
Plant Natural Products: Promising Resources for Cancer Chemoprevention

Abstract
1. Introduction
2. Methods
3. An Overview of Carcinogenesis
4. Scientific Principles of Cancer Chemoprevention
5. Agents for Cancer Chemoprevention
5.1. Synthetic Drugs
5.2. Natural Products
5.2.1. Polyphenols
5.2.2. Flavones
5.2.3. Monoterpene and Triterpenoids
5.2.4. Sulfur Compounds
5.2.5. Cellulose
6. Possible Mechanisms of Inhibition of Carcinogenesis
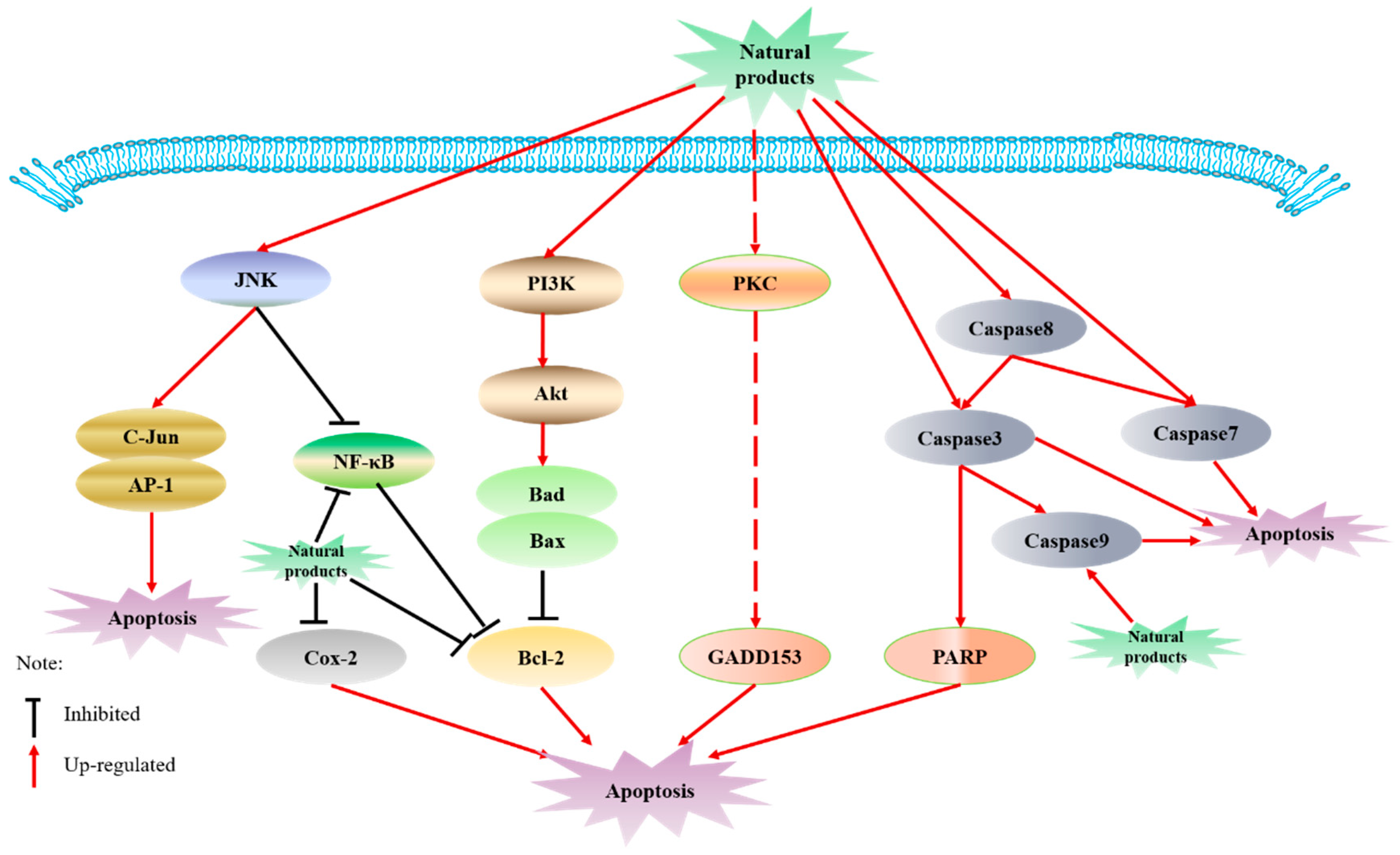
6.1. Induction of Apoptosis
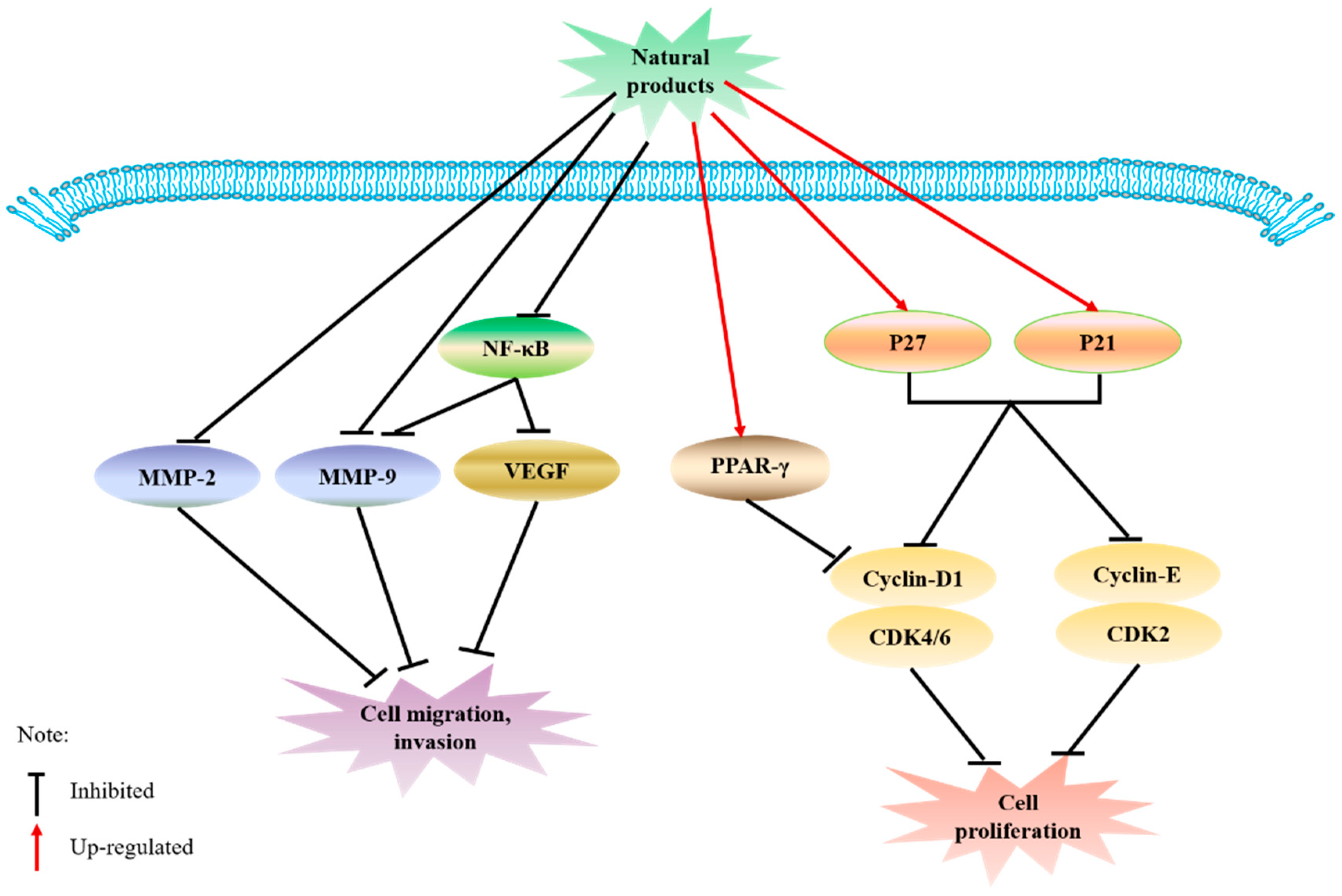
6.2. Inhibition of Cell Signaling Associated with Migration and Proliferation
7. Conclusions and Future Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global Cancer Statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef] [PubMed]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef]
- Karikas, G.A. Anticancer and chemopreventing natural products: Some biochemical and therapeutic aspects. J. BUON 2010, 15, 627–638. [Google Scholar]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural products as sources of new drugs over the period 1981–2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef] [PubMed]
- Sharma, J.; Goyal, P.K. Chemoprevention of chemical-induced skin cancer by Panax ginseng root extract. J. Ginseng Res. 2015, 39, 265–273. [Google Scholar] [CrossRef]
- Vincent, T.L.; Gatenby, R.A. An evolutionary model for initiation, promotion, and progression in carcinogenesis. Int. J. Oncol. 2008, 32, 729–737. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Wu, X.; Patterson, S.; Hawk, E. Chemoprevention—History and general principles. Best Pract. Res. Clin. Gastroenterol. 2011, 25, 445–459. [Google Scholar] [CrossRef] [PubMed]
- Wattenberg, L.W. Chemoprophylaxis of carcinogenesis: A review. Cancer Res. 1966, 26, 1520–1526. [Google Scholar]
- Sporn, M.B.; Dunlop, N.M.; Newton, D.L.; Smith, J.M. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs (retinoids). Fed. Proc. 1976, 35, 1332–1338. [Google Scholar]
- Sporn, M.B. Approaches to prevention of epithelial cancer during the preneoplastic period. Cancer Res. 1976, 36, 2699–2702. [Google Scholar]
- Kelloff, G.J.; Hawk, E.T.; Karp, J.E.; Crowell, J.A.; Boone, C.W.; Steele, V.E.; Lubet, R.A.; Sigman, C.C. Progress in clinical chemoprevention. Semin. Oncol. 1997, 24, 241–252. [Google Scholar] [PubMed]
- Meyskens, F.L.; McLaren, C.E.; Pelot, D.; Brooks, S.F.; Carpenter, P.M.; Hawk, E.; Kelloff, G.; Lawson, M.J.; Kidao, J.; McCracken, J.; et al. Difluoromethylornithine Plus Sulindac for the Prevention of Sporadic Colorectal Adenomas: A Randomized Placebo Controlled. Double-Blind Trial. Cancer Prev. Res. 2008, 1, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Shukla, Y.; Pal, S.K. Dietary cancer chemoprevention: An overview. Int. J. Hum. Genet. 2004, 4, 265–276. [Google Scholar] [CrossRef]
- LaCroix, A.Z.; Powles, T.; Osborne, C.K.; Wolter, K.; Thompson, J.R.; Thompson, D.D.; Allred, D.C.; Armstrong, R.; Cummings, S.R.; Eastell, R.; et al. Breast Cancer Incidence in the Randomized PEARL Trial of Lasofoxifene in Postmenopausal Osteoporotic Women. J. Natl. Cancer Inst. 2010, 102, 1706–1715. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.R.; Bhattoa, H.P.; Neven, P.; Cox, D.A.; Dowsett, S.A.; Alam, J.; Sipos, A.; Muram, D. Gynecologic effects of arzoxifene in postmenopausal women with osteoporosis or low bone mass. Menopause 2012, 19, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Ensrud, K.; Genazzani, A.R.; Geiger, M.J.; McNabb, M.; Dowsett, S.A.; Cox, D.A.; Barrett-Connor, E. Effect of raloxifene on cardiovascular adverse events in postmenopausal women with osteoporosis. Am. J. Cardiol. 2006, 97, 520–527. [Google Scholar] [CrossRef]
- Elgendy, M.; Cirò, M.; Hosseini, A.; Weiszmann, J.; Mazzarella, L.; Ferrari, E.; Cazzoli, R.; Curigliano, G.; DeCensi, A.; Bonanni, B.; et al. Combination of Hypoglycemia and Metformin Impairs Tumor Metabolic Plasticity and Growth by Modulating the PP2A-GSK3b-MCL-1 Axis. Cancer Cell 2019, 35, 1–18. [Google Scholar] [CrossRef]
- Liu, Q.L.; Tong, D.L.; Liu, G.L.; Gao, J.; Wang, L.A.; Xu, J.; Yang, X.X.; Xie, Q.B.; Huang, Y.Q.; Pang, J.; et al. Metformin Inhibits Prostate Cancer Progression by Targeting Tumor-Associated Inflammatory Infiltration. Clin. Cancer Res. 2018, 24, 5622–5634. [Google Scholar] [CrossRef]
- Sporn, M.B.; Liby, K.T. A Mini-Review of Chemoprevention of Cancer—Past, Present, and Future. Prog. Chem. 2013, 25, 1421–1428. [Google Scholar] [CrossRef]
- Shankar, S.; Srivastava, R.K. Curcumin: Structure, biology and clinical applications. In Nutrition, Diet and Cancer; Shankar, S., Srivastava, R.K., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 413–457. [Google Scholar]
- Isemura, M.; Saeki, K.; Kimura, T.; Hayakawa, S.; Minami, T.; Sazuka, M. Tea catechins and related polyphenols as anti-cancer agents. Biofactors 2000, 13, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Shukla, Y. Tea and cancer chemoprevention: A comprehensive review. Asian Pac. J. Cancer Prev. 2007, 8, 155–166. [Google Scholar] [PubMed]
