
Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae) †


Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. CHC Profiles
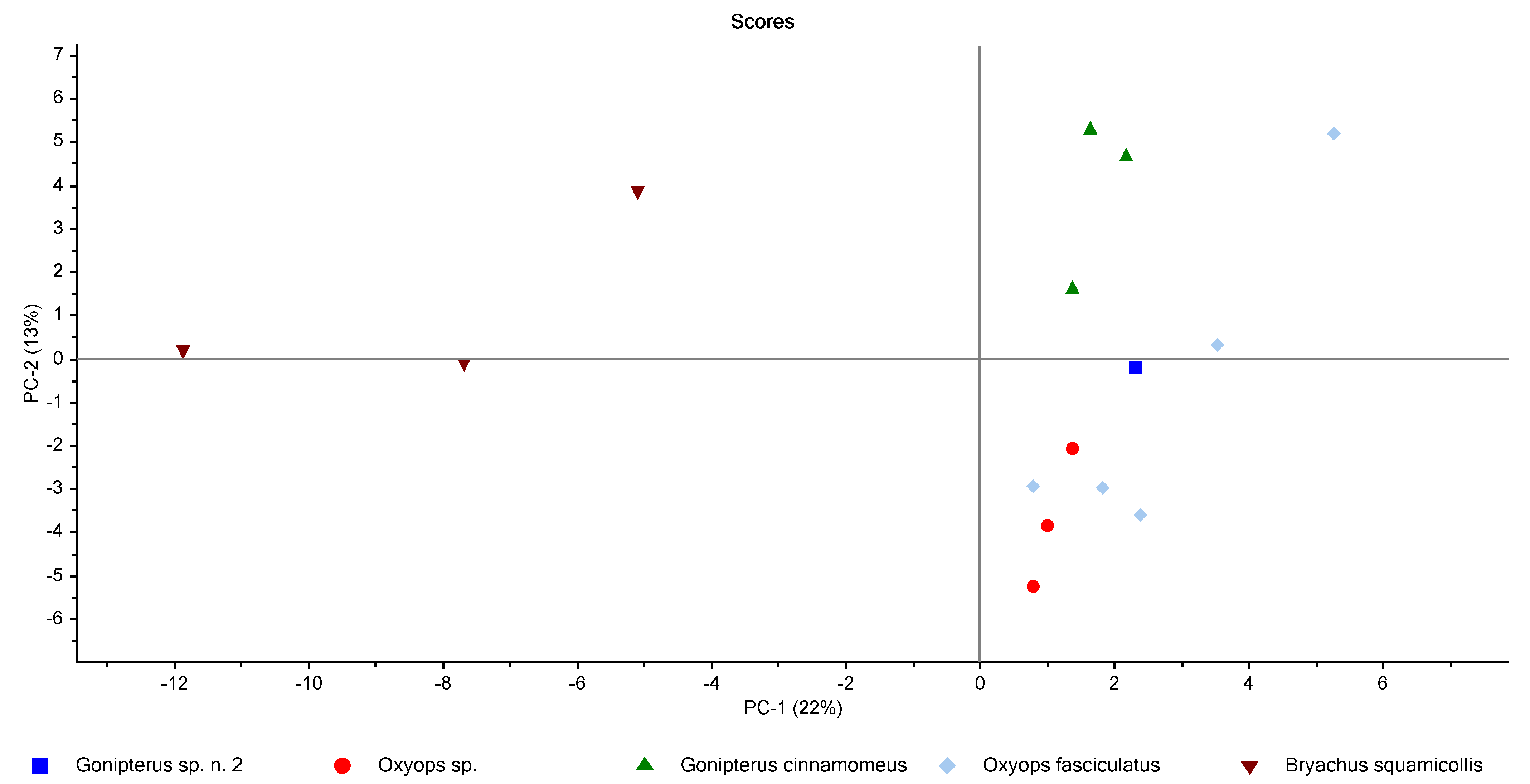
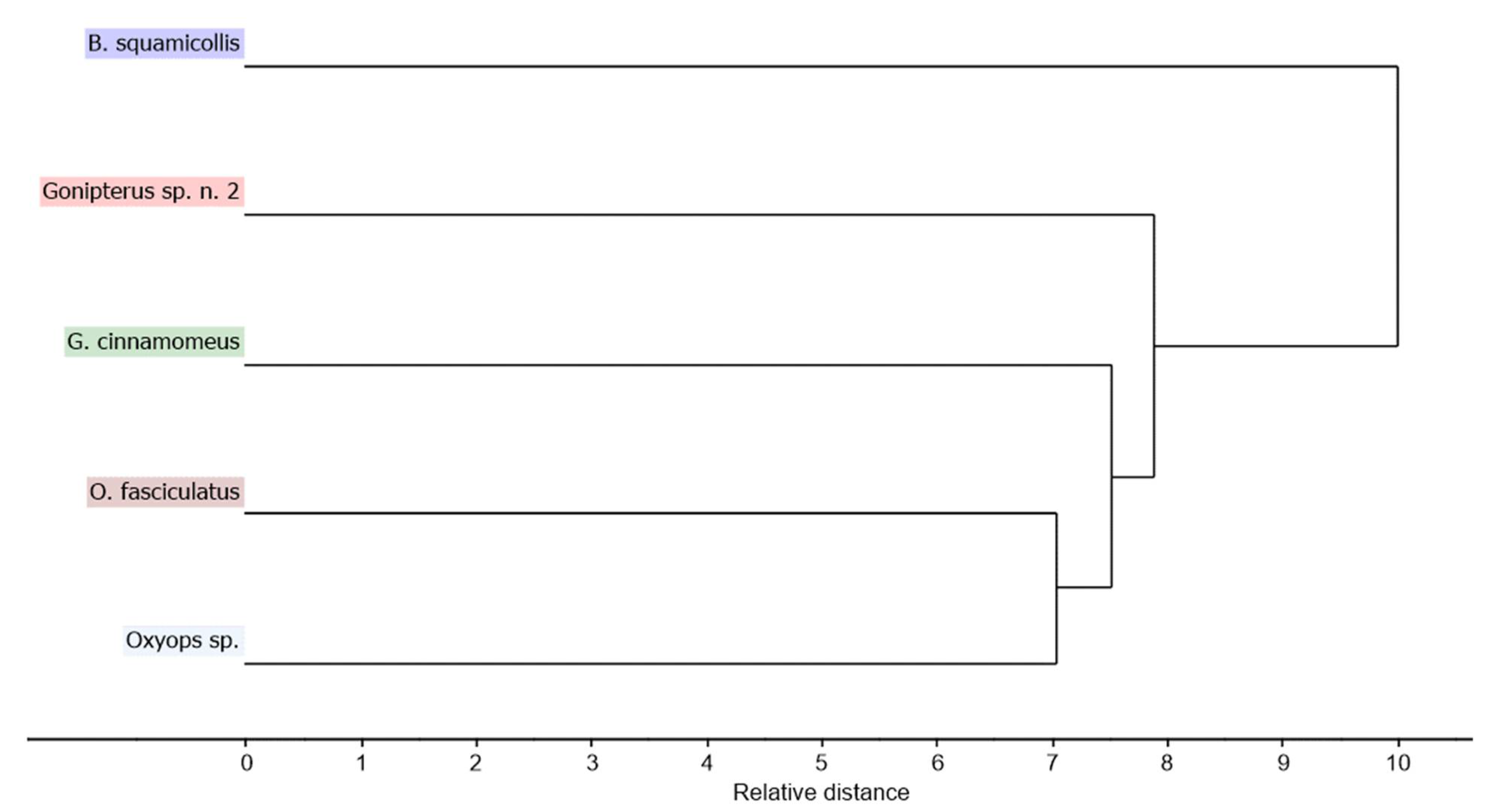
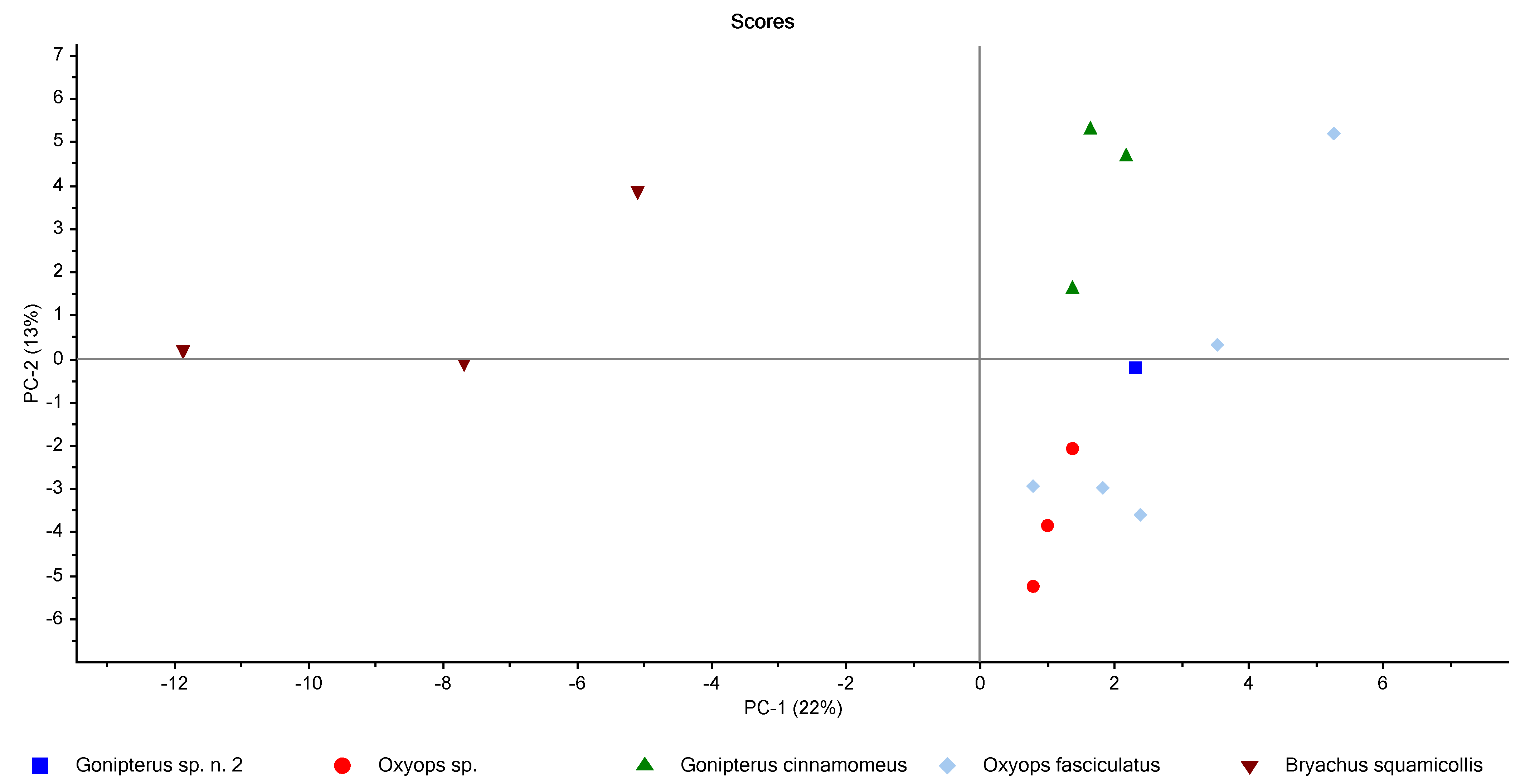
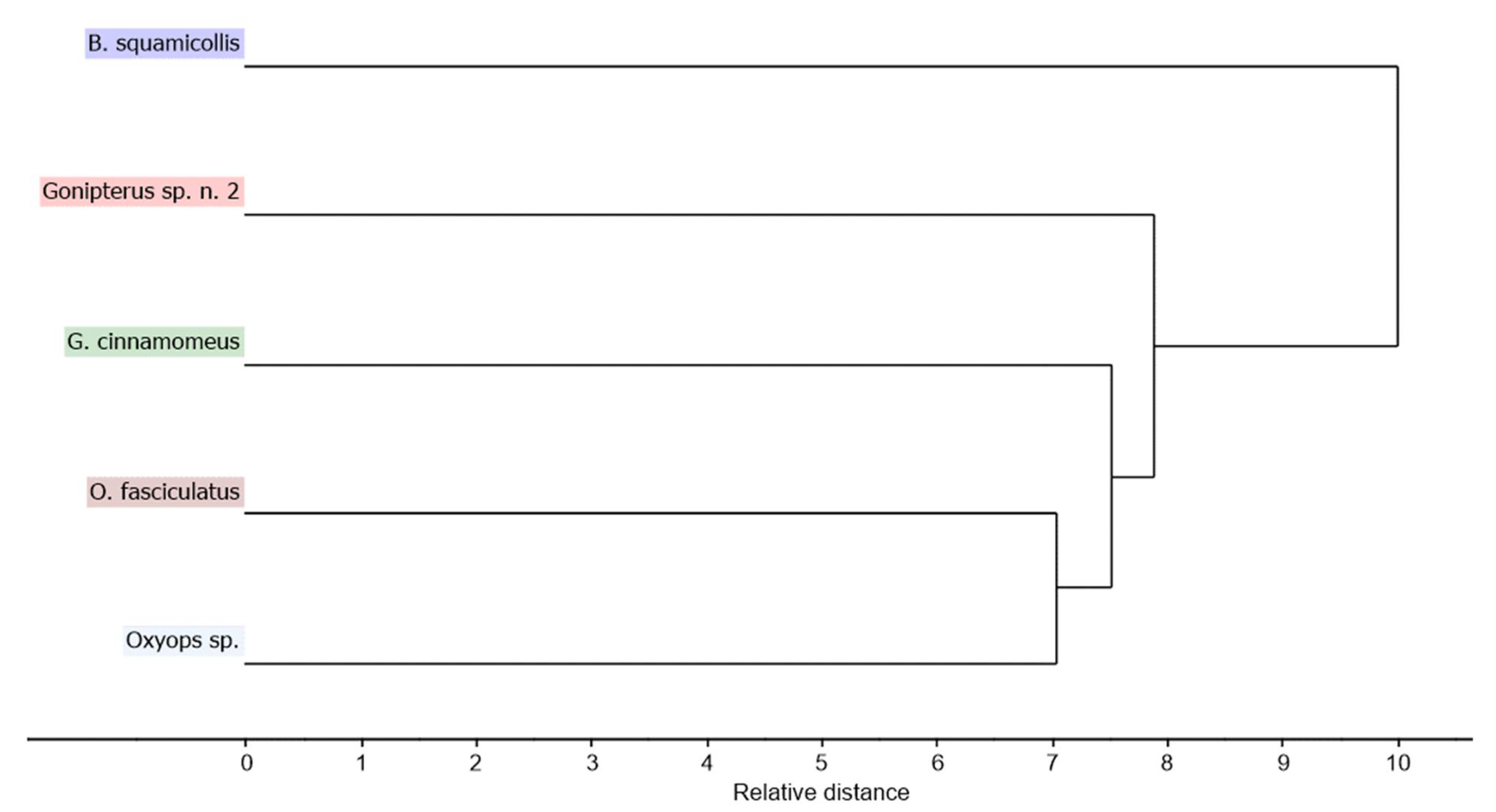
3.2. Chemometric Analysis
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schröder, M.L.; Slippers, B.; Wingfield, M.J.; Hurley, B.P. Invasion history and management of Eucalyptus snout beetles in the Gonipterus scutellatus species complex. J. Pest Sci. 2020, 93, 11–25. [Google Scholar] [CrossRef]
- Mapondera, T.S.; Burgess, T.; Matsuki, M.; Oberprieler, R.G. Identification and molecular phylogenetics of the cryptic species of the Gonipterus scutellatus complex (Coleoptera: Curculionidae: Gonipterini). Aust. J. Entomol. 