
Investigation of the Effects of the Explant Type and Different Plant Growth Regulators on Micropropagation of Five Mediterranean Salvia spp. Native to Greece


Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Culture Establishment Stage
2.2. Shoot Multiplication Stage
2.2.1. Effect of Explant Type and BA Concentration on the Proliferation of the Five Salvia spp.
2.2.2. Effect of Explant Type, and Cytokinin Type and Concentration on the Proliferation of S. fruticosa and S. officinalis
2.2.3. Explant Number and Data Collection at the Shoot Multiplication Stage
2.3. In Vitro Rooting
2.4. Ex Vitro Acclimatization
2.5. In Vitro Culture Conditions
2.6. Statistical Analysis
3. Results
3.1. Establishment Stage
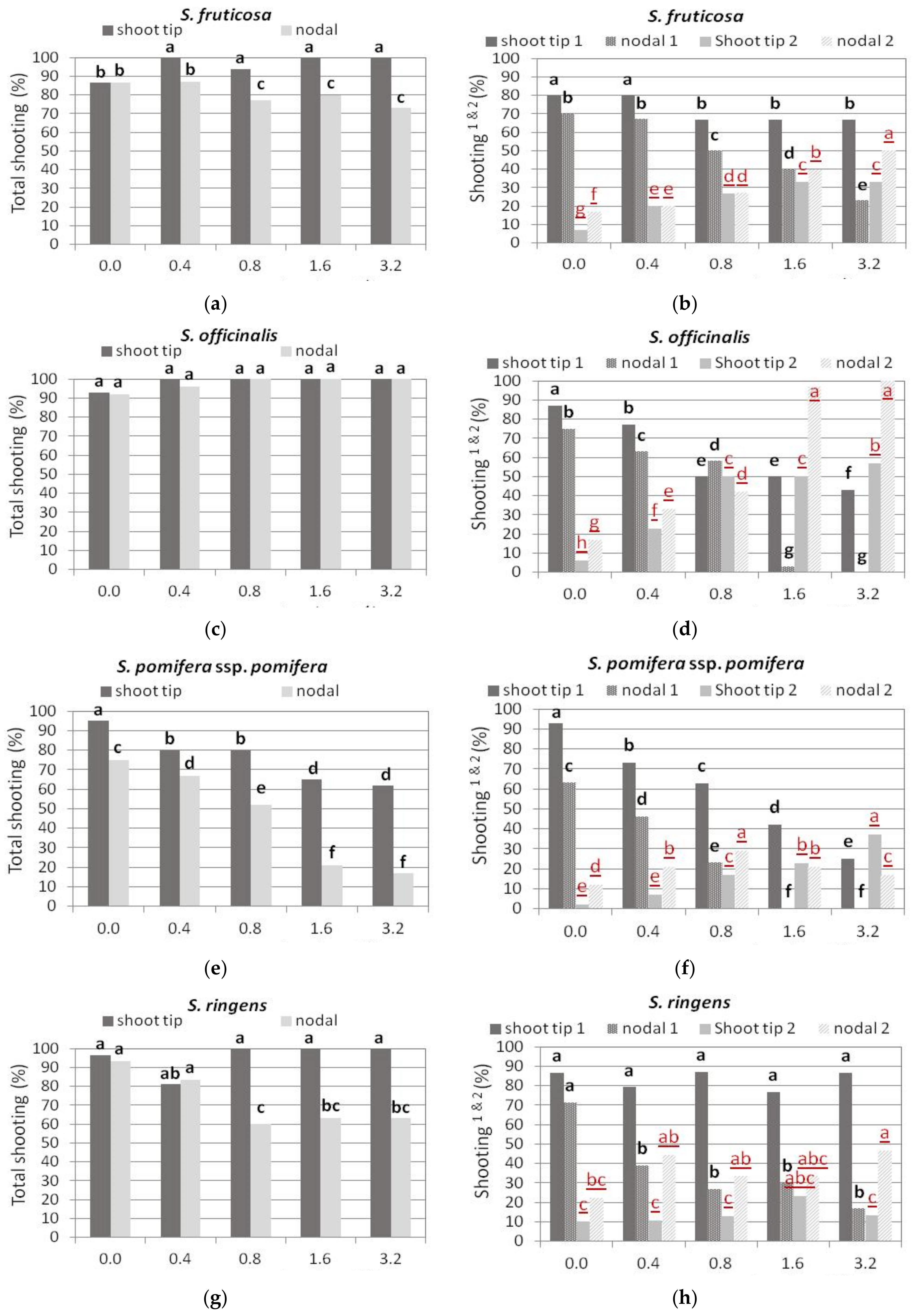
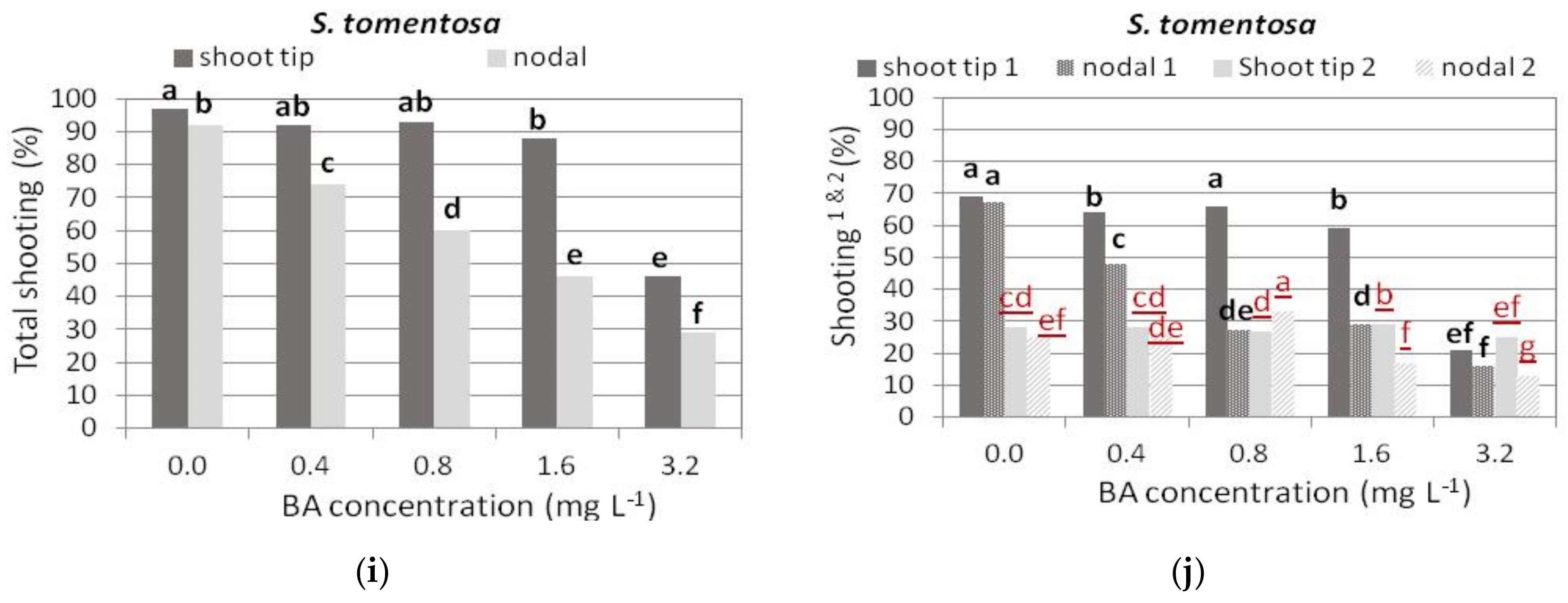
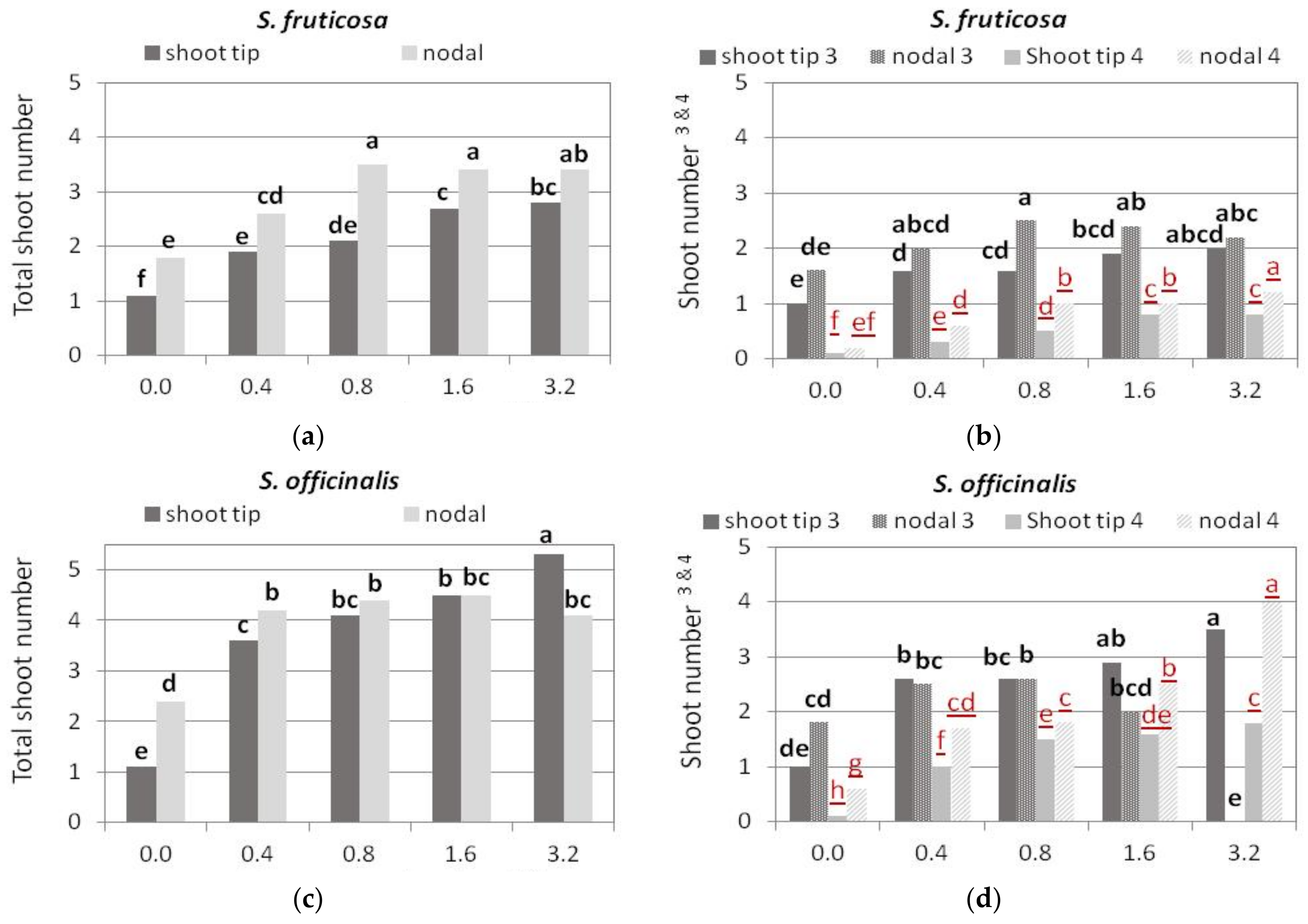
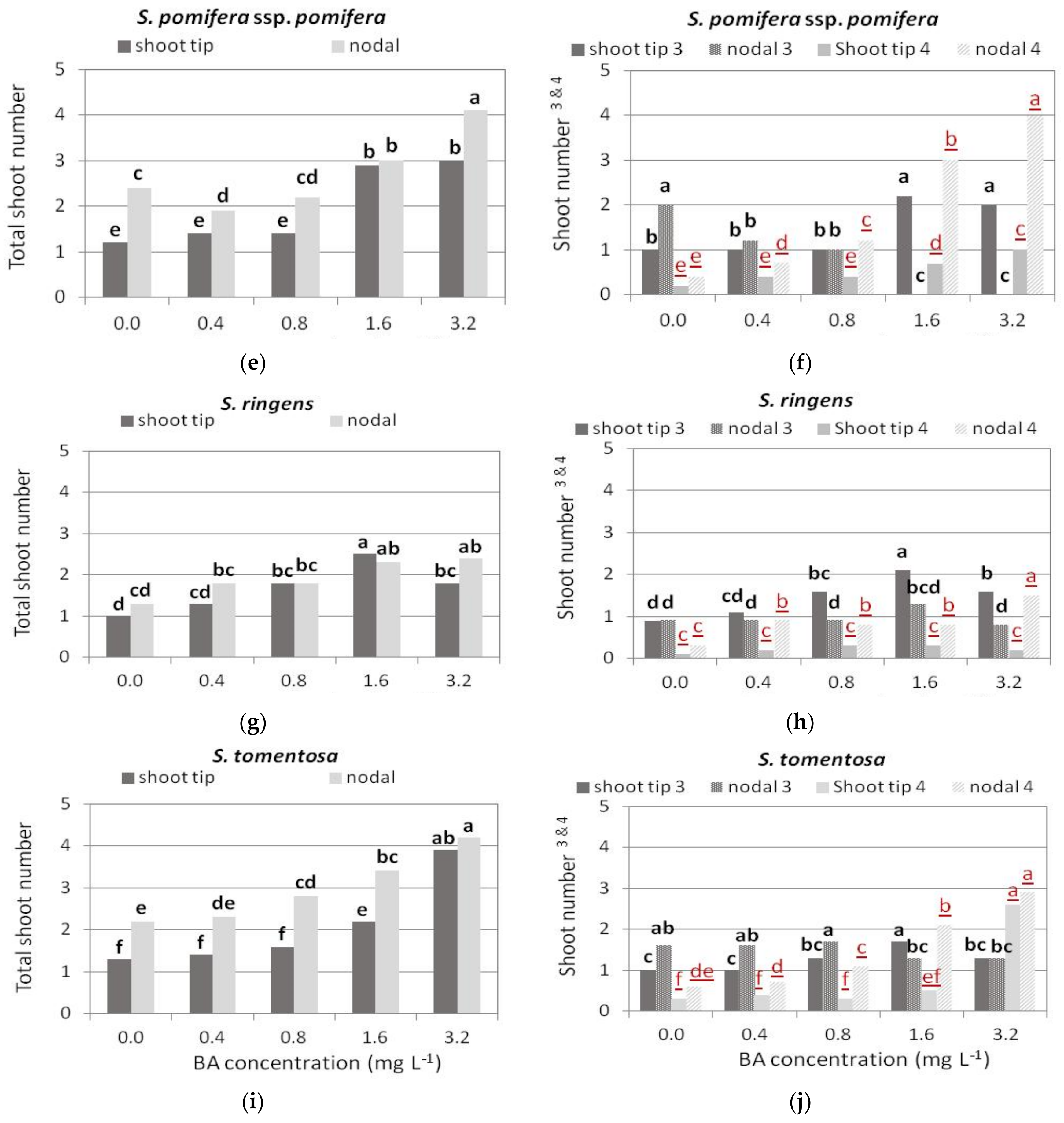
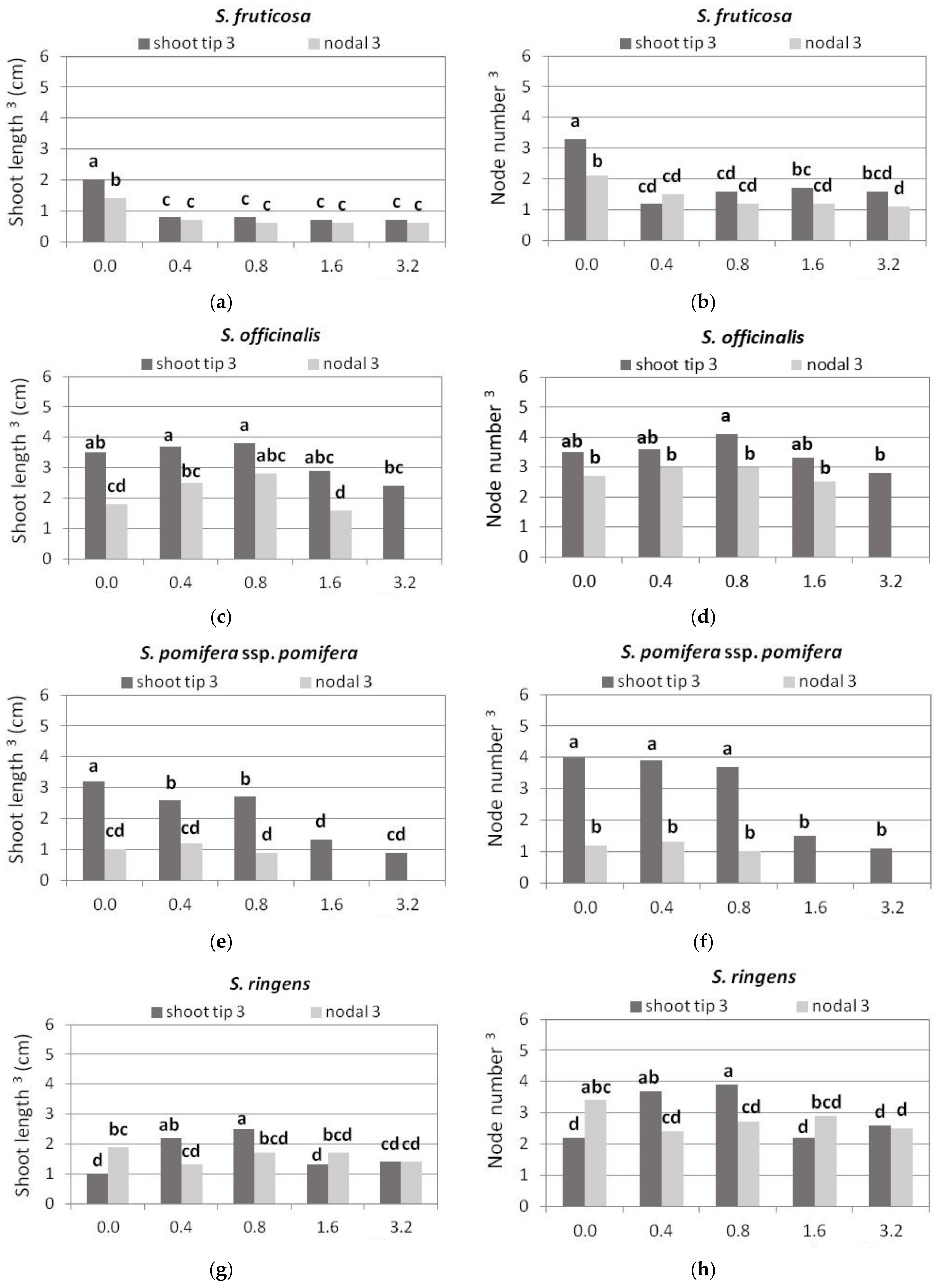
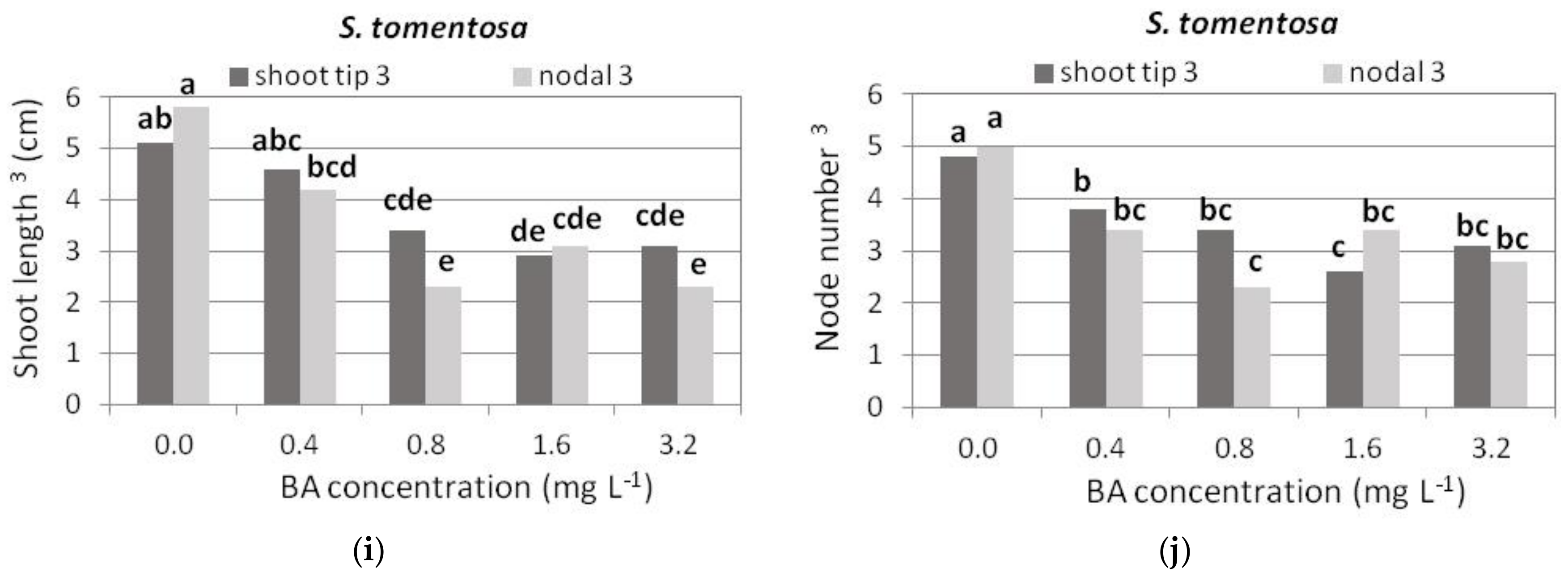
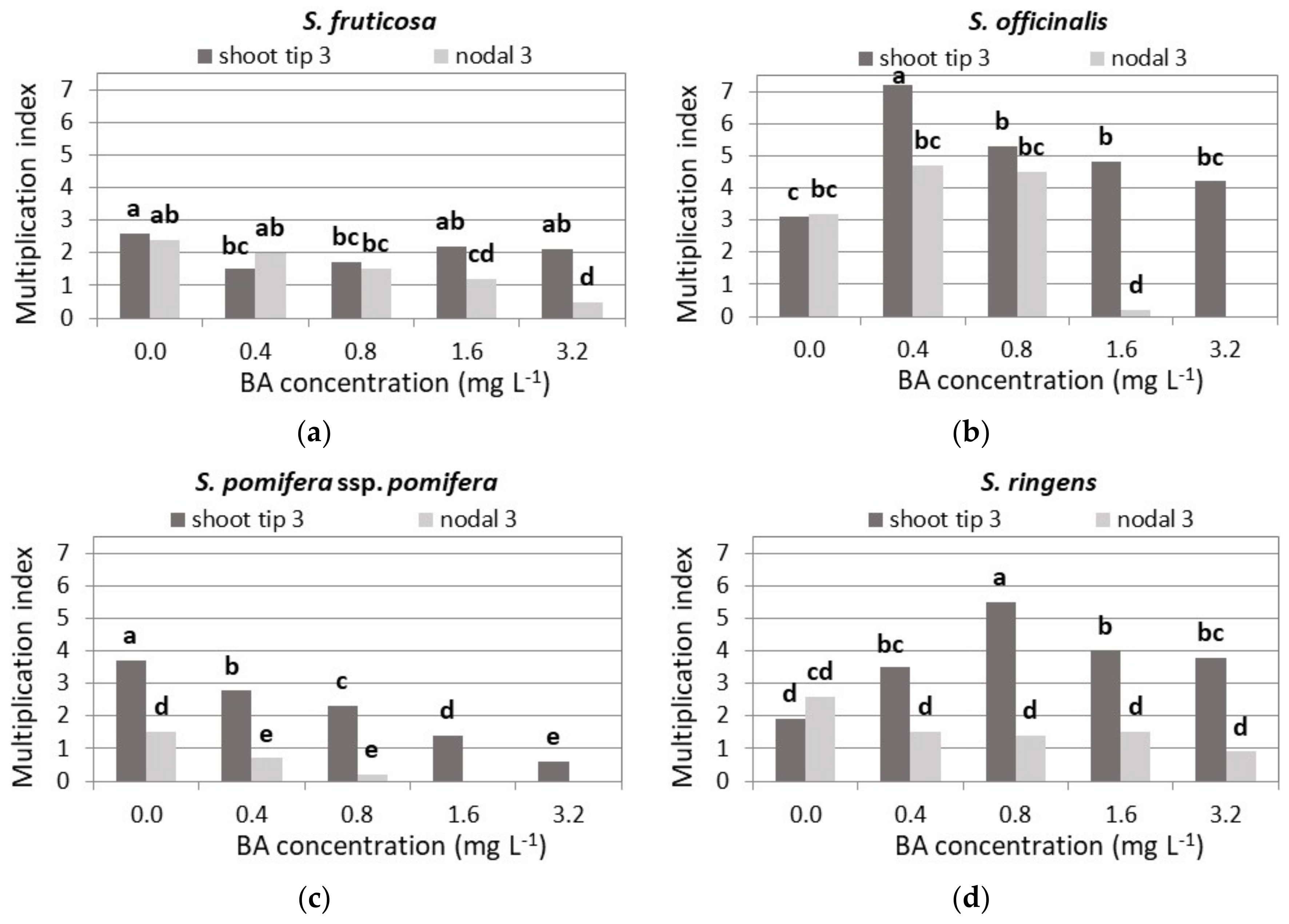
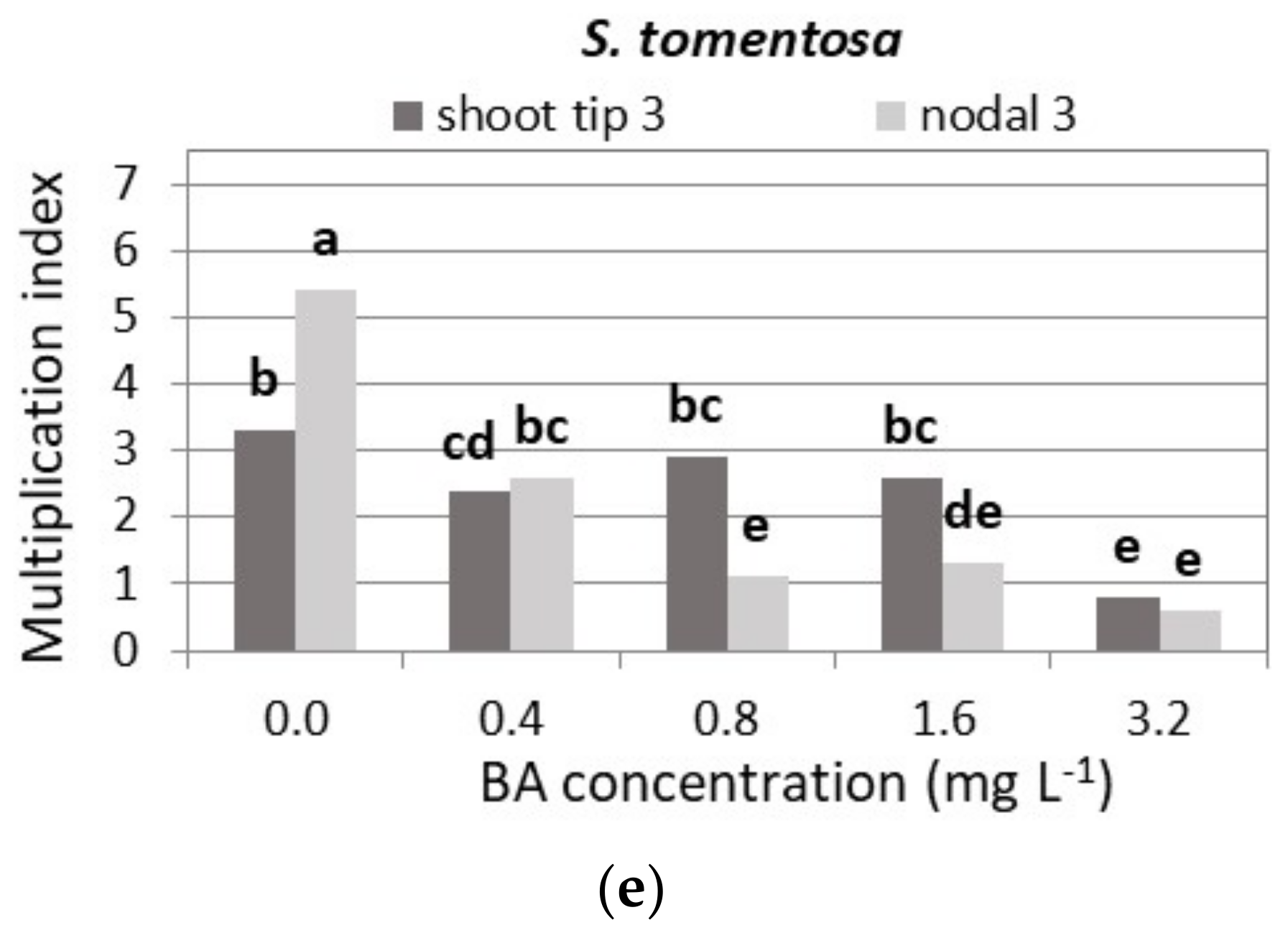
3.2. Shoot Multiplication on BA Media
3.2.1. S. fruticosa
3.2.2. S. officinalis
