
Analysis of Peroxiredoxin 5 (SmPrx5) Function and Expression in Immune and Oxidative Stress Responses of Sepiella maindroni


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Bacterial Challenge of S. maindroni In Vivo
2.3. Thermal Temperature Stress of S. maindroni
2.4. Full-Length cDNA Amplification of SmPrx5
2.5. Sequence Analysis of SmPrx5
2.6. Spatial and Temporal Expression Analysis of SmPrx5
2.7. Recombinant Expression and Purification of SmPrx5
2.8. Antioxidant Properties of SmPrx5 In Vitro
3. Results
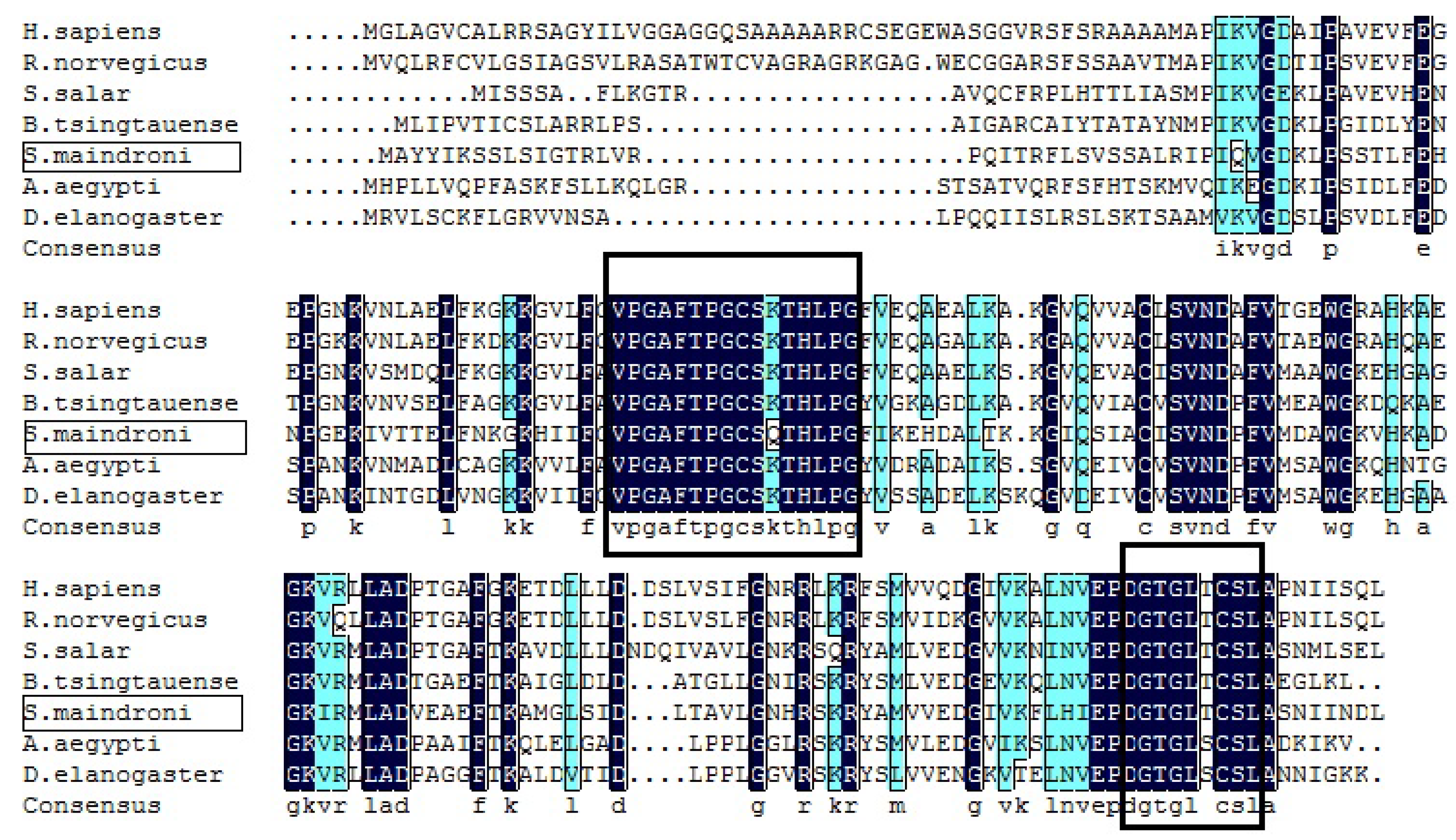
3.1. Sequence and Phylogenetic Analysis of SmPrx5
3.2. Tissue Distribution of SmPrx5
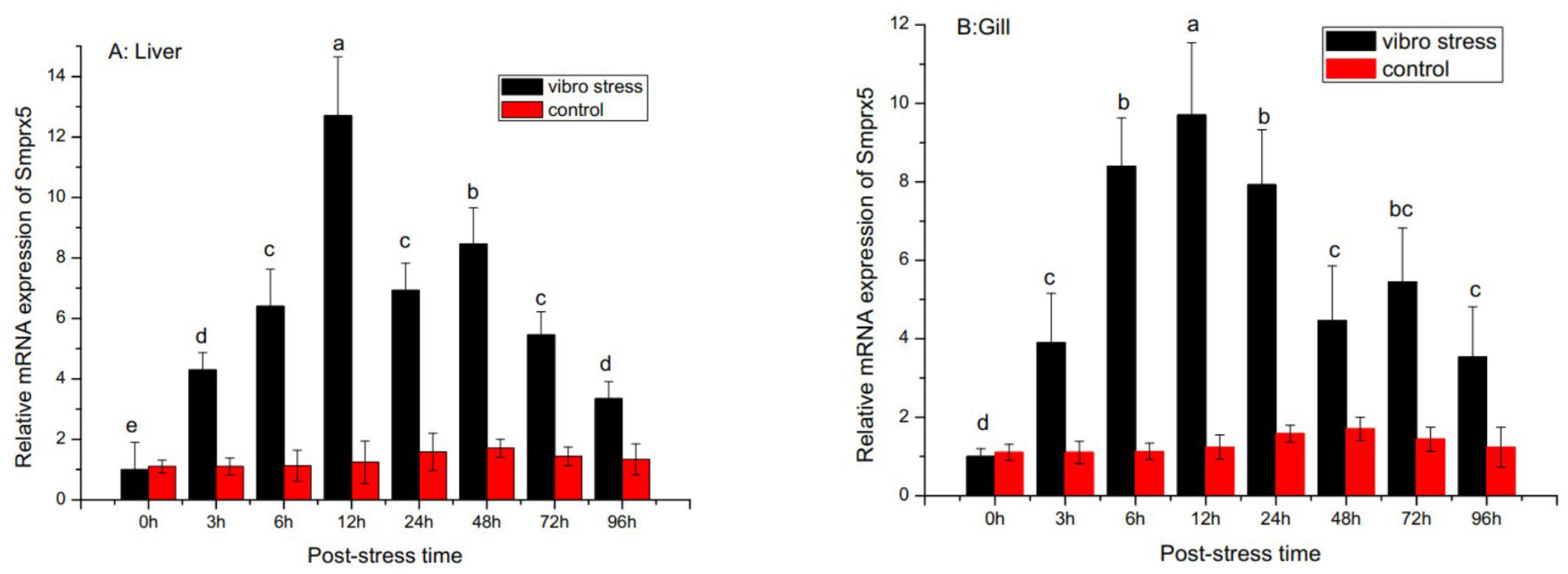
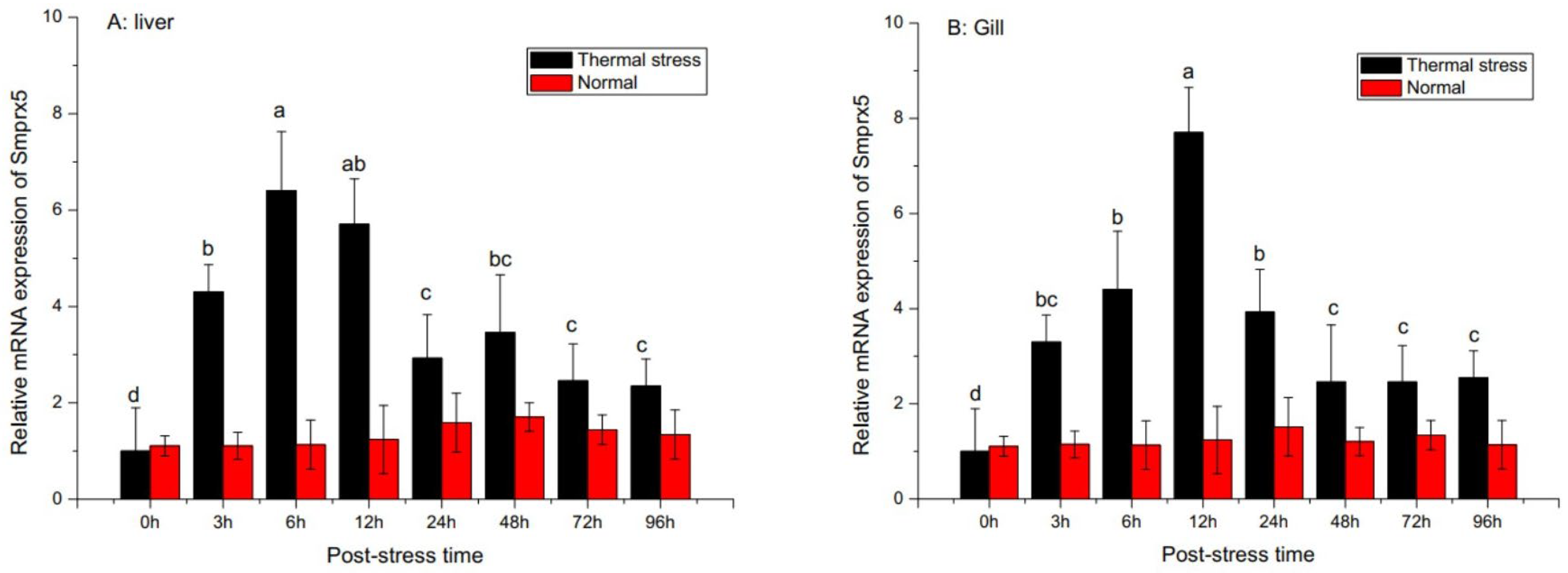
3.3. SmPrx5 Expression Following Bacterial Challenge and Thermal Stress
3.4. Characterization of Recombinant Protein
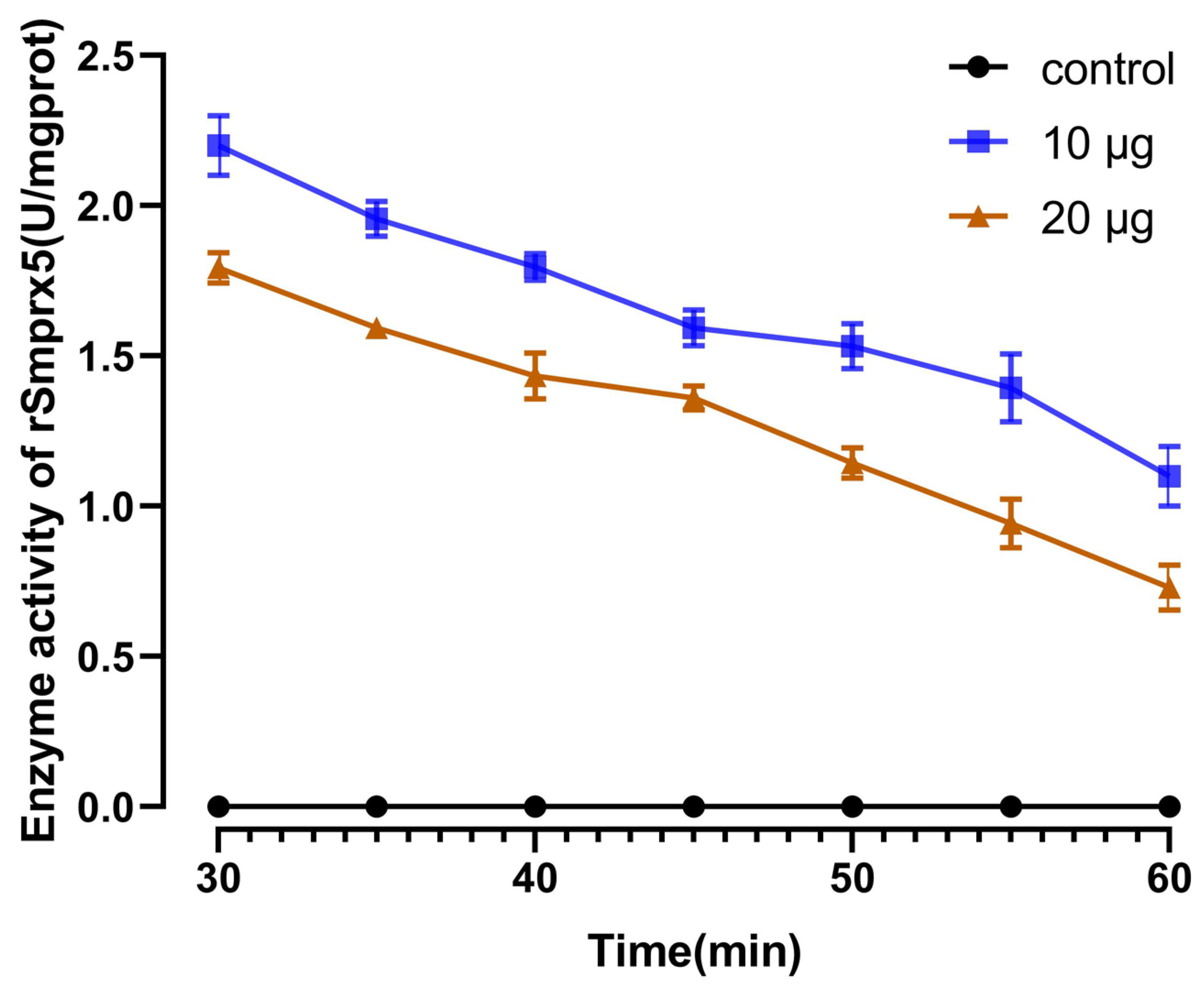
3.5. Antioxidant Activity of rSmPrx5
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roch, P. Defense mechanisms and disease prevention in farmed marine invertebrates. Aquaculture 1999, 172, 125–145. [Google Scholar] [CrossRef]
- Poynton, R.A.; Hampton, M.B. Peroxiredoxins as biomarkers of oxidative stress. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 906–912. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.-J.; Hu, Y.-H.; Zhang, M.; Sun, L. Analysis of the expression and antioxidative property of a peroxiredoxin 6 from Scophthalmus maximus. Fish Shellfish Immunol. 2010, 29, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Wood, Z.A.; Schroder, E.; Harris, J.R.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef]
- Robinson, M.W.; Hutchinson, A.T.; Dalton, J.P.; Donnelly, S. Peroxiredoxin: A central player in immune modulation. Parasite Immunol. 2010, 32, 305–313. [Google Scholar] [CrossRef]
- Seo, M.S.; Kang, S.W.; Kim, K.; Baines, I.C.; Lee, T.H.; Rhee, S.G. Identification of a New Type of Mammalian Peroxiredoxin That Forms an Intramolecular Disulfide as a Reaction Intermediate. J. Biol. Chem. 2000, 275, 20346–20354. [Google Scholar] [CrossRef]
