, C. Macrophage-Derived miRNA-Containing Exosomes Induce Peritendinous Fibrosis after Tendon Injury through the miR-21-5p/Smad7 Pathway. Mol. Ther.-Nucleic Acids 2019, 14, 114–130. [Google Scholar] [CrossRef]
- Zhang, X.; Han, Z.; Han, K.; Zhang, H.; Huang, J.; Huangfu, X.; Zhao, J. Loading Mesenchymal Stem Cell-Derived Exosomes Into a Traditionally Designed Rotator Cuff Patch: A Potential Strategy to Enhance the Repair of Chronic Rotator Cuff Tear Associated With Degenerative Changes. Am. J. Sport. Med. 2022, 50, 2234–2246. [Google Scholar] [CrossRef]
- Ye, T.; Chen, Z.; Zhang, J.; Luo, L.; Gao, R.; Gong, L.; Du, Y.; Xie, Z.; Zhao, B.; Li, Q.; et al. Large extracellular vesicles secreted by human iPSC-derived MSCs ameliorate tendinopathy via regulating macrophage heterogeneity. Bioact. Mater. 2023, 21, 194–208. [Google Scholar] [CrossRef]
- Wellings, E.P.; Huang, T.C.T.; Li, J.; Peterson, T.E.; Hooke, A.W.; Rosenbaum, A.; Zhao, C.D.; Behfar, A.; Moran, S.L.; Houdek, M.T. Intrinsic Tendon Regeneration After Application of Purified Exosome Product: An In Vivo Study. Orthop. J. Sport. Med. 2021, 9, 23259671211062929. [Google Scholar] [CrossRef]
- Wang, C.; Tan, J.; Zhang, Y.; Chen, D.; He, Y. In Situ-Forming Fibrin Gel Encapsulation of MSC-Exosomes for Partial-Thickness Rotator Cuff Tears in a Rabbit Model: Effectiveness Shown in Preventing Tear Progression and Promoting Healing. J. Bone Jt. Surg. 2022, 104, 1492–1502. [Google Scholar] [CrossRef]
- Chen, S.H.; Chen, Z.Y.; Lin, Y.H.; Chen, S.H.; Chou, P.Y.; Kao, H.K.; Lin, F.H. Extracellular vesicles of adipose-derived stem cells promote the healing of traumatized achilles tendons. Int. J. Mol. Sci. 2021, 22, 12373. [Google Scholar] [CrossRef] [PubMed]
- Gissi, C.; Radeghieri, A.; Lamorgese Passeri, C.A.; Gallorini, M.; Calciano, L.; Oliva, F.; Veronesi, F.; Zendrini, A.; Cataldi, A.; Bergese, P.; et al. Extracellular vesicles from rat-bone-marrow mesenchymal stromal/stem cells improve tendon repair in rat Achilles tendon injury model in dose-dependent manner: A pilot study. PLoS ONE 2020, 15, e0229914. [Google Scholar] [CrossRef]
- Li, J.; Yao, Z.; Xiong, H.; Cui, H.; Wang, X.; Zheng, W.; Qian, Y.; Fan, C. Extracellular vesicles from hydroxycamptothecin primed umbilical cord stem cells enhance anti-adhesion potential for treatment of tendon injury. Stem Cell Res. Ther. 2020, 11, 500. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, C.S.; Clements, A.E.B.; Kink, J.A.; Choi, U.; Baer, G.S.; Halanski, M.A.; Hematti, P.; Vanderby, R. Extracellular Vesicle-Educated Macrophages Promote Early Achilles Tendon Healing. Stem Cells 2019, 37, 652–662. [Google Scholar] [CrossRef]
- Wang, C.; Song, W.; Chen, B.; Liu, X.; He, Y. Exosomes Isolated From Adipose-Derived Stem Cells: A New Cell-Free Approach to Prevent the Muscle Degeneration Associated With Torn Rotator Cuffs. Am. J. Sport. Med. 2019, 47, 3247–3255. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; He, G.; Guo, Y.; Tang, H.; Shi, Y.; Bian, X.; Zhu, M.; Kang, X.; Zhou, M.; Lyu, J.; et al. Exosomes from tendon stem cells promote injury tendon healing through balancing synthesis and degradation of the tendon extracellular matrix. J. Cell. Mol. Med. 2019, 23, 5475–5485. [Google Scholar] [CrossRef]
- Song, K.; Jiang, T.; Pan, P.; Yao, Y.; Jiang, Q. Exosomes from tendon derived stem cells promote tendon repair through miR-144-3p-regulated tenocyte proliferation and migration. Stem Cell Res. Ther. 2022, 13, 80. [Google Scholar] [CrossRef]
- Yu, Y.; Sun, B.; Wang, Z.; Yang, M.; Cui, Z.; Lin, S.; Jin, M.; Yi, C. Exosomes From M2 Macrophage Promote Peritendinous Fibrosis Posterior Tendon Injury via the MiR-15b-5p/FGF-1/7/9 Pathway by Delivery of circRNA-Ep400. Front. Cell Dev. Biol. 2021, 9, 595911. [Google Scholar] [CrossRef]
- Han, Q.; Wang, S.; Chen, D.; Gan, D.; Wang, T. Exosomes derived from human umbilical cord mesenchymal stem cells reduce tendon injuries via the miR-27b-3p/ARHGAP5/RhoA signaling pathway. Acta Biochim. Et. Biophys. Sin. 2022, 54, 232. [Google Scholar] [CrossRef]
- Jenner, F.; Wagner, A.; Gerner, I.; Ludewig, E.; Trujanovic, R.; Rohde, E.; von Rechenberg, B.; Gimona, M.; Traweger, A. Evaluation of the Potential of Umbilical Cord Mesenchymal Stromal Cell-Derived Small Extracellular Vesicles to Improve Rotator Cuff Healing: A Pilot Ovine Study. Am. J. Sport. Med. 2023, 51, 03635465221145958. [Google Scholar] [CrossRef]
- Li, X.; Su, Z.; Shen, K.; Wang, Q.; Xu, C.; Wang, F.; Zhang, Y.; Jiang, D. Eugenol-Preconditioned Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Antioxidant Capacity of Tendon Stem Cells in Vitro and in Vivo. Oxidative Med. Cell. Longev. 2022, 2022, 3945195. [Google Scholar] [CrossRef]
- Xu, T.; Lin, Y.; Yu, X.; Jiang, G.; Wang, J.; Xu, K.; Fang, J.; Wang, S.; Dai, X. Comparative Effects of Exosomes and Ectosomes Isolated From Adipose-Derived Mesenchymal Stem Cells on Achilles Tendinopathy in a Rat Model. Am. J. Sport. Med. 2022, 50, 2740–2752. [Google Scholar] [CrossRef]
- Yu, H.; Cheng, J.; Shi, W.; Ren, B.; Zhao, F.; Shi, Y.; Yang, P.; Duan, X.; Zhang, J.; Fu, X.; et al. Bone marrow mesenchymal stem cell-derived exosomes promote tendon regeneration by facilitating the proliferation and migration of endogenous tendon stem/progenitor cells. Acta Biomater. 2020, 106, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhu, Y.; Hsiao, A.W.T.; Xu, J.; Tong, W.; Chang, L.; Zhang, X.; Chen, Y.F.; Li, J.; Chen, W.; et al. Bioactive glass-elicited stem cell-derived extracellular vesicles regulate M2 macrophage polarization and angiogenesis to improve tendon regeneration and functional recovery. Biomaterials 2023, 294, 121998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, A.; Han, K.; Zhang, H.; Huangfu, X.; Huang, J.; Jiang, J.; Zhao, J. Anti-inflammatory and Tendon-Protective Effects of Adipose Stem Cell-Derived Exosomes with Concomitant Use of Glucocorticoids. Stem Cells Int. 2022, 2022, 1455226. [Google Scholar] [CrossRef] [PubMed]
- Fu, G.; Lu, L.; Pan, Z.; Fan, A.; Yin, F. Adipose-derived stem cell exosomes facilitate rotator cuff repair by mediating tendon-derived stem cells. Regen. Med. 2021, 16, 359–372. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, M.; Shi, M.; Zhang, T.; Lu, W.; Yang, S.; Cui, Q.; Li, Z. Adipose-derived mesenchymal stromal cell-derived exosomes promote tendon healing by activating both SMAD1/5/9 and SMAD2/3. Stem Cell Res. Ther. 2021, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Jiang, H.; Zhang, H.; Sun, Z.; Lin, Q.; Wang, T.; Yu, T.; Zhang, Y. Adipose-Derived Mesenchymal Stem Cell-Derived Extracellular Vesicles Rescue Tendon Injury in Rat via the miR-19 a/IGFBP3 Axis. Stem Cells Int. 2022, 2022, 197473. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.; Zhang, G.; Yu, W.; He, Y. Adipose Stem Cell-Derived Exosomes Ameliorate Chronic Rotator Cuff Tendinopathy by Regulating Macrophage Polarization: From a Mouse Model to a Study in Human Tissue. Am. J. Sport. Med. 2021, 49, 2321–2331. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Qian, J.; Yang, M.-Y.; He, Y.; Tao, X.; Shi, Y.-X.; Bian, X.-T.; Chen, W.; Wang, Y.-J.; Wang, H.; et al. MiR-6924-5p-rich exosomes derived from genetically modified Scleraxis-overexpressing PDGFR alpha(+) BMMSCs as novel nanotherapeutics for treating osteolysis during tendon-bone healing and improving healing strength. Biomaterials 2021, 279, 121242. [Google Scholar] [CrossRef]
