Abstract
In recent decades, the use of nanocomposites (NCs) in crop production has been actively studied. We have previously shown that chemically synthesized selenium nanocomposites (Se NCs) based on natural polymeric matrices of arabinogalactan (Se/AG NCs), carrageenan (Se/CAR NCs), and starch (Se/ST NCs) stimulated the growth and development of potatoes in vitro and had an antibacterial effect against the phytopathogen Clavibacter sepedonicus. It is important to confirm that the results obtained in the laboratory are also reproducible in the field conditions of crop cultivation. In addition, the applications of NCs can be expanded if a wider range of their biological activity is revealed, and the effect on other types of cultivated plants and phytopathogens is shown. In this regard, the purpose of this research was to study the effect of nanopriming with Se NCs on the productivity of potatoes in a field experiment, the germination of soybeans, and the viability of the phytopathogen Pectobacterium carotovorum under in vitro conditions. The study included the use of traditional methods of conducting a field experiment in natural growing conditions, also using microbiological cultures and studying the bacterial profile from the endosphere of potato tubers by the imprint method, germinating Glycine max L. soybean seeds after nanopriming in Petri dishes, followed by the measurement of morphometric and biochemical parameters such as the activity of antioxidant enzymes and content of diene conjugates. Based on the results of field experiments, it was found that the preplant treatment of tubers with Se/AG and Se/CAR NCs stimulated a significant increase in the number of stems in potato plants and tubers per plant, but for Se/AG NC, the increase was observed only in 2020 and 2022, and the stimulating effect of Se/AG and Se/CAR NCs on the weight of tubers was observed only in 2020. In the yield structure, the proportion of marketable seed tubers was increased under the influence of all Se NCs. Endosphere cultures from tubers showed that bacteria were present in only 56% of the tubers in the control, while 78% of the tubers obtained from plants grown from seed tubers treated with Se/AG NC and 100% of the tubers from plants grown from seed tubers treated with Se/ST and Se/CAR NCs contained bacteria. The bacteria were represented by both spore-forming Gram-positive and Gram-negative bacteria of various sizes. The results presented in this article and previously published data showed that the maximum diversity of bacteria was observed after treatment with Se/ST NC. Experiments on potatoes showed that Se/AG NC had the greatest biological effect. In addition, the growth-stimulating effect of these NCs on the biomass of the roots of soybean seedlings was shown. It activated the antioxidant enzymes and reduced the level of diene conjugates (DCs), the amount of which increased significantly after seed biopriming with the phytopathogen Pectobacterium carotovorum. The antibacterial effect of Se/AG NC in relation to the P. carotovorum was observed as a reduction in the growth of the bacterial culture, biofilm formation, and dehydrogenase activity of cells. Thus, it has been shown that Se/AG NC has a stimulating effect on such agriculturally important crops as potatoes and soybeans, as well as an antibacterial effect against Gram-negative and Gram-positive phytopathogenic bacteria.
Keywords:
potato; tubers; nanocomposite; selenium; microbial profile; bacteria; seed priming; soybean; Pectobacterium carotovorum 1. Introduction
The largest share of the nutrition of mankind belongs to four agricultural crops: corn, rice, wheat, and potato [1]. Potato is actively cultivated in various countries of the world, Europe [2], and Asia [3,4], and ensures food security [5]. Soybean occupies the fifth position among agricultural crops used for nutrition [6]. Both potato and soybean play an important role in global food security [7,8]. To protect the crops of these cultivated plants, pesticides are widely used, whose excessive use has a negative impact on the environment. An alternative to pesticides is seed priming, i.e., seed treatment with various primers, such as hydropriming, osmopriming, thermopriming, solid matrix priming, biopriming, and chemopriming [9,10,11]. Seed priming increases the yield and quality potential of high-value crops by increasing plant stress resistance, making the plant more capable of coping with multiple stresses in the field [12,13,14,15]. Seed priming has been shown to result in faster seedling emergence, vigorous seedling growth, and faster seedling establishment [16]. Priming causes an increase in the activity of enzymes, such as antioxidant enzymes, amylases, proteases, and lipases, which break down macromolecules for the growth and development of the seed embryo [17,18]. Seed priming also reduces stress during the germination stage and ultimately results in higher emergence and rooting success [10,19,20]. Nanopriming is an innovative seed priming technology that helps to improve seed germination, seed growth, and yield by enhancing plant resistance to various stresses [21]. According to published data, nanopriming induces the formation of nanopores in shoots and promotes water absorption and activates ROS in seeds, which are involved in the production of secondary metabolites and stress tolerance [22]. In addition, this method of seed treatment acts as an inducer of rapid starch hydrolysis and induces aquaporin gene expression, which promotes seed germination [10].
The use of nanosubstances for seed priming is promising due to the high biological activity of nanoparticles (NPs) and their low volume. Nanotechnology has offered an advanced seed priming technology for smart agriculture. For example, nanoemulsions of turmeric oil and silver nanoparticles (Ag NPs) synthesized from agro-industrial by-products have been used as nanoprimers for watermelon, Citrullus lanatus, seeds, raising their germination rate and yield by 32–35% compared to the control [23]. The nanopriming of chickpea seeds, Cicer arietinum, with ZnO NPs significantly increased the antioxidant activity of germinated plants in an experiment with potted plants in a greenhouse, and their infection by the phytopathogen Fusarium oxysporum was 90% lower than in the control [24]. The treatment of chickpea seeds with ZnO NPs promoted the formation of sugars, phenol, total proteins, and superoxide dismutase (SOD), which increased resistance to the wilt pathogen [24]. Other researchers have shown that seed treatment with ZnO NPs significantly improved plant growth, biomass accumulation, and photosynthesis in maize, Zea mays, under cobalt (Co) stress [25]. Nanopriming with ZnO NPs reduced the accumulation of reactive oxygen species (ROS) and malondialdehyde (MDA) in corn shoots. More importantly, ZnO NPs attenuated the toxic effect of Co, reducing its uptake and imparting stability to plant ultracellular structures and the photosynthetic apparatus. In addition, seedlings primed with ZnO NPs were found to have a higher accumulation of nutrients and antioxidant enzymes [25]. Chinese cabbage seed treatment with Ag NP significantly accelerated seed germination and seedling development and increased cabbage yield by 44.3% [26]. It was found that such seed treatment increased the content of essential amino acids in cabbage leaves several times, including alanine, aspartic acid, glutamine, glutamic acid, histidine, isoleucine, leucine, lysine, phenylalanine, proline, serine, threonine, tyrosine, and valine. At the same time, seed priming with Ag NPs significantly increased the content of iron (Fe) by 23.8% in cabbage leaves [26]. The stimulating effect of seed priming with gold nanoparticles (Au NPs) on the photosynthetic apparatus of Triticum aestivum seedlings was shown [27]. In plants grown from treated seeds, ultrastructural changes in chloroplasts were found: an increase in the size of plastids, starch grains, granules in chloroplasts, and the number of thylakoids in granules. The intensity of photosynthesis, the content of chlorophylls, and the proportion of unsaturated fatty acids in the composition of total leaf lipids also increased in the treated plants [27].
Earlier, we studied the biological activity of selenium nanocomposites (Se NCs) packed in natural polymer matrices (carrageenan, arabinogalactan, and starch). It was revealed that these agents reduce the viability of the phytopathogenic bacterium Clavibacter sepedonicus and phytopathogenic fungi Phytophthora [28,29]. It was shown that after the treatment of plants with Se NCs, selenium did not accumulate in their tissues [30,31,32]. It was also shown that chemically synthesized Se NCs based on natural polymer matrices did not inhibit the viability of soil microorganisms, which tentatively indicated that they are environmentally safe for practical use [33].
Based on the unique physical and chemical properties of NPs and the aforementioned advantages of NCs as an effective delivery mechanism in biological systems, we further investigated how tuber nanopriming with Se NCs will affect the vegetation and productivity of potatoes in field experiments and morphometric and biochemical characteristics of soybean seedlings in a laboratory experiment.