- Shirakami, Y.; Shimizu, M.; Adachi, S.; Sakai, H.; Nakagawa, T.; Yasuda, Y. Epigallocatechin gallate suppresses the growth of human hepatocellular carcinoma cells by inhibiting activation of the vascular endothelial growth factor-vascular endothelial growth factor receptor axis. Cancer Sci. 2009, 100, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G. Anti-Cancer Activities of Tea Epigallocatechin-3-Gallate in Breast Cancer Patients under Radiotherapy. Curr. Mol. Med. 2012, 12, 163–176. [Google Scholar] [CrossRef]
- Rashidi, B.; Malekzadeh, M.; Goodarzi, M.; Masoudifar, A.; Mirzaei, H. Green tea and its anti-angiogenesis effects. Biomed. Pharmacother. 2017, 89, 949–956. [Google Scholar] [CrossRef]
- Bigelow, R.L.H.; Cardelli, J.A. The green tea catechins, (−)-Epigallocatechin-3-gallate (EGCG) and (−)-Epicatechin-3-gallate (ECG), inhibit HGF/Met signaling in immortalized and tumorigenic breast epithelial cells. Oncogene 2006, 25, 1922–1930. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.J.; Wu, C.F.; Ali, Z.; Wang, Y.H.; Khan, S.; Walker, L.A.; Khan, I.A.; Efferth, T. Both Phenolic and Non-phenolic Green Tea Fractions Inhibit Migration of Cancer Cells. Front. Pharmacol. 2016, 7, 398. [Google Scholar] [CrossRef] [PubMed]
- Bettuzzi, S.; Brausi, M.; Rizzi, F.; Castagnetti, G.; Peracchia, G.; Corti, A. Chemoprevention of Human Prostate Cancer by Oral Administration of Green Tea Catechins in Volunteers with High-Grade Prostate Intraepithelial Neoplasia: A Preliminary Report from a One-Year Proof-of-Principle Study. Cancer. Res. 2006, 66, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Kumar1, N.B.; Pow-Sang, J.; Spiess, P.E.; Park, J.; Salup, R.; Williams, C.R.; Parnes, H.; Schell, M.J. Randomized, placebo-controlled trial evaluating the safety of one-year administration of green tea catechins. Oncotarget 2016, 7, 70794–70802. [Google Scholar] [CrossRef]
- Ogas, T.; Kondratyuk, T.P.; Pezzuto, J.M. Resveratrol analogs: Promising chemopreventive agents. Ann. N. Y. Acad. Sci. 2013, 1290, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Leipert, J.; Kässner, F.; Schuster, S.; Händel, N.; Körner, A.; Kiess, W.; Garten, A. Resveratrol potentiates growth inhibitory effects of rapamycin in PTEN-deficient lipoma cells by suppressing p70S6 kinase activity. Nutr. Cancer 2016, 68, 342–349. [Google Scholar] [CrossRef][Green Version]
- Monteillier, A.; Voisin, A.; Furrer, P.; Allémann, E.; Cuendet, M. Intranasal administration of resveratrol successfully prevents lung cancer in A/J mice. Sci. Rep. 2018, 8, 14257. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.H.; Chen, Y.; Huang, J.; Deng, H.; Tang, X.W.; Wang, X.J. Mkp-1 is required for chemopreventive activity of butylated hydroxyanisole and resveratrol against colitis-associated colon tumorigenesis. Food Chem. Toxicol. 2019, 127, 72–80. [Google Scholar] [CrossRef] [PubMed]
- Kiskova, T.; Ekmekcioglu, C.; Garajova, M.; Orenda, S.P.; Bojkova, B.; Bobrov, N.; Jager, W.; Kassayova, M.; Thalhammer, T. A combination of resveratrol and melatonin exerts chemopreventive effects in N-methyl-N-nitrosourea-induced rat mammary carcinogenesis. Eur. J. Cancer Prev. 2012, 21, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Brown, V.A.; Jones, D.J.L.; Britton, R.G.; Hemingway, D.; Miller, A.S.; West, K.P.; Booth, T.D.; Perloff, M.; Crowell, J.A.; et al. Clinical Pharmacology of Resveratrol and its Metabolites in Colorectal Cancer Patients. Cancer Res. 2010, 70, 7392–7399. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Ma, X.Y.; Song, Y.Q.; Zhang, Y.Y.; Xiong, W.B.; Li, L.; Zhou, L.M. Anti-colorectal cancer targets of resveratrol and biological molecular mechanism: Analyses of network pharmacology, human and experimental data. J. Cell Biochem. 2019, 120, 11265–11273. [Google Scholar] [CrossRef]
- Wang, D.; Hang, T.; Wu, C.; Liu, W. Identification of the major metabolites of resveratrol in rat urine by HPLC-MS/MS. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2005, 829, 97–106. [Google Scholar] [CrossRef]
- Aires, V.; Limagne, E.; Cotte, A.K.; Latruffe, N.; Francois, G.; Delmas, D. Resveratrol metabolites inhibit human metastatic colon cancer cells progression and synergize with chemotherapeutic drugs to induce cell death. Mol. Nutr. Food Res. 2013, 57, 1–12. [Google Scholar] [CrossRef]
- Storniolo, C.; Moreno, J.J. Resveratrol metabolites have an antiproliferative effect on intestinal epithelial cancer cells. Food Chem. 2012, 134, 1385–1391. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Rane, G.; Kanchi, M.M.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Tan, B.K.H.; Kumar, A.P.; Sethi, G. The Multifaceted Role of Curcumin in Cancer Prevention and Treatment. Molecules 2015, 20, 2728–2769. [Google Scholar] [CrossRef]
- Park, W.; Amin, A.R.M.R.; Chen, Z.G.; Shin, D.M. New Perspectives of Curcumin in Cancer Prevention. Cancer Prev. Res. 2013, 6, 387–400. [Google Scholar] [CrossRef]