- Xu, J.; Ye, Z.; Han, K.; Zheng, T.; Zhang, T.; Dong, S.; Jiang, J.; Yan, X.; Cai, J.; Zhao, J. Infrapatellar Fat Pad Mesenchymal Stromal Cell-Derived Exosomes Accelerate Tendon-Bone Healing and Intra-articular Graft Remodeling After Anterior Cruciate Ligament Reconstruction. Am. J. Sport. Med. 2022, 50, 662–673. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Yan, S.; Song, Y.; Chen, C.; Xu, D.; Lu, B.; Xu, Y. Exosomes secreted by hypoxia-stimulated bone-marrow mesenchymal stem cells promote grafted tendon-bone tunnel healing in rat anterior cruciate ligament reconstruction model. J. Orthop. Transl. 2022, 36, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.D.; Kang, L.; Tian, J.; Wu, Y.; Huang, Y.; Liu, J.; Wang, H.; Qiu, G.; Wu, Z. Exosomes derived from magnetically actuated bone mesenchymal stem cells promote tendon-bone healing through the miR-21-5p/SMAD7 pathway. Mater. Today Bio. 2022, 15, 100319. [Google Scholar] [CrossRef]
- Shi, Y.; Kang, X.; Wang, Y.; Bian, X.; He, G.; Zhou, M.; Tang, K. Exosomes derived from bone marrow stromal cells (BMSCs) enhance tendon-bone healing by regulating macrophage polarization. Med. Sci. Monit. 2020, 26, e923328-1. [Google Scholar] [CrossRef]
- Wu, B.; Chen, H.; Shi, X.; Wang, L.; Zhang, T.; Guan, C.; Huang, T.; Yang, Y.; Hu, J.; Lu, H. Exosomes Derived from Bone Marrow Mesenchymal Stem Cell Preconditioned by Low-Intensity Pulsed Ultrasound Stimulation Promote Bone-Tendon Interface Fibrocartilage Regeneration and Ameliorate Rotator Cuff Fatty Infiltration. Med. Sci. Monit. 2021; preprint (Version 1). [Google Scholar] [CrossRef]
- Han, L.; Liu, H.; Fu, H.; Hu, Y.; Fang, W.; Liu, J. Exosome-delivered BMP-2 and polyaspartic acid promotes tendon bone healing in rotator cuff tear via Smad/RUNX2 signaling pathway. Bioengineered 2022, 13, 1459–1475. [Google Scholar] [CrossRef]
- Ren, Y.; Zhang, S.; Wang, Y.; Jacobson, D.S.; Reisdorf, R.L.; Kuroiwa, T.; Behfar, A.; Moran, S.L.; Steinmann, S.P.; Zhao, C. Effects of purified exosome product on rotator cuff tendon-bone healing in vitro and in vivo. Biomaterials 2021, 276, 121019. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; He, B.; Wang, L.; Yuan, B.; Shu, H.; Zhang, F.; Sun, L. Bone marrow mesenchymal stem cell-derived exosomes promote rotator cuff tendon-bone healing by promoting angiogenesis and regulating M1 macrophages in rats. Stem Cell Res. Ther. 2020, 11, 496. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Q.; Tong, K.; Zhu, J.; Wang, H.; Chen, B.; Chen, L. BMSC-derived exosomes promote tendon-bone healing after anterior cruciate ligament reconstruction by regulating M1/M2 macrophage polarization in rats. Stem Cell Res. Ther. 2022, 13, 295. [Google Scholar] [CrossRef]
- Wang, C.; Hu, Q.; Song, W.; Yu, W.; He, Y. Adipose Stem Cell-Derived Exosomes Decrease Fatty Infiltration and Enhance Rotator Cuff Healing in a Rabbit Model of Chronic Tears. Am. J. Sport. Med. 2020, 48, 1456–1464. [Google Scholar] [CrossRef]
- Ning, C.; Li, P.; Gao, C.; Fu, L.; Liao, Z.; Tian, G.; Yin, H.; Li, M.; Sui, X.; Yuan, Z.; et al. Recent advances in tendon tissue engineering strategy. Front. Bioeng. Biotechnol. 2023, 11, 1115312. [Google Scholar] [CrossRef]
- Polly, S.S.; Nichols, A.E.C.; Donnini, E.; Inman, D.J.; Scott, T.J.; Apple, S.M.; Werre, S.R.; Dahlgren, L.A. Adipose-Derived Stromal Vascular Fraction and Cultured Stromal Cells as Trophic Mediators for Tendon Healing. J. Orthop. Res. 2019, 37, 1429–1439. [Google Scholar] [CrossRef]
- Titan, A.L.; Foster, D.S.; Chang, J.; Longaker, M.T. Flexor Tendon: Development, Healing, Adhesion Formation, and Contributing Growth Factors. Plast. Reconstr. Surg. 2019, 144, 639e–647e. [Google Scholar] [CrossRef]
- Chen, X.; Jones, I.A.; Park, C.; Vangsness, C.T., Jr. The Efficacy of Platelet-Rich Plasma on Tendon and Ligament Healing: A Systematic Review and Meta-analysis With Bias Assessment. Am. J. Sport. Med. 2018, 46, 2020–2032. [Google Scholar] [CrossRef]
- Ruiz-Alonso, S.; Lafuente-Merchan, M.; Ciriza, J.; Saenz-Del-Burgo, L.; Pedraz, J.L. Tendon tissue engineering: Cells, growth factors, scaffolds and production techniques. J. Control. Release 2021, 333, 448–486. [Google Scholar] [CrossRef]
- Harding, J.; Mirochnitchenko, O. Preclinical studies for induced pluripotent stem cell-based therapeutics. J. Biol. Chem. 2014, 289, 4585–4593. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. PLoS Med. 2021, 18, 105906. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Shim, I.K.; Choi, J.H.; Lee, E.S.; Lee, Y.N.; Jeon, I.H.; Koh, K.H. Extracellular Vesicles Delivered by Injectable Collagen Promote Bone–Tendon Interface Healing and Prevent Fatty Degeneration of Rotator Cuff Muscle. Arthrosc.-J. Arthrosc. Relat. Surg. 2022, 38, 2142–2153. [Google Scholar] [CrossRef]
- Shi, Z.; Wang, Q.; Jiang, D. Extracellular vesicles from bone marrow-derived multipotent mesenchymal stromal cells regulate inflammation and enhance tendon healing. J. Transl. Med. 2019, 17, 211. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Su, Q.; Li, J.; Xue, C.; Zhu, J.; Cai, Q.; Huang, J.; Ji, S.; Cheng, B.; Ge, H. NAMPT encapsulated by extracellular vesicles from young adipose-derived mesenchymal stem cells treated tendinopathy in a “One-Stone-Two-Birds” manner. J. Nanobiotechnol. 2023, 21, 7. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; de Vries, R.B.M.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. Bmc Med. Res. Methodol. 2014, 14, 43. [Google Scholar] [CrossRef]
- Torres-Torrillas, M.; Rubio, M.; Damia, E.; Cuervo, B.; Del Romero, A.; Peláez, P.; Chicharro, D.; Miguel, L.; Sopena, J.J. Adipose-Derived Mesenchymal Stem Cells: A Promising Tool in the Treatment of Musculoskeletal Diseases. Int. J. Mol. Sci. 2019, 20, 3105. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Lui, P.P.; Rui, Y.F.; Wong, Y.M. Comparison of potentials of stem cells isolated from tendon and bone marrow for musculoskeletal tissue engineering. Tissue Eng. Part A 2012, 18, 840–851. [Google Scholar] [CrossRef]