3.2.3. S. pomifera ssp. pomifera
3.2.4. S. ringens
3.2.5. S. tomentosa
3.3. Effects of Cytokinin Type and Concentration on the Multiplication of S. fruticosa and S. officinalis
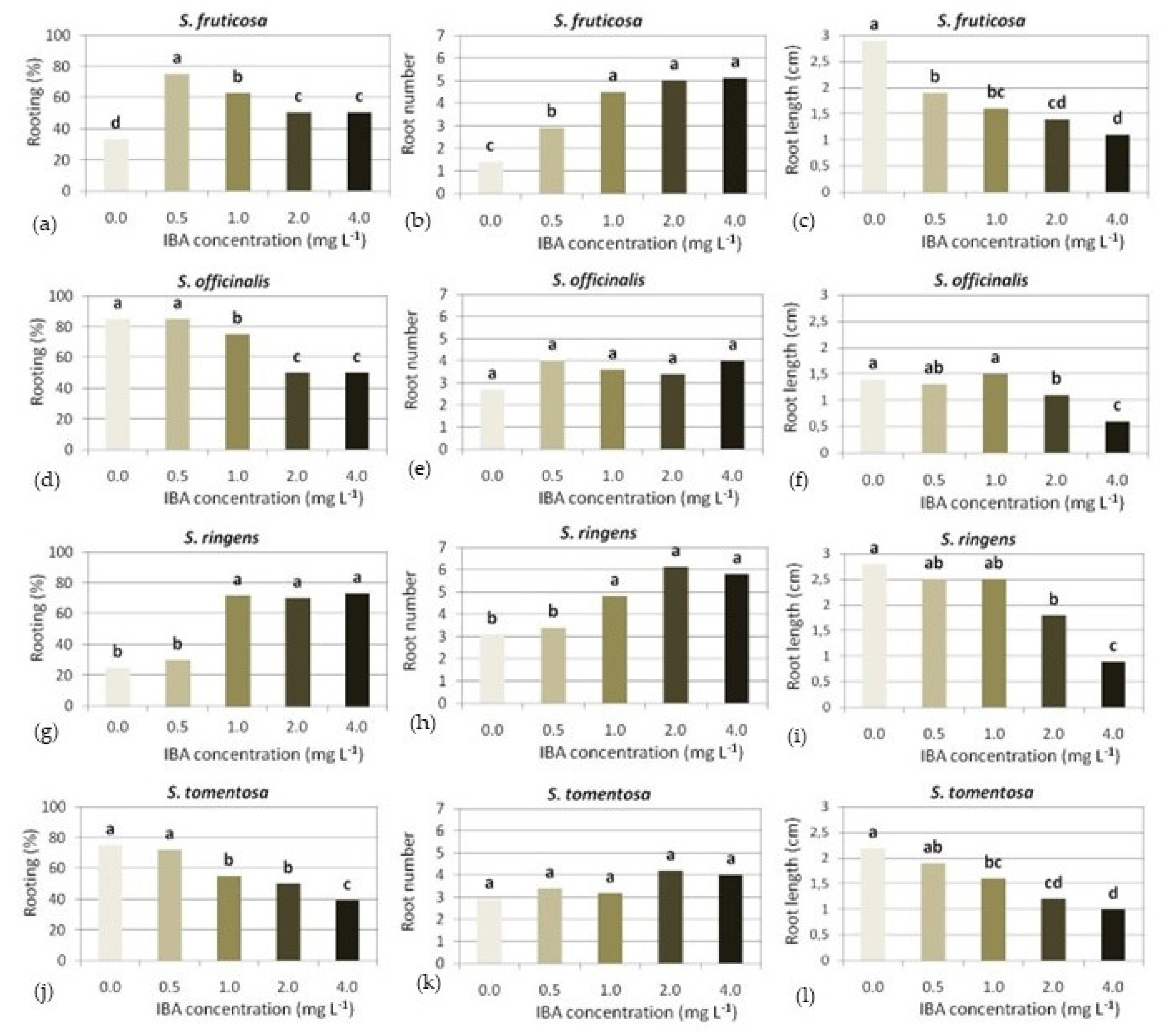
3.4. In Vitro Rooting
3.4.1. S. fruticosa
3.4.2. S. officinalis
3.4.3. S. ringens
3.4.4. S. tomentosa
3.4.5. S. pomifera ssp. pomifera
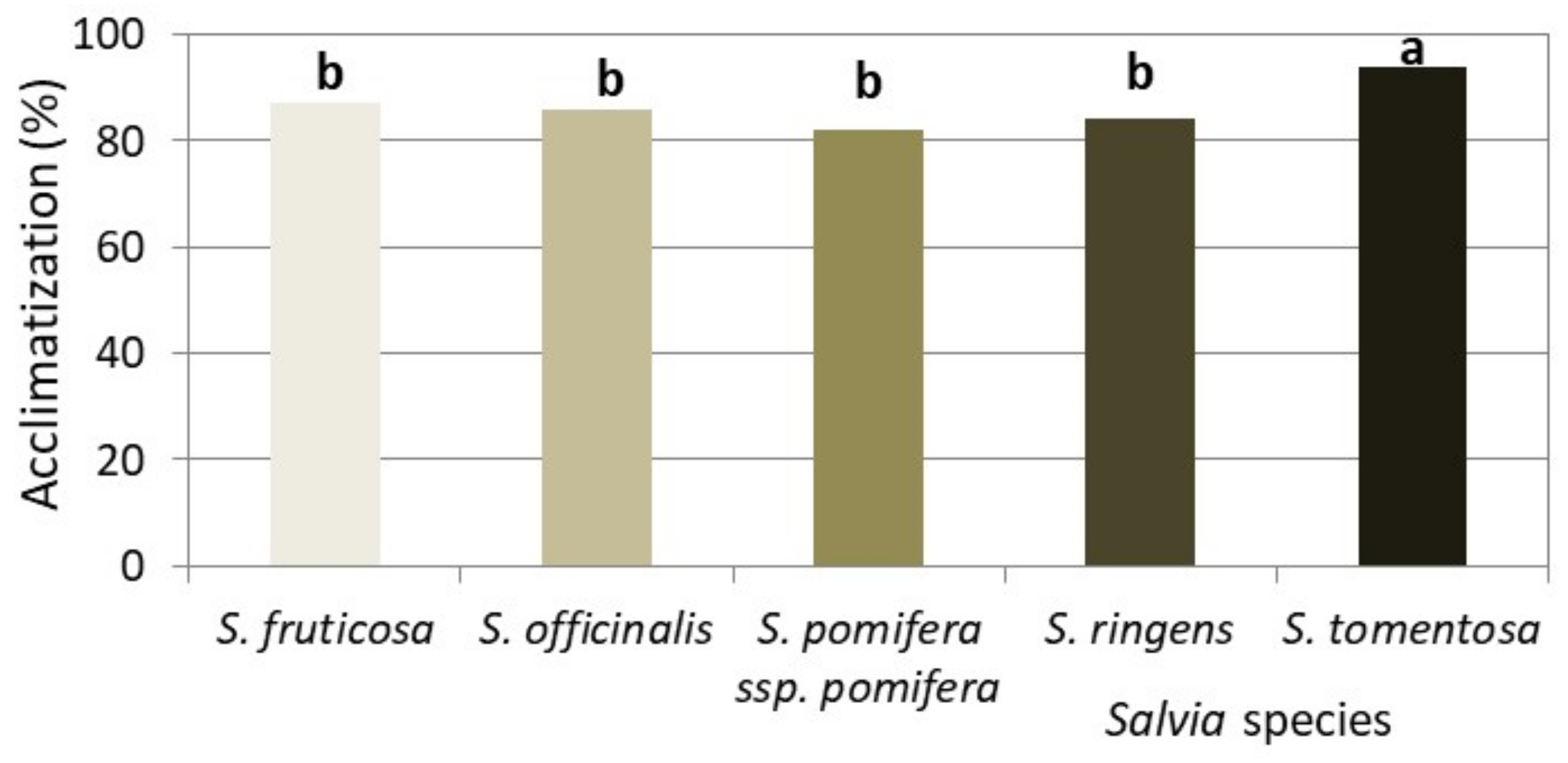
3.5. Ex Vitro Acclimatization
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- The Plant List. Version 1.1. 2013. Available online: http://www.theplantlist.org/ (accessed on 29 April 2022).
- Walker, J.B.; Sytsma, K.J.; Treutlein, J.; Wink, M. Salvia (Lamiaceae) is not monophyletic: Implications for the systematics, radiation, and ecological specializations of Salvia and tribe Mentheae. Am. J. Bot. 2004, 91, 1115–1125. [Google Scholar] [CrossRef] [PubMed]
- Bentham, G. Labiatae. In Genera Plantarum; Bentham, G., Hooker, J.D., Eds.; Reeve and Co.: London, UK, 1876; Volume 2, pp. 1160–1196. [Google Scholar]
- Online Etymology Dictionary. Available online: https://www.etymonline.com/word/salvia (accessed on 11 December 2021).