2012, 51, 175–188. [Google Scholar] [CrossRef]
- Wheeler, G.S.; Massey, L.M.; Southwell, I.A. Antipredator Defense of Biological Control Agent Oxyops vitiosa Is Mediated by Plant Volatiles Sequestered from the Host Plant Melaleuca quinquenervia. J. Chem. Ecol. 2002, 28, 297–315. [Google Scholar] [CrossRef] [PubMed]
- Martin, S.; Drijfhout, F. A Review of Ant Cuticular Hydrocarbons. J. Chem. Ecol. 2009, 35, 1151. [Google Scholar] [CrossRef] [PubMed]
- Holze, H.; Schrader, L.; Buellesbach, J. Advances in deciphering the genetic basis of insect cuticular hydrocarbon biosynthesis and variation. Heredity 2021, 126, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Sprenger, P.P.; Hartke, J.; Schmitt, T.; Menzel, F.; Feldmeyer, B. Candidate genes involved in cuticular hydrocarbon differentiation between cryptic, parabiotic ant species. G3 2021, 11, jkab078. [Google Scholar] [CrossRef] [PubMed]
- Page, M.; Nelson, L.J.; Haverty, M.I.; Blomquist, G.J. Cuticular Hydrocarbons as Chemotaxonomic Characters for Bark Beetles: Dendroctonus ponderosae, D. jeffreyi, D. brevicomis, and D. frontalis (Coleoptera: Scolytidae). Ann. Entomol. Soc. Am. 1990, 83, 892–901. [Google Scholar] [CrossRef]
- Jacob, J.; Hanssen, H.-P. Distribution and variability of cuticular hydrocarbons within the Coleoptera. Biochem. Syst. Ecol. 1986, 14, 207–210. [Google Scholar] [CrossRef]
- Souza, N.M.; Schröder, M.L.; Hayes, R.A.; Bello, J.E.; Nahrung, H.F. Cuticular hydrocarbons of Gonipterus weevils: Are there species differences? Chemoecology. 2021, 31, 159–167. [Google Scholar] [CrossRef]
- Lapointe, S.L.; Hunter, W.B.; Alessandro, R.T. Cuticular hydrocarbons on elytra of the Diaprepes root weevil Diaprepes abbreviatus (L.) (Coleoptera: Curculionidae). Agric. For. Entomol. 2004, 6, 251–257. [Google Scholar] [CrossRef]
- Mackley, J.W.; Carlson, D.A.; Butler, J.F. Identification of the cuticular hydrocarbons of the horn fly and assays for attraction. J. Chem. Ecol. 1981, 7, 669–683. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M. The Pherobase: Database of Insect Pheromones and Semiochemicals. Available online: https://www.pherobase.com/ (accessed on 7 July 2021).