- Capelli, C.; Gotti, E.; Morigi, M.; Rota, C.; Weng, L.; Dazzi, F.; Spinelli, O.; Cazzaniga, G.; Trezzi, R.; Gianatti, A.; et al. Minimally manipulated whole human umbilical cord is a rich source of clinical-grade human mesenchymal stromal cells expanded in human platelet lysate. Cytotherapy 2011, 13, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Lorenzo, M.J.; Royo-Cañas, M.; Alegre-Aguarón, E.; Desportes, P.; Castiella, T.; García-Alvarez, F.; Larrad, L. Phenotype and chondrogenic differentiation of mesenchymal cells from adipose tissue of different species. J. Orthop. Res. 2009, 27, 1499–1507. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.J.; Lee, M.; Im, W.; Kim, M. Low pH increases the yield of exosome isolation. Biochem. Biophys. Res. Commun. 2015, 461, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Wu, F.; Xue, E.; Huang, L.; Yan, P.; Pan, X.; Zhou, Y. Hypoxia preconditioning promotes bone marrow mesenchymal stem cells survival by inducing HIF-1α in injured neuronal cells derived exosomes culture system. Cell Death Dis. 2019, 10, 134. [Google Scholar] [CrossRef]
- Bu, H.; He, D.; He, X.; Wang, K. Exosomes: Isolation, Analysis, and Applications in Cancer Detection and Therapy. Chembiochem 2019, 20, 451–461. [Google Scholar] [CrossRef]
- Alvarez, M.L.; Khosroheidari, M.; Kanchi Ravi, R.; DiStefano, J.K. Comparison of protein, microRNA, and mRNA yields using different methods of urinary exosome isolation for the discovery of kidney disease biomarkers. Kidney Int. 2012, 82, 1024–1032. [Google Scholar] [CrossRef]
- Nordin, J.Z.; Lee, Y.; Vader, P.; Mäger, I.; Johansson, H.J.; Heusermann, W.; Wiklander, O.P.; Hällbrink, M.; Seow, Y.; Bultema, J.J.; et al. Ultrafiltration with size-exclusion liquid chromatography for high yield isolation of extracellular vesicles preserving intact biophysical and functional properties. Nanomedicine 2015, 11, 879–883. [Google Scholar] [CrossRef]
- Kang, D.; Oh, S.; Ahn, S.M.; Lee, B.H.; Moon, M.H. Proteomic analysis of exosomes from human neural stem cells by flow field-flow fractionation and nanoflow liquid chromatography-tandem mass spectrometry. J. Proteome Res. 2008, 7, 3475–3480. [Google Scholar] [CrossRef]
- Coumans, F.A.; van der Pol, E.; Böing, A.N.; Hajji, N.; Sturk, G.; van Leeuwen, T.G.; Nieuwland, R. Reproducible extracellular vesicle size and concentration determination with tunable resistive pulse sensing. J. Extracell. Vesicles 2014, 3, 25922. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Fraser, K.; Ghaddar, B.; Yang, K.; Kim, E.; Balaj, L.; Chiocca, E.A.; Breakefield, X.O.; Lee, H.; Weissleder, R. Multiplexed Profiling of Single Extracellular Vesicles. ACS Nano 2018, 12, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T., Jr.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- Yuana, Y.; Koning, R.I.; Kuil, M.E.; Rensen, P.C.; Koster, A.J.; Bertina, R.M.; Osanto, S. Cryo-electron microscopy of extracellular vesicles in fresh plasma. J. Extracell. Vesicles 2013, 2, 21494. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef]
- Sunwoo, J.Y.; Eliasberg, C.D.; Carballo, C.B.; Rodeo, S.A. The role of the macrophage in tendinopathy and tendon healing. J. Orthop. Res. 2020, 38, 1666–1675. [Google Scholar] [CrossRef]
- Moqbel, S.A.A.; Xu, K.; Chen, Z.; Xu, L.; He, Y.; Wu, Z.; Ma, C.; Ran, J.; Wu, L.; Xiong, Y. Tectorigenin Alleviates Inflammation, Apoptosis, and Ossification in Rat Tendon-Derived Stem Cells via Modulating NF-Kappa B and MAPK Pathways. Front. Cell Dev. Biol. 2020, 8, 568894. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef]
- Galatz, L.M.; Sandell, L.J.; Rothermich, S.Y.; Das, R.; Mastny, A.; Havlioglu, N.; Silva, M.J.; Thomopoulos, S. Characteristics of the rat supraspinatus tendon during tendon-to-bone healing after acute injury. J. Orthop. Res. 2006, 24, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Chen, Z.; Wu, T.; Li, R.; Chen, C.; Liu, J.; Hou, H.; Zheng, X.; Wang, H. VEGFA-Enriched Exosomes from Tendon-Derived Stem Cells Facilitate Tenocyte Differentiation, Migration, and Transition to a Fibroblastic Phenotype. BioMed Res. Int. 2022, 2022, 537959. [Google Scholar] [CrossRef] [PubMed]
- Koch, D.W.; Schnabel, L.V.; Ellis, I.M.; Bates, R.E.; Berglund, A.K. TGF-β2 enhances expression of equine bone marrow-derived mesenchymal stem cell paracrine factors with known associations to tendon healing. Stem Cell Res. Ther. 2022, 13, 477. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.H.; Sun, Y.H. TGF-β1 stimulated mesenchymal stem cells-generated exosomal miR-29a promotes the proliferation, migration and fibrogenesis of tenocytes by targeting FABP3. Cytokine 2023, 162, 156090. [Google Scholar] [CrossRef] [PubMed]
- Weinreb, J.H.; Sheth, C.; Apostolakos, J.; McCarthy, M.B.; Barden, B.; Cote, M.P.; Mazzocca, A.D. Tendon structure, disease, and imaging. Muscles Ligaments Tendons J. 2014, 4, 66–73. [Google Scholar] [CrossRef]
- Thankam, F.G.; Evan, D.K.; Agrawal, D.K.; Dilisio, M.F. Collagen type III content of the long head of the biceps tendon as an indicator of glenohumeral arthritis. Mol. Cell. Biochem. 2019, 454, 25–31. [Google Scholar] [CrossRef]
- Tsai, W.C.; Pang, J.H.; Hsu, C.C.; Chu, N.K.; Lin, M.S.; Hu, C.F. Ultrasound stimulation of types I and III collagen expression of tendon cell and upregulation of transforming growth factor beta. J. Orthop. Res. 2006, 24, 1310–1316. [Google Scholar] [CrossRef]
- Blitz, E.; Sharir, A.; Akiyama, H.; Zelzer, E. Tendon-bone attachment unit is formed modularly by a distinct pool of Scx- and Sox9-positive progenitors. Development 2013, 140, 2680–2690. [Google Scholar] [CrossRef]
- Felsenthal, N.; Rubin, S.; Stern, T.; Krief, S.; Pal, D.; Pryce, B.A.; Schweitzer, R.; Zelzer, E. Development of migrating tendon-bone attachments involves replacement of progenitor populations. Development 2018, 145, dev165381. [Google Scholar] [CrossRef]
- Du, L.; Qin, C.; Zhang, H.; Han, F.; Xue, J.; Wang, Y.; Wu, J.; Xiao, Y.; Huan, Z.; Wu, C. Multicellular Bioprinting of Biomimetic Inks for Tendon-to-Bone Regeneration. Adv. Sci. (Weinh) 2023, e2301309. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Song, Z.; Chen, D.; Album, Z.; Green, S.; Deng, X.; Rodeo, S.A. GLI1 Deficiency Impairs the Tendon-Bone Healing after Anterior Cruciate Ligament Reconstruction: In Vivo Study Using Gli1-Transgenic Mice. J. Clin. Med. 2023, 12, 999. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Kim, S.; Lin, B.; Wu, B.M.; Lee, M. Cartilaginous Extracellular Matrix-Modified Chitosan Hydrogels for Cartilage Tissue Engineering. Acs Appl. Mater. Interfaces 2014, 6, 20110–20121. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Bliss, J.P.; Bruce, W.J.A.; Walsh, W.R. Bone morphogenetic proteins and Smad expression in ovine tendon-bone healing. Arthrosc.-J. Arthrosc. Relat. Surg. 2007, 23, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Hjorthaug, G.A.; Søreide, E.; Nordsletten, L.; Madsen, J.E.; Reinholt, F.P.; Niratisairak, S.; Dimmen, S. Negative effect of zoledronic acid on tendon-to-bone healing. Acta Orthop. 2018, 89, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.N.; Zhang, Q.Q.; Geng, M.R.; Wang, W.Z.; Yang, J.; Khan, A.U.R.; Du, H.B.; Sha, Z.; Zhou, X.J.; He, C.L. Construction of nanofibrous scaffolds with interconnected perfusable microchannel networks for engineering of vascularized bone tissue. Bioact. Mater. 2021, 6, 3254–3268. [Google Scholar] [CrossRef]