- Knoops, B.; Loumaye, E.; Van Der Eecken, V. Evolution of the peroxiredoxins. Subcell. Biochem. 2007, 44, 27–40. [Google Scholar]
- Declercq, J.-P.; Evrard, C.; Clippe, A.; Stricht, D.V.; Bernard, A.; Knoops, B. Crystal structure of human peroxiredoxin 5, a novel type of mammalian peroxiredoxin at 1.5 Å resolution. J. Mol. Biol. 2001, 311, 751–759. [Google Scholar] [CrossRef]
- Knoops, B.; Goemaere, J.; Van der Eecken, V.; Declercq, J.-P. Peroxiredoxin 5: Structure, Mechanism, and Function of the Mammalian Atypical 2-Cys Peroxiredoxin. Antioxidants Redox Signal. 2011, 15, 817–829. [Google Scholar] [CrossRef]
- Perez-Sanchez, J.; Bermejo-Nogales, A.; Calduch-Giner, J.A.; Kaushik, S.; Sitja-Bobadilla, A. Molecular characterization and expression analysis of six peroxiredoxin paralogous genes in gilthead sea bream (Sparus aurata): Insights from fish exposed to dietary, pathogen and confinement stressors. Fish Shellfish Immunol. 2011, 31, 294–302. [Google Scholar] [CrossRef]
- Duan, Y.; Liu, P.; Li, J.; Li, J.; Gao, B.; Chen, P. cDNA cloning, characterization and expression analysis of peroxiredoxin 5 gene in the ridgetail white prawn Exopalaemon carinicauda. Mol. Biol. Rep. 2013, 40, 6569–6577. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.; Zhang, S.; Zhang, G. Cloning, genomic structure, and expression analysis of peroxiredoxin V from bay scallop Argopecten irradians. Fish Shellfish Immunol. 2011, 30, 309–316. [Google Scholar] [CrossRef]
- Wan, H.; Kang, T.; Zhan, S.; You, H.; Zhu, F.; Lee, K.S.; Zhao, H.; Jin, B.R.; Li, J. Peroxiredoxin 5 from common cutworm (Spodoptera litura) acts as a potent antioxidant enzyme. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 175, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.-M.; Yoo, J.-W.; Lee, S.; Lee, H.J.; Lee, H.-S.; Lee, D.-S. Peroxiredoxin 5 promotes the epithelial-mesenchymal transition in colon cancer. Biochem. Biophys. Res. Commun. 2017, 487, 580–586. [Google Scholar] [CrossRef]
