Abstract
Although using non-Saccharomyces yeasts during alcoholic fermentation can improve the wine aroma, most of them are not ethanol tolerant; therefore, in 2017, this study screened 85 non-Saccharomyces yeasts isolated and identified from 24 vineyards in seven Chinese wine-producing regions, obtaining Pichia terricola strain H5, which displayed 8% ethanol tolerance. Strain H5 was subjected to ultraviolet (UV) irradiation and diethyl sulfate (DES) mutagenesis treatment to obtain mutant strains with different fermentation characteristics from the parental H5. Compared with strain H5, the UV-irradiated strains, UV5 and UV8, showed significantly higher ethanol tolerance and fermentation capacity. Modified aroma profiles were also evident in the fermentation samples exposed to the mutants. Increased ethyl caprate, ethyl caprylate, and ethyl dodecanoate content were apparent in the UV5 samples, providing the wine with a distinctly floral, fruity, and spicy profile. Fermentation with strain UV8 produced a high ethyl acetate concentration, causing the wine to present a highly unpleasant odor. To a certain extent, UV irradiation improved the ethanol tolerance and fermentation ability of strain H5, changing the wine aroma profile. This study provides a theoretical basis for the industrial application of non-Saccharomyces yeasts that can improve wine flavor.
1. Introduction
Wine making involves microbial interaction. Although bacteria, fungi, and yeasts participate during the fermentation process, yeasts play a leading role []. Saccharomyces cerevisiae is mainly responsible for alcoholic fermentation and is the dominant strain during the wine-making process; however, in addition to Saccharomyces cerevisiae, many non-Saccharomyces yeasts are also involved []. Non-Saccharomyces yeasts refer to a large group of yeasts other than Saccharomyces cerevisiae, which exist during all the stages of wine making after the grapes are harvested, including Metschnikowia pulcherrima, Starmerella stellata, Torulaspora delbrueckii, Debaryomyces hansenii, Pichia fermentans, Hanseniaspora uvarum, Pichia terricola, etc. [,,]. They mainly improve the aroma of wine via metabolism and autolysis, and participate in the formation of complex, fresh flavor substances []. These yeasts improve the taste of wine, providing it with fruity, floral, and nutty aroma characteristics, which positively impacts the wine quality [,]. Previous studies have shown an increase in the acetate, phenethyl acetate, and 2-phenylethanol content in wine produced via the combined fermentation of Hanseniaspora vineae and Saccharomyces cerevisiae, while the ethyl ester proportion decreases, enhancing the fruity flavor and complexity of the wine []. Whitener et al. [] studied Kazachstania gamospora fermentation, revealing that it produced more floral-scented esters (such as phenethyl propionate) than commercial Saccharomyces cerevisiae. Mixed fermentation with Pichia kluyveri and Saccharomyces cerevisiae can increase the concentration of volatile mercaptans, especially hexyl 3-mercaptoacetate, which is recommended for the production of Sauvignon Blanc, Chardonnay, and Riesling wines [].
Non-Saccharomyces yeasts represent the dominant microbiota during the early stage of wine making []; however, many factors affect the growth and metabolism of non-Saccharomyces yeasts during the complex fermentation process, such as ethanol, SO2, temperature, sugar, and acid. Of these, ethanol stress is a vital influencing factor. The ethanol concentration continues to rise as fermentation progresses, leading to a continuous decline in the number of non-Saccharomyces yeasts during the middle and late stages, with some even dying [], reducing their positive effect on the final wine quality. Different methods have been adopted for selection and cultivation to improve the yeast tolerance to high ethanol stress conditions. Using chemical mutagens and ultraviolet (UV) irradiation for random mutagenetic breeding presents advantages, such as a short mutagenetic cycle and high mutagenetic efficiency, providing practical strategies to modify the physiological yeast characteristics [,]. UV mutagenesis can induce cytosine to thymine nucleotide base substitution and DNA photoproducts produced by UV radiation, such as cyclobutane pyrimidine dimers (CPDS) and (6-4) pyrimidine–pyrimidone photoproducts (6-4 PPs), to block the DNA replication process [,]. Diethyl sulfate (DES) can alkylate some oxygen sites, including the ethylation of the sixth guanine oxygen atom. The mutagenicity of DES manifests in the occurrence of mutations via direct mispairing [,].
This study subjected strain H5 to UV irradiation and DES mutagenesis to obtain mutants with higher ethanol tolerance. Applying these mutants to fermentation revealed that their fermentation characteristics differed from the parental H5. Mutant UV5 added a distinctly floral, fruity, and spicy aroma to the sample, while strain UV8 produced a high ethyl acetate concentration, causing a highly unpleasant odor.
2. Materials and Methods
2.1. Yeast Strains and Growth Conditions
In 2017, the 85 non-Saccharomyces yeasts used in this experiment were identified and isolated from 24 vineyards in seven Chinese wine-producing regions. The yeast information is shown in Table S1.
The strains were stored at −80 °C in cryovials containing 20% (v/v) glycerol. The strains were cultured and propagated in YPD liquid medium (2% glucose, 2% peptone, and 1% yeast extract (all w/v)). During the study, the strains were cultured on the YPD agar medium and stored at 4 °C.
2.2. Screening the Non-Saccharomyces Yeasts for Ethanol Tolerance
2.2.1. High-Throughput 2,3,5-Triphenyltetrazolium Chloride (TTC) Medium Method for First Stage Selection
The method described in the previous study [] was used with minor changes. Yeast cell suspensions were diluted in gradients and plated on the TTC lower medium (YPD agar medium at pH 5.5), while three parallel inoculations were performed for each yeast. The plates were incubated at 28 °C for 48 h and stained by a lay of TTC upper medium (0.5% glucose, 1.5% agar, and 0.05% TTC). The red colonies were selected after incubation at 28 °C in the dark for 3 h. TTC, a redox substance that can react with yeast metabolites, can be reduced to red by hydrogen. It can be used to judge the dehydrogenase activity in yeast, that is, the alcohol production capacity of yeast.
2.2.2. Detecting Non-Saccharomyces Yeast Activity under Ethanol Stress
The loopful volume of the cell suspensions (OD600 = 1.2) were streaked on YPD solid plates at ethanol volume fractions of 4%, 6%, 8%, 9%, 10%, 11%, and 12% (v/v). Three parallels were set for each treatment and incubated at 28 °C for 72 h. The colony growth on the plate was monitored, with the untreated plate as a control.
2.2.3. The Influence of Ethanol Addition on Yeast Cell Growth
Here, 2% of the yeast cell suspensions, which were diluted to OD600 of 1.2, were inoculated into the YPD liquid medium at ethanol concentrations of 0%, 4%, 6%, 8%, and 10% (v/v), and cultured at 28 °C in a rotary shaker at 180 rpm. Each treat was set in three parallels, and the untreated medium was used as a control. The absorbance of the yeast cultures was measured at 600 nm at 3-h intervals (at 6-h intervals in stationary phase) for 30 h to determine their growth rates under ethanol stress.