- Wynn, T.A.; Ramalingam, T.R. Mechanisms of fibrosis: Therapeutic translation for fibrotic disease. Nat. Med. 2012, 18, 1028–1040. [Google Scholar] [CrossRef]
- Jiang, D.; Xu, C.; Li, Z.; Zhang, Y.; Han, F.; Jiang, Z. Protective action of hepatocyte growth factor on transforming growth factor beta-1-induced alpha-smooth muscle actin and extracellular matrix in cultured human peritoneal fibroblasts. Med. Sci. Monit. 2010, 16, Br250–Br254. [Google Scholar]


{kind=link}
| Injury Types | Treatment Options |
|---|---|
| Acute tendon injury | Existing treatment options for acute tendon injury are mostly surgical treatment. Surgical treatment includes open, minimally invasive, and percutaneous repair techniques [14]. Currently, biological agents such as platelet-rich plasma are also used in surgical treatment to promote postoperative recovery of tendons [15]. |
| Chronic tendon injury | Existing treatment options for chronic tendon injury include activity modification, relative rest, drug treatment and rehabilitative exercise [16]. Traditional drugs used to treat chronic tendon injuries include non-steroidal anti-inflammatory drugs (NSAIDs), corticosteroids and topical nitroglycerin [17]. New methods such as ultrasound-guided debridement and platelet-rich plasma are also used to address chronic tendon injuries [18,19]. |
| Cellular Therapy Options | Overview of the Options |
|---|---|
| Mesenchymal stem cells (MSCs) | MSCs are of great significance for tendon regeneration. MSCs have a wide range of sources and can be obtained from tissues such as fat, umbilical cord, and bone marrow. MSCs promote tendon regeneration by secreting cytokines, regulating inflammation, and differentiating into tendon cells [91]. Currently, MSCs used for treatment include Adipose-derived mesenchymal stem cells (ASCs), Bone marrow mesenchymal stem cells (BMSCs), and tendon stem cells (TSCs). The conventional vacuum fraction (SVF) also contains a certain amount of MSCs, which are currently being studied [92]. |
| Growth factors | Growth factors can promote cell proliferation and differentiation, and stimulate the synthesis of extracellular matrix (ECM), which are beneficial for tendon regeneration. Common growth factors related to tendon regeneration include insulin-like growth factor 1 (IGF-1), vascular endothelial growth factor (VEGF), bone morphogenetic proteins (BMPs) and so on [93]. |
| Platelet-rich plasma (PRP) | PRP contains various growth factors that can promote tendon regeneration by promoting cell proliferation and angiogenesis [94]. PRP has certain clinical application potential. |
| Extracellular vesicles (EVs) | Currently, the EVs mainly used for studying the promotion of tissue regeneration are exosomes. EVs contain a variety of proteins, lipids and nucleic acids such as microRNA (miRNA), mRNA and DNA, which play an important role in cell-to-cell communication [40]. |
| Differentiated cells | Differentiated cells include Tenocytes and fibroblasts. Differentiated cells have strong proliferation and differentiation abilities, and they will not lead to teratoma, which is their major advantage [95]. |
| Pluripotent stem cells (PSCs) | PSCs include embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs), they have a strong ability for proliferation and differentiation. PSCs differentiate to functional tendon cells to promote tendon regeneration [96]. |
| Author | Year | Animal | Sample Size | Gender | Animal Model |
|---|---|---|---|---|---|
| Chamberlain et al. [64] | 2019 | Nude mice | 27 | Male | The SDF was removed from the Achilles tendon, and the Achilles tendon was completely transected at the midpoint, followed by repair. |
| Chen et al. [61] | 2021 | NZ rabbits | NR | NR | The Achilles tendon was transected, followed by repair with the Modified Kessler four-core technique. |
| Cui et al. [56] | 2019 | C57BL/6J mice | NR | Male | Complete transection and repair of the FDL tendon were conducted in the right hind paw |
| Davies et al. [54] | 2022 | C57BL/6J mice | 12 | NR | The supraspinatus and infraspinatus tendons were transected and the suprascapular nerve was identified and resected. |
| Fu et al. [76] | 2021 | SD rats | 96 | NR | The rats’ supraspinatus tendon was cut off on Bilateral shoulders and cartilage was worn down. Repair was conducted 4 weeks later. |
| Gao et al. [51] | 2022 | SD rats | 9 | Male | Carrageenan was injected into quadriceps tendon under ultrasound guidance. |
| Gissi et al. [62] | 2020 | Lewis rats | 16 | Male | A bilateral Achilles tendon defect 2 mm in diameter was conducted. |
| Han et al. [69] | 2022 | SD rats | 18 | Male | The superficial Achilles tendon was removed, and transection was made in the middle of the deep Achilles tendon, followed by repair. |
| Hayashi et al. [48] | 2022 | C57BL/6J mice | 39 | Male | A transverse incision was made at the midpoint of the Achilles tendon with the scissors. |
| Jenner et al. [70] | 2023 | Sheep | 12 | NR | A full-thickness defect was created in the center of the infraspinatus tendon at the enthesis. |
| Li et al. [63] | 2020 | SD rats | 33 | Male | The thin strand of the Achilles tendon was resected, and the thick strand was transected in the middle, followed by repair of the tendon. |
| Li et al. [71] | 2022 | SD rats | 64 | Male | One-third of the central part of the patellar tendon was removed. |
| Liu et al. [77] | 2021 | SD rats | 63 | Male | One-third of the central part of the patellar tendon was removed. |
| Liu et al. [53] | 2021 | Rats | 20 | NR | Type I collagenase solution was injected into the right hind leg of the rats every 2 days for 2weeks. |
| Shen et al. [50] | 2019 | NGL mice | 32 | Both sexes | At the midpoint between the musculotendinous junction of the Achilles tendon and the calcaneal insertion, the Achilles tendon was transected. |
| Shi et al. [52] | 2020 | Dogs | 48 | NR | The tendon was transected in the middle of a 30mm length centered at the proximal interphalangeal joint level. |