- Feng, X.; Liu, J.; Fan, S.; Liu, F.; Li, Y.; Jin, Y.; Bai, L.; Yang, Z. The identification of goat peroxiredoxin-5 and the evaluation and enhancement of its stability by nanoparticle formation. Sci. Rep. 2016, 6, 24467. [Google Scholar] [CrossRef]
- Sienko, J.; Gaj, P.; Czajkowski, K.; Nowis, D. Peroxiredoxin-5 is a negative survival predictor in ovarian cancer. Ginekol. Polska 2019, 90, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Castellanos-Martínez, S.; Gestal, C. Pathogens and immune response of cephalopods. J. Exp. Mar. Biol. Ecol. 2013, 447, 14–22. [Google Scholar] [CrossRef]
- Vidal, E.A.; Villanueva, R.; Andrade, J.P.; Gleadall, I.G.; Iglesias, J.; Koueta, N.; Rosas, C.; Segawa, S.; Grasse, B.; Franco-Santos, R.M.; et al. Cephalopod culture: Current status of main biological models and research priorities. Adv. Mar. Biol. 2014, 67, 90–98. [Google Scholar]
- Müller, V.; de Boer, R.J.; Bonhoeffer, S.; Szathmáry, E. An evolutionary perspective on the systems of adaptive immunity. Biol. Rev. 2017, 93, 505–528. [Google Scholar] [CrossRef]
- Fiorito, G.; Affuso, A.; Basil, J.; Cole, A.; de Girolamo, P.; D’Angelo, L.; Mark, F. Guidelines for the care and welfare of cephalopods in research: A consensus based on an initiative by CephRes, FELASA and the Boyd Group. Lab Anim. 2015, 49 (Suppl. 2), 1–90. [Google Scholar] [CrossRef]
- Cui, M.; Zhang, Q.; Yao, Z.; Zhang, Z.; Zhang, H.; Wang, Y. Immunoglobulin M gene expression analysis of orange-spotted grouper, Epinephelus coioides, following heat shock and Vibrio alginolyticus challenge. Fish Shellfish Immunol. 2010, 29, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.-M.; Lu, Z.-R.; He, H.-J.; Ye, P.-L.; Ying, Z.; Wang, C.-L. Effects of several ecological factors on the hatching of Sepiella maindroni wild and cultured eggs. Chin. J. Appl. Ecol. 2010, 21, 1321–1326. [Google Scholar]