- R Core Team. R: A Language and Environment for Statistical Computing (Version 4.0.2); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Species | No. Specimens | Length (mm) | Width (mm) |
|---|---|---|---|
| Oxyops fasciculatus | 5 | 6.4 ± 0.4 | 3.4 ± 0.2 |
| Oxyops sp. (undescribed) | 3 | 7.6 ± 0.5 | 4.1 ± 0.3 |
| Gonipterus sp. n. 2 (tentative ID) | 1 | 7.8 | 4.2 |
| Gonipterus cinnamomeus | 3 | 6.6 ± 0.2 | 3.7 ± 0.2 |
| Bryachus squamicollis | 3 | 9.9 ± 0.8 | 5.0 ± 0.5 |
| No. | Compound | LRI | M+ (m/z) | Ident. ^ | B. squamicollis (n = 3) | G. cinnamomeus (n = 3) | G. sp. n. 2 (n = 1) | O. fasciculatus (n = 5) | Oxyops sp. (n = 3) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 3-hexanone | 787 | 100 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.08 | 0.05 ± 0.04 | 0.04 ± 0.00 |
| 2 | 2-hexanone | 791 | 100 | MS, LRI | 0.02 ± 0.02 | 0.07 ± 0.01 | 0.07 | 0.06 ± 0.04 | 0.04 ± 0.01 |
| 3 | 2,4-dimethylheptane | 820 | 128 | MS, LRI | 0.02 ± 0.01 | 0.02 ± 0.02 | 0.05 | 0.03 ± 0.02 | 0.02 ± 0.00 |
| 4 | Heptanal | 902 | 114 | MS, LRI | 0.03 ± 0.01 | 0.03 ± 0.02 | 0 | 0 | 0 |
| 5 | Octanal | 1002 | 128 | MS, LRI | 0.01 ± 0.01 | 0 | 0 | 0 | 0 |
| 6 | Eucalyptol | 1033 | 154 | MS, LRI | 0 | 0 | 0.10 | 0.01 ± 0.02 | 0 |
| 7 | 3,6-dimethyldecane | 1055 | 170 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.02 | 0.06 | 0.05 ± 0.03 | 0.03 ± 0.01 |
| 8 | 2,6,8-trimethyldecane | 1099 | 184 | MS, LRI | 0.02 ± 0.02 | 0.02 ± 0.03 | 0 | 0.01 ± 0.03 | 0.01 ± 0.01 |
| 9 | Nonanal | 1104 | 142 | MS, LRI | 0.13 ± 0.05 | 0 | 0 | 0 | 0 |
| 10 | Decanal | 1205 | 156 | MS, LRI | 0.01 ± 0.01 | 0.02 ± 0.04 | 0 | 0 | 0 |
| 11 | Exo-2-hydroxycineole | 1228 | 170 | MS, LRI | 0.04 ± 0.02 | 0.08 ± 0.04 | 0 | 0.02 ± 0.02 | 0 |
| 12 | Tentative: 2,6,10-trimethylundecane | 1275 | 198 | MS, LRI | 0.03 ± 0.01 | 0.06 ± 0.01 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0 |
| 13 | 10-undecenal | 1282 | 168 | MS, LRI | 0.14 ± 0.17 | 0 | 0 | 0 | 0 |
| 14 | Tentative: carvacrol | 1297 | 150 | MS, LRI | 2.42 ± 3.83 | 0.04 ± 0.07 | 0 | 0.02 ± 0.04 | 0 |
| 15 | Tentative: isoascaridole | 1312 | 168 | MS, LRI | 0.01 ± 0.01 | 0 | 0 | 0 | 0 |
| 16 | Tentative: 4a-methyldecahydro-1-naphthalenol | 1319 | 168 | MS, LRI | 0.21 ± 0.23 | 0 | 0 | 0 | 0 |
| 17 | 4,6-dimethyldodecane | 1321 | 198 | MS, LRI | 0.03 ± 0.01 | 0.02 ± 0.03 | 0 | 0.03 ± 0.03 | 0.01 ± 0.02 |
| 18 | Tentative: cis-p-menth-1-en-3,8-diol | 1358 | 170 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 19 | (+)-cis,trans-nepetalactone | 1364 | 166 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 20 | Dodecanal | 1408 | 184 | MS, LRI | 0 | 0.02 ± 0.03 | 0 | 0.01 ± 0.01 | 0.01 ± 0.02 |