| Song et al. [67] | 2022 | SD rats | 72 | Male | One-third of the central part of the patellar tendon was removed |
| Tao et al. [8] | 2021 | SD rats | 96 | Male | One third of the central part of the left knee patellar tendon was removed. |
| Wang et al. [60] | 2022 | NZ rabbits | 112 | Male | The bursal-side defects on the supraspinatus tendon were made to establish the 50% PTRCT model. |
| Wang et al. [65] | 2019 | SD rats | 42 | Female | The supraspinatus and infraspinatus tendons were cut close to the greater tuberosity of the humerus. |
| Wang et al. [66] | 2019 | SD rats | 18 | Male | Type I collagenase solution was injected into Achilles tendons. |
| Wang et al. [79] | 2021 | C57BL/6J mice | 72 | Male | The mice were trained on a treadmill for 1 week. |
| Wellings et al. [59] | 2021 | NZ rabbits | 45 | Female | The Achilles tendon bundle was isolated and transected approximately 1.5 cm proximal to the calcaneal tubercle. Then, tendon was repaired with Kessler core suture technique. |
| Xu et al. [72] | 2022 | SD rats | 36 | Male | Type I collagenase solution was injected into the Achilles tendon. |
| Xu et al. [74] | 2023 | SD rats | 100 | Male | Achilles tendon was completely full-thickness ruptured, followed by repair. |
| Yao et al. [9] | 2020 | SD rats | 60 | Male | The superficial Achilles tendon was removed, and transection was made in the middle of the deep Achilles tendon, followed by repair. |
| Yao et al. [55] | 2021 | SD rats | 180 | Male | A rectangular full-thick-ness defect was introduced to the left Achilles tendon. |
| Ye et al. [58] | 2023 | SD rats | 30 | Male | Carrageenan solution was injected around the right quadriceps tendon. |
| Yu et al. [68] | 2021 | C57BL/6J mice | 24 | Male | The FDL tendons in the mouse right hind paw were transected and repaired |
| Yu et al. [73] | 2020 | SD rats | 52 | Male | One-third of the central part of the patellar tendon was removed. |
| Zhang et al. [49] | 2020 | SD rats | 54 | Male | One third of the central part of the Achilles tendon was removed. |
| Zhang et al. [57] | 2022 | NZ rabbits | 108 | Male | The supraspinatus tendon was cut from the greater tubercle, and the torn tendon was left unrepaired for 12 weeks. |
| Zhang et al. [75] | 2022 | SD rats | 36 | Male | The supraspinatus tendon was completely cut from the greater tubercle to create a full-thickness injury, which was left unrepaired for 12 weeks. |
| Zhao et al. [78] | 2022 | SD rats | NR | NR | The rat tendons were cut off for one week. |
| Zhu et al. [47] | 2022 | SD rats | 20 | Female | Carrageenan solution was injected around the quadriceps tendon. |
| Author | Year | Animal | Sample Size | Gender | Animal Model |
|---|---|---|---|---|---|
| Han et al. [86] | 2022 | Rabbits | 30 | Male | The supraspinatus tendon was cut, and a 0.5 cm × 0.5 cm tendon tissue was removed. |
| Huang et al. [88] | 2020 | SD rats | 54 | Male | The supraspinatus insertion was resected, and some of the supraspinatus was cut off. |
| Li et al. [89] | 2022 | Wistar rats | 90 | Male | Unilateral ACL resection (at the right side), and bone tunnels were created. |
| Ren et al. [87] | 2021 | SD rats | 36 | NR | The supraspinatus tendon was transected at its insertion site on the greater tuberosity. |
| Shi et al. [84] | 2020 | C57BL/6J mice | 90 | Male | The Achilles tendon was cut off, and the cartilage layer at the insertion was removed. |
| Wang et al. [80] | 2021 | C57BL/6J mice | NR | NR | The Achilles tendon was released from the calcaneal tuberosity to the calf muscle and a 3 mm midline knee incision was made. A tunnel was drilled in the proximal tibial metaphysis to the long axis of the tibia. |
| Wang et al. [90] | 2020 | Rabbits | 35 | Male | The supraspinatus tendon was detached at the insertion on the greater tuberosity of the humerus. |
| Wu et al. [83] | 2022 | SD rats | 108 | Male | Unilateral ACL resection, and bone tunnels were created. |
| Wu et al. [85] | 2021 | C57BL/6J mice | 120 | Male | The supraspinatus tendon was transected at its insertion site on the greater tuberosity, and the remaining tendon and fibrocartilage layer of the footprint were gently abraded. |
| Xu et al. [81] | 2022 | SD rats | 90 | Male | The intra-articular ACL was sectioned, and bone tunnels were created. |
| Zhang et al. [82] | 2022 | SD rats | 78 | Male | Unilateral ACL resection (at the right side), and bone tunnels were created. |
| Author | Year | In Vivo Outcomes | In Vitro Outcomes |
|---|---|---|---|
| Chamberlain et al. [64] | 2019 | EEM treatment significantly increased ultimate stress and Young’s modulus, and EVs treatment was not as effective as EEM treatment in improving biomechanical properties. The treatment of macrophages, EEMs, and EVs all significantly down-regulated type I collagen expression and decreased the M1/M2 macrophage ratio. | NR |
| Chen et al. [61] | 2021 | The biomechanical properties were enhanced with the treatment of rabbit ASC exosomes. Rabbit ASC exosomes reduced the inflammatory hardening, promoted the expression of D CN, COLI, TNMD and BGN, inhibited the expression of type III collagen, and made the collagen fibers more orderly. | HU-MSC exosomes promoted the proliferation and migration of tenocytes in a dose-dependent manner. |
| Cui et al. [56] | 2019 | There were no significant differences in the biomechanical properties of the tendons between the 2 groups. Mouse BMDM exosomes induced fibrosis of injured tendon, upregulated the expression of COL I, COL III, α-SMA and TGF-β1, and caused adhesion between tendon and surrounding tissues. | Mouse BMDM exosomes promoted proliferation and migration of fibroblasts and tenocytes and improved their fibrotic activity, and up-regulated the expression levels of COL I, COL III, α-SMA and TGF-β1 via miR-21-5p which directly targets Smad7. |
| Davies et al. [54] | 2022 | Fat infiltration was significantly reduced in the exosomes treated group. | Mouse FAP exosomes promoted cell proliferation, migration and differentiation. |
| Fu et al. [76] | 2021 | Hydrogel group and EHC group had less inflammatory reaction, while EHC group had more regular fiber arrangement and the best biomechanical properties. EHC significantly up-regulated the expression of RUNX-2, SOX-9 and TNC. Hydrogel also up-regulated their expression, but not as obvious as EHC. | Human ASC exosomes promoted the proliferation and differentiation of TSCs in a dose-dependent manner. |
| Gao et al. [51] | 2022 | Human IP-MSC exosomes down-regulate the expression of CGRP, iNOS and other inflammatory cytokines, and can significantly relieve the pain caused by tendon lesions. The group treated with exosomes showed a higher histological score. | Human IP-MSC exosomes inhibited mast cell activation and down-regulated inflammatory cytokines by regulating HIF-1 signaling pathway. |