- Schultz, J.; Milpetz, F.; Bork, P.; Ponting, C.P. SMART, a simple modular architecture research tool: Identification of signaling domains. Proc. Natl. Acad. Sci. USA 1998, 95, 5857–5864. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Godahewa, G.I.; Perera, N.C.N.; Nam, B.-H.; Lee, J. Antioxidative properties and structural features of atypical 2-Cys peroxiredoxin from Sebastes schlegelii. Dev. Comp. Immunol. 2018, 82, 152–164. [Google Scholar] [CrossRef]
- Ren, L.; Sun, Y.; Wang, R.; Xu, T. Gene structure, immune response and evolution: Comparative analysis of three 2-Cys peroxiredoxin members of miiuy croaker, Miichthys miiuy. Fish Shellfish Immunol. 2014, 36, 409–416. [Google Scholar] [CrossRef]
- Revathy, K.S.; Umasuthan, N.; Whang, I.; Jung, H.-B.; Lim, B.-S.; Nam, B.-H.; Lee, J. A potential antioxidant enzyme belonging to the atypical 2-Cys peroxiredoxin subfamily characterized from rock bream, Oplegnathus fasciatus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 187, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.-H.; Kim, S.J.; Kang, S.W.; Lee, K.-K.; Rhee, S.G.; Yu, D.-Y. Molecular Cloning and Characterization of the Mouse Peroxiredoxin V Gene. Biochem. Biophys. Res. Commun. 2000, 270, 356–362. [Google Scholar] [CrossRef]
- Wu, L.; Zhou, Y.; Abbas, M.N.; Kausar, S.; Chen, Q.; Jiang, C.-X.; Dai, L.-S. Molecular structure and functional characterization of the peroxiredoxin 5 in Procambarus clarkii following LPS and Poly I:C challenge. Fish Shellfish Immunol. 2017, 71, 28–34. [Google Scholar] [CrossRef]
- Tu, D.-D.; Jiang, M.; Gu, W.-B.; Zhou, Y.-L.; Zhu, Q.-H.; Zhou, Z.-K.; Chen, Y.-Y.; Shu, M.-A. Identification and characterization of atypical 2-cysteine peroxiredoxins from mud crab Scylla paramamosain: The first evidence of two peroxiredoxin 5 genes in non-primate species and their involvement in immune defense against pathogen infection. Fish Shellfish Immunol. 2017, 69, 119–127. [Google Scholar] [CrossRef]