2.3. Mutation Procedure of Non-Saccharomyces Yeast by DES and UV
To obtain strains with higher ethanol tolerance, the cell suspension of strain H5 was subjected to DES mutagenesis and UV irradiation, using a method described by Yi et al. [] with minor modifications. The treatment conditions with a lethality rate of about 80% were set as the mutation conditions []. During UV irradiation, the cell suspension (OD600 = 1.2) was spread on the YPD agar plate by the gradient dilution coating plate method, and irradiated with UV (20 W) at a distance of 50 cm for 0, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, and 120 s. After irradiation, the plates were immediately shielded from light and incubated at 28 °C in the dark for 48 h. During DES treatment, 10 mL of the H5 bacterial suspension cultured to the mid-log phase (OD600 = 1.2) was centrifuged at 805× g for 10 min. The precipitate was washed twice with phosphate buffer (pH 7.2), and then redissolved in 10 mL of phosphate buffer. The cell suspension was treated with 10 mL of 1% DES in phosphate buffer (pH 7.2) for 0, 2, 4, 6, 8, 10, 12, 14, 16, 18, and 20 min, after which the procedure was terminated with 10 mL of 25% sodium thiosulfate. After the gradient dilution of the treated cells with sterile saline, they were spread on a YPD agar plate and incubated at 28 °C for 48 h.
The lethality was computed as follows:
Lethality (%) = (Number of strains before mutation − Number of strains after mutation)/(Number of strains before mutation) × 100.
2.4. Identification of Yeast
The strains requiring identification were inoculated in YPD liquid medium and incubated for 12 h at 28 °C while exposed to constant shaking at 160 rpm. The yeast cells were harvested after the centrifugation of 2 mL of the cell suspension, and the ribosomal DNA genomes of yeast were extracted using a TIANamp Yeast DNA Kit (by TIANGEN Biotech (Beijing) Co., Ltd., Beijing, China). ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [] primers were used for PCR amplification of the 5.8S-ITS rDNA sequence []. The PCR cycling conditions were as follows: 95 °C pre-degeneration for 5 min; 95 °C degeneration for 1 min, 52 °C annealing for 2 min, 72 °C extension for 2 min 20 s, repeated 35 times; complementary extension at 72 °C for 10 min. The reaction system consisted of 50 μL 2×TSINGKE® MasterMix 25 μL, 10 μ MF-primer 1 μL, 10 μM R-primer 1 μL, template DNA 1 μL, and DD water 22 μL. The PCR products were detected via 1% agarose gel electrophoresis and sent to Beijing Tsingke Biological Technology for sequencing. The gene sequences of the strains were searched on the NCBI website (https://blast.ncbi.nlm.nih.gov/Blast.cgi) (21 February 2022), and compared with similar sequences using MEGABLAST to preliminarily determine the strain species based on the similarity of homologous sequences. The strains were then compared with the ITS sequences of the most similar type strains found in the MEGABLAST search by pairwise Blast alignment.
2.5. The Practical Fermentation of Selected Strains
Cabernet Sauvignon grape juice (2018, Changli Hebei) was used for single-strain fermentation. Grape juice stored at −20 °C was thawed at 4 °C. After centrifugation at 13,919× g for 15 min at 4 °C, the grape juice was sterilized using 0.65 μm and then 0.45 μm filter membranes. Separate 500 mL fermentation bottles were filled with 200 mL of sterile grape juice. The strains were pre-cultured in YPD medium at 28 °C and 120 rpm for 24 h and then statically incubated in sterile grape juice at 28 °C for 72 h to obtain 107 CFU/mL cells. The pre-cultured strains were inoculated with 1% inoculum at 0 d of fermentation. With commercial Saccharomyces cerevisiae S1 as the control group, all strains were subjected to single-strain fermentation, and each fermentation group was triplicated. Each group was subjected to static fermentation at 20 °C, and samples were taken at 0, 2, 4, 7, 10, 13, and 16 d of fermentation to determine the number of yeast colonies, °Brix, and pH value, as well as the residual sugar, glycerin, ethanol, acid, and volatile aroma compound content. The remaining samples were stored at −20 °C for high-performance liquid chromatography (HPLC) and headspace-solid phase microextraction/gas chromatography–mass spectrometry flame-ionization detection (HS-SPME/GC-MS-FID).
2.6. Determining the Physicochemical Indexes via HPLC
A method described by Sun et al. [] was used to determine the content of glucose, fructose, glycerol, and ethanol. The chromatographic conditions included an Aminex HPX-87H (300 mm × 7.8 mm) analytical column at a temperature of 55 °C; the differential detector was Waters-2414 (Waters, Dublin, Ireland); an internal temperature of 40 °C; respective sample injection volumes of 10 μL, a mobile phase of 0.005 mol/L H2SO4, a flow rate of 0.5 mL/min, and isocratic elution. Qualitative analysis was based on retention time. Quantitative calculation occurred according to peak areas.
The oxalic acid, citric acid, tartaric acid, malic acid, succinic acid, lactic acid, and acetic acid content were determined according to a method described by Chen et al. [], with minor modifications. The chromatographic conditions included an Aminex HPX-87H (300 mm × 7.8 mm) analytical column at a temperature of 40 °C; the photodiode array detector was Waters-2996 (Waters, Dublin, Ireland); a detection wavelength of 210 nm; a mobile phase of 0.005 mol/L H2SO4; a flow rate of 0.4 mL/min; respective injection volumes of 20 μL; isocratic elution. Qualitative analysis was based on retention time. The quantification occurred according to the peak area.
2.7. Determining the Aroma Compounds via HS-SPME/GC-MS-FID
The volatile aroma compounds were determined using a method delineated by Chen, Chia, and Liu []. HS-SPME/GC-MS-FID was used to determine the volatile aroma compounds in the sample. The pH of wine samples was adjusted to 2.5 ± 0.1 using 1 mol/L hydrochloric acid. A 20 mL headspace bottle was filled with 5.0 mL of the wine sample and sealed with a cap with a rubber septum. The injection port temperature was 250 °C, the carrier gas (He) flow rate was 1.0 mL/min, and the splitless injection mode was selected. The temperature programming conditions included an initial temperature of 50 °C for 5 min, which was increased to 230 °C at a rate of 5 °C/min, and maintained for 30 min. The signals were acquired in full scan mode via electron ionization (EI) at 70 eV, an interface temperature of 280 °C, an ion source temperature of 230 °C, a quadrupole temperature of 150 °C, a scanning mass range (m/z) of 35–500 amu, and a scan speed of 5.2 times/S, and a solvent delay of 3 min. The qualitative analysis of aroma components was performed using the NIST 14 mass spectrum library, while the relative percentages of each volatile compound were determined according to the GC-FID peak area. All samples were tested in triplicate.
2.8. Sensory Evaluation
The wine aroma evaluation panel consisted of 12 members who had extensive wine tasting experience. Members awarded the samples scores ranging from 1–5 and evaluated six aromatic qualities of the Cabernet Sauvignon wine (alcohol, floral, fruity, preserved fruit, spicy, and fermented aroma). Furthermore, the participants performed a preference ranking and difference comparison. The samples were randomly numbered and presented to the panelists. The aroma characteristics were characterized via the five-point intensity method (blank—no odor, 1—just detected the odor, 2—weak, 3—medium, 4—strong, 5—intense).