- Teng, C.F.; Yu, C.H.; Chang, H.Y.; Hsieh, W.C.; Wu, T.H.; Lin, J.H.; Wu, H.C.; Jeng, L.B.; Su, I.J. Chemopreventive effect of phytosomal curcumin on hepatitis B virus-related hepatocellular carcinoma in a transgenic mouse model. Sci. Rep. 2019, 9, 10338. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Tastan, H.; Ozercan, I.H.; Guler, O.; Kahraman, N.; Kucuk, O.; Ozpolat, B. Chemopreventive and Antitumor Efficacy of Curcumin in a Spontaneously Developing Hen Ovarian Cancer Model. Cancer Prev. Res. 2017, 11, 59–67. [Google Scholar] [CrossRef]
- Puliyappadamba, V.T.; Thulasidasan, A.K.T.; Vijayakurup, V.; Antony, J.; Bava, S.V.; Anwar, S.; Sundaram, S.; Anto, R.J. Curcumin inhibits B[a]PDE-induced procarcinogenic signals in lung cancer cells, and curbs B[a]P-induced mutagenesis and lung carcinogenesis. Biofactors 2015, 41, 431–442. [Google Scholar] [CrossRef]
- Liu, B.; Cui, L.S.; Zhou, B.; Zhang, L.L.; Liu, Z.H.; Zhang, L. Monocarbonyl curcumin analog A2 potently inhibits angiogenesis by inducing ROS-dependent endothelial cell death. Acta Pharmacol. Sin. 2019, 1412–1423. [Google Scholar] [CrossRef]
- Jiao, D.; Wang, J.; Lu, W.; Tang, X.; Chen, J.; Mou, H.; Chen, Q.Y. Curcumin inhibited HGF-induced EMT and angiogenesis through regulating c-Met dependent PI3K/Akt/mTOR signaling pathways in lung cancer. Mol. Ther. Oncolytics 2016, 3, 16018. [Google Scholar] [CrossRef]
- Vageli, D.P.; Doukas, S.G.; Spock, T.; Sasaki, C.T. Curcumin prevents the bile reflux-induced NF-jB-related mRNA oncogenic phenotype, in human hypopharyngeal cells. J. Cell. Mol. Med. 2018, 22, 4209–4220. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Yu, X.J.; Zhang, L.; Wang, L.; Peng, Z.H.; Chen, Y. The pharmacokinetics and tissue distribution of curcumin and its metabolites in mice. Biomed. Chromatogr. 2018, 32, e4267. [Google Scholar] [CrossRef]
- Boven, L.; Holmes, S.P.; Latimer, B.; McMartin, K.; Ma, X.H.; Moore-Medlin, T.; Khandelwal, A.R.; McLarty, J.; Nathan, C.A.O. Curcumin Gum Formulation for Prevention of Oral Cavity Head and Neck Squamous Cell Carcinoma. Laryngoscope 2019, 129, 1597–1603. [Google Scholar] [CrossRef]
- Gibellini, L.; Pinti, M.; Nasi, M.; Montagna, J.P.; Biasi, S.D.; Roat, E.; Bertoncelli, L.; Cooper, E.L.; Cossarizza, A.A. Quercetin and Cancer Chemoprevention. Evid. Based Complement. Alternat. Med. 2011, 2011, 591356. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, G.; Dai, J.G.; Sun, Y.; Hoffman, R.M.; Yang, Z.J.; Fan, Y. Chemoprevention by Quercetin of Oral Squamous Cell Carcinoma by Suppression of the NF-ĸB Signaling Pathway in DMBA-treated Hamsters. Anticancer Res. 2017, 37, 4041–4050. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, G.; Athirai, T.; Kiruthiga, B.; Senthilkumar, K.; Elumalai, P.; Arunkumar, R.; Arunakaran, J. Chemopreventive Effect of Quercetin in MNU and Testosterone Induced Prostate Cancer of Sprague-Dawley Rats. Nutr. Cancer 2014, 66, 38–46. [Google Scholar] [CrossRef]
- Patel, D.H.; Sharma, N. Inhibitory effect of quercetin on epithelial to mesenchymal transition in SK-MEL -28 human melanoma cells defined by in vitro analysis on 3D collagen gels. OncoTargets Ther. 2016, 9, 6445–6459. [Google Scholar] [CrossRef]
- Cassia, C.C.; Corrêa, M.P.; Carvalho, T.T.; Zarpelon, A.C.; Hohmann, M.S.; Rossaneis, A.C.; Coelho-Silva, L.; Pavanelli, W.R.; Pinge-Filho, P.; Crespigio, J. Quercetin reduces Ehrlich tumor-induced cancer pain in mice. Anal. Cell. Pathol. 2015, 2015, 285708. [Google Scholar] [CrossRef]
- Wang, P.; Vadgama, J.V.; Said, J.W.; Magyar, C.E.; Doan, N.; Heber, D.; Henning, S.M. Enhanced inhibition of prostate cancer xenograft tumor growth by combining quercetin and green tea. J. Nutr. Biochem. 2014, 25, 73–80. [Google Scholar] [CrossRef]
- Wang, P.; Heber, D.; Henning, S.M. Quercetin increased bioavailability and decreased methylation of green tea polyphenols in vitro and in vivo. Food Funct. 2012, 3, 635–642. [Google Scholar] [CrossRef]
- Rich, G.T.; Buchweitz, M.; Winterbone, M.S.; Kroon, P.A.; Wilde, P.J. Towards an understanding of the low bioavailability of quercetin: A study of its interaction with intestinal lipids. Nutrients 2017, 9, 111. [Google Scholar] [CrossRef]
- Ader, P.; Wessmann, A.; Wolffram, S. Bioavailability and metabolism of the flavonol quercetin in the pig. Free Radic. Biol. Med. 2000, 28, 1056–1067. [Google Scholar] [CrossRef]
- Xiao, C.; Yin, O.Q.P.; Zhong, Z.; Chow, M.S.S. Pharmacokinetics and Modeling of Quercetin and Metabolites. Pharm. Res. 2005, 22, 892–901. [Google Scholar] [CrossRef]
- Li, H.L.; Zhao, X.B.; Ma, Y.K.; Zhai, G.X.; Li, L.B.; Lou, H.X. Enhancement of gastrointestinal absorption of quercetin by solid lipid nanoparticles. J. Control Release 2009, 133, 238–244. [Google Scholar] [CrossRef]
- Wu, Y.Y. Research Progress in Anabolic Control Mechanisms of Plant Carotenoids. Bot. Res. 2020, 9, 217–225. [Google Scholar] [CrossRef]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer Chemoprevention by Carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Sarkar, A.; Chatterjee, M. Further evidence for chemopreventive potential of beta-carotene against experimental carcinogenesis: Diethylnitrosamine-initiated and phenobarbital-promoted hepatocarcinogenesis is prevented more effectively by beta-carotene than by retinoic acid. Nutr. Cancer 2000, 37, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Obermüller-Jevic, U.C.; Hellmis, E.; Koch, W.; Jacobi, G.; Biesalski, H.K. Lycopene inhibits disease progression in patients with benign prostate hyperplasia. J. Nutr. 2008, 138, 49–53. [Google Scholar] [CrossRef]