- Hamidpour, M.; Hamidpour, R.; Hamidpour, S.; Shahlari, M. Chemistry, Pharmacology, and Medicinal Property of Sage (Salvia) to Prevent and Cure Illnesses such as Obesity, Diabetes, Depression, Dementia, Lupus, Autism, Heart Disease, and Cancer. J. Tradit. Complement. Med. 2014, 4, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Bonesi, M.; Loizzo, M.R.; Acquaviva, R.; Malfa, G.A.; Aiello, F.; Tundis, R. Anti-inflammatory and antioxidant agents from Salvia genus (Lamiaceae): An assessment of the current state of knowledge. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2017, 16, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Lopresti, A.L. Salvia (Sage): A review of its potential cognitive-enhancing and protective effects. Drugs R&D 2017, 17, 53–64. [Google Scholar] [CrossRef]
- Hao, D.C.; Ge, G.B.; Xiao, P.G. Anticancer drug targets of Salvia phytometabolites: Chemistry, biology and omics. Curr. Drug Targets 2018, 19, 1–20. [Google Scholar] [CrossRef]
- Ververis, A.; Savvidou, G.; Ioannou, K.; Nicolaou, P.; Christodoulou, K.; Plioukas, M. Greek sage exhibits neuroprotective activity against amyloid beta-induced toxicity. Evid. Based Complement. Altern. Med. 2020, 2020, 2975284. [Google Scholar] [CrossRef]
- Afonso, A.F.; Pereira, O.R.; Cardoso, S.M. Salvia species as nutraceuticals: Focus on antioxidant, antidiabetic and anti-obesity properties. Appl. Sci. 2021, 11, 9365. [Google Scholar] [CrossRef]
- Mervić, M.; Štefan, M.B.; Kindl, M.; Blažeković, B.; Marijan, M.; Vladimir-Knežević, S. Comparative Antioxidant, Anti-Acetylcholinesterase and Anti-α-Glucosidase Activities of Mediterranean Salvia Species. Plants 2022, 11, 625. [Google Scholar] [CrossRef]
- Karalija, E.; Dahija, S.; Tarkowski, P.; Ćavar Zeljkovic, S. Influence of Climate-Related Environmental Stresses on Economically Important Essential Oils of Mediterranean Salvia sp. Front. Plant Sci. 2022, 13, 864807. [Google Scholar] [CrossRef]
- Jiang, S. Therapeutic landscapes and healing gardens: A review of Chinese literature in relation to the studies in western countries. Front. Archit. Res. 2014, 3, 141–153. [Google Scholar] [CrossRef]
- Savi, T.; Andri, S.; Nardini, A. Impact of different green roof layering on plant water status and drought survival. Ecol. Eng. 2013, 57, 188–196. [Google Scholar] [CrossRef]
- Raimondo, F.; Trifilò, P.; Lo Gullo, M.A.; Andri, S.; Savi, T.; Nardini, A. Plant performance on Mediterranean green roofs: Interaction of species-specific hydraulic strategies and substrate water relations. AoB Plants 2015, 7, plv007. [Google Scholar] [CrossRef]
- Martini, A.N.; Tassoula, L.; Papafotiou, M. Adaptation of Salvia fruticosa, S. officinalis, S. ringens and interspecific hybrids in an extensive green roof under two irrigation frequencies. Notulae 2022, 50, 12767. [Google Scholar] [CrossRef]
- Papafotiou, M.; Martini, A.N.; Tassoula, L.; Stylias, E.G.; Kalantzis, A.; Dariotis, E. Acclimatization of Mediterranean native sages (Salvia spp.) and interspecific hybrids in an urban green roof under regular and reduced irrigation. Sustainability 2022, 14, 4978. [Google Scholar] [CrossRef]
- Rahimi, E.; Barghjelveh, S.; Dong, P. A review of diversity of bees, the attractiveness of host plants and the effects of landscape variables on bees in urban gardens. Agric. Food Secur. 2022, 11, 6. [Google Scholar] [CrossRef]
- Dimopoulos, P.; Raus, T.; Bergmeier, E.; Constantinidis, T.; Iatrou, G.; Kokkini, S.; Strid, S.; Tzanoudakis, D. Vascular Plants of Greece: An Annotated Checklist; Botanischer Garten und Botanisches Museum Berlin-Dahlem; Hellenic Botanical Society: Berlin, Germany, 2013. [Google Scholar]
- Chatzopoulou, F.M.; Makris, A.M.; Argiriou, A.; Degenhardt, J.; Kanellis, A.K. EST analysis and annotation of transcripts derived from a trichome-specific cDNA library from Salvia fruticosa. Plant Cell Rep. 2010, 29, 523–534. [Google Scholar] [CrossRef]
- Ghorbani, A.; Esmaeilizadeh, M. Pharmacological properties of Salvia officinalis and its components. J. Tradit. Complement Med. 2017, 7, 433–440. [Google Scholar] [CrossRef]
- Kemp, S.; Hadley, P.; Blanuša, T. The influence of plant type on green roof rainfall retention. Urban Ecosyst. 2019, 22, 355–366. [Google Scholar] [CrossRef]
- Göçer, H.; Yetişir, H.; Ulaş, A.; Arslan, M.; Aydin, A. Plant growth, ion accumulation and essential oil content of Salvia officinalis Mill. and S. tomentosa L. grown under different salt stress. KSU J. Agric. Nat. 2021, 24, 505–514. [Google Scholar]
- Papafotiou, M.; Martini, A.N.; Papanikolaou, E.; Stylias, E.G.; Kalantzis, A. Hybrids Development between Greek Salvia Species and Their Drought Resistance Evaluation along with Salvia fruticosa, under Attapulgite-Amended Substrate. Agronomy 2021, 11, 2401. [Google Scholar] [CrossRef]
- Putievsky, E.; Ravid, U.; Diwan-Rinzler, N.; Zohary, D. Genetic affinities and essential oil composition of Salvia officinalis L., S. fruticosa Mill., S. tomentosa Mill. and their hybrids. Flavour Fragr. J. 1990, 5, 121–123. [Google Scholar] [CrossRef]
- Karousou, R.; Vokou, D.; Kokkini, S. Distribution and essential oils of Salvia pomifera subsp. pomifera (Labiatae) on the island of Crete (S Greece). Biochem. Syst. Ecol. 1998, 26, 889–897. [Google Scholar] [CrossRef]
- Haznedaroglu, M.; Karabay, N.; Zeybek, U. Antibacterial activity of Salvia tomentosa essential oil. Fitoterapia 2001, 72, 829–831. [Google Scholar] [CrossRef] [PubMed]
- Tepe, B.; Daferera, D.; Sokmen, A.; Sokmen, M.; Polissiou, M. Antimicrobial and antioxidant activities of the essential oil and various extracts of Salvia tomentosa Miller (Lamiaceae). Food Chem. 2005, 90, 333–340. [Google Scholar] [CrossRef]
- Arslan, M. Cultivation potential of Salvia tomentosa and S. aramiensis under the Eastern Mediterranean conditions. Sci. Pap. Ser. A Agron. 2016, 59, 174–177. [Google Scholar]
- Hanlidou, E.; Karousou, R.; Lazari, D. Essential-oil diversity of Salvia tomentosa Mill. in Greece. Chem. Biodivers. 2014, 11, 1205–1215. [Google Scholar] [CrossRef]
- Bardakci, H.; Servi, H.; Polatoglu, K. Essential oil composition of Salvia candidissima Vahl. occidentalis Hedge, S. tomentosa Miller and S. heldreichiana Boiss. Ex Bentham from Turkey. J. Essent. Oil-Bear. Plants 2019, 22, 1467–1480. [Google Scholar] [CrossRef]
- Özcan, M.M.; Figueredo, G.; Chalchat, J.C.; Chalard, P.; Tugay, O.; Ceylan, D.A. Chemical constituents of essential oils of Salvia heldreichiana Boiss. Ex Bentham and Salvia tomentosa Mill. J. Agroaliment. Process. Technol. 2019, 25, 106–110. [Google Scholar]
- Grigoriadou, K.; Trikka, F.A.; Tsoktouridis, G.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Maloupa, E.; Makris, A.M. Micropropagation and cultivation of Salvia sclarea for essential oil and sclareol production in northern Greece. In Vitro Cell. Dev. Biol. Plant 2020, 56, 51–59. [Google Scholar] [CrossRef]
- Thanos, C.A.; Doussi, M.A. Ecophysiology of seed germination in endemic labiates of Crete. Isr. J. Plant Sci. 1995, 43, 227–237. [Google Scholar] [CrossRef]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea, Volume 3 Diapenstaceae to Myoporaceae; Cambridge University Press: Cambridge, UK, 1972; pp. 188–190. [Google Scholar]
- Guner, A.; Ozhatay, N.; Ekim, T.; Baser, K.H.C. Flora of Turkey and the East Aegean Islands, Supplement II; Edinburg University Press: Edinburg, UK, 2000; Volume 11. [Google Scholar]
- Martini, A.N.; Bertsouklis, K.F.; Vlachou, G.; Dariotis, E.; Papafotiou, M. Comparative evaluation of rooting cuttings of five Mediterranean sage species (Salvia sp.) native to Greece. Acta Hortic. 2020, 1298, 587–592. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. Seed germination, micropropagation from adult and juvenile origin explants and address of hyperhydricity of the Cretan endemic herb Calamintha cretica. Not. Bot. Horti. Agrobot. 2020, 48, 1504–1518. [Google Scholar] [CrossRef]