2.9. Statistical Analysis
The mean and standard variance were calculated using the data of triplicate fermentations, and the results were expressed as mean ± standard deviation (SD). The significant differences were carried out using SPSS 21.0 software (SPSS Inc., Chicago, IL, USA) based on a one-way analysis of variance (ANOVA, significance level was 0.05) with Duncan’s multiple range test. The Shapiro–Wilk’s and Levene’s tests were used in the test for the normality and homogeneity of variances. Principal component analysis (PCA) was performed using Matlab R2013b software (MathWorks, Natick, MA, USA).
3. Results
3.1. Selecting a Non-Saccharomyces Yeast with High-Alcoholicity Endurance
The TTC medium chromogenic method was employed to examine the red color intensity of 85 non-Saccharomyces strains. The results demonstrated that the chromogenic levels of the strains were significantly different. The strains B21, D1, D6, F26, E31, and H5 displayed a prominent dark red color and had better consistency between the three parallel. Commercial Saccharomyces cerevisiae S1 was used as a positive control, and the strain H67 was used as a negative control. The colorimetric results are shown in Figure 1a. Although the TTC medium method could quickly screen out the strains displaying better color, quantitative evaluation was impossible; therefore, six selected yeasts were utilized in the follow-up study to determine their tolerance at different ethanol concentrations. As shown in Table 1, strain D1 exhibited the worst ethanol tolerance and only proliferated at a 4% ethanol concentration. Strains B21 and E31 showed good tolerance of a 6% ethanol concentration. The strains displaying the highest ethanol tolerance were F26, H5, and D6, which could form colonies on the medium containing 8% ethanol.
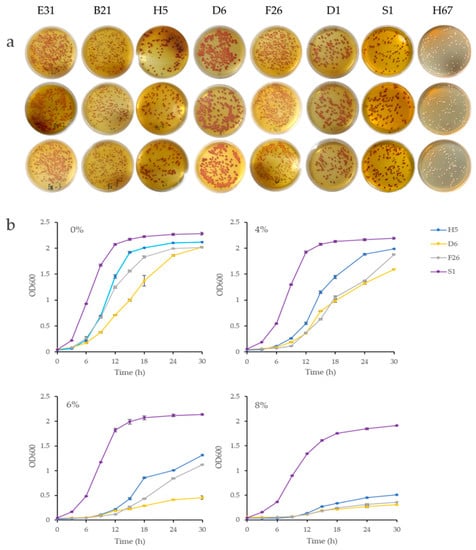
Figure 1.
(a) The coloration results obtained via the TTC medium chromogenic screening method. The red intensity level of colonies in the culture medium can indirectly reflect the ethanol tolerance of the strains; (b) the growth curves of the screened strains at ethanol concentrations of 0%, 4%, 6%, and 8% (v/v).

Table 1.
The ethanol tolerance results of the screened strains.
To further evaluate the tolerance of the D6, F26, and H5 strains against ethanol, their growth curves at different ethanol concentrations were measured at OD600, as shown in Figure 1b. Commercial Saccharomyces cerevisiae S1 was used as the control group. In ethanol conditions, all strains displayed extended lag phases, reduced growth rates, and significantly lower maximal OD values than in non-ethanol conditions (p < 0.05). At an ethanol concentration of 6% (v/v), the proliferation of strains D6, F26, and H5 was noticeably inhibited. The absorbance values of strains H5 and F26 at 30 h were only half that of the control group, while strain D6 was reduced to one-fourth. At an ethanol concentration of 8% (v/v), the absorbance values of the three strains when reaching the stationary phase were less than one-fourth of the control group; however, in general, the growth rate of H5 was significantly faster than F26 and D6 at different ethanol concentrations; therefore, H5 was selected for subsequent experiments.
3.2. Screening for Non-Saccharomyces Yeasts with High Ethanol Tolerance Using DES and UV
The effect of a mutagen dosage on the lethality of strain H5 is shown in Figure 2. The cells cultivated to the logarithmic phase were subjected to UV irradiation and DES mutagenesis. Figure 2a shows that the lethality increased with extended UV irradiation time. The lethality reached 76.28% and 90.29% at UV irradiation times of 40 s and 50 s, respectively. Figure 2b shows that the lethality increased with extended DES mutagenesis time. The lethality reached 69.38% and 83.14% at DES mutagenesis times of 8 min and 10 min, respectively. The treatment condition that produced a lethality of about 80% was set as the mutation concentration []; therefore, a UV irradiation time of 40 s (lethality 76.28%) and a 1% DES mutagenesis time of 10 min (lethality 83.14%) were set as the mutation conditions for strain H5.
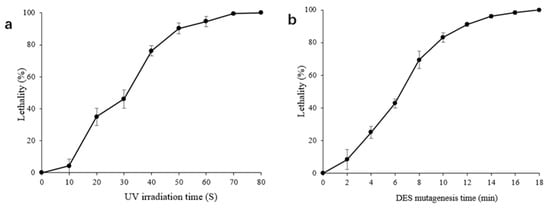
Figure 2.
The lethality curve during mutagenesis. (a) The lethality curve of strain H5 subjected to UV mutagenesis after exposure to different irradiation times. A UV irradiation time of 40 s was set as the mutation conditions for strain H5; (b) the lethality curve of strain H5 subjected to DES mutagenesis after exposure to different times. Treatment with 1% DES for 10 min was set as the mutation conditions for S. cerevisiae. The results expressed as mean ± SD.
After treatment with 1% DES for 10 min and UV irradiation for 40 s, 15 mutants (UV1-UV8 and DES1-DES7) displaying a dark red colony color and screened via high-throughput TTC medium were collected. These strains were streaked onto the YPD solid medium containing ethanol to evaluate the ethanol tolerance of the mutant strains. Strain H5 and commercial Saccharomyces cerevisiae S1 were used as controls, and the results are shown in Table 2. The ethanol tolerance of the UV5 and UV8 mutants improved significantly after UV irradiation, increasing to 11% and 10%, respectively. After DES treatment, at the same ethanol concentration (8%), only DES7 exhibited better proliferation than strain H5, while the ethanol tolerance of the other strains did not improve.
Table 2.
The ethanol tolerance results of mutants H5 and S1 introduced as control strains.
To further explore the ethanol tolerance of the mutants, their growth curves were measured at different ethanol concentrations, using strain H5 and commercial Saccharomyces cerevisiae S1 as controls, as shown in Figure 3. The growth rates of strain UV5 and UV8 were higher than that of strain H5 at different ethanol concentrations, but they did not exceed that of commercial Saccharomyces cerevisiae S1. At an ethanol concentration of 6%, the absorbance values of UV5 and UV8 exceeded 2.00 at 30 h, while that of strain H5 was 0.84. At an ethanol concentration of 8%, the absorbance values of UV5 and UV8 remained higher than 1.00 after 30 h, while that of strain H5 was only 0.46. When the ethanol concentration reached 10%, strain H5 was completely inhibited, and the OD value did not increase, while strains UV5 and UV8 continued to proliferate, and the absorbance values exhibited significant differences (p < 0.05); however, the growth of strain DES7 at 6% and 8% ethanol concentrations was slightly superior to that of strain H5, while the absorbance values were significantly different (p < 0.05) at 30 h. DES7 showed no growth at 10% ethanol concentration, which was consistent with the results in the solid YPD medium. The findings indicated that the ethanol tolerance of strains UV5 and UV8 surpassed that of strain H5.