- Ip, B.C.; Hu, K.Q.; Liu, C.; Smith, D.E.; Obin, M.S.; Ausman, L.M.; Wang, X.D. Lycopene metabolite, apo-10′-lycopenoic acid, inhibits diethylnitrosamine-initiated, high fat diet-promoted hepatic inflammation and tumorigenesis in mice. Cancer Prev. Res. 2013, 6, 1304–1316. [Google Scholar] [CrossRef]
- Goswami, S.; Srivastava, A.; Pushker, N. Abstract 3812: Induction of apoptosis by zeaxanthin in human uveal melanoma cells. Cancer Res. 2015, 75, 3812. [Google Scholar] [CrossRef]
- Zhang, W.L.; Zhao, Y.N.; Shi, Z.Z.; Cong, D.; Bai, Y.S. Lutein Inhibits Cell Growth and Activates Apoptosis via the PI3K/AKT/mTOR Signaling Pathway in A549 Human Non-Small-Cell Lung Cancer Cells. J. Environ. Pathol. Toxicol. Oncol. 2018, 37, 341–350. [Google Scholar] [CrossRef]
- Yonekura, L.; Nagao, A. Intestinal absorption of dietary carotenoids. Mol. Nutr. Food Res. 2007, 51, 107–115. [Google Scholar] [CrossRef]
- Brown, M.J.; Ferruzzi, M.G.; Nguyen, M.L.; Cooper, D.A.; Eldridge, A.L.; Schwartz, S.J.; White, W.S. Carotenoid bioavailability is higher from salads ingested with full-fat than fat-reduced salad dressings as measured with electrochemical detection. Am. J. Clin. Nutr. 2004, 80, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Elgendy, M.; Ciro, M.; Hosseini, A.; Weiszmann, J.; Mazzarella, L.; Ferrari, E.; Cazzoli, R.; Curigliano, G.; DeCensi, A.; Bonanni, B.; et al. Studies on the chemical constitution of Egyptian, N. sativaL. Seeds. Planta Med. 1963, 11, 465–470. [Google Scholar] [CrossRef]
- Sayed-Ahmed, M.M.; Aleisa, A.M.; Al-Rejaie, S.S.; Al-Yahya, A.A.; Al-Shabanah, O.A.; Hafez, M.M.; Nagi, M.N. Thymoquinone Attenuates Diethylnitrosamine Induction of Hepatic Carcinogenesis Through Antioxidant Signaling. Oxid. Med. Cell. Longev. 2010, 3, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Yi, T.; Cho, S.G.; Yi, Z.; Pang, X.; Rodriguez, M.; Wang, Y.; Sethi, G.; Aggarwal, B.B.; Liu, M. Thymoquinone inhibits tumor angiogenesis and tumor growth through suppressing AKT and ERK signaling pathways. Mol. Cancer Ther. 2008, 7, 1789–1796. [Google Scholar] [CrossRef]
- Aziza, S.; Hussein, S.A.; Khalaf, H.A. Chemopreventive effect of thymoquinone on benzo(a)pyrene-induced lung cancer in male swiss albino mice. Benha Vet. Med. J. 2014, 27, 330–340. [Google Scholar]
- Alkharfy, K.M.; Ahmad, A.; Khan, R.M.A.; Al-Shagha, W.M. Pharmacokinetic plasma behaviors of intravenous and oral bioavailability of thymoquinone in a rabbit model. Eur. J. Drug Metab. Pharmacokinet. 2015, 40, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Vieira, A.; Heidor, R.; Cardozo, M.T.; Scolastici, C.; Purgatto, E.; Shiga, T.M.; Barbisan, L.F.; Ong, T.P.; Moreno, F.S. Efficacy of geraniol but not of β-ionone or their combination for the chemoprevention of rat colon carcinogenesis. Braz. J. Med. Biol. Res. 2011, 44, 538–545. [Google Scholar] [CrossRef]
- Kim, S.H.; Bae, H.C.; Park, E.J.; Lee, C.R.; Kim, B.J.; Lee, S.; Park, H.H.; Kim, S.J.; So, I.; Kim, T.W. Geraniol inhibits prostate cancer growth by targeting cell cycle and apoptosis pathways. Biochem. Biophys. Res. Commun. 2011, 407, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Patlolla, J.M.R.; Rao, C.V. Triterpenoids for Cancer Prevention and Treatment: Current Status and Future Prospects. Curr. Pharm. Biotechnol. 2012, 13, 147–155. [Google Scholar] [CrossRef]
- Mahato, S.B.; Sarkar, S.K.; Poddar, G. Triterpenoid saponins. Phytochemistry 1988, 27, 3037–3067. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Indranie, C.; Malisetty, S.V.; Jagan, P.; Steele, V.E.; Rao, C.V. Chemoprevention of Colon Carcinogenesis by Oleanolic Acid and Its Analog in Male F344 Rats and Modulation of COX-2 and Apoptosis in Human Colon HT-29 Cancer Cells. Pharm. Res. 2008, 25, 2151–2157. [Google Scholar] [CrossRef]
- LI, L.; Lin, J.; Sun, G.; Wei, L.; Shen, A.; Zhang, M.; Peng, J. Oleanolic acid inhibits colorectal cancer angiogenesis in vivo and in vitro via suppression of STAT3 and Hedgehog pathways. Mol. Med. Rep. 2016, 13, 5276–5282. [Google Scholar] [CrossRef]
- Dong, W.J.; Kim, Y.H.; Hui, H.K.; Ji, H.Y.; Yoo, S.D.; Choi, W.R.; Lee, S.M.; Han, C.K.; Lee, H.S. Dose-Linear Pharmacokinetics of Oleanolic Acid after Intravenous and Oral Administration in Rats. Biopharm. Drug Dispos. 2007, 28, 51–57. [Google Scholar] [CrossRef]
- Konopleva, M.; Zhang, W.; Shi, Y.X.; McQueen, T.; Tsao, T.; Abdelrahim, M.; Munsell, M.F.; Johansen, M.; Yu, D.; Madden, T.; et al. Synthetic triterpenoid 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid induces growth arrest in HER2-overexpressing breast cancer cells. Mol. Cancer Ther. 2006, 5, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Tran, K.; Risingsong, R.; Royce, D.B.; Williams, C.R.; Sporn, M.B.; Pioli, P.A.; Gediya, L.K.; Njar, V.C.; Liby, K.T. The combination of the histone deacetylase inhibitor vorinostat and synthetic triterpenoids reduces tumorigenesis in mouse models of cancer. Carcinogenesis 2013, 34, 199–210. [Google Scholar] [CrossRef]