- Cong, M.; Ni, D.; Song, L.; Wang, L.; Zhao, J.; Qiu, L.; Li, L. Molecular cloning, characterization and mRNA expression of peroxiredoxin in Zhikong scallop Chlamys farreri. Mol. Biol. Rep. 2008, 36, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, S.; Lee, B.-L. Recent Advances in the Innate Immunity of Invertebrate Animals. BMB Rep. 2005, 38, 128–150. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Murrell, G.A.; Trickett, A.; Landtmeters, M.; Knoops, B.; Wang, M.X. Overexpression of antioxidant enzyme peroxiredoxin 5 protects human tendon cells against apoptosis and loss of cellular function during oxidative stress. Biochim. Biophys. Acta 2004, 1693, 37–45. [Google Scholar] [CrossRef]
- Lu, J.L.; Vallat, J.-M.; Pollard, J.D.; Knoops, B.; Ouvrier, R. Expression of the antioxidant enzyme peroxiredoxin 5 in the human peripheral nervous system. J. Peripher. Nerv. Syst. 2006, 11, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Baković, J.; Yu, B.Y.K.; Silva, D.; Chew, S.P.; Kim, S.; Ahn, S.-H.; Palmer, L.; Aloum, L.; Stanzani, G.; Malanchuk, O.; et al. A key metabolic integrator, coenzyme A, modulates the activity of peroxiredoxin 5 via covalent modification. Mol. Cell. Biochem. 2019, 461, 91–102. [Google Scholar] [CrossRef]
- Hu, H.; Tian, M.; Li, P.; Bao, Y.; Guan, X.; Lian, Z.; Yin, Y.; Ding, C.; Yu, S. Brucella infection regulates peroxiredoxin-5 protein expression to facilitate intracellular survival by reducing the production of nitric oxide and reactive oxygen species. Biochem. Biophys. Res. Commun. 2019, 516, 82–88. [Google Scholar] [CrossRef]
- Pushpamali, W.A.; De Zoysa, M.; Kang, H.S.; Oh, C.H.; Whang, I.; Kim, S.J.; Lee, J. Comparative study of two thioredoxin peroxidases from disk abalone (Haliotis discus discus): Cloning, recombinant protein purification, characterization of antioxidant activities and expression analysis. Fish Shellfish Immunol. 2008, 24, 294–307. [Google Scholar] [CrossRef]