2. Materials and Methods
2.1. Nanocomposites
The following NCs were used in the research: arabinogalactan-based Se NC (Se/AG NC, 5.92% Se), starch-based Se NC (Se/ST NC, 1.46% Se), and kappa-carrageenan-based Se NC (Se/CAR NC, 3.67% Se), which were described in detail in Perfileva et al. [33,34]. The most effective concentrations were selected based on previous studies by Perfileva et al. [33].
2.2. Potato Nanopriming and Field Experiment Conditions
Potato tubers, Solanum tuberosum L., representing the early maturing and high-yielding variety “Gala” were used in the study. There were control and three nanopriming treatments of tubers with Se/AG, Se/ST NC, and Se/CAR NCs in April 2020, 2021, and 2022 by spraying with aqueous solutions of Se NCs and 30 tubers per treatment and control. Control tubers were sprayed with water. Then, they were germinated in the dark for 14 days at 22 °C. Every year the tubers were planted in the same field at the same experimental site of the Siberian Institute of Plant Physiology and Biochemistry, Siberian Branch of the Russian Academy of Sciences (SIFIBR SB RAS, Irkutsk, Russia). There were four variants in total, with three plots for each variant and ten plants per plot. All experiments were carried out on a natural infectious background in a field with three plots (repeats) per field with a randomized plot design. The plot area was 100 m2 (20 × 5 m). The plots were located in the same conditions on a flat, unshaded area. The duration of each experiment was 90 days. The experiment was set up on gray forest soil, typical for the southern part of Eastern Siberia, with the following agrochemical parameters: humus content—7–8% (C), total nitrogen—0.13%, mobile phosphorus (P2O5)—159 mg/kg, exchangeable potassium (K2O)—139 mg/kg, pH KCl = 5.6. In general, in terms of fertility, the soil was characterized as slightly acidic, rather humus-rich. The soil was loam and clay, highly cultivated with a humidity level of 10% and a density of 1.39 g/cm3. Agrochemical analyses of soil and plants were performed according to [35]. The common crop cultivation agrotechniques for this region were used in the natural conditions of the sharply continental climate of Eastern Siberia (Irkutsk, Russia): the cultivation sites were periodically weeded and hilled without additional irrigation and fertilization. The number of branches in each plant was estimated in the middle of the growing season. The weight and number of tubers obtained from experimental plants, as well as the structure of the crop and the number of rotted tubers, were analyzed at the end of the growing season. To determine the structure of the crop, the tubers were ranked by weight according to the federal standards (GOST 33996-2016 and 7176-2017) into categories used in potato production: tubers weighing 85–150 g were ranked as commercial and 50–80 g—as seed tubers. After analysis, the tubers obtained from the field experiment were placed in mesh bags and stored in containers at an air temperature of +4 °C and a humidity of 75%.
2.3. Analysis of Microorganisms Collected from Potato Tubers
In October-November 2022, the microorganisms from potato tubers were analyzed using the modified replica plating method [36]. Briefly, the tubers were sterilized for 1 min in 96% ethanol, then 20 min in a 3% peroxide solution, then they were washed three times with sterile water and dried. With a sterile scalpel, the tuber was cut across. A slice of potato tuber was laid out on a Petri dish containing meat-peptone (MP) agar media with nystatin. Nystatin was used to prevent the overgrowth of fungal cultures. In a series of sowing, 10 tubers from each of the 3 plots for each of the 3 treatment variants and control were tested, or 120 in total (10 × 3 × 4 = 120). After 3 days, the appearance of colonies of microorganisms was monitored. Next, the isolation of pure cultures was carried out by their repeated subculturing on MP agar media. The bacterial colonies were stained according to Gram and described morphologically using microscopy performed on an inverted fluorescent microscope AxioObserver Z1 (Carl Zeiss, Jena, Germany), evaluating the shape, length, and width of cells and the presence of spores.
2.4. Experiments with Bacteria Pectobacterium carotovorum
The study was also conducted using specifically pathogenic bacteria Pectobacterium carotovorum, which causes soft rot disease in various plant hosts and blackleg in potato by degradation of the plant cell wall. The bacterial strain VKM B-1274 was obtained from the All-Russian Collection of Microorganisms, Pushchino, Moscow Region. The bacterial culture of P. carotovorum was cultivated on MP agar media (State Research Center for Applied Microbiology and Biotechnology, Obolensk, Moscow Region, Russia). Bacteria were transferred to a fresh nutrient medium after 7–10 days and cultivated in a thermostat in the dark, at a temperature of 25 °C. After 5–7 passages, the bacteria were sown on a potato-glucose decoction from 200 g of potatoes with the addition of glucose (20 g/L) and agar-agar (17–20 g/L) to preserve their virulent properties. A liquid culture of P. carotovorum was grown on a shaker (80 rpm) in flasks with MP broth in the dark at 26 °C.
The effect of Se NCs was assessed using several methods: (1) determining the turbidity of the optical density of a bacterial suspension (bacteria were incubated on MP agar media with Se NC for 72 h, periodically measuring the optical density of the bacterial suspension on a Bio-Rad iMark Microplate Absorbance Reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA) at 595 nm), (2) the method of circles (wells were made in a Petri dish of MP agar media, and aqueous solution of Se NCs was poured into the wells while water was poured into the control wells; then, a bacterial culture was planted on the dish, and the inhibition of bacterial growth was assessed by the presence of an inhibition zone around the well [37]), (3) the study of biofilm formation (bacteria were incubated on MP agar media with Se NC for 24 h, then dropped onto a sterile polystyrene plate, which was incubated for 2 days at 26 °C; then the biofilms were stained with gentian violet dye (Lenreaktiv, St. Petersburg, Russia) followed by three washings of the cells with water to remove the unabsorbed dye; finally, the gentian violet remaining in the bacterial cells was extracted with 96% ethanol and its optical density was measured on a spectrophotometer at a wavelength of 595 nm) [38], and (4) assessing the intensity of cell respiration by measuring the dehydrogenase activity of cells using 2,3,5-triphenyltetrazolium chloride (TTC) (Sigma-Aldrich, Saint Louis, MO, USA) [39]. To do this, the bacterial suspension was incubated for 24 h with Se NC, then centrifuged at 14,500 rpm, the precipitate was poured with TTC for 24 h, and then it was washed twice with water, poured with 96% ethanol, and incubated on a thermoshaker at 60 °C for 15 min. Finally, the optical density of the samples was measured on a spectrophotometer at a wavelength of 490 nm. TTC is able to bind with reducing agents formed during the respiration of NAD(P)H2, forming an insoluble precipitate of intense crimson color [39].
2.5. Soybean Nanopriming
The nanopriming study was carried out on seedlings of soybean (Glycine max (L.)) variety “Sayana”, which is characterized by increased cold and frost resistance. It belongs to the varieties with an early ripeness that mature in 98 days, on average. The height of this plant variety is up to 105–147 cm. The northern ecotype “Sayana” is characterized by significant resistance to low air and soil temperatures during the germination phase and increased productivity in long-day conditions of insufficient heat supply and cold stress [40].
To study the effect of nanopriming of soybean seeds on the intensity of seed germination, the following series of experiments was carried out. For each treatment and control, 30 seeds were used. Seeds were sterilized for 1 min in 96% ethanol, then 20 min in 3% hydrogen peroxide. Then, the seeds were washed three times with sterile water. For biopriming, the seeds were soaked in a bacterial suspension of P. carotovorum for two hours; the control was soaked in a beef-peptone broth. Next, nanopriming was carried out; the seeds were soaked for 30 min in an aqueous solution of Se/AG NC, and the control was soaked in water. After that, the seeds were dried and planted on two layers of moistened filter paper in Petri dishes with 10 seeds per dish. The seeds were incubated in a thermostat at 23 °C under a humidity of 70%. After 7 days, the number of non-germinated seeds was counted, and growth (length and weight of the seedling and roots) and biochemical traits were measured. The peroxidase activity in potato tissues was determined according to the Boyarkin method [41]. The analysis of catalase activity in plants was carried out using spectrophotometry by carrying out a color reaction between hydrogen peroxide and ammonium molybdate with the measurement of the optical density of the reaction products at λ = 470 nm [42]. The primary products of lipid peroxidation (LPO) and diene conjugates (DCs) in shoot and root tissues of soybean seedlings were determined using hexane and isopropanol [43].