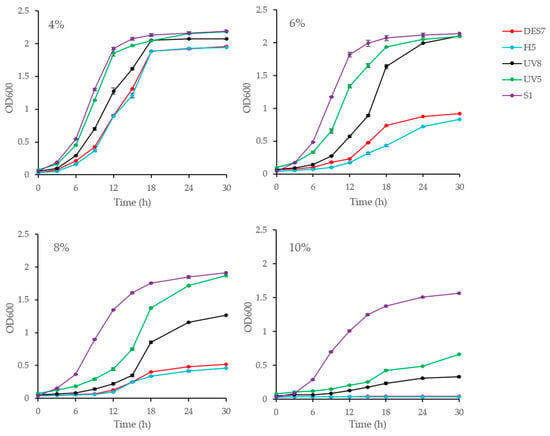
Figure 3.
The growth curves of the mutants at ethanol concentrations of 4%, 6%, 8%, and 10% (v/v). Strain H5 and S1 served as controls at various ethanol concentrations.
3.3. Evaluation of the Fermentation Ability of the Mutagenized Strains
3.3.1. The Performance of the Mutants during Fermentation
The yeast cell dynamics during the fermentation process are shown in Figure 4a, while the °Brix change is shown in Figure 4b. As shown in Figure 4a, the yeasts continued to proliferate as the fermentation progressed, reaching their maximum count at 7 d and maintaining high cell viability until the end of the process. The °Brix value results showed that neither strain H5 nor the three mutants could fully utilize the sugar content in grape must.
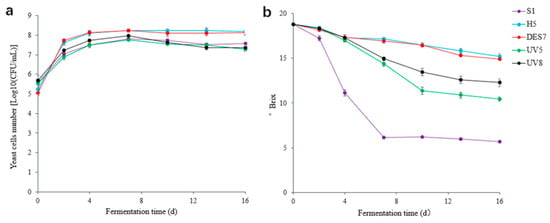
Figure 4.
(a) The changes in the colony numbers during fermentation; (b) changes in the °Brix values of the different fermentation groups during fermentation.
The impact of the yeast metabolism on the wine composition was evaluated, quantifying the main analytical components of the grape must and wine at the end of fermentation. The chemical analysis results are summarized in Table 3. Except for commercial Saccharomyces cerevisiae S1, the remaining four yeast strains still contained residual sugar that could not be ignored at 16 d fermentation (>0.4 g/L). This confirmed that they could not complete the wine fermentation process independently, which was consistent with the °Brix value test results. The ability of strain UV5 to utilize glucose and fructose improved significantly compared with strain H5, reducing the content of these substances to 15.17 g/L and 40.10 g/L, respectively. Strain UV8 also used more glucose, reducing the residual glucose content to 31.83 g/L after fermentation. At the end of fermentation, the ethanol and glycerol content produced by the three mutant strains and strain H5 were significantly lower than those of strain S1. The ethanol yield of strain UV5 was 3.20 times that of the H5, while the glycerol yield increased by 16.27%. Although strain UV8 consumed more glucose than strain H5, the ethanol production only increased by 0.39% (v/v), while no significant differences were evident in glycerol production (p < 0.05). The glucose, fructose, ethanol, and glycerol levels indicated that the fermentation performance of UV5 and UV8 was surpassed that of strain H5, while the fermentation performance of UV5 was stronger. The data indicated that UV irradiation improved the fermentation performance of strain H5, which was not significantly different from that of strain DES7 (p < 0.05).
Table 3.
The physicochemical indexes in the grape must and wine.
Organic acids are important compounds found in grape must and wine. After alcoholic fermentation, the citric acid content of strain UV8 was the highest at 0.88 g/L, while that of DES7 was the lowest at 0.58 g/L. Tartaric acid represents the dominant organic acid in wine, primarily contributing to its acidity and tart taste []. The tartaric acid content of strains UV5, UV8, and S1 was significantly reduced after fermentation compared with grape must. This was attributed to a decline in the solubility and precipitation of tartaric acid crystals due to alcohol accumulation during fermentation, decreasing the tartaric acid content [,]. Malic acid provides a green, astringent taste that can be synthesized or degraded by yeast during alcoholic fermentation [,]. Significant differences were apparent between the malic acid levels of the five experimental groups. Compared with grape must, the malic acid content declined substantially after fermentation. The lactic acid in wine is mainly produced during malolactic fermentation. Lactic acid bacteria convert part of the malic acid into lactic acid, rendering the taste of the wine softer and less harsh. Only some yeast strains can produce trace amounts of lactic acid during the fermentation process due to the lack of an effective lactic acid pathway []. During this experiment, lactic acid was only detected in strains H5 and DES7, providing milky and buttery flavors to the wine. The results suggested that UV irradiation affected the metabolic capacity of the organic acids in yeasts.
3.3.2. The Influence of Mutants on the Volatile Aroma Compounds
The volatile metabolites in grape must and wine samples were identified via HS-SPME/GC-MS-FID. A total of 102 volatile aroma compounds were detected, which could be divided into six categories: alcohols, acids, esters, aldehydes and ketones, terpenes, and other compounds. The volatile aroma compounds in the samples were semi-quantitatively analyzed using the area normalization method. The peak area and relative content of each substance are shown in Table S2.
Twenty-six aroma compounds were detected in the grape must, including eight alcohols, one acid, one ester, eight aldehydes and ketones, one terpene, and seven other aroma compounds. After fermentation, the aroma compound diversity detected in each sample was significantly increased compared with the grape must. Fifty-five volatile aroma compounds were identified in the S1 fermentation samples, 59 in strain H5, 56 in DES7, 54 in UV5, and 48 in UV8. The aroma compounds detected during the fermentation processes mainly consisted of esters, alcohols, and acids. Although the total number of aroma compounds in each sample was similar, the types and relative content were significantly different. Compared with the S1 group, the unique volatile compounds detected in other groups included citronellol, 2-methylpropanol, benzyl alcohol, isovaleric acid, hexyl caproate, methyl salicylate, etc. Compared with strain H5, mutant DES7 displayed fewer aroma compound species and relative peak areas; however, although the aroma compound types decreased in UV5 and UV8, the relative peak areas were higher.
As vital aroma compounds produced during alcoholic fermentation, esters present strong floral, fruity flavors and significantly contribute to the fruity aroma of young wines [,]. The total peak area of the esters in group S1 was 2357.37 × 106, which was significantly higher than in other groups. The total peak area of the esters in group H5 was 294.87 × 106, accounting for 58.74% of the total aroma content, among which ethyl caproate was the most abundant, accounting for 19.04%, providing tropical fruit aromas, such as bananas and pineapples. The total peak area of the esters in the DES7 group was 202.57 × 106, accounting for 49.56% of the total aroma content, of which ethyl caproate was the highest, accounting for 22.83%. The relative ester content in UV5 was significantly higher than in H5. The total peak area was 1790.31 × 106, accounting for 88.19% of the total aroma content. Furthermore, the ethyl caprate, ethyl caprylate, and ethyl dodecanoate were higher, accounting for 32.22%, 15.47%, and 15.47%, respectively. Ethyl decanoate provided oily, floral, grape, and other aromas, while ethyl caprylate was responsible for pleasant fruity, buttery, mushroom, and other aromas. Ethyl dodecanoate presented a strong aroma of peanuts []. The total peak area of the esters in the UV8 group was 938.43 × 106, accounting for 91.85% of the total aroma content, of which ethyl acetate was the most abundant, accounting for 74.85%. The peak area of the ethyl acetate in the UV8 group was significantly higher than in other groups, possibly indicating that UV8 can produce high ethyl acetate concentrations. Although ethyl acetate can add grape, cherry, and other fruit characteristics, when its content exceeds 100 mg/L, the wine presents a solvent/nail varnish-like and other unpleasant aromas [].