- Kwon, H.J.; Shim, J.S.; Kim, J.H.; Cho, H.Y.; Yum, Y.N.; Kim, S.H.; Yu, J. Betulinic acid inhibits growth factor-induced in vitro angiogenesis via the modulation of mitochondrial function in endothelial cells. Jpn. J. Cancer Res. 2002, 93, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Qin, L.; Xu, Y.; Cheng, N. Inhibition of betulinic acid to growth and angiogenesis of human colorectal cancer cell in nude mice. Chin. Ger. J. Clin. Oncol. 2010, 9, 153–157. [Google Scholar] [CrossRef]
- Ribeiro, C.M.S.; Savelkoul, H.F.J.; Wiegertjes, G.F. Immune responses of carp against parasites. Cheminform 2009, 35, 3169–3172. [Google Scholar] [CrossRef]
- Udeani, G.O.; Zhao, G.M.; Geun, S.Y.; Cooke, B.P.; Graham, J.; Beecher, C.W.; Kinghorn, A.D.; Pezzuto, J.M. Pharmacokinetics and tissue distribution of betulinic acid in CD-1 mice. Biopharm. Drug Dispos. 1999, 20, 379–383. [Google Scholar] [CrossRef]
- Herman-Antosiewicz, A.; Powolny, A.A.; Singh, S.V. Molecular targets of cancer chemoprevention by garlic-derived organosulfides. Acta Pharmacol. Sin. 2007, 28, 1355–1364. [Google Scholar] [CrossRef]
- Wang, W.; Sukamtoh, E.; Xiao, H. Allicin Inhibits Lymphangiogenesis in vitro and in vivo. Mol. Nutr. Food Res. 2015, 59, 2345–2354. [Google Scholar] [CrossRef]
- Xiang, Y.; Zhao, J.; Zhao, M. Allicin activates autophagic cell death to alleviate the malignant development of thyroid cancer. Exp. Ther. Med. 2018, 15, 3537–3543. [Google Scholar] [CrossRef]
- Lai, K.C.; Hsu, S.C.; Kuo, C.L.; Yang, J.S.; Ma, C.Y.; Lu, H.F.; Tang, N.Y.; Hsia, T.C.; Ho, H.C.; Chung, J.G. Diallyl Sulfide, Diallyl Disulfide, and Diallyl Trisulfide Inhibit Migration and Invasion in Human Colon Cancer Colo 205 Cells Through the Inhibition of Matrix Metalloproteinase-2, -7, and -9 Expressions. Environ. Toxicol. 2013, 28, 479–488. [Google Scholar] [CrossRef] [PubMed]
- Thejass, P.; Kuttan, G. Immunomodulatory activity of Sulforaphane, a naturally occurring isothiocyanate from broccoli (Brassica oleracea). Phytomedicine 2007, 14, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Jakubikova, J.; Sedlak, J. Garlic-derived organosulfides induce cytotoxicity, apoptosis, cell cycle arrest and oxidative stress in human colon carcinoma cell lines. Neoplasma 2006, 53, 191–199. [Google Scholar]
- Ong, C.; Elbarbry, F. A new validated HPLC method for the determination of sulforaphane: Application to study pharmacokinetics of sulforaphane in rats. Biomed. Chromatogr. 2016, 30, 1016–1021. [Google Scholar] [CrossRef] [PubMed]
- Burnett, J.P.; Lim, G.; Li, Y.; Shah, R.B.; Lim, R.; Paholak, H.J.; McDermott, S.P.; Sun, L.; Tsume, Y.; Bai, S.; et al. Sulforaphane enhances the anticancer activity of taxanes against triple negative breast cancer by killing cancer stem cells. Cancer Lett. 2017, 394, 52–64. [Google Scholar] [CrossRef]
- Razis, A.F.A.; Noor, N.M. Sulforaphane is superior to glucoraphanin in modulating carcinogen-metabolising enzymes in Hep G2 cells. Asian Pac. J. Cancer Prev. 2013, 14, 4235–4238. [Google Scholar] [CrossRef] [PubMed]
- Cornblatt, B.S.; Ye, L.; Dinkova-Kostova, A.T.; Erb, M.; Fahey, J.W.; Singh, N.K.; Chen, M.S.A.; Stierer, T.; Garrett-Meyer, E.; Argani, P.; et al. Preclinical and Clinical Evaluation of Sulforaphane for Chemoprevention in the Breast. Carcinogenesis 2007, 28, 1485–1490. [Google Scholar] [CrossRef]
- Veeranki, O.L.; Bhattacharya, A.; Tang, L.; Marshall, J.R.; Zhang, Y. Cruciferous Vegetables, Isothiocyanates, and Prevention of Bladder Cancer. Curr. Pharmacol. Rep. 2015, 1, 272–282. [Google Scholar] [CrossRef]
- Davis, C.D. Selenium Supplementation and Cancer Prevention. Curr. Nutr. Rep. 2012, 1, 16–23. [Google Scholar] [CrossRef]
- Kandaş, N.O.; Randolph, C.; Bosland, M.C. Differential effects of selenium on benign and malignant prostate epithelial cells: Stimulation of LNCaP cell growth by noncytotoxic, low selenite concentrations. Nutr. Cancer 2009, 61, 251–264. [Google Scholar] [CrossRef]
- Letavayová, L.; Vlcková, V.; Brozmanová, J. Selenium: From cancer prevention to DNA damage. Toxicology 2006, 227, 1–14. [Google Scholar] [CrossRef]
- Schwarz, E.C.; Qu, B.; Hoth, M. Calcium, cancer and killing: The role of calcium in killing cancer cells by cytotoxic T lymphocytes and natural killer cells. Biochim. Biophys. Acta 2013, 1833, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Sesink, A.L.A.; Termont, D.S.M.L.; Kleibeuker, J.H.; Meer, R.V.D. Red meat and colon cancer: Dietary haem-induced colonic cytotoxicity and epithelial hyperproliferation are inhibited by calcium. Carcinogenesis 2001, 22, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Grothey, A.; Nikcevich, D.A.; Sloan, J.A.; Kugler, J.W.; Silberstein, P.T.; Dentchev, T.; Wender, D.B.; Novotny, P.J.; Chitaley, U.; Alberts, S.R.; et al. Intravenous calcium and magnesium for oxaliplatin-induced sensory neurotoxicity in adjuvant colon cancer: NCCTG N04C7. J. Clin. Oncol. 2011, 29, 421–427. [Google Scholar] [CrossRef]