- Radyuk, S.N.; Michalak, K.; Klichko, V.I.; Benes, J.; Orr, W.C. Peroxiredoxin 5 modulates immune response in Drosophila. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2010, 1800, 1153–1163. [Google Scholar] [CrossRef]
- Wang, X.; Hu, B.; Wen, C.; Zhang, M.; Jian, S.; Yang, G. Molecular cloning, expression and antioxidative activity of 2-cys-peroxiredoxin from freshwater mussel Cristaria plicata. Fish Shellfish Immunol. 2017, 66, 254–263. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, F.; Zhang, J.; Wang, B.; Gao, H.; Huang, B.; Jiang, H.; Xiang, J. Molecular cloning, expression of a peroxiredoxin gene in Chinese shrimp Fenneropenaeus chinensis and the antioxidant activity of its recombinant protein. Mol. Immunol. 2007, 44, 3501–3509. [Google Scholar] [CrossRef]
- Arockiaraj, J.; Easwvaran, S.; Vanaraja, P.; Singh, A.; Othman, R.Y.; Bhassu, S. Immunological role of thiol-dependent peroxiredoxin gene in Macrobrachium rosenbergii. Fish Shellfish Immunol. 2012, 33, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Park, H.; Ahn, I.-Y.; Kim, H.; Cheon, J.; Kim, M. Analysis of ESTs and expression of two peroxiredoxins in the thermally stressed Antarctic bivalve Laternula elliptica. Fish Shellfish Immunol. 2008, 25, 550–559. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.N.; Kausar, S.; Cui, H. The biological role of peroxiredoxins in innate immune responses of aquatic invertebrates. Fish Shellfish Immunol. 2019, 89, 91–97. [Google Scholar] [CrossRef]
- Kim, M.H.; Lee, H.J.; Lee, S.R.; Lee, H.S.; Huh, J.W.; Bae, Y.C.; Lee, D.S. Peroxiredoxin 5 inhibits glutamate-induced neuronal cell death through the regulation of calcineurin-dependent mitochondrial dynamics in HT22 cells. Mol. Cell Biol. 2019, 39, e00045-19. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.G.; Kam, M.K.; Kim, K.M.; Kim, H.S.; Kwon, O.-S.; Lee, H.-S.; Lee, D.-S. Peroxiredoxin 5 prevents iron overload-induced neuronal death by inhibiting mitochondrial fragmentation and endoplasmic reticulum stress in mouse hippocampal HT-22 cells. Int. J. Biochem. Cell Biol. 2018, 102, 10–19. [Google Scholar] [CrossRef]