Higher alcohols are aroma compounds produced by yeast via amino acid metabolism and can enhance the wine aroma complexity to a certain extent []. Compared with the S1 group, more types of higher alcohols were evident in the other groups. Fourteen alcohols were detected in the H5 group, the relative content accounting for 32.44%. Although more than ten alcohols were detected in each mutant group, the relative alcohol content in the UV5 and UV8 groups were significantly reduced, at 3.69% and 5%, respectively. Phenethyl alcohol and isoamyl alcohol, representing higher alcohols synthesized by yeast, were the most abundant in each sample after fermentation, mainly providing aromas of roses and fruit, such as bananas. In addition, compared with strain H5, the unique alcohols detected in the DES7 group were 1,2-butanediol, trans-2-hexenol, octanol, while amyl alcohol was found in the UV5 group, and geraniol and 2, 6-octanediol in the UV8 group.
High acid levels may cause undesirable flavors in wine []. This experiment showed low acid content levels in each group after fermentation. Aldehydes and ketones are metabolites found during the early fermentation phase and are generally abundant in grape must. In this study, eight aldehydes and ketones were detected in the grape must, among which hexanal, hexenal, and β-damascenone displayed higher levels. After fermentation, the relative aldehyde and ketone content in each group was below 2%. These results indicated that the metabolic ability of the volatile aroma compounds of the strains was modified by DES mutagenesis and UV irradiation.
PCA was performed on the 43 primary aroma compounds to delineate the differences between these components in the different samples more intuitively. The results are illustrated in Figure 5a, showing that the variance contribution rate was 43.42% in the first principal component (PC1), and 22.48% in the second principal component (PC2), while the cumulative variance contribution rate was 65.90%. The aromas presented on the positive semi-axis of PC1 is mainly included fermented notes provided by alcohols, esters, and acids. UV5 and S1 were designated to the positive semi-axis of the PC1, while H5, DES7, UV8, and grape must were assigned to the negative semi-axis of PC1. The aroma mainly included that of the original grape must variety and some fermented aromas provided by hexanal, trans-2-hexenol, damascene, phenylacetaldehyde, citronellol, and phenethyl acetate. This may be related to the fermentation ability of the five strains, of which only S1 completed the alcoholic fermentation process. Of the remaining four strains, the fermentation ability of UV5 was the highest.
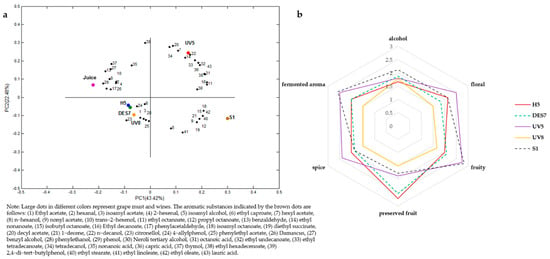
Figure 5.
(a) PCA of selected volatile compounds in Cabernet Sauvignon must and wine; (b) the aroma radar map of the sensory evaluation.
The positive semi-axis area of PC2 included ethyl caprylate, ethyl nonanoate, nerolid alcohol, lauric acid, n-hexanol, and hexyl acetate, while the negative semi-axis area was represented by nonyl acetate and caprylic acid, isoamyl esters, isoamyl alcohol, citronellol, phenethyl alcohol, and ethyl caproate. The five groups of samples were divided into four categories according to the scores of the two principal components. UV5 appeared in the upper right quadrant, S1 in the lower right quadrant, grape must in the upper left quadrant, and H5, DES7, and UV8 in the lower left quadrant. Although UV8 was in the third quadrant with H5 and DES7, as shown in Table S2, this strain contained a higher level of ethyl acetate, providing the wine with the unpleasant smell of nail polish. In addition, the grape must and UV5 were designated to the positive semi-axis of the Y-axis according to PC2, indicating that this strain retained the original grape must aroma within a certain range.
The aroma radar map of the sensory evaluation is shown in Figure 5b, indicating that the volatile profile of the UV5 group presented prominent floral, fruity, fermented, and spicy aromas. The S1 group displayed higher alcohol, fermented, and fruity aromas scores. The volatile profiles of H5 and DES7 were similar, with the preserved fruit aroma scoring the highest. Previous studies have shown that the β-glucosidase extracted from Pichia terricola can significantly increase the phenol and isoprenoid concentrations in wine, providing preserved fruit and raisin aromas []. The UV8 group displayed the smallest volatile profile and presented an unpleasant nail polish smell. The sensory evaluation and HS-SPME/GC-MS-FID results could be mutually verified.
3.4. Identification of Strain UV5
Strain UV5 was identified via molecular taxonomy via sequence similarity analysis of the ITS region of the ribosomal DNA gene. The sequence of the UV5 ITS region was consistent with the type of strain of Pichia terricola (CBS 2617), and the similarity percentage with the ITS sequence of the type strain was 99.51%; therefore, strain UV5 was identified as Pichia terricola.
4. Discussion
The aroma properties in wine are complex and essential for evaluating its flavor resulting from the interaction between various volatile components, such as esters, alcohols, aldehydes and ketones, acids, and terpenes []. Alcohols and esters mostly add rich floral and fruity characteristics to the wine. Aldehydes provide a floral, green apple aroma, while some acids and volatile phenols enhance wine aroma complexity; however, excessive amounts of these volatile aroma substances can cause undesirable and unanticipated odors []. Furthermore, wine can present different aroma characteristics and styles due to variation in metabolic pathways, enzymatic activity, and yeast strain metabolite type and content []. For example, the mono-culture fermentation of Torulaspora delbrueckii produces lower levels of volatile acids than Saccharomyces cerevisiae []. Metschnikowia pulcherrima generates more ethyl caprylate, providing the wine with pineapple or pear aromas []. Yeasts are in continuous and extensive competition during the alcoholic fermentation process. Temperature, alcohol, oxygen, SO2, carbon source, nitrogen source, and other factors affect this competitive relationship as fermentation progresses, impacting the taste and aroma characteristics of wine []. Toxic metabolites mediate ethanol inhibition. The toxic effect of ethanol on yeast is mainly reflected in the impact of cell morphology and physiological cell activity. Morphological cell modifications are primarily evident in cell enlargement and skeleton evacuation. Changes in the physiological cell activity are mainly represented by the destruction of the cell membrane structure, affecting the synthesis and metabolism of biological macromolecules, and modifying the activity of related enzymes during the glycolysis process [,]. Different yeast species display varying ethanol tolerance levels. Compared with Saccharomyces cerevisiae, non-Saccharomyces yeasts display a weaker tolerance to alcohol. Studies show that Saccharomyces cerevisiae 101, ITB, and AAV2 can tolerate 15% ethanol, while Issatchenkia occidentalis APC and Issatchenkia orientalis cl1132 can only tolerate 10% []. The present study showed that Pichia terricola H5 could tolerate 8% ethanol. Strains D6 (Wickerhamomyces anomalus) and F26 (Meyerozyma caribbica) were tolerant to an 8% ethanol concentration. This was consistent with the results obtained by Kim et al. [], who showed that W. anomalus and M. caribbica could proliferate at 8% ethanol.