- Li, J.; Koh, W.P.; Jin, A.Z.; Yuan, J.M.; Yu, M.C.; Butler, L.M. Calcium intake is not related to breast cancer risk among Singapore Chinese women. Int. J. Cancer 2013, 133, 680–686. [Google Scholar] [CrossRef]
- Colston, K.; Colston, M.J.; Feldman, D. 1,25-Dihydroxyvitamin D3 and malignant melanoma: The presence of receptors and inhibition of cell growth in culture. Endocrinology 1981, 108, 1083–1086. [Google Scholar] [CrossRef]
- Krishnan, A.V.; Peehl, D.M.; Feldman, D. Inhibition of Prostate Cancer Growth by Vitamin D: Regulation of Target Gene Expression. J. Cell. Biochem. 2003, 88, 363–371. [Google Scholar] [CrossRef]
- Pommergaard, H.C.; Raskov, H.; Rosenberg, B.J. Chemoprevention with Acetylsalicylic Acid, Vitamin D and Calcium Reduces Risk of Carcinogen-induced Lung Tumors. Anticancer Res. 2013, 33, 4767. [Google Scholar] [CrossRef][Green Version]
- Welsh, J. Cellular and molecular effects of vitamin D on carcinogenesis. Arch. Biochem. Biophys. 2012, 523, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Khan, Q.J.; Kimler, B.F.; Fabian, C.J. The relationship between vitamin D and breast cancer incidence and natural history. Curr. Oncol. Rep. 2010, 12, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.L.; Campbell, S.E.; Krishnan, K. The Role of Vitamin E in Prostate Cancer. In Oxidative Stress in Cancer Biology and Therapy; Basu, S., Wiklund, L., Eds.; Human Press: Totowa, NJ, USA, 2012; pp. 333–354. [Google Scholar]
- Sun, S.Y.; Numsen, H.; Reuben, L. Apoptosis as a Novel Target for Cancer Chemoprevention. J. Natl. Cancer Inst. 2004, 96, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Collett, G.P.; Campbell, F.C. Curcumin induces c-jun N-terminal kinase-dependent apoptosis in HCT116 human colon cancer cells. Carcinogenesis 2004, 25, 2183–2189. [Google Scholar] [CrossRef]
- Chhunchha, B.; Fatma, N.; Kubo, E.; Rai, P.; Singh, S.P.; Singh, D.P. Curcumin abates hypoxia-induced oxidative stress based-ER stress-mediated cell death in mouse hippocampal cells (HT22) by controlling Prdx6 and NF-κB regulation. Am. J. Physiol. Cell Physiol. 2013, 304, C636–C655. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, C.; Song, D.; Xia, R.; Yu, W.; Dang, Y.; Fei, Y.; Yu, L.; Wu, J. Epigallocatechin-3-gallate enhances ER stress-induced cancer cell apoptosis by directly targeting PARP16 activity. Cell Death Discov. 2017, 10, 17034. [Google Scholar] [CrossRef]
- Wu, P.P.; Kuo, S.C.; Huang, W.W.; Yang, J.S.; Lai, K.C.; Chen, H.J.; Lin, K.L.; Chiu, Y.J.; Huang, L.J.; Chung, J.G. (−)-Epigallocatechin gallate induced apoptosis in human adrenal cancer NCI-H295 cells through caspase-dependent and caspase-independent pathway. Anticancer Res. 2009, 29, 1435–1442. [Google Scholar] [PubMed]
- Lin, Z.H. Epicallocatechin Gallate Induces Apoptosis of Pancreatic Cancer Cells Through PI3K-Akt Signaling Pathway. Master’s Thesis, Shanxi Medical University, Taiyuan, China, 2011. (In Chinese). [Google Scholar]
- Sheth, S.; Jajoo, S.; Kaur, T.; Mukherjea, D.; Sheehan, K.; Rybak, L.P.; Ramkumar, V. Resveratrol reduces prostate cancer growth and metastasis by inhibiting the Akt/MicroRNA-21 pathway. PLoS ONE 2012, 7, e51655. [Google Scholar] [CrossRef]
- Frazzi, R.; Valli, R.; Tamagnini, I.; Casali, B.; Latruffe, N.; Merli, F. Resveratrol-mediated apoptosis of hodgkin lymphoma cells involves SIRT1 inhibition and FOXO3a hyperacetylation. Int. J. Cancer 2013, 132, 1013–1021. [Google Scholar] [CrossRef]
- Min, J.R.; Chung, H.S. [10]-Gingerol induces mitochondrial apoptosis through activation of MAPK pathway in HCT116 human colon cancer cells. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 92–101. [Google Scholar] [CrossRef]
- Johnson, J.J.; Mukhtar, H. Curcumin for chemoprevention of colon cancer. Cancer Lett. 2007, 255, 170–181. [Google Scholar] [CrossRef]
- Narayanaswamy, R.; Wai, L.K.; Abas, F. Molecular docking analysis of Curcumin analogues as Matrix metalloproteinases (MMP 2 and MMP 9) inhibitors. J. Appl. Cosmetol. 2017, 35, 61–68. [Google Scholar]
- Yang, J.; Wang, C.; Zhang, Z.; Chen, X.; Jia, Y.; Wang, B.; Kong, T. Curcumin inhibits the survival and metastasis of prostate cancer cells via the Notch-1 signaling pathway. APMIS 2017, 125, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Wang, X. Furanodiene blocks NF-kB-dependent MMP-9 and VEGF activation and inhibits cellular invasiveness and angiogenesis of breast cancer cells in vitro and in vivo. Biomed. Res. 2012, 23, 231–237. [Google Scholar]
- Liao, S.; Xia, J.; Chen, Z.; Zhang, S.; Ahmad, A.; Miele, L.; Sarkar, F.H.; Wang, Z. Inhibitory effect of curcumin on oral carcinoma CAL-27 cells via suppression of Notch-1 and NF-κB signaling pathways. J. Cell. Biochem. 2011, 112, 1055–1065. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Zhang, X.; Gao, D.; Jiang, X.; Dong, W. Resveratrol inhibits MMP-9 expression by up-regulating PPAR alpha expression in an oxygen glucose deprivation-exposed neuron model. Neurosci. Lett. 2009, 451, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Chen, X.; Yu, H. Resveratrol Inhibits MMP-2 Expression of Hepatoma in Nude Mice. J. Anim. Vet. Adv. 2011, 10, 33–37. [Google Scholar] [CrossRef][Green Version]