| Gissi et al. [62] | 2020 | High concentration of EVs significantly promoted the expression of type I collagen fibers and inhibited the expression of type III collagen fibers, and the group had higher histological scores and a more obvious promotion effect on tendon healing. | High concentration of EVs promoted the expression of type I collagen and the proliferation of tenocytes, and both high and low concentration of EVs promoted the migration of tenocytes. MMP 14 was present in EVs. |
| Han et al. [69] | 2022 | HU-MSC exosomes made Achilles tendon arrange more orderly, cells proliferated well, and effectively alleviated tendon injury. | HU-MSC exosomes promoted the proliferation and migration of tenocytes and activate RhoA in a dose-dependent manner. HU-MSC exosomes reduced tendon injury via miR-27b-3p-mediated suppression of ARHGAP5, resulting in RhoA activation. |
| Hayashi et al. [48] | 2022 | P5 EVs promoted the formation of collagen fibers and the growth of fibrous tissue was good. The P5 EVs treatment group had better histological scores, good tendon healing, and no significant adhesions. | P5 EVs promoted the proliferation and migration of tenocytes more significantly. |
| Jenner et al. [70] | 2023 | Sheep treated with HU-MSC exosomes had improved orientation of collagen fibers and less osteophyte formation at the injury site. The fibrocartilaginous transition zone was formed, inflammation at the lesion site was alleviated and fibrotic adhesions were significantly reduced | HU-MSC exosomes inhibited the proliferation of CD3/CD28 stimulated T-cells. |
| Li et al. [63] | 2020 | Tendon maximum tensile strength remained the same in all three groups. Both EVs and HCPT-EVs reduced the degree of adhesion between tendon and surrounding tissues, while the group treated with HCPT-EVs had the best histological score. | HCPT-EVs up-regulated the expression of GRP78, CHOP and Bax, and down-regulated the expression of Bcl-2, COL III, -SMA, which might activate ERS pathway to inhibit adhesion. HCPT-EVs reduced the activity of fibroblasts and inhibited their proliferation more effectively. |
| Li et al. [71] | 2022 | TSC treated with EUG-BM-MSC-EVs significantly improved tendon fiber arrangement, promoted the expression of type I collagen and type III collagen. The expressions of PCNA, TNMD, bFGF and SCXA were higher than those in other groups. | EUG-BM-MSC-EVs promoted the proliferation and migration of TSC and increased the expression of PCNA. Additionally, EUG-BM-MSC-EVs reversed the down-regulation of Col I, TNC, TNMD and SCXA expression induced by H2O2, and decreased the apoptosis rate, PARP1 expression and ROS content. |
| Liu et al. [77] | 2021 | Rat ASC exosomes significantly increased ultimate load, stiffness, and Young’s modulus, reduced inflammatory reaction, made collagen fibers more organized and tightly packed, and upregulated the expressions of TNMD, COL I, SCXA, and M2 macrophage markers. | Rat ASC exosomes promoted TSCs proliferation and migration and upregulated the expression of TNMD, COL I and SCXA by activating Smad2/3 and Smad1/5/9 signaling pathways. |
| Liu et al. [53] | 2021 | Rat TSC exosomes promoted the expression of type I collagen, inhibited the expression of type III collagen, and inhibited inflammatory reaction. The treatment effect of EXO/MBA group was better than that of EXO group. | EXO/MBA promoted the proliferation and differentiation of tenocytes, promoted the expression of collagen fibers, and effectively inhibited the degradation of extracellular matrix and inflammatory reaction. Exosomes also down-regulated the expression of MMP-3 and MMP-13. |
| Shen et al. [50] | 2019 | Mouse ASC IEVs decreased the activity of NF-κB, up-regulated the expression of COL I, COL II, COL III, SOX-9, and down-regulated the expression of MMP-1. | Both IEVs and EVs effectively inhibited the inflammatory response, but the effect of IEVs was stronger. |
| Shi et al. [52] | 2020 | TEPEP patch significantly improved the load-failure strength and tensile stiffness of the tendon, and increased the expression of collagen fibers. In the group treated with TEPEP patch, there were a large number of fibroblasts migrated to the injury site, and the number of cells was the largest among the three groups. | TEPEP patch up-regulated the expression levels of COL III, MMP-2, MMP-3 and MMP-14 in tenocytes and inhibited inflammatory response. |
| Song et al. [67] | 2022 | The injection of rat TSC exosomes significantly promoted tendon healing and the recovery of biomechanical properties which included the ultimate stress and Young’s modulus of injured tendon. Histologically, the group treated with exosomes had the best histological score with more regular fiber alignment. | Rat TSC exosomes promoted the proliferation and migration of tenocytes and the expression of type I collagen, SCX, COL I and DCN through miR-144-3P. High concentration of exosomes could protect tenocytes from oxidative stress and serum deprivation in vitro. |
| Tao et al. [8] | 2021 | Different EVs have different effects on the biomechanical properties of the interface and the formation of various tissues. H19-OL-EVs demonstrated the best effect on promoting the formation and arrangement of matrix and collagen. | Three kinds of pretreated EVs had more significant effects on the proliferation, migration and differentiation of TSPCs and the activation of YAP. H19 regulated YAP phosphorylation and translocation through H19-pp1-YAP interactions, thereby promoting proliferation, migration, and expression of tendon-related genes. |
| Wang et al. [60] | 2022 | Human ASC exosomes significantly improves ultimate failure load, stiffness, and ultimate tensile strength. Histologically, the group treated with exosomes contained more well-aligned collagen fibers, had a better histological score, and had a higher intensity and amount of type I collagen than the other groups. | NR |
| Wang et al. [65] | 2019 | The injection of exosomes can prevent the decrease of biomechanical properties. Exosomes reduced the fatty infiltration, inflammatory reaction and apoptosis rate of the tissues. The degree of vascularization in exosomes group was lower than that in saline group. | NR |
| Wang et al. [66] | 2019 | Rat TSC exosomes enhanced the maximum loading and ultimate stress of injured tendon. Histologically, the group treated with exosomes had more homogeneous collagen arrangement, better histological score, lower MMP-3 and higher TIMP-3 and COL I expression. | Rat TSC exosomes reversed the inflammatory reaction induced by IL-b, inhibited the expression of MMP-3, and promoted the expression of TIMP-3 and COL I. |