The potential of genetically modified yeasts has been greatly weakened due to the severe restriction of wine markets to the application of genetically modified organisms. Non-genetically modified organism techniques have been used to improve the performance of yeasts to meet the demands of the wine industry []. To improve the ethanol tolerance of yeasts, the performance of the strains was modified via mutagenetic breeding and genetic recombination. Some studies constructed hybrids between Kluyveromyces fragilis and Saccharomyces cerevisiae using protoplast fusion technology to directionally change the ethanol tolerance of Kluyveromyces fragilis []. Compared with genetic recombination, mutation breeding presents advantages, such as simple operation, low technology and equipment requirements, and high mutation frequency; therefore, mutation breeding is widely used for yeast cultivation. Pattanakittivorakul et al. [] obtained a thermotolerant ethanol-fermenting yeast strain via UV radiation and ethyl methane sulfonate mutagenesis that effectively improved the efficiency of ethanol production. Watanabe et al. [] treated Pichia stipitis with UV irradiation for 25 min to obtain the Pichia stipitis NBRC1687 mutant after screening. It displayed significant ethanol tolerance, with the ethanol content produced via fermentation in the same conditions was 1.4 times that of the parental strain. In this experiment, Pichia terricola H5 was used as the mutagenic material and subjected to UV irradiation and DES mutagenesis to improve its ethanol tolerance. The results showed that the ethanol tolerance of UV5 and UV8 increased significantly after UV irradiation, reaching 11% and 10%, respectively, which was consistent with previous studies indicating that UV irradiation could modify the ethanol tolerance of yeast. Of the DES mutants, only the ethanol tolerance of DES7 was slightly higher than the parental H5, differing from the findings in existing literature []. This may be attributed to the DES concentration and the processing time used during the experiment. Less strain variability is evident at a lower mutagen concentration and shorter mutagenesis time, reducing the likelihood of changes in the strain performance [].
This study selected three mutants for alcoholic fermentation with commercial Saccharomyces cerevisiae S1 and strain H5 after analyzing their ethanol tolerance while evaluating the fermentation performance and volatile aromatic substances. The fermentation results showed that the three mutants and H5 could not complete alcoholic fermentation independently, while their fermentation ability was weaker than commercial Saccharomyces cerevisiae, which was consistent with the research results of Clemente-Jimenez et al. []. Although the residual sugar content exceeded 4 g/L when only Pichia terricola was used for alcoholic fermentation, the fermentation capacity of the UV5 mutant was improved. Compared with strain H5, the residual sugar content in the UV5 group was reduced by 32.83%, while the ethanol content was 3.75 times that of the H5 group. UV irradiation improved the ethanol tolerance and alcohol production of yeast, which was consistent with previous studies []. Furthermore, Pichia terricola could reduce acid levels by degrading malic acid and citric acid to improve the taste of the wine []. This study showed that the citric and malic acid levels were significantly lower in the H5 strain and DES7 mutant than in S1 while exhibiting higher levels in the UV5 group. It is speculated that while the UV-irradiated yeast develops ethanol resistance, it may also affect the metabolic cell pathways related to citric and malic acid; however, the specific reasons for this require further analysis. Clemente-Jimenez et al. [] indicated that Pichia terricola displayed low fermentation ability while producing high ethyl acetate levels, causing the wine to present an unpleasant odor. The H5 group did not exhibit high ethyl acetate content, while the sensory evaluation results did not indicate the presence of an unpleasant odor; however, high ethyl acetate levels were evident in the UV8 mutant group, displaying a severely unpleasant odor, which was consistent with the literature. This may be because UV irradiation not only induces the ability of cells to resist ethanol stress but also promotes the expression of genes related to ethyl acetate synthesis in cells; however, further research is necessary to provide confirmation. Moreover, the aroma determination results indicated that the fermentation of strain UV5 with improved ethanol tolerance produced higher levels of ethyl caprate, ethyl caprylate, and ethyl dodecanoate, providing a stronger floral, fruity fragrance.
Since this study only measured the ethanol tolerance of the strain at the macro level, the related stress resistance pathways and mechanisms require further exploration. Studies have shown that 359 genes in Saccharomyces cerevisiae cells are associated with ethanol tolerance []. The mutant genotypes and mutation sites require further exploration to clarify the differences in ethanol tolerance, fermentation characteristics, and aroma characteristics between the mutant and strain H5.
5. Conclusions
This research aims to improve the ethanol tolerance of non-Saccharomyces yeast and apply it to wine fermentation. An appropriate UV irradiation dose increased the ethanol tolerance of strain H5 and improved its fermentation performance. In particular, the production of ethyl caprate, ethyl caprylate, and other volatile aroma compounds in strain UV5 were significantly higher, which increased the aroma structure complexity of the wine.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/fermentation8040149/s1, Table S1: The region and vineyards of strains used; Table S2: The average peak area and relative peak area of volatile aroma compounds in each sample.