| 21 | Aromadendrene | 1444 | 204 | MS, LRI | 0 | 0 | 0 | 0.01 ± 0.03 | 0 |
| 22 | Unidentified alkane 1 | 1488 | - | MS | 0.01 ± 0.01 | 0.01 ± 0.02 | 0.06 | 0.05 ± 0.02 | 0.03 ± 0.00 |
| 23 | Bicyclogermacrene | 1501 | 204 | MS, LRI | 0.03 ± 0.05 | 0 | 0 | 0.02 ± 0.02 | 0 |
| 24 | Tentative: 2,6,10-trimethyltridecane | 1534 | 226 | MS, LRI | 0.01 ± 0.01 | 0.01 ± 0.02 | 0 | 0 | 0 |
| 25 | Globulol | 1592 | 222 | MS, LRI | 0 | 0 | 0.07 | 0.01 ± 0.03 | 0 |
| 26 | Tetradecanal | 1612 | 212 | MS, LRI | 0.04 ± 0.02 | 0 | 0.05 | 0.01 ± 0.02 | 0.02 ± 0.03 |
| 27 | Heptadecane | 1699 | 240 | MS, LRI | 0.03 ± 0.01 | 0.05 ± 0.01 | 0.05 | 0.04 ± 0.03 | 0.02 ± 0.02 |
| 28 | Phytane | 1743 | 282 | MS, LRI | 0.01 ± 0.02 | 0 | 0 | 0 | 0 |
| 29 | Cis-9-hexadecenal | 1795 | 220 | MS, LRI | 0.05 ± 0.01 | 0 | 0 | 0 | 0 |
| 30 | Hexadecanal | 1816 | 240 | MS, LRI | 0.17 ± 0.05 | 0.09 ± 0.04 | 0.10 | 0.16 ± 0.13 | 0.13 ± 0.08 |
| 31 | 6,10,14-trimethyl-2-pentadecanone | 1840 | 268 | MS, LRI | 0 | 0 | 0 | 0 | 0.01 ± 0.02 |
| 32 | 2-heptadecanone | 1899 | 254 | MS, LRI | 0.09 ± 0.02 | 0.07 ± 0.03 | 0.14 | 0.04 ± 0.05 | 0.06 ± 0.02 |
| 33 | Tentative: 2,2-dimethyloctadecane | 1910 | 282 | MS, LRI | 0.04 ± 0.02 | 0.06 ± 0.06 | 0.09 | 0.03 ± 0.03 | 0.02 ± 0.03 |
| 34 | Heptadecanal | 1918 | 254 | MS, LRI | 0.05 ± 0.01 | 0 | 0 | 0 | 0 |
| 35 | Tentative: 3-ethyl-3-methylheptadecane | 1953 | 283 | MS, LRI | 0.03 ± 0.02 | 0.06 ± 0.02 | 0.05 | 0.06 ± 0.03 | 0.03 ± 0.00 |
| 36 | 9-octadecanone | 1990 | 268 | MS, LRI | 0 | 0 | 0 | 0.1 ± 0.1 | 0 |
| 37 | Cis-13-octadecenal | 1995 | 266 | MS, LRI | 0.12 ± 0.03 | 0 | 0 | 0 | 0 |
| 38 | Tentative: cis-9-octadecenal | 2014 | 266 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 39 | Octadecanal | 2019 | 268 | MS, LRI | 0.61 ± 0.14 | 0.07 ± 0.04 | 0.13 | 0.19 ± 0.18 | 0.16 ± 0.13 |
| 40 | Cis-2-octadecen-1-ol acetate | 2074 | 310 | MS, LRI | 0 | 0 | 0 | 0.01 ± 0.01 | 0 |
| 41 | 2-nonadecanone | 2098 | 282 | MS, LRI | 0.09 ± 0.02 | 0.04 ± 0.04 | 0.18 | 0.02 ± 0.05 | 0.08 ± 0.05 |
| 42 | Nonadecanal | 2117 | 282 | MS, LRI | 0.06 ± 0.01 | 0 | 0 | 0 | 0 |
| 43 | Unidentified alkane 2 | 2128 | - | MS | 0.23 ± 0.21 | 0.25 ± 0.43 | 0 | 0.03 ± 0.07 | 0.12 ± 0.22 |
| 44 | Unidentified alkane 3 | 2139 | - | MS | 0 | 0.35 ± 0.60 | 0 | 0 | 0.23 ± 0.40 |
| 45 | Unidentified alkane 4 | 2148 | - | MS | 0.48 ± 0.46 | 0.42 ± 0.73 | 0 | 0.06 ± 0.14 | 0.28 ± 0.48 |
| 46 | Unidentified alkane 5 | 2160 | - | MS | 0.10 ± 0.08 | 0.14 ± 0.24 | 0.07 | 0.04 ± 0.02 | 0.01 ± 0.02 |
| 47 | Unidentified alkane 6 | 2168 | - | MS | 0.08 ± 0.08 | 0.09 ± 0.15 | 0 | 0 | 0.28 ± 0.49 |
| 48 | Docosane | 2197 | 310 | MS, LRI | 0.02 ± 0.02 | 0 | 0 | 0.06 ± 0.05 | 0 |