The obtained results were statistically processed using the Microsoft Excel software package and the SigmaPlot v.12.5 program (SYSTAT Software, Chicago, IL, USA). The Shapiro–Wilk test was used to check the sample for normality. The data obtained after treatment were statistically compared with controls using the nonparametric Mann–Whitney U test because not all data followed normality.
3. Results
3.1. Effect of NCs on Potato Productivity
Figure 1 presents the results of the effect of the Se/AG, Se/ST and Se/CAR NCs based on the natural polymer matrices on the average weight and number of tubers per plant. It was found that the treatment with Se/AG and Se/CAR NCs in the first year of testing increased the biomass of potato tubers but did not affect this trait in the second and third years. Se/ST NC had no effect on the weight of tubers (Figure 1).
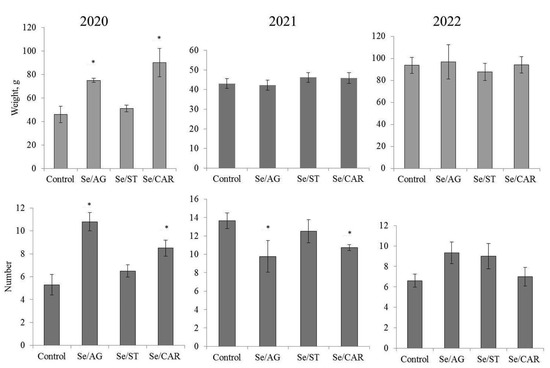
Figure 1.
Results of the field experiments in 2020, 2021, and 2022 on the effect of nanopriming of tubers with three Se NCs based on the natural polymer matrices (Se/AG, Se/ST, and Se/CAR NCs, respectively) on potato productivity (average weight and number of tubers per plant). Significant differences from control according to the Mann–Whitney U-test: * p < 0.01.
It was found that Se/AG and Se/CAR NCs significantly increased the number of tubers in the first and third years of testing but reduced it in the second year of testing, while Se/ST NC had no effect on this trait (Figure 1).
Table 1 presents the results of the analysis of the potato crop structure in terms of the percentage of marketable and seed tubers. The Se NC treatment did not affect the percentage of seed tubers. Although Se/AG NC increased the percentage of marketable and seed tubers in the crop structure in 2020 and 2021, and Se/CAR NC in 2020 and Se/ST NC in 2021 compared to the control, the effect was very heterogeneous across different years and insignificant, on average, for three years.

Table 1.
The structure of the potato crop (the percentage of marketable and seed tubers) according to the results of three years of field trials.
In addition, in 2022, we estimated the aboveground number of stems in potato plants. Se NC treatment was found to stimulate it. The average number of branches per plant was 2.7 ± 0.3 in the control, 3.1 ± 0.2 after treatment with Se/AG NC, 3.6 ± 0.4 with Se/ST NC, and 3.4 ± 0.4 with Se/CAR NC. It seems that the Se NC treatment promoted this trait, but more studies in multiple years are needed to confirm this trend.
3.2. Effect of NC on the Microbial Profile of Potato Tubers
The results show that both Gram-positive and Gram-negative bacilli of various sizes, single, V-shaped, and connected in pairs in chains, were sown from the tubers. Coccobacilli were rarely sown. Almost all the inoculated bacteria contained exospore endoliths. Part of the inoculated bacteria grew better on potato agar (PA) than on MP. In the control variant (tubers obtained from seed material not subjected to the NC treatment), bacteria were sown from 56%. From the tubers of this variant, eight different types of bacteria differing in morphology were sown (Figure 2).
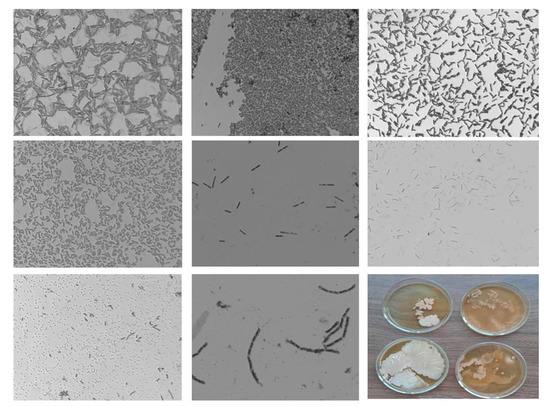
Figure 2.
Photos of bacteria sown from control potato tubers (1000× zoom).
The cell size in the control variant varied from 1.89 to 5.02 μm in length and from 0.49 to 1.26 μm in width. Different species on a Petri dish with MP mostly grew separately from each other, without competition. Visually, some cultures in the control were morphologically similar to bacteria of the genera Geobacillus, Gammaproteobacteria, Bacillus ssp., and Bacillus subtilis.
In the Se/AG NC variant, bacterial colonies were seeded from 89% of the studied samples (Figure 3). According to the morphological characteristics of the cells, nine species of bacteria were observed after treatment with Se/ST NC. They were visually similar to bacteria of the genera Bacillus subtilis, Geodermatophilaceae, Stenotrophomonas, and Actinobacteria. The cell size varied from 2.29 to 3.11 μm in length and from 0.4 to 0.67 μm in width. Only one sample of all planted crops grew on the PA.
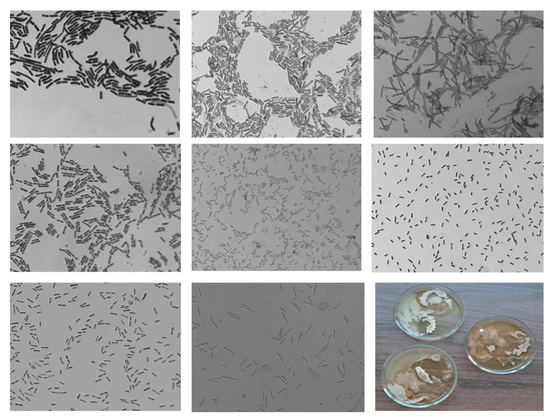
Figure 3.
Photo of bacteria sown from potato tubers treated with Se/AG NC (1000× zoom).
After treatment with Se/ST and Se/CAR NCs, bacterial colonies were developed from all the studied samples (Figure 4 and Figure 5). We found the maximum diversity in morphological features after the Se/ST NC treatment.
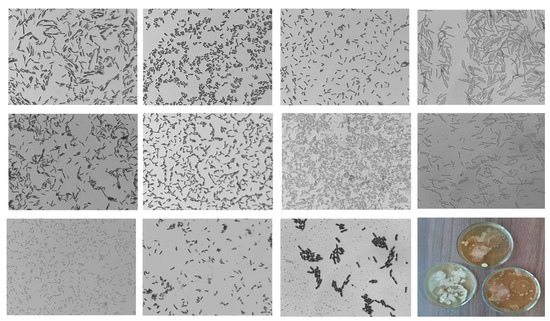
Figure 4.
Photo of bacteria sown from potato tubers treated with Se/ST NC (1000× zoom).
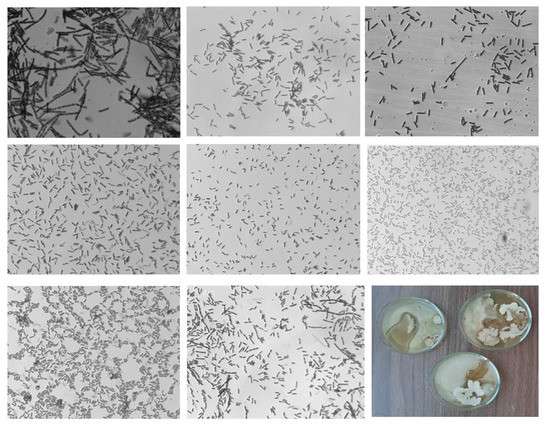
Figure 5.
Photo of bacteria sown from potato tubers treated with Se/CAR NC (1000× zoom).
The cell size after treatment with Se/ST NC varied from 0.89 to 3.38 μm in length and from 0.42 to 0.79 μm in width. Coccobacilli were observed. Bacteria sown from half of the potato tubers successfully grew on PA. Moreover, their growth rate on PA was higher, and their morphological features differed from those grown on MP. The zones of inhibition of bacterial growth were noted at the border of contact between various growing bacterial cultures (such as in Figure 6D). There were species similar to Bacillus subtilis, Gammaproteobacteria, Stenotrophomonas, and Geobacillus.