- Kanellou, E.; Vlachou, G.; Martini, A.N.; Bertsouklis, K.F.; Papafotiou, M. Seed germination of five sage species (Salvia sp.) of populations native to Greece. Acta Hortic. 2022, 1345, 439–444. [Google Scholar] [CrossRef]
- Jaafar, A.; Mohsen, E.; Hossein, A.R.; Ali, A.J.; Mostafa, E.; Yaser, S.M.; Mohammad, A.S.B.G. Seed germination as the major conservation issue of endemic Iranian salvia species. J. Med. Plants Res. 2012, 6, 37–46. [Google Scholar] [CrossRef]
- Jaafar, A.; Seyed, H.T.; Mohsen, E.; Hossein, A.R.; Ahamad, R.D.T.; Mehdi, M.; Mostafa, E.; Amir, H.D.; Asma, B. Effect of drought stress on germination and seedling growth of Salvia species. Afr. J. Agric. Res. 2012, 7, 5719–5725. [Google Scholar] [CrossRef]
- Kintzios, S.; Nikolau, A.; Skoula, M. Somatic embryogenesis and in vitro rosmarinic acid accumulation in Salvia officinalis and S. fruticosa leaf callus cultures. Plant Cell Rep. 1999, 18, 462–466. [Google Scholar] [CrossRef]
- Santos-Gomes, P.C.; Seabra, R.M.; Andrade, P.B.; Ferreira, M.F. Phenolic antioxidant compounds produced by in vitro shoots of sage (Salvia officinalis L.). Plant Sci. 2002, 162, 981–987. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Bilichowski, I.; Mikiciuk-Olasik, E.; Wysokinska, H. The effect of triacontanol on shoot multiplication and production of antioxidant compounds in shoot cultures of Salvia officinalis L. Acta Soc. Bot. Pol. 2006, 75, 11–15. [Google Scholar] [CrossRef][Green Version]
- Avato, P.; Fortunato, I.M.; Ruta, C.; D’ Elia, R. Glandular hairs and essential oils in micropropagated plants of Salvia officinalis L. Plant Sci. 2005, 169, 29–36. [Google Scholar] [CrossRef]
- Tawfik, A.A.; Mohamed, F.M. Regeneration of salvia (Salvia officinalis L.) via induction of meristematic callus. In Vitro Cell. Dev. Biol. Plant 2007, 43, 21–27. [Google Scholar] [CrossRef]
- Gostin, I. Effects of different plant hormones on Salvia officinalis cultivated in vitro. Int. J. Bot. 2008, 4, 430–436. [Google Scholar]
- Grzegorczyk, I.; Wysokinska, H. Liquid shoot culture of Salvia officinalis L. for micropropagation and production of antioxidant compounds; effect of triacontanol. Acta Soc. Bot. Pol. 2008, 77, 99–104. [Google Scholar] [CrossRef]
- Cristea, O.T.; Prisecaru, M.; Ambarus, S.; Calin, M.; Brezeanu, C.; Brezeanu, M.; Florin, G. Vegetative multiplication of Salvia officinalis L. for the obtaining of true-to-type plants. Biologie 2014, 23, 104–107. [Google Scholar]
- Petrova, M.; Nikolova, M.; Dimitrova, L.; Zayova, E. Micropropagation and evaluation of flavonoid content and antioxidant activity of Salvia officinalis L. Genet. Plant Physiol. 2015, 5, 48–60. [Google Scholar]
- Mohamed, M.A.-H.; Aly, M.K.; Ahmed, E.T.; Abd El-latif, S.A.H. Effect of plant growth regulators on organogenesis of Salvia officinalis L. plants. Minia J. Agric. Res. Develop. 2019, 3, 401–414. [Google Scholar] [CrossRef]
- Arikat, N.A.; Jawad, F.M.; Karam, N.S.; Shibli, R.A. Micropropagation and accumulation of essential oils in wild sage (Salvia fruticosa Mill.). Sci. Hortic. 2004, 100, 193–202. [Google Scholar] [CrossRef]
- Martini, A.N.; Vlachou, G.; Papafotiou, M. Effect of Explant Origin and Medium Plant Growth Regulators on In Vitro Shoot Proliferation and Rooting of Salvia tomentosa, a Native Sage of the Northeastern Mediterranean Basin. Agronomy 2022, 12, 1889. [Google Scholar] [CrossRef]
- Kitto, S.L. Commercial micropropagation. HortScience 1997, 32, 1012–1014. [Google Scholar] [CrossRef]
- Sidhu, Y. In vitro micropropagation of medicinal plants by tissue culture. Plymouth Stud. Sci. 2011, 4, 432–449. [Google Scholar]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In vitro propagation of medicinal and aromatic plants: The case of selected Greek species with conservation priority. In Vitro Cell. Dev. Biol. Plant 2019, 55, 635–646. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Grzegorczyk, I.; Wysokinska, H. Micropropagation of Salvia officinalis L. by shoot tips. Biotechnologia 2004, 2, 212–218. [Google Scholar]
- CachiŢĂ-Cosma, D. The effect of the nature and origin of explants on micropropagation. In High-Tech and Micropropagation I. Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1991; Volume 17. [Google Scholar]
- Papafotiou, M.; Martini, A. Effect of position and orientation of leaflet explants with respect to plant growth regulators on micropropagation of Zamioculcas zamiifolia Engl. (ZZ). Sci. Hortic. 2009, 120, 115–120. [Google Scholar] [CrossRef]
- Kartsonas, E.; Papafotiou, M. Mother plant age and seasonal influence on in vitro propagation of Quercus euboica Pap., an endemic, rare and endangered oak species of Greece. Plant Cell Tissue Organ Cult. (PCTOC) 2007, 90, 111–116. [Google Scholar] [CrossRef]
- Martini, A.N.; Papafotiou, M.; Vemmos, S.N. Season and explant origin affect phenolic content, browning of explants, and micropropagation of ×Malosorbus florentina (Zucc.) Browicz. HortScience 2013, 48, 102–107. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. In Vitro Propagation of Calamintha nepeta. Acta Hort. 2016, 1113, 189–194. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. In Vitro Propagation of Ballota acetabulosa. Acta Hort. 2016, 1113, 171–174. [Google Scholar] [CrossRef]
- Cuenca, S.; Amo-Marco, J.B. In vitro propagation of two Spanish endemic species of Salvia through bud proliferation. In Vitro Cell. Dev. Biol. Plant 2000, 36, 225–229. [Google Scholar] [CrossRef]
- Ghanbar, T.; Hosseini, B.; Jabbarzadeh, Z.; Farokhzad, A.; Sharafi, A. High-frequency in vitro direct shoots regeneration from axillary nodal and shoot tip explants of clary sage (Salvia sclarea L.). Bulg. J. Agric. Sci. 2016, 22, 73–78. [Google Scholar]
- Raja, H.D.; Arockiasamy, D.I. In vitro propagation of Mentha viridis L. from nodal and shoot tip explants. Plant Tissue Cult. Biotechnol. 2008, 18, 1–6. [Google Scholar] [CrossRef]
- Sujana, P.; Naidu, C.V. High frequency rapid plant regeneration from shoot tip and nodal explants of Mentha piperita (L.)—An important multipurpose medicinal plant. J. Phytol. 2011, 3, 9–13. [Google Scholar]
- Moharami, L.; Hosseini, B.; Ghotbi Ravandi, E.; Jafari, M. Effects of plant growth regulators and explant types on in vitro direct plant regeneration of Agastache foeniculum, an important medicinal plant. In Vitro Cell. Dev. Biol. Plant 2014, 50, 707–711. [Google Scholar] [CrossRef]
- Sharma, U.; Mohan, J.S.S. Reduction of vitrification in in vitro raised shoots of Chlorophytum borilivinum (Sant. and Fernand.), a rare potent medicinal herb. Indian J. Exp. Biol. 2006, 44, 499–505. [Google Scholar] [PubMed]
- Islam, A.T.M.R.; Islam, M.M.; Alam, M.F. Rapid in vitro clonal propagation of herbal spice, Mentha piperita L. using shoot tip and nodal explants. Res. Plant Biol. 2017, 5, 43–50. [Google Scholar] [CrossRef]
- Papafotiou, M.; Martini, A.N. In vitro seed and clonal propagation of the Mediterranean aromatic and medicinal plant Teucrium capitatum. HortScience 2016, 51, 403–411. [Google Scholar] [CrossRef]
- Papafotiou, M.; Bertsouklis, K.F.; Trigka, M. Micropropagation of Arbutus unedo, A. andrachne, and their natural hybrid, A. x andrachnoides from seedling explants. J. Hort. Sci. Biot. 2013, 88, 768–775. [Google Scholar] [CrossRef]
- Sharma, S.; Shahzad, A.; Kumar, J.; Anis, M. In vitro propagation and synseed production of scarlet salvia (Salvia splendens). Rend. Fis. Acc. Lincei 2014, 25, 359–368. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K. Studies on seed germination and micropropagation of Clinopodium nepeta: A medicinal and aromatic plant. HortScience 2019, 54, 1558–1564. [Google Scholar] [CrossRef]