| 49 | Eicosanal | 2222 | 296 | MS, LRI | 0.09 ± 0.04 | 0 | 0.71 | 0.03 ± 0.07 | 0 |
| 50 | Unidentified alkane 7 a | 2260 | - | MS | 0 | 0 | 0 | 0 | 0.03 ± 0.03 |
| 51 | Tricosane | 2297 | 324 | MS, LRI | 0.74 ± 0.19 | 0.37 ± 0.09 | 0 | 1.35 ± 0.54 | 0.27 ± 0.06 |
| 52 | Unidentified ketone 1 | 2304 | - | MS | 0 | 0 | 0.09 | 0 | 0 |
| 53 | Henicosanal | 2326 | 310 | MS, LRI | 0 | 0 | 0.50 | 0 | 0 |
| 54 | 11-methyltricosane | 2331 | 338 | MS, LRI | 0.20 ± 0.34 | 0 | 0 | 0 | 0.02 ± 0.04 |
| 55 | Unidentified aldehyde | 2367 | - | MS | 0.02 ± 0.01 | 0 | 0 | 0 | 0 |
| 56 | 3-methyltricosane | 2374 | 339 | MS, LRI | 0.02 ± 0.03 | 0 | 0 | 0 | 0 |
| 57 | Tetracosane | 2400 | 338 | MS, LRI | 0.03 ± 0.02 | 0.07 ± 0.01 | 0.16 | 0.09 ± 0.05 | 0.02 ± 0.02 |
| 58 | Docosanal | 2430 | 324 | MS, LRI | 0.01 ± 0.01 | 0.58 ± 0.31 | 1.12 | 0 | 0 |
| 59 | Tentative: 9-methyltetracosane | 2437 | 352 | MS, LRI | 0.01 ± 0.02 | 0 | 0 | 0 | 0 |
| 60 | 2-methyltetracosane | 2473 | 352 | MS, LRI | 0 | 0.94 ± 1.64 | 0 | 0 | 0.69 ± 1.19 |
| 61 | Tentative: x-pentacosene | 2479 | 352 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 62 | Pentacosane | 2499 | 352 | MS, LRI | 1.67 ± 0.93 | 1.83 ± 0.37 | 1.82 | 4.26 ± 2.02 | 0.97 ± 0.88 |
| 63 | Unidentified ketone 2 b | 2509 | - | MS | 0 | 0.05 ± 0.09 | 0.13 | 0 | 0 |
| 64 | Tentative: 7-methylpentacosane | 2522 | 367 | MS, LRI | 0 | 0.27 ± 0.46 | 0 | 0.48 ± 0.42 | 0 |
| 65 | Tentative: 11-methylpentacosane | 2530 | 395 | MS, LRI | 0 | 0.15 ± 0.26 | 0 | 0.11 ± 0.14 | 0 |
| 66 | Tentative: 13-methylpentacosane | 2569 | 367 | MS, LRI | 0.03 ± 0.05 | 0 | 0 | 0 | 0 |
| 67 | 3-methylpentacosane | 2574 | 366 | MS, LRI | 0.26 ± 0.23 | 1.09 ± 0.64 | 0.27 | 0 | 0 |
| 68 | Tentative: 11,15-dimethylpentacosane | 2584 | 409 | MS, LRI | 0 | 0 | 0 | 0.70 ± 1.57 | 0 |
| 69 | Hexacosane | 2600 | 366 | MS, LRI | 9.04 ± 6.87 | 13.26 ± 7.6 | 14.70 | 8.30 ± 7.10 | 11.01 ± 8.76 |
| 70 | Tetracosanal | 2637 | 352 | MS, LRI | 1.95 ± 2.56 | 4.22 ± 0.91 | 0 | 0 | 0.16 ± 0.27 |
| 71 | 2-methylhexacosane | 2663 | 380 | MS, LRI | 0 | 0 | 0 | 0.03 ± 0.07 | 0 |
| 72 | Unidentified alkane 8 | 2672 | - | MS | 0 | 0.51 ± 0.46 | 0 | 0 | 0.15 ± 0.15 |
| 73 | Tentative: 13-methylhexacosane | 2682 | 381 | MS, LRI | 0.65 ± 1.12 | 0.02 ± 0.04 | 0 | 0.88 ± 1.37 | 0 |
| 74 | Unidentified alkane 9 | 2690 | - | MS | 0.36 ± 0.62 | 0 | 0 | 0 | 0 |
| 75 | Heptacosane | 2704 | 380 | MS, LRI | 11.32 ± 2.95 | 4.3 ± 4.94 | 2.27 | 1.54 ± 3.44 | 0.36 ± 0.62 |
| 76 | Tentative: 7-methylheptacosane | 2712 | 395 | MS, LRI | 21.49 ± 5.33 | 28.94 ± 7.24 | 25.45 | 27.58 ± 6.29 | 16.77 ± 6.68 |
| 77 | Unidentified ketone 3 | 2723 | - | MS | 0 | 0.75 ± 0.31 | 0 | 0 | 0 |