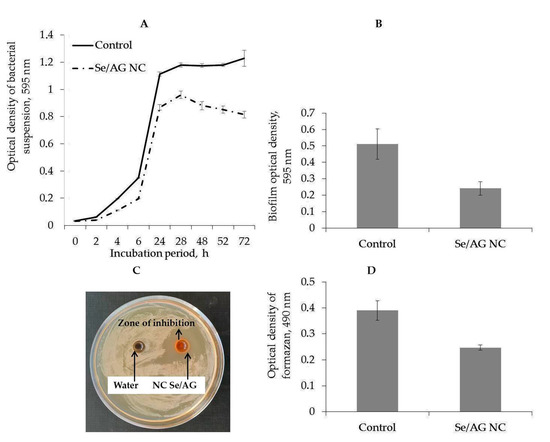
Figure 6.
Effect of Se/AG NC on viability (A), biofilm formation after 24 h of incubation with Se/AG NC (B), growth (C), and dehydrogenase activity after 24 h of incubation with Se/AG NC (D) of Pectobacterium carotovorum.
Despite the high density of bacterial colonies in the Petri dishes sown after the Se/CAR NC treatment, the diversity was less than after the Se/ST NC treatment. At the same time, among the species, an antibacterial effect of some cultures on others was found (Figure 5). The cell size in the Se/CAR NC variant varied from 1.9 to 4.22 μm in length and from 0.54 to 1.3 μm in width. The samples of potatoes treated with Se/ST NC contained the largest bacterial cells of all that were sown from all other tubers. Only one sample sown from all tubers grew on the PA. Zones of inhibition of bacterial growth were also found at the border of contact between various growing bacterial cultures in this experiment (such as in Figure 6D). Based on morphological traits, bacteria from this treatment visually resembled bacteria from the genera Bacillus, Stenotrophomonas, Variovorax, and Geobacillus.
3.3. Influence of Se/AG NC on Phytopathogen Pectobacterium carotovorum
Thus, based on the results described above, Se/AG NC, as a nanocomposite with the highest biological activity, was chosen to study its effect on the viability of the phytopathogen P. carotovorum and the germination of soybean seeds. P. carotovorum is a Gram-negative, motile, phytopathogenic bacterium that has been reported to cause infectious soft rot disease in various crop species, including post-harvest vegetables, such as Chinese cabbage, lettuce, radish, potatoes, tomatoes, and onions [44]. In potatoes, P. carotovorum causes blackleg disease, which leads to wilting of the entire plant, as well as blackening and necrosis of the stem [45]. The search for effective agents to control this phytopathogen is extremely important since most of the pesticides used have antifungal, but not antibacterial, effects.
The bacteriostatic effect of Se/AG NC on P. carotovorum was assessed in the early stages. The turbidity of the bacterial suspension was assessed in dynamics over 72 h.
It was found that Se/AG NC had a bactericidal effect already at the logarithmic phase of bacterial growth after 2 h of co-incubation of bacteria with Se/AG NC. After 24 h of co-incubation, their number decreased by 26% compared with the control, and after 72 h by 35% (Figure 6A). After 24 h of co-incubation, the intensity of bacterial biofilm formation decreased by almost twice compared with the control (Figure 6B).
The study of the bactericidal effect of Se/AG NC on P. carotovorum showed the zone of inhibition around Se/AG NC with 1.04 ± 0.03 cm in size (Figure 6C).
The bacteriostatic, antibiofilm, and bactericidal effects of Se/AG NC may be associated with the suppression of bacterial respiration. This is evidenced by the decrease in the activity of NAD(P)H dehydrogenase that was also observed (Figure 6D).
3.4. Influence of Se/AG NC on Soybean Germination
After 7 days of germination of soybean seeds, and after their soaking for 30 min in an aqueous solution of Se/AG NC, seed germination was visually assessed (Figure 7), and growth traits, such as root and hypocotyl length, and root and hypocotyl mass, were measured (Table 2).

Figure 7.
Effect of biopriming with Pectobacterium carotovorum and nanopriming with Se/AG NC on soybean germination after 7 days of germination.
Table 2.
Influence of Se/Ag NC on the length and weight hypocotyl and roots in soybean seedlings.
The results of the study showed that Se/AG NC had both stimulating and healing effects on seed germination. The biopriming of soybean seeds with a bacterial suspension of P. carotovorum increased the number of unsprouted seeds compared to the control by 75%. While nanopriming of bioprimed seeds with Se/AG NC significantly reduced the negative effect of the phytopathogen (Table 2). Se/AG NC also had a stimulating effect on the growth traits of NC by the weight of the roots. Biopriming of P. carotovorum seeds led to a significant decrease in all growth traits measured in soybean seedlings. However, nanopriming of bioprimed seeds with Se/AG NC reduced the negative effect of the phytopathogen on the growth and development of soybean seedlings (Table 2).
Measurements of biochemical traits in the tissues of soybean roots and hypocotyls are presented in Table 3. Antioxidant enzymes are indicative markers of stress, both biotic and abiotic. Therefore, peroxidase activity and catalase activity in the tissues of the hypocotyl and root of soybean seedlings were used as biochemical indicators of the stress response of the seedlings. It was found that after nanopriming, peroxidase activity significantly increased in soybean roots, and catalase activity increased in hypocotyl but decreased in roots compared with the control (Table 3). Seed biopriming with P. carotovorum caused a significant increase in peroxidase activity in roots and a five-fold increase in catalase activity in hypocotyls (Table 3). The combined effect of biopriming and nanopriming on soybean seedlings also stimulated peroxidase activity in hypocotyls and catalase activity in both hypocotyls and roots. However, this increase was lower than with biopriming alone, although not statistically significantly except for the DC content, which indicates induced lipids’ peroxidation (Table 3).
Table 3.
Influence of NC Se/Ag on the biochemical traits of soybean seedlings.
LPO is a consequence of oxidative stress caused by ROS. These free radicals trigger a cascade of reactions leading to the disruption of the barrier functions of biological membranes and the appearance of DCs. DCs are the primary products of LPO, which lead to the appearance of secondary products and complete loss of cell membranes. Therefore, this will lead to cell apoptosis [43]. We have estimated the content of DC in the tissues of soybean seedlings. It was found that Se/AG NC significantly reduced the amount of DC in hypocotyls (Table 3). Biopriming significantly, by five-fold, increased the amount of DC in the roots compared to the control. The combined effect of biopriming and nanopriming on soybean seedlings reduced the negative impact of the phytopathogen in roots, reducing the amount of DCs (Table 3).
4. Discussion
In this work, the effect of nanopriming of planting potato tubers with Se NCs based on natural polymeric matrices was studied for three years. It was found that during the growing season in 2022, the number of branches in plants grown from treated tubers was higher than in the control by 15–33%. Stimulation of the development of the biomass of aboveground organs under the influence of NPs is widely described in the literature. For example, an increase in vegetative growth and seed yield of Arabidopsis thaliana after growing seeds on a medium with mesoporous silica NPs and watering plants with it during the growing season was shown [46]. The use of foliar nanofertilizers based on Mn-Zn ferrite NPs when sowing pumpkin (Cucurbita pepo L.) led to an increase in yield of up to 53% compared to the control [47]. Seed treatment of wheat Triticum aestivum with Fe3O4 NPs stimulated seedling growth by increasing photosynthesis and respiration, antioxidant activity, and improved availability of minerals Fe and P [48]. It has been demonstrated that Fe NPs promote the growth of cucumber (Capsicum annuum) by changing the organization of leaves and increasing the number of chloroplasts and granule stacking, as well as regulating the development of vascular bundles [49]. Fe NPs can be taken up by roots and then transported to the central cylinder in bioavailable forms, where they are translocated and utilized by leaves and stems [49]. Stimulation of the intensity of growth of Basella alba seedlings and elongation of their roots under the influence of Fe NPs in combination with zinc (Zn) NPs was shown [50]. Stevia rebaudiana biomass accumulation was observed when NPs of bimetallic copper (Cu) and gold (Au) alloys were added to the Murashige–Skoog medium [51]. ZnO NPs [52] and Ag NPs [53] increased the productivity of various cultivated plants by influencing physiological, biochemical, and antioxidant activities. Nanosulfur obtained by green synthesis using Melia azedarach aqueous leaf extract and citric acid stimulated the growth of the biomass of the roots and aboveground parts of Cucurbita pepo plants in the field experiment [54].