- Kim-Park, W.K.; Allam, E.S.; Palasuk, J.; Kowolik, M.; Park, K.K.; Windsor, L.J. Green tea catechin inhibits the activity and neutrophil release of Matrix Metalloproteinase-9. J Tradit. Complem. Med. 2016, 6, 343–346. [Google Scholar] [CrossRef]
- Chao, P.; Luo, Y.M.; Chen, Y.Q. Epigallocatechin gallate inhibits activity of matrix metalloproteinase-2 and NF-κB in U87MG glioblastoma multiforme cells. Chin. J. Mod. Med. 2015, 25, 27–31. [Google Scholar]
- Lim, T.G.; Lee, S.Y.; Huang, Z. Curcumin suppresses proliferation of colon cancer cells by targeting CDK2. Cancer Prev. Res. 2014, 7, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Deng, S.W.; Yin, Q.H.; Qi, S.U.; Zhuang, Y.Z. Influence of autophagic apoptosis induced by curcumin on human gastric carcinoma SGC7901 cells. Chin. J. Mod. Med. 2012, 22, 30–35. [Google Scholar]
- Zhai, X.X.; Ding, J.C.; Tang, Z.M. Effects of resveratrol on the proliferation, apoptosis and telomerase ability of human A431 epidermoid carcinoma cells. Oncol. Lett. 2016, 11, 3015–3018. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Cekanova, M.; Baek, S.J. Multiple mechanisms are involved in 6-gingerol-induced cell growth arrest and apoptosis in human colorectal cancer cells. Mol. Carcinogen. 2008, 47, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Min, K.W.; Wimalasena, J.; Baek, S.J. Cyclin D1 degradation and p21 induction contribute to growth inhibition of colorectal cancer cells induced by epigallocatechin-3-gallate. J. Cancer Res. Clin. 2012, 138, 2051–2060. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Common Phytochemicals | |||||||
|---|---|---|---|---|---|---|---|
| Groups | Phytochemicals | Structure | Cancer Type | The Subjects | Chemopreventive Property | Common Source | Reference |
| Phenols | (−)-epicatechin (EC) |  | Prostate cancer | Human | Oral administration of 600 mg/d for 1 year reduced the incidence of diagnosed cancers in volunteers with high-grade intraepithelial neoplasia of the prostate (H GPIN) | Green tea | [30] |
| (−)-epigalocatachin (EGC) |  | Breast cancer | MCF10A cells | Completely inhibited Met, AKT and ERK phosphorylation at 0.6 mM | [27] | ||
| (−)-epicatechin-3-gallate (ECG) |  | Liver cancer | HepG2 cells | Interrupted closures by the disassembly of microtubules | [28] | ||
| Osteosarcoma cancer | U2OS cells | ||||||
| (−)-epigallocatechin-3-gallate (EGCG) |  | Hepatocellular carcinoma | HuH7 cells | Inhibited the growth of HuH7 xenografts | [25] | ||
| Breast cancer | Tumorigenic breast epithelial cells | Blocked the ability of hepatocyte growth factor (HGF) to induce cell motility and invasion | [27] | ||||
| Prostate cancer | Human | Not produce treatment related adverse effects in men with baseline HGPIN or ASAP | [31] | ||||
| Resveratrol |  | Lung cancer | A/J mice | Tumor diversity and volume decreased in mice | Polygonum cuspidatum, red grapes, berries, peanuts, pines etc. | [34] | |
| Colorectal cancer | BALB/c wild-type mice | Marked suppression of dextran sulfate sodium (DSS)-associated tumorigenesis | [35] | ||||
| Human | It can be used as a potential chemoprophylaxis for colorectal cancer tract | [37] | |||||
| Curcumin |  | Liver cancer | Transgenic mice | Inhibition of hepatocellular carcinoma formation, improvement of liver histopathology, and reduction of total tumor volume in transgenic mice | [44] | ||
| Ovarian cancer | Hens | Reduced the overall ovarian cancer incidence to 31% and 57% | [45] | ||||
| Lung cancer | Lung cancer cells (H1299, A549) | An inhibitory effect in lung carcinogenesis induced by B[a]P, a procarcinogen present in environment and cigarette smoke | Curcuma Longa | [46] | |||
| Curcuma Longa A2 |  | ROS-dependent endothelial cell | Human umbilical vein endothelial cells (HUVECs) | Suppresses the migration and tube formation of human umbilical vein endothelial cells (HUVECs) in vitro | [47] | ||
| —————— | Rat | Suppresses newly formed microvessels in chicken chorioallantoic membranes (CAMs) and Matrigel plus in vivo | |||||
| Flavonoids | Quercetin |  | Oral squamous cell carcinoma (OSCC) | Hamster | Decreased incidence of oral squamous cell carcinoma (OSCC) and severity of hyperplasia and dysplasia | Apples, onions, tomatoes, broccoli, citrus fruit, etc. | [34] |
| Melanoma | SK-MEL-28 human melanoma cells | Decreased migration rates (26.36% vs. 64.36%) and motility rate by approximately tenfold in SK-MEL-28 cells cultured on collagen I matrices | [55] | ||||
| Lutein |  | Lung and colon cancers | Human | Reduce the risk of lung and colon cancers by the suppression of k-Ras and β-catenin expression | Papaya, pumpkin, citrus, wolfberry, peach, spinach, leek, corn, Chinese cabbage, etc. | [68] | |
| Zeaxanthin |  | ||||||