- Zayova, E.; Nikolova, M.; Dimitrova, L.; Petrova, M. Comparative study of in vitro, ex vitro and in vivo propagated Salvia hispanica (Chia) plants: Morphometric analysis and antioxidant activity. AgroLife Sci. J. 2016, 5, 166–174. [Google Scholar]
- Vlachou, G.; Martini, A.N.; Dariotis, E.; Papafotiou, M. Comparative evaluation of seed germination of five Mediterranean sage species (Salvia sp.) native to Greece. Acta Hortic. 2020, 1298, 593–598. [Google Scholar] [CrossRef]
- Lai, C.C.; Lin, H.M.; Nalawade, S.M.; Fang, W.; Tsay, H.S. Hyperhydricity in shoot cultures of Scrophularia yoshimurae can be effectively reduced by ventilation of culture vessels. J. Plant Physiol. 2005, 162, 355–361. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, M.; van Staden, J. Effect of ammonium ions and cytokinins on hyperhydricity and multiplication rate of in vitro regenerated shoots of Aloe polyphylla. Plant Cell Tissue Organ Cult. (PCTOC) 2008, 92, 227–231. [Google Scholar] [CrossRef]
- Ivanova, M.; van Staden, J. Influence of gelling agent and cytokinins on the control of hyperhydricity in Aloe polyphylla. Plant Cell Tissue Organ Cult. (PCTOC) 2011, 104, 13–21. [Google Scholar] [CrossRef]
- Badr-Elden, A.M.; Nower, A.A.; Ibrahim, I.A.; Ebrahim, M.K.; Abd Elaziem, T.M. Minimizing the hyperhydricity associated with in vitro growth and development of watermelon by modifying the culture conditions. Afr. J. Biotechnol. 2012, 11, 8705–8717. [Google Scholar] [CrossRef]
- Pence, V.C.; Finke, L.R.; Niedz, R.P. Evaluating a DOE screen to reduce hyperhydricity in the threatened plant, Cycladenia humilis var. jonesii. In Vitro Cell. Dev. Biol. Plant 2020, 56, 215–229. [Google Scholar] [CrossRef]
- Reyes-Vera, I.; Potenza, C.; Barrow, J. Hyperhydricity reversal and clonal propagation of four-wing saltbush (Atriplex canescens, Chenopodiaceae) cultivated in vitro. Aust. J. Bot. 2008, 56, 358–362. [Google Scholar] [CrossRef]
- Wu, H.; Yu, X.; Teixeira da Silva, J.A.; Lu, G. Direct shoot induction of Paeonia lactiflora ‘Zhong Sheng Fen’and rejuvenation of hyperhydric shoots. N. Zeal. J. Crop Hortic. Sci. 2011, 39, 271–278. [Google Scholar] [CrossRef]
- Bertsouklis, K.; Papafotiou, M.; Balotis, G. Effect of medium on in vitro growth and ex vitro establishment of Globularia alypum L. Acta Hortic. 2003, 616, 177–180. [Google Scholar] [CrossRef]
- Papafotiou, M.; Kalantzis, A. Studies on in vitro propagation of Lithodora zahnii. Acta Hortic. 2009, 813, 465–470. [Google Scholar] [CrossRef]
- Vlachou, G.; Papafotiou, M.; Bertsouklis, K.F. Studies on in vitro propagation of Anthyllis barba-jovis L. Acta Hortic. 2017, 1155, 317–320. [Google Scholar] [CrossRef]
- Trigka, M.; Papafotiou, M. In vitro propagation of Anthyllis barba-jovis from seedling tissues. Acta Hortic. 2017, 1189, 473–748. [Google Scholar] [CrossRef]
- Kataeva, N.V.; Alexandrova, I.G.; Butenko, R.G.; Dragavtceva, E.V. Effect of applied and internal hormones on vitrification and apical necrosis of different plants cultured in vitro. Plant Cell Tissue Organ Cult. (PCTOC) 1991, 27, 149–154. [Google Scholar] [CrossRef]
- Ravanfar, S.A.; Salim, S.; Aziz, M.A.; Abdullah, S.N.A.; Rashid, A.A. Influence of phenyl-urea and adenine-type cytokinins on direct adventitious shoot regeneration of cabbage (Brassica oleracea subsp. capitata) “KCross”. Biotechnology 2014, 31, 1–6. [Google Scholar] [CrossRef][Green Version]
- Liu, M.; Jiang, F.; Kong, X.; Tian, J.; Wu, Z.; Wu, Z. Effects of multiple factors on hyperhydricity of Allium sativum L. Sci. Hortic. 2017, 217, 285–296. [Google Scholar] [CrossRef]
- Murashige, T. Principles of rapid propagation. In Propagation of Higher Plants through Tissue Culture: A Bridge between Research and Application; Hughes, K.W., Henke, R., Constantin, M., Eds.; Tech. Information Center, U.S. Dept. of Energy: Oak Ridge, TN, USA, 1979; pp. 14–24. [Google Scholar]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Salvia sp. | Explant Type | Shoot-Producing Explants 1/2 (%) | Mean Number of NSh ₸/HSh ₸₸ | Mean Shoot Length (cm) | Mean NSh Node Number | Multi-Plication Index ₸ |
|---|---|---|---|---|---|---|
| S. fruticosa (n = 18) | Shoot tip | 67.0 c z/17 de | 1.2 bc/0.2 cd | 1.6 cd | 3.2 bc | 2.6 c |
| Single node | 56.0 d/22 cd | 1.9 a/0.7 b | 0.8 d | 1.1 d | 1.2 de | |
| S. officinalis (n = 30) | Shoot tip | 93.3 a/7 f | 1.1 c/0.1 d | 3.6 a | 4.7 a | 4.8 b |
| Single node | 75.0 b/23 c | 2.1 a/0.8 ab | 3.0 b | 3.7 b | 5.8 a | |
| S. pomifera ssp. pomifera (n = 12) | Shoot tip | 92.0 a/8 f | 1.0 c/0.1 d | 1.6 d | 2.1 cd | 1.9 cd |
| Single node | 46.0 e/50 a | 1.8 a/1.0 a | 1.0 d | 1.6 d | 1.3 cde | |
| S. ringens (n = 40) | Shoot tip | 87.0 a/0 g | 1.0 c/0.0 d | 3.0 ab | 5.2 a | 4.5 b |
| Single node | 27.1 f/18 cd | 1.4 b/0.4 c | 2.4 bc | 4.5 a | 1.6 cde | |
| S. tomentosa (n = 7) | Shoot tip | 26.0 f/10 ef | 1.0 c/0.2 cd | 1.9 bcd | 2.4 cd | 0.6 de |
| Single node | 10.0 g/40 b | 2.0 a/0.9 ab | 1.4 cd | 2.0 cd | 0.4 e | |
| FSalvia sp. | -/- | -/- | *** | *** | - | |
| Fexplant type | -/- | -/- | ** | *** | - | |
| FSalvia sp. × expl. type | ***/*** | **/* | NS | NS | *** | |
| Fone-way ANOVA | ***/*** | ***/*** | *** | *** | *** |
| Three-Way ANOVA | Shoot- Producing Explants 1 | Shoot- Producing Explants 2 | Mean Number of NSh ₸ | Mean NSh Length ₸ | Mean NSh Node Number ₸ | Mean Number of HSh ₸₸ | Multi-Plication Index ₸ |
|---|---|---|---|---|---|---|---|
| 0.4 | 74.5 | 17.6 | 1.5 | 1.4 a z | 2.1 a | 0.4 | 2.1 |
| 0.8 | 66.6 | 23.6 | 1.7 | 1.3 ab | 2.0 a | 0.5 | 2.2 |
| 1.6 | 58.1 | 31.3 | 1.6 | 1.2 b | 1.8 b | 0.8 | 1.6 |
| 3.2 | 44.5 | 41.2 | 1.8 | 1.1 b | 1.7 b | 1.1 | 1.3 |
| BA | 57.6 | 31.3 | 2.0 | 0.7 c | 1.4 d | 0.8 | 1.6 |
| ZEA | 51.2 | 32.8 | 1.7 | 1.1 b | 1.9 bc | 0.9 | 1.5 |
| KIN | 70.9 | 21.0 | 1.4 | 1.7 a | 2.1 ab | 0.5 | 2.0 |
| 2iP | 62.8 | 22.4 | 1.3 | 1.1 b | 1.8 c | 0.4 | 1.4 |
| mT | 62.1 | 34.6 | 1.8 | 1.6 a | 2.3 a | 1.0 | 2.6 |
| Shoot tip | 70.7 | 25.1 | 1.5 | 1.4 a | 2.1 a | 0.5 | 2.1 |
| Single node | 51.2 | 31.8 | 1.8 | 1.1 b | 1.7 b | 0.9 | 1.5 |
| Significance § | |||||||
| FCytokinin type | - | - | - | *** | *** | - | - |
| FCytok. concentration | - | - | - | ** | *** | - | - |
| FExplant type | - | - | - | *** | *** | - | - |
| FCytok. × Concentration | - | - | NS | NS | NS | - | *** |
| FCytok. × Explant type | - | - | ** | NS | NS | - | * |
| FCytok. × Expl. type | - | - | * | NS | NS | - | NS |
| FCytok. × Conc. × Expl. type | *** | *** | NS | NS | NS | *** | NS |
| Three-Way ANOVA | Shoot- Producing Explants 1 | Shoot- Producing Explants 2 | Mean Number of NSh ₸ | Mean NSh Length ₸ | Mean NSh Node Number ₸ | Mean Number of HSh ₸₸ | Multi-Plication Index ₸ |
|---|---|---|---|---|---|---|---|
| 0.4 | 72.6 | 25.8 | 1.9 | 2.8 a z | 2.9 a | 0.6 | 3.8 |
| 0.8 | 58.2 | 41.0 | 2.0 | 2.3 b | 3.1 a | 0.8 | 3.1 |
| 1.6 | 43.2 | 55.9 | 2.0 | 2.1 b | 2.8 a | 1.1 | 2.1 |
| 3.2 | 33.4 | 65.6 | 1.8 | 1.6 c | 2.1 b | 1.9 | 1.8 |
| BA | 43.0 | 56.5 | 2.4 | 2.5 b | 2.8 bc | 2.0 | 3.7 |
| ZEA | 50.0 | 50.0 | 1.7 | 2.8 a | 3.2 a | 0.9 | 2.4 |
| KIN | 70.1 | 27.1 | 1.5 | 2.0 c | 2.3 d | 0.6 | 2.3 |
| 2iP | 65.0 | 33.5 | 1.5 | 1.9 c | 2.9 b | 0.6 | 2.6 |
| mT | 30.5 | 68.3 | 2.5 | 1.8 c | 2.4 cd | 1.4 | 2.4 |
| Shoot tip | 60.0 | 42.1 | 1.9 | 2.6 a | 3.2 a | 0.8 | 3.0 |
| Single node | 46.8 | 52.1 | 1.9 | 1.8 b | 2.3 b | 1.4 | 2.4 |
| Significance § | |||||||
| FCytokinin type | - | - | - | *** | *** | - | - |
| FCytok. concentration | - | - | - | ** | *** | - | - |
| FExplant type | - | - | - | *** | *** | - | - |
| FCytok. × Concentration | - | - | NS | NS | NS | - | *** |
| FCytok. × Explant type | - | - | ** | NS | NS | - | * |
| FCytok. × Expl. type | - | - | * | NS | NS | - | NS |