It was demonstrated that nanoselenium compounds could also have a stimulating effect on plant biomass. For example, the surface area of the leaf blade of cucumber plants grown using Se NPs was 50% higher compared to the control variant [55]. Melissa officinalis plants grown after treatment with Se NPs were characterized by a sharp increase in biomass, activation of lateral buds, and stimulation of lateral root development [56]. When studying the mechanisms of influence of Se NPs, it is assumed that an increase in the growth of higher plants under their influence occurs due to an increase in the productivity of photosynthesis [57]. A change in the fatty acid profile of lipids in plant cells was also shown under the influence of Se NPs [58].
Tuber biomass indicators in our field experiments varied depending on the Se NC and the year of the experiment. Such an indicator as the weight of tubers per plant increased only in 2020 in the variants of Se/AG and Se/CAR NCs. At the same time, the average number of tubers per plant was also higher in 2020 for the same NCs compared to the control. However, on the contrary, this parameter decreased in 2021 in the treatments with Se/AG and Se/CAR NCs compared to the control. There was no difference between control and treatments in 2022. Preplant treatment of tubers with Se/ST NC had no effect on the weight nor the number of tubers in each of the three years.
It is likely that the revealed differences in the effect of the same NC on potato productivity in different years can be associated with different climatic conditions. According to the meteorological observations, anomalous weather conditions prevailed in the Irkutsk Region during 2020, 2021, and 2022 (https://www.irmeteo.ru/index.php?id=697, accessed on 20 February 2023), but the most favorable weather conditions for potato cultivation were observed in 2020. It was in this year that the treatment with Se/AG and Se/CAR NCs stimulated both the weight and number of tubers per plant.
Thus, the results of field studies of the treatment of potato tubers with Se NCs presented in this work confirmed the previously obtained observations in vitro that Se NCs favorably affect the viability of potatoes, and some of them even stimulate growth and biomass. Treatment with Se/AG NC was the most effective among the Se NCs used in the study.
Analysis of the crop structure during three years of the field study demonstrated the absence of a negative effect of Se NC. Preplant treatment of tubers with Se NC did not increase the number of small tubers in the structure. Moreover, nanopriming with Se/AG NC in 2020 and 2022 increased the number of marketable tubers in the crop structure in 2020 and 2021—the number of seed tubers. Treatment with Se/ST NC stimulated the formation of seed tubers in 2020 and 2021 and commercial tubers in 2022. Treatment with Se/CAR NC contributed to the growth of large tubers in the crop structure in 2020 and commercial tubers in 2021 and 2022. The stimulation effect of metallic NPs on root formation accompanied by the activation of physiological processes in plant tissues was observed in several recent studies [59,60,61,62]. For example, Al-NPs stimulated root growth in radish and rapeseed [63]. Fe3O4 NPs enhanced the growth of tomato roots [64]. Se NPs increased the biomass of Melissa officinalis roots [56].
There are studies on the microbiome of potato seed tubers of various varieties that describe that the microflora from the parent tuber can pass into a new tuber [65]. Among the microorganisms isolated were Actinobacteria, accounting for ~22% of the total bacterial community, Alphaproteobacteria (~16%), Bacilli (~11%), Betaproteobacteria (~10%), and Gammaproteobacteria (~7%) [65]. Kõiv et al. [66] compared the bacterial microbiota in potato tubers infected with Pectobacterium, Atrospeticum, and uninfected tubers and found that Actinobacteria dominated the community. This is followed by Betaproteobacteria and Bacillus [66]. Moreover, Actinobacteria, Alphaproteobacteria, and Gammaproteobacteria have been described as consistently present in tubers of various potato varieties [67].
The number of morphologically different bacteria in the tubers treated with Se/CAR NC was less than in the tubers treated with Se/ST NC. It is worth noting the presence of inhibition zones at the sites of contact between colonies of different bacterial species. Apparently, such bacteria secrete some antimicrobial substances, the nature of which would be important to find out in future studies.
In a previous study, we have shown the fungicidal and antibacterial effect of Se NCs on the phytopathogenic fungus Phytophthora cactorum [28] and the Gram-positive phytopathogenic bacterium Clavibacter sepedonicus [29,33], respectively. It was found that Se NCs can reduce the growth of the C. sepedonicus bacterial culture, inhibit its biofilm formation, and increase the number of dead cells in bacterial suspension [33]. The Gram-negative bacterium P. carotovorum is widespread among phytopathogenic bacteria that infect many crops, including potatoes [43,44]. This pathogen is distributed throughout the world and has an extremely strong survival rate, being able to persist in soil or plants as a saprophyte for more than a year [68]. The applied pesticides do not effectively affect this phytopathogen; therefore, a search is underway for new substances to control this bacterium [68]. In our experiments, it was found that Se/AG NC has a bacteriostatic, antibiofilm, and bactericidal effect. Biofilm formation is a key factor in the virulence of P. carotovorum, leading to the formation of biofilms in plant pathways, blocking them. The negative effect can be caused by the suppression of bacterial respiration, as evidenced by the decrease in the dehydrogenase activity of cells under the influence of Se/AG NC. The antibacterial activity of biosynthesized chitosan NPs against P. carotovorum has been shown in [69]. It was demonstrated recently that CuS NPs inhibited the mobility of P. carotovorum [70], and Ag NPs exhibited high antibacterial activity against P. carotovorum by completely suppressing bacterial growth and biofilm formation and destroying the bacterial cell membrane [71].
In addition to the negative effect on phytopathogens, nanocompounds can promote seed germination and improve some morphological traits of seedlings [10]. A convenient species for studying the growth-stimulating effect of NCs is soybean. Both positive and negative effects of nanocompounds on soybean germination have been shown [72]. Se/AG NC had a growth-stimulating effect on the weight of the soybean seedling root and stimulated the activity of antioxidant enzymes. Nanopriming with Se/AG NC significantly reduced the amount of DCs in root tissues, both in healthy seeds and those subjected to biopriming with pathogenic fungus. The decrease in DC levels may be associated with the antioxidant property of Se NPs [73,74,75]. Biopriming of soybean seeds with a phytopathogenic P. carotovorum caused a significant decrease in their germination and hypocotyl and root traits and a significant increase in the activity of antioxidant enzymes and the level of DC, which indicates a state of high stress in the plant. Apparently, the negative effect of biopriming is associated with exoenzymes secreted by P. carotovorum. Pectinases, polygalacturonases, cellulases, and proteases secreted by the bacterium are the virulence factors that can lead to the maceration of plant tissues [68]. It was shown in our experiments that the nanopriming of seeds subjected to biopriming reduced the negative effect of biopriming. This can be likely explained by the antibacterial effect of Se/AG NCs on the phytopathogenic P. carotovorum.
5. Conclusions
Thus, the conducted studies showed the biological activity of Se NC in laboratory and field conditions and in different objects, such as vegetating potato plants, soybean seedlings, and Gram-negative phytopathogenic bacterium. Field experiments carried out over three years demonstrated that among the studied NCs, the most pronounced effect was observed for Se/AG NC. Nanopriming of potato tubers before planting with Se/AG NC increased the number of tubers per plant and, in some years, stimulated the mass of tubers obtained from the plant. In the yield structure, Se/AG NC contributed to an increase in the number of marketable and seed tubers. These data correlate with the stimulatory effect of Se/AG NC on potato viability under in vitro conditions.