- Pirson, M.; Clippe, A.; Knoops, B. The curious case of peroxiredoxin-5: What its absence in aves can tell us and how it can be used. BMC Evol. Biol. 2018, 18, 18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | Application |
|---|---|---|
| SmPrx5-3GSP1 | GTTCCTGCATATTGAGCCTGAT | 3′RACE |
| SmPrx5-3GSP2 | TACCGTCGTTCCACTAGTGATTT | 3′RACE |
| SmPrx5-F1 | ATCCAGAAAACCTTTGCATGCAGAA | qRT-PCR |
| SmPrx5-R1 | CAAAAGATATATGAGGGAGTGC | qRT-PCR |
| GAPDH-F | TGGTTCCTTGGCTTTTGCT | qRT-PCR |
| GAPDH-F | GGTGGTGGTGCGGGTAGT | qRT-PCR |
| rSmPrx5-F | CATATGGCGTACTACATAAAATCGTCCT | Recombinant expression |
| rSmPrx5-R | CTCGAGTTAGTGGTGGTGGTGGTGGTGCAAGTCATTAATGATGTTGGAA | Recombinant expression |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, C.; Song, W.; Wang, C. Analysis of Peroxiredoxin 5 (SmPrx5) Function and Expression in Immune and Oxidative Stress Responses of Sepiella maindroni. Fishes 2025, 10, 289. https://doi.org/10.3390/fishes10060289
Shao C, Song W, Wang C. Analysis of Peroxiredoxin 5 (SmPrx5) Function and Expression in Immune and Oxidative Stress Responses of Sepiella maindroni. Fishes. 2025; 10(6):289. https://doi.org/10.3390/fishes10060289
Chicago/Turabian StyleShao, Chu, Weiwei Song, and Chunlin Wang. 2025. "Analysis of Peroxiredoxin 5 (SmPrx5) Function and Expression in Immune and Oxidative Stress Responses of Sepiella maindroni" Fishes 10, no. 6: 289. https://doi.org/10.3390/fishes10060289
APA StyleShao, C., Song, W., & Wang, C. (2025). Analysis of Peroxiredoxin 5 (SmPrx5) Function and Expression in Immune and Oxidative Stress Responses of Sepiella maindroni. Fishes, 10(6), 289. https://doi.org/10.3390/fishes10060289