| Wang et al. [79] | 2021 | Human ASC exosomes significantly increased the maximum failure load of the tendon. Human ASC exosomes up-regulated the expression of type I collagen and the ratio of collagen I/III, down-regulated the expression of type III collagen, and inhibited the formation of adhesion and contracture. Human ASC exosomes promoted M2 polarization of macrophages and down-regulated the expression of MMP-3, MMP-13 and SOX-9. | NR |
| Wellings et al. [59] | 2021 | The failure load and ultimate tensile stress of the three groups were similar. PEP-treated tendons contained more well-aligned collagen fibers, lower adhesion grades, and a ratio of type I and type III collagen more similar to normal tendon. | NR |
| Xu et al. [72] | 2022 | Rats treated with exosomes showed less inflammatory reaction, more mature collagen fibers, less angiogenesis, higher expression of type I collagen, lower expression of type III collagen, higher biomechanical properties and lower percentage of lesions than rats in other groups. | MiR-29a, miR-21-5p and miR-148a-3p are highly expressed in Exosomes |
| Xu et al. [74] | 2023 | In the rat Achilles tendon injury model, both EVN and EVB significantly improved the biomechanical properties of the tendon, but EVB restored the intrinsic failure pattern. EVB promoted capillary formation and vessel maturation during tendon regeneration, promoted M2 polarization of macrophages, up-regulated the expression of SCX and TNMD, and reduced scar formation and detrimental morphological changes in the Achilles tendon. | EVN-educated macrophages promoted endothelial cell migration and angiogenesis in vitro. EVB-educated macrophages upregulated VEGF expression and promoted M2 polarization of macrophages. Levels of HSA-miR-125a-5p, HSA-miR-199b-3p, and miR-92b-5p were elevated in EVB, the first two of which were reported to regulate macrophage polarization and promote angiogenesis. |
| Yao et al. [9] | 2020 | There was no significant difference in maximum tensile strength among the three groups. The group treated with HU-MSC exosomes had the best histological score, the least collagen deposition, and the adhesion of tendon and surrounding tissue was effectively relieved. | HU-MSC exosomes manipulated p65 activity by delivering low-abundance miR-21a-3p, thus the expression of COL III and α-SMA was down-regulated, and the proliferation of fibroblasts was inhibited. Ultimately, tendon adhesion was inhibited. |
| Yao et al. [55] | 2021 | HU-MSC exosomes up-regulated the expression of tendon markers such as COL I, TNMD and SCXA, down-regulated the expression of COL III, and promoted the deposition of extracellular matrix in tendon, thus promoting tendon healing. Additionally, HU-MSC exosomes significantly improve the biomechanical properties of tendons through miR-29a-3p. | HU-MSC exosomes regulated PTEN/mTOR/TGF-β1 pathway through miR-29a-3p to up-regulate the expression of tendon-related genes and promote TSC differentiation into tendon. |
| Ye et al. [58] | 2023 | Large EVs and Small EVs had similar functions, including relieving the pain of rat tendon, inhibiting the inflammatory reaction and improving the histological score of diseased tendons. | Large EVs promoted M2 polarization of macrophages in a dose-dependent manner and inhibited inflammatory responses. Large EVs delivered encapsulated DUSP2 and DUSP3 to macrophages, inhibited the activation of P38 MAPK signaling pathway, and thus promoted M2 polarization of macrophages. |
| Yu et al. [68] | 2021 | Ultimate stress, Young’s modulus, and tensile strength were increased with the treatment of mouse BMDM exosomes, while this therapeutic effect was inhibited by the downregulation of CircRNA-EP400. Mouse BMDM exosomes promoted peritendinous fibrosis by CircRNA-EP400. | Mouse BMDM exosomes, especially mouse BMDM exosomes with high CircRNA-EP400 expression, promoted the proliferation and migration of fibroblasts and tenocytes through miR-15b-5p/FGF-1/7/9 pathway, and increased the expression levels of FGF-1, FGF-7, FGF-9, TGF-β1, TGF-I and α-SMA. |
| Yu et al. [73] | 2020 | Rat BM-MSC exosomes increased the deposition of type I collagen and the density and arrangement of cells in the injured area were more similar to those in normal tendon. The histological score was better, and the expression of TNMD, number of TSPCs and biomechanical properties were increased. | Rat BM-MSC Exosomes promoted the proliferation, migration and differentiation of TSPCs and the expression of TNMD, MKX and COL I genes. |
| Zhang et al. [49] | 2020 | Rat TSC exosomes promoted the formation of collagen fibrils, and their arrangement was more continuous and regular. Additionally, exosomes promoted M2 polarization of macrophages and inhibited inflammatory response. | Rat TSC exosomes significantly promoted tenocytes proliferation and migration in a dose-dependent manner through the activation of PI3K/AKT and MAPK/ERK1/2 signaling molecules. Rat TSC exosomes up-regulated the expression of COL I, COL III and TIMP-1, down-regulated the expression of MMP-9 and α-SMA. |
| Zhang et al. [57] | 2022 | The REPA group contained more well-aligned fibers and fibrocartilage, and always showed a lower fat infiltration than other groups | Rabbit BM-MSC exosomes with patch promoted the proliferation and migration of tenocytes. |
| Zhang et al. [75] | 2022 | Injection of exosomes and GC counteracted the negative effect of GC on the biomechanical properties of injured tendons. The rats injected with GC showed fatty infiltration and collagen degeneration, and their histological characteristics were significantly worse than those of rats in the control group and rats injected with GC and exosomes. | GC reduced the inflammatory reaction, but inhibited the proliferation and migration of tenocytes, down-regulated the expression of collagen, up-regulated the expression of MMP-2, MMP-9 and MMP-13, and promoted cell senescence and apoptosis. Exosomes further reduced the inflammatory reaction and counteracted the negative effects of GC. In addition, exosomes promoted cell proliferation, up-regulated collagen expression and type I/III ratio |
| Zhao et al. [78] | 2022 | The group treated with exosomes had less inflammatory reaction, better structure, lower caspase-3 expression and higher PCNA expression and activity of tendon cells. Exosomes decreased the levels of CK, LDH, MDA and oxidative stress. | IGFBP3 promoted the expression of CK, LDH, MDA and caspase-3, and inhibited the expression of PCNA. Exosomes inhibited IGFBP3 expression through MiR-19a, which promoted tenocytes proliferation and decreased apoptosis rate. |