The results obtained for Se/ST and Se/CAR NCs were inconsistent across three years and ambiguous. Therefore, further laboratory tests on soybeans and the phytopathogenic bacterium P. carotovorum were carried out only with Se/AG NC. It demonstrated a detrimental effect on P. carotovorum. The data obtained in this study and previously published results allow us to conclude that Se/AG NC has a wide spectrum of antiphytopathogenic activity: it inhibits the growth and development of the Gram-positive phytopathogenic bacterium Clavibacter sepedonicus, the Gram-negative phytopathogenic bacterium P. carotovorum, and the phytopathogenic fungus Phytophthora cactorum. The results obtained are important because substances with pronounced antibacterial and fungicidal activity are extremely rare. In addition, Se/AG NC had a growth-stimulating effect on soybean roots, activated the antioxidant enzymes, and significantly reduced the amount of DC in root tissues. Nanopriming reduced the negative effect of the phytopathogen on the physiological and biochemical parameters of soybean seedlings. The present data and previously obtained results on potato plants in vitro showed that nanocomposites are able to increase the survival of cultivated plants under biotic stress due to the modulation of the activity and content of the components of the plant cell antioxidant system.
The obtained results demonstrated a great potential for the use of Se/AG NC in practical agriculture, in particular, for plant growth stimulation and health improvement.
Author Contributions
Conceptualization, A.I.P. and I.A.G.; methodology, O.A.N., A.V.S. and A.R.K.; software, A.V.S.; validation, A.I.P. and O.A.N.; formal analysis, A.I.P.; investigation, I.A.G. and A.I.P.; resources, I.A.G.; data curation, A.I.P., A.V.S. and A.R.K.; writing—original draft preparation, A.I.P. and K.V.K.; writing—review and editing, A.I.P. and K.V.K.; visualization. A.I.P., K.V.K. and A.R.K.; supervision, K.V.K.; project administration, A.I.P.; funding acquisition, I.A.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the federal program No. 0277-2021-0004 (121031300011-7) funded by the Ministry of Science and Higher Education of the Russian Federation within the framework of the basic project “Study of molecular mechanisms of physiological processes and allelopathy in plant-microbial relationships”.
Data Availability Statement
All data are presented in the paper.
Acknowledgments
The authors are grateful to Nikolai Dorofeev for the support of this study and providing soybean seeds.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Füleky, G. Cultivated plants, primarily as food sources. In Cultivated Plants, Primarily as Food Sources; Fuleky, G., Ed.; EOLSS Publishers Co Ltd.: Oxford, UK, 2009; Volume 1, pp. 1–41. [Google Scholar]
- Goffart, J.-P.; Haverkort, A.; Storey, M.; Haase, N.; Martin, M.; Lebrun, P.; Ryckmans, D.; Florins, D.; Demeulemeester, K. Potato production in Northwestern Europe (Germany, France, the Netherlands, United Kingdom, Belgium): Characteristics, issues, challenges and opportunities. Potato Res. 2022, 65, 503–547. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.J.; Suarez, V. The rise of Asia as the centre of global potato production and some implications for industry. Potato J. 2012, 39, 1–22. [Google Scholar]
- Wang, Z.-J.; Liu, H.; Zeng, F.-K.; Yang, Y.-C.; Xu, D.; Zhao, Y.-C.; Liu, X.-F.; Kaur, L.; Liu, G.; Singh, J. Potato processing industry in China: Current scenario, future trends and global impact. Potato Res. 2022, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Devaux, A.; Goffart, J.-P.; Kromann, P.; Andrade-Piedra, J.; Polar, V.; Hareau, G. The potato of the future: Opportunities and challenges in sustainable agri-food systems. Potato Res. 2021, 64, 681–720. [Google Scholar] [CrossRef]
- Sun, J.; Mooney, H.; Wu, W.; Tang, H.; Tong, Y.; Xu, Z.; Huang, B.; Cheng, Y.; Yang, X.; Wei, D.; et al. Importing food damages domestic environment: Evidence from global soybean trade. Proc. Natl. Acad. Sci. USA 2018, 115, 5415–5419. [Google Scholar] [CrossRef]
- Campos, H.; Ortiz, O. (Eds.) The Potato Crop. Its Agricultural, Nutritional and Social Contribution to Humankind; Springer: Cham, Switzerland, 2020; 518p. [Google Scholar] [CrossRef]
- Dilawari, R.; Kaur, N.; Priyadarshi, N.; Prakash, I.; Patra, A.; Mehta, S.; Singh, B.; Jain, P.; Islam, M.A. Soybean: A key player for global food security. In Soybean Improvement; Wani, S.H., Sofi, N.u.R., Bhat, M.A., Lin, F., Eds.; Springer: Cham, Switzerland, 2022; pp. 1–46. [Google Scholar] [CrossRef]
- Paparella, S.; Araújo, S.S.; Rossi, G.; Wijayasinghe, M.; Carbonera, D.; Balestrazzi, A. Seed priming: State of the art and new perspectives. Plant Cell Rep. 2015, 34, 1281–1293. [Google Scholar] [CrossRef]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture—Recent developments and future perspectives. J. Nanobiotechnol. 2022, 20, 254. [Google Scholar] [CrossRef]
- Pagano, A.; Macovei, A.; Xia, X.; Padula, G.; Hołubowicz, R.; Balestrazzi, A. Seed priming applied to onion-like crops: State of the art and open questions. Agronomy 2023, 13, 288. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Zulfiqar, F. Effect of seed priming on horticultural crops. Sci. Hortic. 2021, 286, 110197. [Google Scholar] [CrossRef]
- Liu, X.; Quan, W.; Bartels, D. Stress memory responses and seed priming correlate with drought tolerance in plants: An overview. Planta 2022, 255, 45. [Google Scholar] [CrossRef] [PubMed]
- Singhal, R.K.; Kumar, M.; Bose, B.; Mondal, S.; Srivastava, S.; Dhankher, O.P.; Tripathi, R.D. Heavy metal (loid)s phytotoxicity in crops and its mitigation through seed priming technology. Int. J. Phytoremediation 2023, 25, 187–206. [Google Scholar] [CrossRef] [PubMed]
- Adetunji, A.E.; Adetunji, T.L.; Varghese, B.; Sershen, N.; Pammenter, N.W. Oxidative Stress, Ageing and Methods of Seed Invigoration: An Overview and Perspectives. Agronomy 2021, 11, 2369. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Influence of different seed priming techniques on oxidative and antioxidative responses during the germination of Oryza sativa varieties. Physiol. Mol. Biol. Plants 2020, 26, 551–565. [Google Scholar] [CrossRef] [PubMed]
- Mombeini, M.; Ansari, N.A.; Abdossi, V.; Naseri, A. Effects of chemical seed priming on germination and antioxidant enzymes activity of two cucumber (Cucumis sativus L.) cultivars. Int. J. Hortic. Sci. Technol. 2022, 9, 165–175. [Google Scholar] [CrossRef]
- Szőllősi, R.; Molnár, Á.; Kondak, S.; Kolbert, Z. Dual effect of nanomaterials on germination and seedling growth: Stimulation vs. phytotoxicity. Plants 2020, 9, 1745. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Rizwan, M.; Hassan, Z.U.; Akram, M.A.; Tariq, R.; Brestic, M.; Xie, W. Nanoparticle’s uptake and translocation mechanisms in plants via seed priming. foliar treatment. and root exposure: A review. Environ. Sci. Pollut. Res. Int. 2022, 29, 89823–89833. [Google Scholar] [CrossRef]