Author Contributions
Conceptualization, W.H. and J.Z.; Formal analysis, J.G., X.H. and Y.Y.; Funding acquisition, W.H. and J.Z.; Investigation, Y.Y.; Methodology, J.G. and X.H.; Project administration, J.G. and X.H.; Supervision, W.H. and J.Z.; Writing—review and editing, J.G., Y.Y. and J.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National “Thirteenth Five-Year” Plan for Science and Technology Support (2016YFD0400500).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gobbi, M.; De Vero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative Aptitude of Non-Saccharomyces Wine Yeast for Reduction in the Ethanol Content in Wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Prior, K.J.; Bauer, F.F.; Divol, B. The Utilisation of Nitrogenous Compounds by Commercial Non-Saccharomyces Yeasts Associated with Wine. Food Microbiol. 2019, 79, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled Mixed Culture Fermentation: A New Perspective on the Use of Non-Saccharomyces Yeasts in Winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Sadoudi, M.; Tourdot-Marechal, R.; Rousseaux, S.; Steyer, D.; Gallardo-Chacon, J.J.; Ballester, J.; Vichi, S.; Guerin-Schneider, R.; Caixach, J.; Alexandre, H. Yeast-Yeast Interactions Revealed by Aromatic Profile Analysis of Sauvignon Blanc Wine Fermented by Single or Co-Culture of Non-Saccharomyces and Saccharomyces Yeasts. Food Microbiol. 2012, 32, 243–253. [Google Scholar] [CrossRef]
- Cordero-Bueso, G.; Esteve-Zarzoso, B.; Cabellos, J.M.; Gil-Diaz, M.; Arroyo, T. Biotechnological Potential of Non-Saccharomyces Yeasts Isolated During Spontaneous Fermentations of Malvar (Vitis Vinifera Cv. L.). Eur. Food Res. Technol. 2013, 236, 193–207. [Google Scholar] [CrossRef]
- Giorello, F.; Valera, M.J.; Martin, V.; Parada, A.; Salzman, V.; Camesasca, L.; Farina, L.; Boido, E.; Medina, K.; Dellacassa, E.; et al. Genomic and Transcriptomic Basis of Hanseniaspora vineae’s Impact on Flavor Diversity and Wine Quality. Appl. Environ. Microb. 2019, 85, e01959-18. [Google Scholar] [CrossRef] [Green Version]
- Whitener, M.E.B.; Carlin, S.; Jacobson, D.; Weighill, D.; Divol, B.; Conterno, L.; Du Toit, M.; Vrhovsek, U. Early Fermentation Volatile Metabolite Profile of Non-Saccharomyces Yeasts in Red and White Grape Must: A Targeted Approach. LWT Food Sci. Technol. 2015, 64, 412–422. [Google Scholar] [CrossRef]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-Fermentation with Pichia Kluyveri Increases Varietal Thiol Concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Liu, P.T.; Lu, L.; Duan, C.Q.; Yan, G.L. The Contribution of Indigenous Non-Saccharomyces Wine Yeast to Improved Aromatic Quality of Cabernet Sauvignon Wines by Spontaneous Fermentation. LWT Food Sci. Technol. 2016, 71, 356–363. [Google Scholar]
- Heard, G.M.; Fleet, G.H. The Effects of Temperature and Ph on the Growth of Yeast Species During the Fermentation of Grape Juice. J. Appl. Bacteriol. 1988, 65, 23–28. [Google Scholar] [CrossRef]
- Addo-Quaye, C.; Tuinstra, M.; Carraro, N.; Weil, C.; Dilkes, B.P. Whole-Genome Sequence Accuracy Is Improved by Replication in a Population of Mutagenized Sorghum. G3 Genes Genomes Genet. 2018, 8, 1079–1094. [Google Scholar]
- Zhang, G.Q.; Lin, Y.P.; Qi, X.N.; Wang, L.X.; He, P.; Wang, Q.H.; Ma, Y.H. Genome Shuffling of the Nonconventional Yeast Pichia anomala for Improved Sugar Alcohol Production. Microb. Cell Fact. 2015, 14, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, L.J.; Lee, P.H.; Small, E.M.; Hillyer, C.; Guo, Y.; Osley, M.A. Regulation of UV Damage Repair in Quiescent Yeast Cells. DNA Repair 2020, 90, 102861. [Google Scholar] [CrossRef] [PubMed]
- Ikehata, H.; Ono, T. The Mechanisms of UV Mutagenesis. J. Radiat. Res. 2011, 52, 115–125. [Google Scholar] [PubMed] [Green Version]
- Hoffmann, G.R. Genetic Effects of Dimethyl Sulfate, Diethyl Sulfate, and Related-Compounds. Mutat. Res. 1980, 75, 63–129. [Google Scholar] [CrossRef]
- Abrol, V.; Kushwaha, M.; Arora, D.; Mallubhotla, S.; Jaglan, S. Mutation, Chemoprofiling, Dereplication, and Isolation of Natural Products from Penicillium oxalicum. ACS Omega 2021, 6, 16266–16272. [Google Scholar] [CrossRef]
- Pang, Z.W.; Liang, J.J.; Qin, X.J.; Wang, J.R.; Feng, J.X.; Huang, R.B. Multiple Induced Mutagenesis for Improvement of Ethanol Production by Kluyveromyces marxianus. Biotechnol. Lett. 2010, 32, 1847–1851. [Google Scholar] [CrossRef]
- Yi, S.; Zhang, X.; Li, H.X.; Du, X.X.; Liang, S.W.; Zhao, X.H. Screening and Mutation of Saccharomyces cerevisiae UV-20 with a High Yield of Second Generation Bioethanol and High Tolerance of Temperature, Glucose and Ethanol. Indian J. Microbiol. 2018, 58, 440–447. [Google Scholar] [CrossRef]
- Kang, L.Z.; Han, F.; Lin, J.F.; Guo, L.Q.; Bai, W.F. Breeding of New High-Temperature-Tolerant Strains of Flammulina velutipes. Sci. Hortic. 2013, 151, 97–102. [Google Scholar]
- Hesham, A.E.L.; Wambui, V.; Jo, H.O.; Maina, J.M. Phylogenetic Analysis of Isolated Biofuel Yeasts Based on 5.8S-ITS rDNA and D1/D2 26S rDNA Sequences. J. Genet. Eng. Biotechnol. 2014, 12, 37–43. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T.; Robert, V. Methods for Isolation, Phenotypic Characterization and Maintenance of Yeasts. In The Yeasts, a Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2011; pp. 87–110. [Google Scholar]
- Sun, X.Y.; Zhao, Y.; Liu, L.L.; Jia, B.; Zhao, F.; Huang, W.D.; Zhan, J.C. Copper Tolerance and Biosorption of Saccharomyces cerevisiae During Alcoholic Fermentation. PLoS ONE 2015, 10, e0128611. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Chia, J.Y.; Liu, S.Q. Impact of Addition of Aromatic Amino Acids on Non-Volatile and Volatile Compounds in Lychee Wine Fermented with Saccharomyces cerevisiae Merit.Ferm. Int. J. Food Microbiol. 2014, 170, 12–20. [Google Scholar] [CrossRef]
- Volschenk, H.; Van Vuuren, H.J.J.; Viljoen-Bloom, M. Malic Acid in Wine: Origin, Function and Metabolism During Vinification. S. Afr. J. Enol. Vitic. 2006, 27, 123–136. [Google Scholar] [CrossRef] [Green Version]
- Peinado, R.A.; Mauricio, J.C.; Moreno, J. Aromatic Series in Sherry Wines with Gluconic Acid Subjected to Different Biological Aging Conditions by Saccharomyces cerevisiae Var. Capensis. Food Chem. 2006, 94, 232–239. [Google Scholar] [CrossRef]
- Lelova, Z.; Ivanova-Petropulos, V.; Masar, M.; Lisjak, K.; Bodor, R. Optimization and Validation of a New Capillary Electrophoresis Method with Conductivity Detection for Determination of Small Anions in Red Wines. Food Anal. Methods 2018, 11, 1457–1466. [Google Scholar] [CrossRef]