| Zhu et al. [47] | 2022 | Human IP-MSC exosomes can significantly relieve chronic pain caused by tendinopathy. The group treated with exosomes has higher histological scores, and the inflammatory response and capillary proliferation were inhibited. | Human IP-MSC exosomes promoted the proliferation of tenocytes and the expression of anti-inflammatory cytokines, and down-regulated the expression of pro-inflammatory cytokines |
| Author | Year | In Vivo Outcomes | In Vitro Outcomes |
|---|---|---|---|
| Han et al. [86] | 2022 | BMD, TMD, BV/TV were the highest in the BMP-2-EXO group where ultimate load strength and stiffness were also significantly increased. BMP-2-Exosomes inhibited the inflammatory reaction and promoted the formation of fibrocartilage. Chondrocytes arranged in order and the interface was similar to the natural tendon–bone interface. BMP-2-Exosomes up-regulated the expression of Smad4, Smad5, RUNX2, Aggrecan, COL II, SOX-9 and TIMP-1. | NR |
| Huang et al. [88] | 2020 | Rat BM-MSC exosomes up-regulated the expression of COL I, COL II and proteoglycan, and significantly increased the maximum breaking load and stiffness of the tendon. The group treated with exosomes had more neovascularization and better growth of tendon–bone interface. | Rat BM-MSC exosomes activated the Hippo signaling pathway through VGEF, which promoted the proliferation and migration of HUVECs. Exosomes inhibited the M1 polarization of macrophages and inhibited the release of pro-inflammatory cytokines. |
| Li et al. [89] | 2022 | The group treated by exosomes secreted by miR-23a-3p-overexpressing BM-MSCs had the smallest mean bone tunnel area, the largest BV/TV, the decreased relative width of the interface, and the largest amount of fibrocartilage. These exosomes promoted the proliferation of chondrocytes and the expression of COL II, and the maximal failure load and stiffness of the tendon were significantly increased. | Rat BM-MSC Exosomes inhibited inflammation by promoting M2 polarization of macrophages via miR-23a-3p. |
| Ren et al. [87] | 2021 | PEP promoted fibrocartilage and angiogenesis, increased stiffness and maximum tensile load, and inhibited inflammation. The interface was similar to the natural tendon–bone interface. PEP upregulated the expression of COL I, COL III, SCX, TNMD, TNC, DCN and IGF. | PEP significantly promoted the proliferation and migration of tenocytes and osteoblasts, and the fusion time was the shortest. PEP upregulated the expression of COL I, COL III, TNC, DCN, SCX, Spp1, EGR and PPARG. |
| Shi et al. [84] | 2020 | Rat BM-MSC exosomes promoted the formation of transitional structures at the tendon–bone interface and enhanced the biomechanical properties of the tendon–bone interface. The chondrocytes increased and the collagen fibers arranged in order. Exosomes up-regulated the expression of COL II, aggrecan, TGF-β 3, IGF-1 and IGF-2. | Rat BM-MSC exosomes promoted M2 polarization of macrophages and inhibited inflammatory response. |
| Wang et al. [80] | 2021 | Both miR-6924-5p and exosomes secreted by SCX-overexpressing BM-MSCs significantly inhibited osteolysis, prevented osteolysis and improved the biomechanical strength of tendon–bone interface. | Exosomes secreted by SCX-overexpressing BM-MSCs targeted OCSTAMP and CXCL12 via miR-6924-5p, inhibiting osteoclastogenesis. |
| Wang et al. [90] | 2020 | Human ASC exosomes significantly increased ultimate failure load, stiffness and stress. Compared with the group treated with saline, the group treated with exosomes had less fatty infiltration, milder inflammatory reaction, more fibrocartilage and type I collagen fibers, and the tendon–bone interface was more continuous and uniform. | NR |
| Wu et al. [83] | 2022 | The groups treated with exosomes demonstrated smaller mean bone tunnel area, larger BV/TV, and better graft to bone fusion. Exosomes reduced the width of the interface, promoted the formation of fibrocartilage, up-regulated the expression of α-SMA and OCN, and enhanced the biomechanical strength of the tendon–bone interface. | IONP-exosomes promoted NIH3T3 fibroblasts proliferation and migration by down-regulating Smad7 via miR-21-5p, which up-regulated the expression of Col I, Col III and α-SMA. |
| Wu et al. [85] | 2021 | The ultimate failure load and stiffness of the LIPUS-BM-MSC-Exosomes group were significantly higher than other two groups. LIPUS-BM-MSC exosomes significantly inhibited fat infiltration, promoted cell proliferation and formation of fibrocartilage and proteoglycan, further promoted extracellular matrix deposition and repaired the tendon–bone interface. | LIPUS-BM-MSC exosomes up-regulated the expression of chondrogenic genes such as COL II, SOX-9, and aggregate, and down-regulated the expression of adipogenic genes such as Adipo, Retn, and Pparg by miR-140. |
| Xu et al. [81] | 2022 | In the IMEI group, the width of the interface was smaller, the inflammatory reaction was lighter, the fibrocartilage formation was obvious, and the collagen fibers were more orderly arranged. The IMEI group had the smallest mean bone tunnel area and the highest BV/TV. Exosomes significantly increased the ultimate failure load and stiffness, and promoted the M2 polarization of macrophages. | NR |
| Zhang et al. [82] | 2022 | The Hypo-Exo group had the smallest mean bone tunnel area, the largest BV/TV, more fibrocartilage formation, and the highest histological score. Exosomes increased the ultimate failure load and stiffness of tendon, and promoted the formation of CD31+/Emcn+ blood vessels at the tendon–bone interface. Hypo-exosomes were the most effective. | Rat BM-MSC exosomes can promote the proliferation, migration and tube formation of HUVECs, and Hypo-exosomes were the most effective. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, M.; Wang, J.; Shao, Z. Therapeutic Potential of Exosomes in Tendon and Tendon–Bone Healing: A Systematic Review of Preclinical Studies. J. Funct. Biomater. 2023, 14, 299. https://doi.org/10.3390/jfb14060299
Zou M, Wang J, Shao Z. Therapeutic Potential of Exosomes in Tendon and Tendon–Bone Healing: A Systematic Review of Preclinical Studies. Journal of Functional Biomaterials. 2023; 14(6):299. https://doi.org/10.3390/jfb14060299
Chicago/Turabian StyleZou, Mingrui, Jingzhou Wang, and Zhenxing Shao. 2023. "Therapeutic Potential of Exosomes in Tendon and Tendon–Bone Healing: A Systematic Review of Preclinical Studies" Journal of Functional Biomaterials 14, no. 6: 299. https://doi.org/10.3390/jfb14060299
APA StyleZou, M., Wang, J., & Shao, Z. (2023). Therapeutic Potential of Exosomes in Tendon and Tendon–Bone Healing: A Systematic Review of Preclinical Studies. Journal of Functional Biomaterials, 14(6), 299. https://doi.org/10.3390/jfb14060299