- Kandhol, N.; Singh, V.P.; Ramawat, N.; Prasad, R.; Chauhan, D.K.; Sharma, S.; Grillo, R.; Sahi, S.; Peralta-Videa, J.; Tripathi, D.K. Nano-priming: Impression on the beginner of plant life. Plant Stress 2022, 5, 100091. [Google Scholar] [CrossRef]
- do Espirito Santo Pereira, A.; Oliveira, H.C.; Fernandes Fraceto, L.; Santaella, C. Nanotechnology potential in seed priming for sustainable agriculture. Nanomaterials 2021, 11, 267. [Google Scholar] [CrossRef]
- Acharya, P.; Jayaprakasha, G.K.; Crosby, K.M.; Jifon, J.L.; Patil, B.S. Nanoparticle-mediated seed priming improves germination. growth. yield. and quality of watermelons (Citrullus lanatus) at multi-locations in Texas. Sci. Rep. 2020, 10, 5037. [Google Scholar] [CrossRef]
- Farhana; Munis, M.F.H.; Alamer, K.H.; Althobaiti, A.T.; Kamal, A.; Liaquat, F.; Haroon, U.; Ahmed, J.; Chaudhary, H.J.; Attia, H. ZnO nanoparticle-mediated seed priming induces biochemical and antioxidant changes in chickpea to alleviate Fusarium Wilt. J. Fungi 2022, 8, 753. [Google Scholar] [CrossRef] [PubMed]
- Salam, A.; Khan, A.R.; Liu, L.; Yang, S.; Azhar, W.; Ulhassan, Z.; Zeeshan, M.; Wu, J.; Fan, X.; Gan, Y. Seed priming with zinc oxide nanoparticles downplayed ultrastructural damage and improved photosynthetic apparatus in maize under cobalt stress. J. Hazard Mater. 2022, 423 Pt A, 127021. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, X.; Zhang, Z.; Chen, K.; Wang, L.; Chen, H.; Yang, Z.; Li, C.; Zhao, L. AgNPs seed priming accelerated germination speed and altered nutritional profile of Chinese cabbage. Sci. Total Environ. 2022, 808, 151896. [Google Scholar] [CrossRef] [PubMed]
- Venzhik, Y.; Deryabin, A.; Popov, V.; Dykman, L.; Moshkov, I. Priming with gold nanoparticles leads to changes in the photosynthetic apparatus and improves the cold tolerance of wheat. Plant Physiol. Biochem. 2022, 190, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Perfileva, A.I.; Tsivileva, O.M.; Nozhkina, O.A.; Karepova, M.S.; Graskova, I.A.; Ganenko, T.V.; Sukhov, B.G.; Krutovsky, K.V. Effect of natural polysaccharide matrix-based selenium nanocomposites on Phytophthora cactorum and rhizospheric microorganisms. Nanomaterials 2021, 11, 2274. [Google Scholar] [CrossRef] [PubMed]
- Perfileva, A.I.; Nozhkina, O.A.; Graskova, I.A.; Sidorov, A.V.; Lesnichaya, M.V.; Aleksandrova, G.P.; Dolmaa, G.; Klimenkov, I.V.; Sukhov, B.G. Synthesis of selenium and silver nanobiocomposites and their influence on phytopathogenic bacterium Clavibacter michiganensis subsp. sepedonicus. Russ. Chem. Bull. 2018, 67, 157–163. [Google Scholar] [CrossRef]
- Perfileva, A.I.; Nozhkina, O.A.; Tretyakova, M.S.; Graskova, I.A.; Klimenkov, I.V.; Sudakov, N.P.; Alexandrova, G.P.; Sukhov, B.G. Biological activity and environmental safety of selenium nanoparticles encapsulated in starch macromolecules. Nanotechnol. Russ. 2020, 15, 96–104. [Google Scholar] [CrossRef]
- Nozhkina, O.A.; Perfileva, A.I.; Graskova, I.A.; Nurminsky, V.N.; Klimenkov, I.V.; Dyakova, A.V.; Ganenko, T.V.; Borodina, T.N.; Aleksandrova, G.P.; Sukhov, B.G.; et al. The biological activity of a selenium nanocomposite encapsulated in carrageenan macromolecules with respect to ring rot pathogenesis of potato plants. Nanotechnol. Russ. 2019, 14, 255–262. [Google Scholar] [CrossRef]
- Graskova, I.A.; Perfilyeva, A.I.; Nozhkina, O.A.; Dyakova, A.V.; Nurminsky, V.N.; Klimenkov, I.V.; Sudakov, N.P.; Borodina, T.N.; Aleksandrova, G.P.; Lesnichaya, M.V.; et al. Impact of nano-sized selenium the ring rott pathogus and in vitro potatoes. Chem. Plant Raw Mater. 2019, 3, 345–354, (In Russian with English Abstract). [Google Scholar] [CrossRef]
- Perfileva, A.I.; Nozhkina, O.A.; Ganenko, T.V.; Sukhov, B.G.; Artem’ev, A.V.; Trofimov, B.A.; Krutovsky, K.V. Selenium nanocomposites in natural matrices as potato recovery agent. Int. J. Mol. Sci. 2021, 22, 4576. [Google Scholar] [CrossRef]
- Perfileva, A.I.; Graskova, I.A.; Sukhov, B.G.; Krutovsky, K.V. Effect of selenium nanocomposites based on natural polymer matrices on the biomass and storage of potato tubers in a field experiment. Agronomy 2022, 12, 1281. [Google Scholar] [CrossRef]
- Faithfull, T. Methods in Agricultural Chemical Analysis: A Practical Handbook; CABI Publishing: Oxford, UK, 2002; 304p. [Google Scholar] [CrossRef]
- Sanders, E.R. Aseptic Laboratory Techniques: Plating Methods. J. Vis. Exp. 2012, 63, e3064. [Google Scholar] [CrossRef]
- Sagdic, O.; Aksoy, A.; Ozkan, G. Evaluation of the antibacterial and antioxidant potentials of gilaburu (Viburnum opulus L.) fruit extract. Acta Aliment. 2006, 35, 487–492. [Google Scholar] [CrossRef]
- Shaginyan, I.A.; Danilina, G.A.; Chernukha, M.Y.; Alekseeva, G.V.; Batov, A.B. Biofilm formation by strains of Burkholderia cepacia complex in dependence of their phenotypic and genotypic characteristics. J. Microbiol. Epidemiol. Immunobiol. 2007, 1, 3–9, (In Russian with English Abstract). [Google Scholar]
- Defez, R.; Andreozzi, A.; Bianco, C. Quantification of triphenyl-2h-tetrazoliumchloride reduction activity in bacterial cells. Bio-protocol 2017, 7, e2115. [Google Scholar] [CrossRef] [PubMed]
- Zelentsov, S.V.; Moshnenko, E.V.; Trunova, M.V.; Bubnova, L.A.; Budnikov, E.N.; Lukomets, A.V.; Savichenko, V.G.; Dorofeev, N.V.; Katysheva, N.B.; Pomortsev, A.V. A cold-resistant soybean cultivar of the northern ecotype Sayana. Oil Crops 2021, 1, 95–102, (In Russian with English Abstract). [Google Scholar] [CrossRef]
- Boyarkin, A.N. A quick method for determining the activity of peroxidase. Biochemistry 1951, 16, 352. [Google Scholar]
- Hadwan, M.H.; Abed, H.N. Data supporting the spectrophotometric method for the estimation of catalase activity. Data Brief 2016, 6, 194–199. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Nam, K.; Ahn, D.U. Analytical methods for lipid oxidation and antioxidant capacity in food systems. Antioxidants 2021, 10, 1587. [Google Scholar] [CrossRef]
- Kang, M.; Kim, S.; Lee, J.Y.; Yoon, S.; Kim, S.H.; Ha, J. Inactivation of Pectobacterium carotovorum subsp. carotovorum on Chinese cabbage (Brassica rapa L. subsp. pekinensis) by wash treatments with phenolic compounds. LWT 2018, 93, 229–236. [Google Scholar] [CrossRef]
- Markovic, S.; Milić Komić, S.; Jelušić, A.; Iličić, R.; Bagi, F.; Stanković, S.; Popović, T. First report of Pectobacterium versatile causing blackleg of potato in Serbia. Plant Dis. 2022, 106, 312. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Sun, D.; Zhang, X.; Hu, H.; Kong, L.; Rookes, J.E.; Xie, J.; Cahill, D.M. Stimulation of photosynthesis and enhancement of growth and yield in Arabidopsis thaliana treated with amine-functionalized mesoporous silica nanoparticles. Plant Physiol. Biochem. 2020, 156, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Shebl, A.; Hassan, A.; Salama, D.M.; Abd El-Aziz, M.E.; Abd Elwahed, M.S. Template-free microwave-assisted hydrothermal synthesis of manganese zinc ferrite as a nanofertilizer for squash plant (Cucurbita pepo L). Heliyon 2020, 6, e03596. [Google Scholar] [CrossRef]