| 78 | 13-methylheptacosane | 2737 | 394 | MS, LRI | 0.32 ± 0.06 | 0.07 ± 0.13 | 0 | 0.08 ± 0.15 | 0.73 ± 0.24 |
| 79 | Unidentified alkane 10 | 2755 | - | MS | 1.14 ± 1.2 | 0 | 0 | 0.01 ± 0.02 | 0.10 ± 0.17 |
| 80 | Tentative: 11-methylheptacosane | 2759 | 395 | MS, LRI | 0 | 4.60 ± 4.16 | 0 | 1.18 ± 2.49 | 0.14 ± 0.23 |
| 81 | 2-methylheptacosane | 2764 | 394 | MS, LRI | 0 | 0 | 0 | 0 | 0.08 ± 0.07 |
| 82 | Docosyl pentyl ether | 2770 | 396 | MS, LRI | 0 | 0 | 0 | 0 | 0.27 ± 0.47 |
| 83 | 3-methylheptacosane | 2774 | 394 | MS, LRI | 0 | 3.14 ± 1.61 | 2.82 | 0 | 0 |
| 84 | Tentative: 5,15- or 5,17-dimethylheptacosane | 2777 | 409 | MS, LRI | 1.06 ± 1.07 | 0 | 0 | 0 | 0 |
| 85 | Tentative: 5,11-dimethylheptacosane | 2784 | 409 | MS, LRI | 0.36 ± 0.62 | 1.93 ± 3.35 | 0 | 3.93 ± 7.21 | 0.79 ± 1.37 |
| 86 | Octacosane | 2800 | 394 | MS, LRI | 6.89 ± 2.27 | 3.55 ± 0.89 | 19.10 | 4.16 ± 0.85 | 4.43 ± 1.34 |
| 87 | Squalene | 2811 | 384 | MS, LRI | 0.38 ± 0.65 | 0.36 ± 0.37 | 0.71 | 0.43 ± 0.20 | 0.27 ± 0.08 |
| 88 | Tentative: 12-methyloctacosane | 2830 | 409 | MS, LRI | 0.22 ± 0.14 | 0.05 ± 0.08 | 0 | 0 | 0 |
| 89 | Hexacosanal | 2837 | 380 | MS, LRI | 0.94 ± 0.24 | 0.94 ± 0.3 | 0 | 0 | 0 |
| 90 | Tentative: x-methyloctacosane | 2858 | 408 | MS, LRI | 0.05 ± 0.05 | 0 | 0.11 | 0 | 0 |
| 91 | 2-methyloctacosane | 2865 | 408 | MS, LRI | 0.24 ± 0.14 | 0.93 ± 0.22 | 0.96 | 0 | 8.28 ± 6.05 |
| 92 | Nonacosene | 2881 | 407 | MS, LRI | 1.03 ± 0.40 | 0 | 0 | 0 | 0 |
| 93 | 1-hexacosanol | 2890 | 382 | MS, LRI | 0 | 0.21 ± 0.20 | 0 | 0 | 0 |
| 94 | Nonacosane | 2918 | 408 | MS, LRI | 26.54 ± 5.05 | 17.34 ± 3.08 | 24.90 | 42.99 ± 14.09 | 51.41 ± 13.58 |
| 95 | Triacontane | 2982 | 422 | MS, LRI | 5.72 ± 0.63 | 7.31 ± 3.07 | 2.79 | 0.51 ± 0.37 | 1.38 ± 0.68 |
| 96 | Tentative: x,12-dimethylnonacosane | 3002 | 437 | MS, LRI | 0.30 ± 0.50 | 0 | 0 | 0.01 ± 0.02 | 0 |
| 97 | Tentative: 2-methyltriacontane | 3039 | 437 | MS, LRI | 1.38 ± 0.43 | 0 | 0 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, J.B. Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biol. Life Sci. Forum 2022, 15, 5. https://doi.org/10.3390/IECD2022-12383
Johnson JB. Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biology and Life Sciences Forum. 2022; 15(1):5. https://doi.org/10.3390/IECD2022-12383
Chicago/Turabian StyleJohnson, Joel B. 2022. "Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae)" Biology and Life Sciences Forum 15, no. 1: 5. https://doi.org/10.3390/IECD2022-12383
APA StyleJohnson, J. B. (2022). Cuticular Hydrocarbon Profiling Reveals Chemotaxonomic Diversity among Gonipterini weevils (Coleoptera: Curculionidae). Biology and Life Sciences Forum, 15(1), 5. https://doi.org/10.3390/IECD2022-12383