| FCytok. × Conc. × Expl. type | *** | *** | NS | NS | NS | *** | NS |
| PGRs Concentration (mg L−1) | Shoot- Producing Explants 1/2 (%) | Mean Number of NSh ₸/HSh ₸₸ | Mean NSh Length ₸ (cm) | Mean Node Number ₸ | Multi-Plication Index |
|---|---|---|---|---|---|
| Shoot tip explant | |||||
| 0.0 (Hf ₸₸₸) | 84 b z/8 n | 1.0 j/0.1 n | 2.2 a | 3.3 a | 2.8 cd |
| 0.4 BA | 80 bc/20 kl | 1.6 def/0.3 lmn | 0.8 fgh | 1.2 ij | 1.5 ijk |
| 0.8 BA | 67 fgh/27 jk | 1.6 def/0.5 kl | 0.8 fgh | 1.6 fgh | 1.7 hi |
| 1.6 BA | 67 fgh/33 hij | 1.9 cde/0.8 hij | 0.7 gh | 1.7 efg | 2.2 efg |
| 3.2 BA | 67 fgh/33 hij | 2.0 bcd/0.8 hij | 0.7 gh | 1.6 fgh | 2.1 fgh |
| 0.4 ZEA | 83 b/13 mn | 1.1 ij/0.2 mn | 1.6 bcd | 2.5 bcd | 2.3 efg |
| 0.8 ZEA | 67 fgh/21 kl | 1.4 fgh/0.3 lmn | 1.3 def | 2.5 bcd | 2.3 efg |
| 1.6 ZEA | 59 ij/29 ij | 1.3 ghi/0.6 j | 1.1 ef | 2.3 cde | 1.8 ghi |
| 3.2 ZEA | 21 op/58 b | 2.2 abc/2.3 b | 0.7 gh | 1.2 ij | 0.6 klm |
| 0.4 KIN | 92 a/8 n | 1.0 j/0.2 mn | 2.1 a | 2.5 bcd | 2.3 efg |
| 0.8 KIN | 83 b/17 lm | 1.2 hij/0.3 lmn | 2.0 ab | 2.5 bcd | 2.5 def |
| 1.6 KIN | 71 def/21 kl | 1.3 ghi/0.5 kl | 1.9 abc | 2.0 def | 1.8 ghi |
| 3.2 KIN | 58 ij/30 ij | 1.5 efg/0.8 hij | 1.7 bcd | 2.0 def | 1.7 hi |
| 0.4 2iP | 83 b/17 lm | 1.0 j/0.2 mn | 1.5 bcd | 2.2 cde | 1.8 ghi |
| 0.8 2iP | 75 cd/21 kl | 1.1 ij/0.2 mn | 1.2 def | 1.9 efg | 1.6 ij |
| 1.6 2iP | 75 cd/21 kl | 1.2 hij/0.3 lmn | 1.1 ef | 1.8 efg | 1.6 ij |
| 3.2 2iP | 71 def/29 ij | 1.2 hij/0.3 lmn | 0.8 fgh | 2.1 def | 1.8 ghi |
| 0.4 mT | 73 de/21 kl | 1.7 def/0.6 j | 2.0 ab | 2.9 ab | 3.6 b |
| 0.8 mT | 77 c/23 jkl | 2.2 abc/0.6 j | 1.8 bc | 2.7 bc | 4.6 a |
| 1.6 mT | 65 gh/35 hij | 1.8 cde/0.9 gh | 1.6 bcd | 2.4 bcd | 2.8 cd |
| 3.2 mT | 62 hi/38 fgh | 2.1 abc/1.0 fgh | 1.4 cde | 1.7 efg | 2.2 efg |
| Single node explant | |||||
| 0.0 (Hf ₸₸₸) | 71 def/17 lm | 1.7 def/0.3 lmn | 1.4 cde | 2.3 cde | 2.8 cd |
| 0.4 BA | 67 fgh/20 kl | 2.0 bcd/0.6 j | 0.7 gh | 1.5 ghi | 2.0 fgh |
| 0.8 BA | 50 kl/27 jk | 2.5 a/1.0 fgh | 0.6 h | 1.2 ij | 1.5 ijk |
| 1.6 BA | 40 mn/40 ef | 2.4 ab/1.0 fgh | 0.6 h | 1.2 ij | 1.2 jk |
| 3.2 BA | 23 op/50 c | 2.2 abc/1.2 ef | 0.6 h | 1.1 j | 0.5 lm |
| 0.4 ZEA | 50 kl/29 ij | 2.0 bcd/0.7 ij | 1.2 def | 1.7 efg | 1.7 hi |
| 0.8 ZEA | 46 lm/33 hij | 1.9 cde/1.0 fgh | 1.1 ef | 1.7 efg | 1.5 ijk |
| 1.6 ZEA | 46 lm/37 ghi | 2.0 bcd/1.0 fgh | 1.0 efg | 1.7 efg | 1.6 ij |
| 3.2 ZEA | 13 p/62 a | 2.0 bcd/2.4 a | 0.8 fgh | 1.3 hij | 0.3 m |
| 0.4 KIN | 83 b/13 mn | 1.4 fgh/0.3 lmn | 1.6 bcd | 2.0 def | 2.3 efg |
| 0.8 KIN | 71 def/21 kl | 1.6 def/0.4 kl | 1.5 bcd | 2.0 def | 2.3 efg |
| 1.6 KIN | 63 ghi/25 jkl | 1.6 def/0.7 ij | 1.5 bcd | 2.1 def | 2.1 fgh |
| 3.2 KIN | 46 lm/33 hij | 1.8 cde/1.0 fgh | 1.4 cde | 1.9 efg | 1.6 ij |
| 0.4 2iP | 78 c/8 n | 1.6 def/0.2 mn | 1.2 def | 1.4 ghi | 1.7 hi |
| 0.8 2iP | 63 ghi/17 lm | 1.6 def/0.2 mn | 1.0 ef | 1.7 efg | 1.7 hi |
| 1.6 2iP | 33 n/33 hij | 1.5 efg/0.8 hij | 0.9 efg | 1.4 ghi | 0.7 klm |
| 3.2 2iP | 25 o/33 hij | 1.5 efg/0.7 ij | 0.9 efg | 1.8 efg | 0.7 klm |
| 0.4 mT | 56 jk/27 jk | 1.7 def/0.6 j | 1.9 abc | 2.7 bcd | 2.6 def |
| 0.8 mT | 67 fgh/29 ij | 2.2 abc/0.9 gh | 1.4 cde | 2.4 bcd | 3.5 b |
| 1.6 mT | 58 ij/42 de | 1.7 def/1.4 d | 1.2 def | 1.6 fgh | 1.6 ij |
| 3.2 mT | 38 mn/62 a | 1.5 ghi/1.6 cd | 1.0 ef | 2.0 def | 1.1 jk |
| Fone-way ANOVA | ***/*** | ***/*** | *** | *** | *** |
| PGRs Concentration (mg L−1) | Shoot- Producing Explants 1/2 (%) | Mean Number of NSh ₸/HSh ₸₸ | Mean NSh Length ₸ (cm) | Mean Node Number ₸ | Multi-Plication Index |
|---|---|---|---|---|---|
| Shoot tip explant | |||||
| 0.0 (Hf ₸₸₸) | 93 a z/7 p | 1.0 l/0.1 j | 3.4 c | 3.7 bc | 3.4 g |
| 0.4 BA | 77 cd/23 op | 2.6 de/1.0 ef | 3.7 bc | 3.6 bc | 7.2 a |
| 0.8 BA | 50 jk/50 hi | 2.6 de/1.5 de | 3.8 bc | 4.1 ab | 5.3 c |
| 1.6 BA | 50 jk/50 hi | 2.9 bc/1.6 de | 2.9 cd | 3.3 cd | 4.8 cd |
| 3.2 BA | 43 jkl/57 gh | 3.5 a/1.8 d | 2.4 fg | 2.8 ef | 4.2 ef |
| 0.4 ZEA | 79 bcd/21 op | 1.1 kl/0.2 ij | 5.6 a | 4.3 a | 3.7 fg |
| 0.8 ZEA | 58 hi/42 jk | 1.0 l/0.4 hi | 2.6 de | 4.2 a | 2.4 jk |
| 1.6 ZEA | 55 ij/45 ij | 1.2 jk/0.7 gh | 2.9 cd | 3.6 bc | 2.4 jk |
| 3.2 ZEA | 29 mn/71 d | 2.3 fgh/1.3 e | 2.0 ijk | 2.5 fgh | 1.7 mn |
| 0.4 KIN | 75 de/17 p | 1.0 l/0.2 ij | 2.5 ef | 2.6 fg | 2.0 lm |
| 0.8 KIN | 75 de/25 n | 1.1 kl/0.5 hi | 2.6 de | 2.8 ef | 2.3 jk |
| 1.6 KIN | 71 ef/29 mn | 1.1 kl/0.7 gh | 2.5 ef | 2.5 fgh | 2.0 lm |
| 3.2 KIN | 67 fg/33 lm | 1.8 hi/0.8 fg | 2.5 ef | 2.8 ef | 3.4 g |
| 0.4 2iP | 83 b/17 p | 1.0 l/0.2 ij | 1.8 jk | 2.5 fgh | 2.1 kl |
| 0.8 2iP | 71 ef/25 n | 1.0 l/0.3 ij | 1.9 jk | 2.9 ef | 2.1 kl |
| 1.6 2iP | 62 gh/38 klm | 1.0 l/0.4 hi | 1.9 jk | 3.3 cd | 2.0 lm |
| 3.2 2iP | 58 hi/42 jk | 1.0 l/0.4 hi | 2.0 ijk | 4.0 ab | 2.3 jk |
| 0.4 mT | 67 fg/33 lm | 2.5 efg/0.4 hi | 2.6 de | 3.5 bc | 5.9 b |
| 0.8 mT | 40 kl/60 f | 2.9 bc/0.6 gh | 2.4 fg | 3.3 cd | 3.8 g |
| 1.6 mT | 20 n/80 bc | 3.0 b/1.1 ef | 2.3 g | 2.8 ef | 1.7 mn |
| 3.2 mT | 10 o/83 b | 3.3 b/1.2 e | 1.4 m | 1.5 l | 0.5 p |
| Single node explant | |||||
| 0.0 (Hf ₸₸₸) | 79 bcd/17 p | 1.9 hi/0.4 hi | 1.9 jk | 2.4 gh | 3.6 fg |
| 0.4 BA | 63 gh/33 lm | 2.5 efg/1.7 d | 2.5 ef | 3.0 de | 4.7 cd |
| 0.8 BA | 58 hi/42 jk | 2.6 de/1.8 d | 2.8 d | 3.0 de | 4.5 de |
| 1.6 BA | 3 p/97 a | 2.0 gh/2.5 c | 1.5 lm | 2.5 fgh | 0.2 p |
| 3.2 BA | 0 q/100 a | 0.0 m/4.1 b | - | - | 0.0 r |
| 0.4 ZEA | 67 fg/33 lm | 1.8 hi/0.9 fg | 3.9 b | 3.3 cd | 4.0 ef |
| 0.8 ZEA | 54 ij/46 ij | 2.0 gh/0.8 fg | 2.6 de | 3.1 de | 3.3 gh |
| 1.6 ZEA | 35 lmn/65 e | 2.0 gh/1.3 e | 1.6 lm | 2.3 hij | 1.6 n |
| 3.2 ZEA | 23 n/77 c | 2.0 gh/1.5 de | 1.5 lm | 2.4 gh | 1.1 o |
| 0.4 KIN | 79 bcd/17 p | 1.6 i/0.3 ij | 1.7 klm | 1.7 kl | 2.1 kl |
| 0.8 KIN | 83 b/17 p | 1.7 i/0.5 hi | 1.7 klm | 1.9 jk | 2.7 i |
| 1.6 KIN | 63 gh/33 lm | 1.8 hi/0.7 gh | 1.8 jk | 1.7 kl | 1.9 mn |
| 3.2 KIN | 54 ij/46 ij | 1.9 hi/1.1 ef | 1.9 jk | 2.2 hij | 2.3 jk |
| 0.4 2iP | 79 bcd/21 op | 2.0 gh/0.4 hi | 1.7 klm | 2.0 ij | 3.2 gh |
| 0.8 2iP | 63 gh/33 lm | 2.0 gh/0.7 gh | 1.9 jk | 2.7 ef | 3.4 g |
| 1.6 2iP | 54 ij/42 jk | 1.9 hi/0.9 fg | 2.0 ijk | 2.8 ef | 2.9 hi |
| 3.2 2iP | 50 jk/50 hi | 2.0 gh/1.1 ef | 2.1 hi | 2.8 ef | 2.8 hi |
| 0.4 mT | 57 hi/43 jk | 2.7 cde/0.6 gh | 2.0 ijk | 2.6 fg | 4.0 ef |
| 0.8 mT | 30 mn/70 d | 2.7 cde/0.7 gh | 1.6 lm | 2.4 gh | 1.9 mn |
| 1.6 mT | 20 n/80 bc | 2.8 bc/1.2 e | 1.7 klm | 3.2 cd | 1.8 mn |
| 3.2 mT | 0 q/97 a | 0.0 m/5.5 a | - | - | 0.0 r |
| Fone-way ANOVA | ***/*** | ***/*** | *** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papafotiou, M.; Vlachou, G.; Martini, A.N. Investigation of the Effects of the Explant Type and Different Plant Growth Regulators on Micropropagation of Five Mediterranean Salvia spp. Native to Greece. Horticulturae 2023, 9, 96. https://doi.org/10.3390/horticulturae9010096
Papafotiou M, Vlachou G, Martini AN. Investigation of the Effects of the Explant Type and Different Plant Growth Regulators on Micropropagation of Five Mediterranean Salvia spp. Native to Greece. Horticulturae. 2023; 9(1):96. https://doi.org/10.3390/horticulturae9010096
Chicago/Turabian StylePapafotiou, Maria, Georgia Vlachou, and Aikaterini N. Martini. 2023. "Investigation of the Effects of the Explant Type and Different Plant Growth Regulators on Micropropagation of Five Mediterranean Salvia spp. Native to Greece" Horticulturae 9, no. 1: 96. https://doi.org/10.3390/horticulturae9010096
APA StylePapafotiou, M., Vlachou, G., & Martini, A. N. (2023). Investigation of the Effects of the Explant Type and Different Plant Growth Regulators on Micropropagation of Five Mediterranean Salvia spp. Native to Greece. Horticulturae, 9(1), 96. https://doi.org/10.3390/horticulturae9010096