- Feng, Y.; Kreslavski, V.D.; Shmarev, A.N.; Ivanov, A.A.; Zharmukhamedov, S.K.; Kosobryukhov, A.; Allakhverdiev, S.I.; Shabala, S. Effects of iron oxide nanoparticles (Fe3O4) on growth. photosynthesis. antioxidant activity and distribution of mineral elements in wheat (Triticum aestivum). Plants 2022, 11, 1894. [Google Scholar] [CrossRef]
- Yuan, J.; Chen, Y.; Li, H.; Lu, J.; Zhao, H.; Liu, M.; Nechitaylo, G.S.; Glushchenko, N.N. New insights into the cellular responses to iron nanoparticles in Capsicum annuum. Sci. Rep. 2018, 8, 3228. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Singh, P.M.; Sagar, V.; Pandya, A.; Chinnappa, M.; Kumar, R.; Bahadur, A. Seed priming with ZnO and Fe3O4 nanoparticles alleviate the lead toxicity in Basella alba L. through reduced lead uptake and regulation of ROS. Plants 2022, 11, 2227. [Google Scholar] [CrossRef] [PubMed]
- Ghazal, B.; Saif, S.; Farid, K.; Khan, A.; Rehman, S.; Reshma, A.; Fazal, H.; Ali, M.; Ahmad, A.; Rahman, L.; et al. Stimulation of secondary metabolites by copper and gold nanoparticles in submerge adventitious root cultures of Stevia rebaudiana (Bert.). IET Nanobiotechnol. 2018, 12, 569–573. [Google Scholar] [CrossRef]
- Faizan, M.; Yu, F.; Chen, C.; Faraz, A.; Hayat, S. Zinc oxide nanoparticles help to enhance plant growth and alleviate abiotic stress: A review. Curr. Protein Pept. Sci. 2021, 22, 362–375. [Google Scholar] [CrossRef]
- Mehmood, A. Brief overview of the application of silver nanoparticles to improve growth of crop plants. IET Nanobiotechnol. 2018, 12, 701–705. [Google Scholar] [CrossRef]
- Salem, N.M.; Albanna, L.S.; Awwad, A.M.; Ibrahim, Q.M.; Abdeen, A.O. Green synthesis of nano-sized sulfur and its effect on plant growth. J. Agric. Sci. 2016, 8, 188–194. [Google Scholar] [CrossRef]
- Gudkov, S.V.; Shafeev, G.A.; Glinushkin, A.P.; Shkirin, A.V.; Barmina, E.V.; Rakov, I.I.; Simakin, A.V.; Kislov, A.V.; Astashev, M.E.; Vodeneev, V.A.; et al. Production and use of selenium nanoparticles as fertilizers. ACS Omega 2020, 5, 17767–17774. [Google Scholar] [CrossRef] [PubMed]
- Ghasemian, S.; Masoudian, N.; Nematpour, F.S.; Afshar, A.S. Selenium nanoparticles stimulate growth, physiology, and gene expression to alleviate salt stress in Melissa officinalis. Biologia 2021, 76, 2879–2888. [Google Scholar] [CrossRef]
- Feng, T.; Chen, S.; Gao, D.; Liu, G.; Bai, H.; Li, A.; Peng, L.; Ren, Z. Selenium improves photosynthesis and protects photosystem II in pear (Pyrus bretschneideri), grape (Vitis vinifera), and peach (Prunus persica). Photosynthetica 2015, 53, 609–612. [Google Scholar] [CrossRef]
- Hussein, H.A.; Darwesh, O.M.; Mekki, B.B.; El-Hallouty, S.M. Evaluation of cytotoxicity, biochemical profile and yield components of groundnut plants treated with nano-selenium. Biotechnol. Rep. 2019, 12, e00377. [Google Scholar] [CrossRef]
- Chouhan, D.; Mandal, P. Applications of chitosan and chitosan based metallic nanoparticles in agrosciences—A review. Int. J. Biol. Macromol. 2021, 166, 1554–1569. [Google Scholar] [CrossRef] [PubMed]
- Landa, P. Positive effects of metallic nanoparticles on plants: Overview of involved mechanisms. Plant Physiol. Biochem. 2021, 161, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Hashmi, S.S.; Palma, J.M.; Corpas, F.J. Influence of metallic, metallic oxide, and organic nanoparticles on plant physiology. Chemosphere 2022, 290, 133329. [Google Scholar] [CrossRef] [PubMed]
- Verma, K.K.; Song, X.P.; Joshi, A.; Tian, D.D.; Rajput, V.D.; Singh, M.; Arora, J.; Minkina, T.; Li, Y.R. Recent trends in nano-fertilizers for sustainable agriculture under climate change for global food security. Nanomaterials 2022, 12, 173. [Google Scholar] [CrossRef]
- Lin, D.; Xing, B. Phytotoxicity of nanoparticles: Inhibition of seed germination and root growth. Environ. Pollut. 2007, 150, 243–250. [Google Scholar] [CrossRef]
- Antisari, L.V.; Carbone, S.; Gatti, A.; Vianello, G.; Nannipieri, P. Uptake and translocation of metals and nutrients in tomato grown in soil polluted with metal oxide (CeO2, Fe3O4, SnO2, TiO2) or metallic (Ag, Co, Ni) engineered nanoparticles. Environ. Sci. Pollut. Res. 2015, 22, 1841–1853. [Google Scholar] [CrossRef]
- Buchholz, F.; Junker, R.; Samad, A.; Antonielli, L.; Sarić, N.; Kostić, T.; Sessitsch, A.; Mitter, B. 16S rRNA gene-based microbiome analysis identifies candidate bacterial strains that increase the storage time of potato tubers. Sci. Rep. 2021, 11, 3146. [Google Scholar] [CrossRef]
- Kõiv, V.; Roosaare, M.; Vedler, E.; Ann Kivistik, P.; Toppi, K.; Schryer, D.; Remm, M.; Tenson, T.; Mäe, A. Microbial population dynamics in response to Pectobacterium atrosepticum infection in potato tubers. Sci. Rep. 2015, 5, 11606. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kobayashi, Y.O.; Someya, N.; Ikeda, S. Community analysis of root- and tuber-associated bacteria in field-grown potato plants harboring different resistance levels against common scab. Microbes Environ. 2015, 30, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fu, L.; Chen, C.; Sun, W.; Tian, Y.; Xie, H. Characteristics and rapid diagnosis of Pectobacterium carotovorum ssp. associated with bacterial soft rot of vegetables in China. Plant Dis. 2020, 104, 1158–1166. [Google Scholar] [CrossRef]
- El-Naggar, N.E.-A.; Bashir, S.I.; Rabei, N.H.; Saber, W.E.I.A. Innovative biosynthesis, artificial intelligence-based optimization, and characterization of chitosan nanoparticles by Streptomyces microflavus and their inhibitory potential against Pectobacterium carotovorum. Sci. Rep. 2022, 12, 21851. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Fu, X.; Gao, Y.; Shi, L.; Liu, Q.; Yang, W.; Feng, J. Synthesis, antibacterial evaluation, and safety assessment of CuS NPs against Pectobacterium carotovorum subsp. carotovorum. Pest Manag. Sci. 2022, 78, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Jia, H.; Zhang, H. Green synthesis of silver nanoparticles from Mahonia fortunei extracts and characterization of its inhibitory effect on chinese cabbage soft rot pathogen. Front. Microbiol. 2022, 13, 1030261. [Google Scholar] [CrossRef]
- Coman, V.; Oprea, I.; Leopold, L.F.; Vodnar, D.C.; Coman, C. Soybean interaction with engineered nanomaterials: A literature review of recent data. Nanomaterials 2019, 9, 1248. [Google Scholar] [CrossRef] [PubMed]
- Valgimigli, L.; Baschieri, A.; Amorati, R. Antioxidat activity of nanomaterials. J. Mater. Chem. B 2018, 6, 2036–2051. [Google Scholar] [CrossRef]
- Konopko, A.; Kusio, J.; Litwinienko, G. Antioxidant activity of metal nanoparticles coated with tocopherol-like residues—The importance of studies in homo- and heterogeneous systems. Antioxidants 2020, 9, 5. [Google Scholar] [CrossRef]
- Mahlangeni, N.T.; Moodley, R. Biosynthesis of manganese oxide nanoparticles using Urginea sanguinea and their effects on cytotoxicity and antioxidant activity. Adv. Nat. Sci. Nanosci. Nanotechnol. 2021, 12, 015015. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).