| Lycopene |  | Benign prostate hyperplasia (BPH) | Human | Inhibited serum prostate-specific antigen (PSA) increase, and further improving clinical diagnostic markers and symptoms of BPH | Tomato, tomato products, watermelon, grapefruit etc. | [66] | |
| Apo-10-lycopenoic |  | Liver tumors | HFD-fed mice | APO10LA can effectively inhibit HFD-promoted hepatic tumorigenesis by stimulating SIRT1 signaling while reducing hepatic inflammation | [67] | ||
| Monoterpene | Thymoquinone |  | Human prostate cancer | PC3 cancer cells | Inhibits tumor angiogenesis and tumor growth and could be used as a potential drug candidate for cancer therapy | The seed oil of Nigella sativa L. | [74] |
| —————— | Human umbilical vein endothelial cell (HUVEC) | Effectively inhibited migration, invasion, and tube formation of human umbilical vein endothelial cell (HUVEC) | [74] | ||||
| Human prostate cancer (PC3) | Male mice | Inhibited human prostate tumor growth in both size and weight in a xenograft human prostate cancer (PC3) model in mice | [74] | ||||
| Triterpenoids | Oleanolic acid |  | Colon cancer | Male mice | Oleanolic acid inhibited, in a dose-dependent manner, the average azomethane (AOM)-induced abnormal colonic cavitation lesions in male F344 rats (36–52%) | Hedyotis Herbaherba, hawthorn, Syzygium Aromaticum, loquat leaf etc. | [81] |
| Colorectal cancer | Mice | Inhibitory tumor growth of xenograft tumor tissue in mice with colorectal cancer | [82] | ||||
| 2-cyano-3,12-dioxooleana-1,9(11)-dien-28-oic acid |  | Breast cancer | Immunodeficient mice | CDDO (20 mg/kg, i.v.) treatment for 3 weeks abrogated the growth of both MCF7/HER2 and MDA-MB-435/HER2 tumors types in immunodeficient mice, by inhibiting HER2 phosphorylation and decreasing HER2 kinase activity | [84] | ||
| Betulinic acid |  | Colorectal cancer | Nude mice | Possess antiangiogenic effects by inhibiting aminopeptidase N | White birch bark, Ziziphi Spinosae, semen | [87] | |
| Sulfur compounds | Allicin |  | —————— | —————— | Inhibited lymphangiogenesis suppressing activation of vascular endothelial growth factor (VEGF) receptor | Garlic, allium, vegetables etc. | [91] |
| Thyroid cancer | SW1736 and HTh-7 cells | Served as an adjunctive therapy for thyroid cancer, as it induces autophagic cell death to alleviate the malignant development of cancer | [92] | ||||
| Diallyl sulfide |  | —————— | C57BL/6 mice | Compared to the control group, through enhancing the production of antiangiogenic factors such as IL-2 and TIMP | [94] | ||
| Diallyl tetrasulfide |  | Colon cancer | CoLo 205 cells | Three oil-soluble compounds including DAS, DADS, and DAT at 10 and 25 μM have an inhibitory effect on the migration and invasion of human colon cancer cells with the order of DATS < DADS < DAS | [93] | ||
| Diallyl disulfide |  | ||||||
| Diallyl trisulfide |  | ||||||
| Sulforaphane |  | Breast cancer | Rats | As a conceptually promising agent in breast cancer prevention. It can be rapidly absorbed and achieved a peak level before 1 h | Broccoli | [96] | |
| Triple negative breast cancer | Orthotopic mouse xenograft model | The addition of sulforaphane can prevent the expansion and clearance of breast CSCs, which will greatly benefit the treatment of TNBC with cytotoxic chemotherapy | [97] | ||||
| Cellulose | Selenium | ———— | Prostate cancer | LNCaP cells | Selenium-induced growth inhibition and apoptosis in PC-3 prostate cancer cells were found to be dose dependent | ———— | [102] |
| Calcium | ———— | Colon cancer | Rats | Inhibited colonic epithelial cell proliferation induced by heme in rats, which suggested that calcium might decrease the colon cancer risk related to high intake of red meat | [105] | ||
| Colon cancer | Human | Intravenous CA/MG can be used as an effective neuroprotectant against the accumulation of SNT in adjuvant colon cancer induced by oxaliplatin | ———— | [106] | |||
| 1, 25-Dihydroxy vitamin D3 |  | Melanoma tumor | Melanoma cells | Decreased cell proliferation was found in melanoma cells | ———— | [108] | |
| Prostate cancer | Human | Slow the rate of prostate specific antigen (PSA) rise in PCa patients demonstrating proof of concept that 1,25(OH)2D3 exhibits therapeutic activity in men with PCa | [109] | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, L.; Zhang, M.; Zhao, R.; Wang, D.; Ma, Y.; Ai, L. Plant Natural Products: Promising Resources for Cancer Chemoprevention. Molecules 2021, 26, 933. https://doi.org/10.3390/molecules26040933
Ma L, Zhang M, Zhao R, Wang D, Ma Y, Ai L. Plant Natural Products: Promising Resources for Cancer Chemoprevention. Molecules. 2021; 26(4):933. https://doi.org/10.3390/molecules26040933
Chicago/Turabian StyleMa, Li, Mengmeng Zhang, Rong Zhao, Dan Wang, Yuerong Ma, and Li Ai. 2021. "Plant Natural Products: Promising Resources for Cancer Chemoprevention" Molecules 26, no. 4: 933. https://doi.org/10.3390/molecules26040933
APA StyleMa, L., Zhang, M., Zhao, R., Wang, D., Ma, Y., & Ai, L. (2021). Plant Natural Products: Promising Resources for Cancer Chemoprevention. Molecules, 26(4), 933. https://doi.org/10.3390/molecules26040933