- Redzepovic, S.; Orlic, S.; Majdak, A.; Kozina, B.; Volschenk, H.; Viljoen-Bloom, M. Differential Malic Acid Degradation by Selected Strains of Saccharomyces During Alcoholic Fermentation. Int. J. Food Microbiol. 2003, 83, 49–61. [Google Scholar] [CrossRef]
- Ferreira, A.M.; Mendes-Faia, A. The Role of Yeasts and Lactic Acid Bacteria on the Metabolism of Organic Acids During Winemaking. Foods 2019, 9, 1231. [Google Scholar] [CrossRef]
- Younis, O.S.; Stewart, G.G. Sugar Uptake and Subsequent Ester and Higher Alcohol Production by Saccharomyces cerevisiae. J. Inst. Brew. 1998, 104, 255–264. [Google Scholar] [CrossRef]
- Lambrechts, M.; Pretorius, I. Yeast and Its Importance to Wine Aroma. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial Modulation of Aromatic Esters in Wine: Current Knowledge and Future Prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Pérez-Martín, F.; Seseña, S.; Izquierdo, P.M.; Palop, M.L. Esterase Activity of Lactic Acid Bacteria Isolated from Malolactic Fermentation of Red Wines. Int. J. Food Microbiol. 2013, 163, 153–158. [Google Scholar] [CrossRef]
- Lilly, M.; Styger, G.; Bauer, F.F.; Lambrechts, M.G.; Pretorius, I.S. The Effect of Increased Branched-Chain Amino Acid Transaminase Activity in Yeast on the Production of Higher Alcohols and on the Flavour Profiles of Wine and Distillates. FEMS Yeast Res. 2006, 6, 726–743. [Google Scholar] [PubMed] [Green Version]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and Bacterial Modulation of Wine Aroma and Flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- De Ovalle, S.; Cavello, I.; Brena, B.M.; Cavalitto, S.; González-Pombo, P. Production and Characterization of a β-Glucosidase from Issatchenkia terricola and Its Use for Hydrolysis of Aromatic Precursors in Cabernet Sauvignon Wine. LWT Food Sci. Technol. 2018, 87, 515–522. [Google Scholar] [CrossRef] [Green Version]
- Mauriello, G.; Capece, A.; D’Auria, M.; Garde-Cerdán, T.; Romano, P. SPME-GC Method as a Tool to Differentiate VOC Profiles in Saccharomyces cerevisiae Wine Yeasts. Food Microbiol. 2009, 26, 246–252. [Google Scholar] [CrossRef]
- Dubourdieu, D.; Torninaga, T.; Masneuf, I.; Des Gachons, C.P.; Murat, M.L. The Role of Yeasts in Grape Flavor Development During Fermentation: The Example of Sauvignon Blanc. Am. J. Enol. Vitic. 2006, 57, 81–88. [Google Scholar]
- Romano, P.; Fiore, C.; Paraggio, M.; Caruso, M.; Capece, A. Function of Yeast Species and Strains in Wine Flavour. Int. J. Food Microbiol. 2003, 86, 169–180. [Google Scholar] [CrossRef]
- Renault, P.; Miot-Sertier, C.; Marullo, P.; Hernández-Orte, P.; Lagarrigue, L.; Lonvaud-Funel, A.; Bely, M. Genetic Characterization and Phenotypic Variability in Torulaspora delbrueckii Species: Potential Applications in the Wine Industry. Int. J. Food Microbiol. 2009, 134, 201–210. [Google Scholar] [CrossRef]
- Clemente-Jiménez, J.M.; Mingorance-Cazorla, L.; Martinez-Rodríguez, S.; Las Heras-Vázquez, F.J.; Rodríguez-Vico, F. Molecular Characterization and Oenological Properties of Wine Yeasts Isolated During Spontaneous Fermentation of Six Varieties of Grape Must. Food Microbiol. 2004, 21, 149–155. [Google Scholar] [CrossRef]
- Liu, Y.Z.; Rousseaux, S.; Tourdot-Marechal, R.; Sadoudi, M.; Gougeon, R.; Schmitt-Kopplin, P.; Alexandre, H. Wine Microbiome: A Dynamic World of Microbial Interactions. Crit. Rev. Food Sci. 2017, 57, 856–873. [Google Scholar]
- Bai, F.W.; Chen, L.J.; Zhang, Z.; Anderson, W.A.; Moo-Young, M. Continuous Ethanol Production and Evaluation of Yeast Cell Lysis and Viability Loss under Very High Gravity Medium Conditions. J. Biotechnol. 2004, 110, 287–293. [Google Scholar] [CrossRef]
- Teixeira, M.C.; Raposo, L.R.; Mira, N.P.; Lourenco, A.B.; Sa-Correia, I. Genome-Wide Identification of Saccharomyces cerevisiae Genes Required for Maximal Tolerance to Ethanol. Appl. Environ. Microbiol. 2009, 75, 5761–5772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Archana, K.M.; Ravi, R.; Anu-Appaiah, K.A. Correlation between Ethanol Stress and Cellular Fatty Acid Composition of Alcohol Producing Non-Saccharomyces in Comparison with Saccharomyces cerevisiae by Multivariate Techniques. J. Food Sci. Technol. 2015, 52, 6770–6776. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Lee, S.B.; Jeon, J.Y.; Park, H.D. Development of Air-Blast Dried Non-Saccharomyces Yeast Starter for Improving Quality of Korean Persimmon Wine and Apple Cider. Int. J. Food Microbiol. 2019, 290, 193–204. [Google Scholar] [PubMed]
- Pérez-Torrado, R.; Querol, A.; Guillamón, J.M. Genetic Improvement of non-GMO Wine Yeasts: Strategies, Advantages and Safety. Trends Food Sci. Technol. 2015, 45, 1–11. [Google Scholar] [CrossRef]
- Farahnak, F.; Seki, T.; Ryu, D.D.Y.; Ogrydziak, D. Construction of Lactose-Assimilating and High-Ethanol-Producing Yeasts by Protoplast Fusion. Appl. Environ. Microbiol. 1986, 51, 362–367. [Google Scholar] [CrossRef] [Green Version]
- Pattanakittivorakul, S.; Lertwattanasakul, N.; Yamada, M.; Limtong, S. Selection of Thermotolerant Saccharomyces cerevisiae for High Temperature Ethanol Production from Molasses and Increasing Ethanol Production by Strain Improvement. Antonie Leeuwenhoek 2019, 112, 975–990. [Google Scholar] [CrossRef]
- Watanabe, T.; Watanabe, I.; Yamamoto, M.; Ando, A.; Nakamura, T. A UV-Induced Mutant of Pichia stipitis with Increased Ethanol Production from Xylose and Selection of a Spontaneous Mutant with Increased Ethanol Tolerance. Bioresour. Technol. 2011, 102, 1844–1848. [Google Scholar]
- Dong, Y.; Zhang, N.; Lu, J.H.; Lin, F.; Teng, L.R. Improvement and Optimization of the Media of Saccharomyces cerevisiae Strain for High Tolerance and High Yield of Ethanol. Afr. J. Microbiol. Res. 2012, 6, 2357–2366. [Google Scholar]
- Liang, M.H.; Liang, Y.J.; Wu, X.N.; Zhou, S.S.; Jiang, J.G. Mutation Breeding of Saccharomyces cerevisiae with Lower Methanol Content and the Effects of Pectinase, Cellulase and Glycine in Sugar Cane Spirits. J. Sci. Food Agric. 2015, 95, 1949–1955. [Google Scholar] [CrossRef] [PubMed]
- Sridhar, M.; Kiran Sree, N.; Venkateswar, R.L. Effect of UV Radiation on Thermotolerance, Ethanol Tolerance and Osmotolerance of Saccharomyces cerevisiae VS1 and VS3 Strains. Bioresour. Technol. 2002, 83, 199–202. [Google Scholar] [CrossRef]
- Wen, L.K.; Wang, L.F.; Wang, G.Z. Degradation of L-Malic and Critic Acids by Issatchenkia terricola. Food Sci. 2011, 32, 220–223. [Google Scholar]
- Yoshikawa, K.; Tanaka, T.; Furusawa, C.; Nagahisa, K.; Hirasawa, T.; Shimizu, H. Comprehensive Phenotypic Analysis for Identification of Genes Affecting Growth under Ethanol Stress in Saccharomyces cerevisiae. FEMS Yeast Res. 2009, 9, 32–44. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).