Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food




Abstract

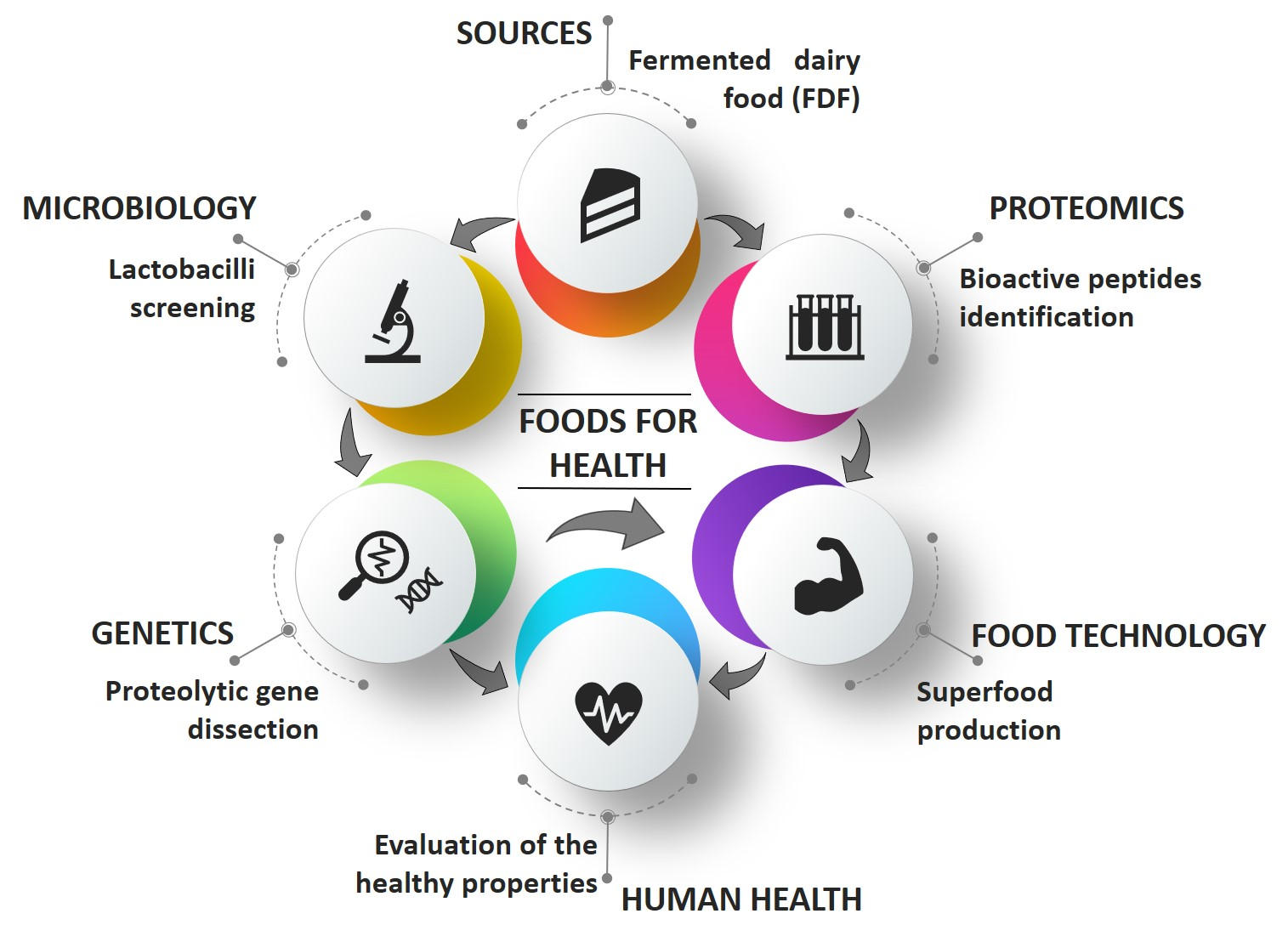
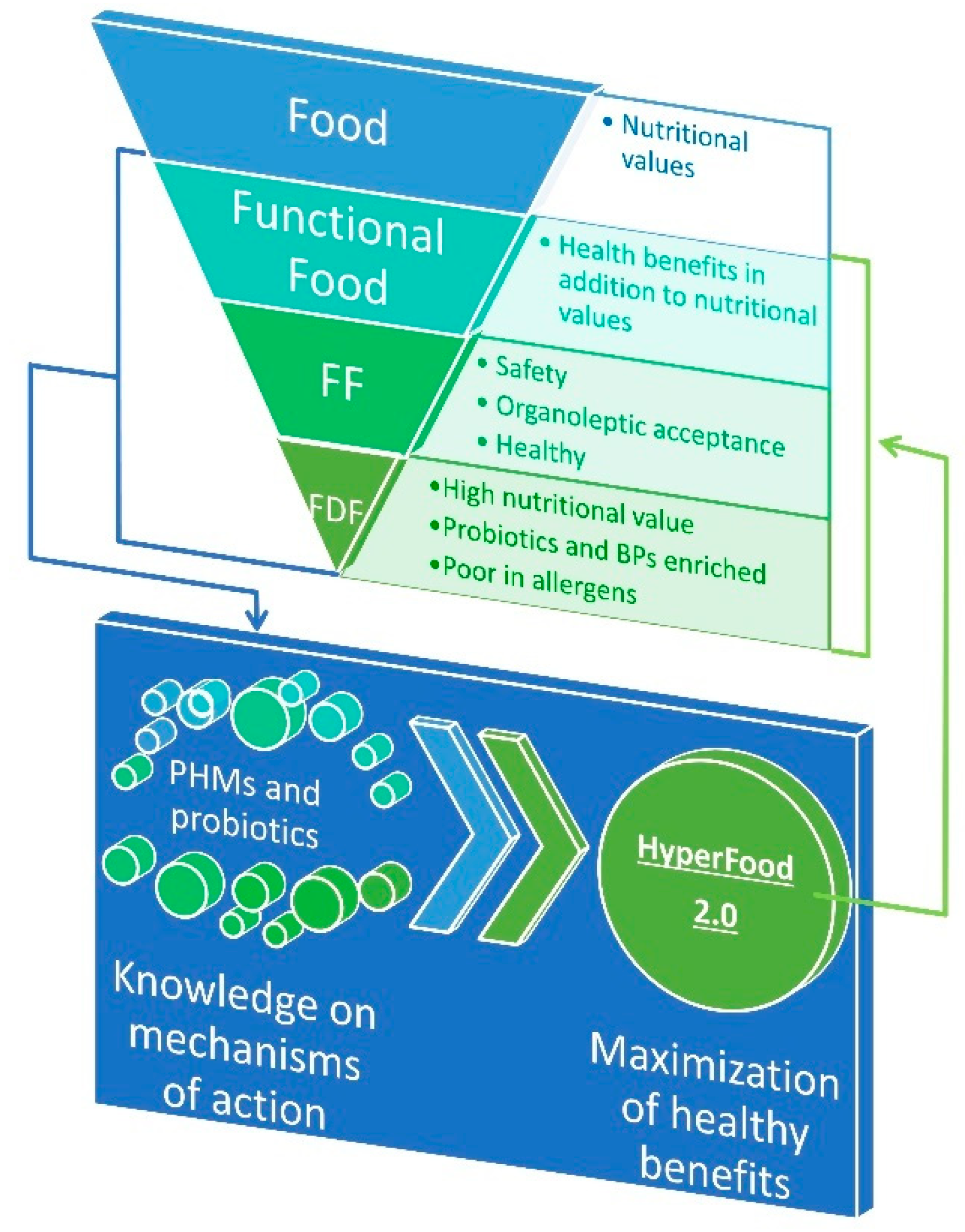
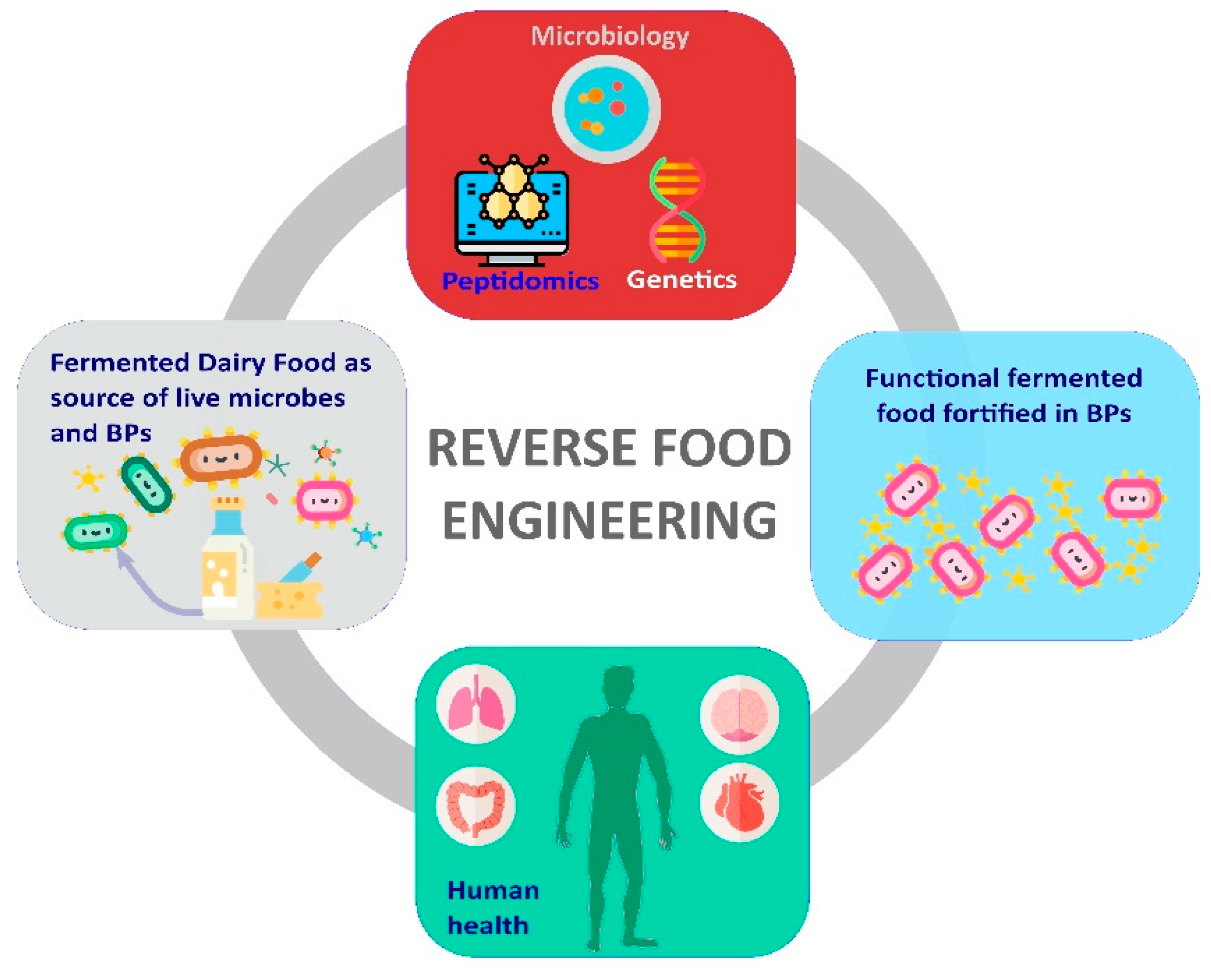
1. Introduction
2. Survey of BPs in FDF
2.1. Anti-Hypertensive Peptides Identified in Commercial FDFs
2.2. Antioxidant Peptides Identified in Commercial Fermented Dairy Products
2.3. Anti-microbial Peptides Identified in Commercial Fermented Dairy Products
2.4. Anti-diabetic Peptides Identified in Commercial Fermented Dairy Products
3. FDF as Source of BPs-Producing LAB
3.1. FDF Microbiota under the Lens of Metagenomics and Genomics
3.2. The Renaissance of Interest Towards Artisanal FDF
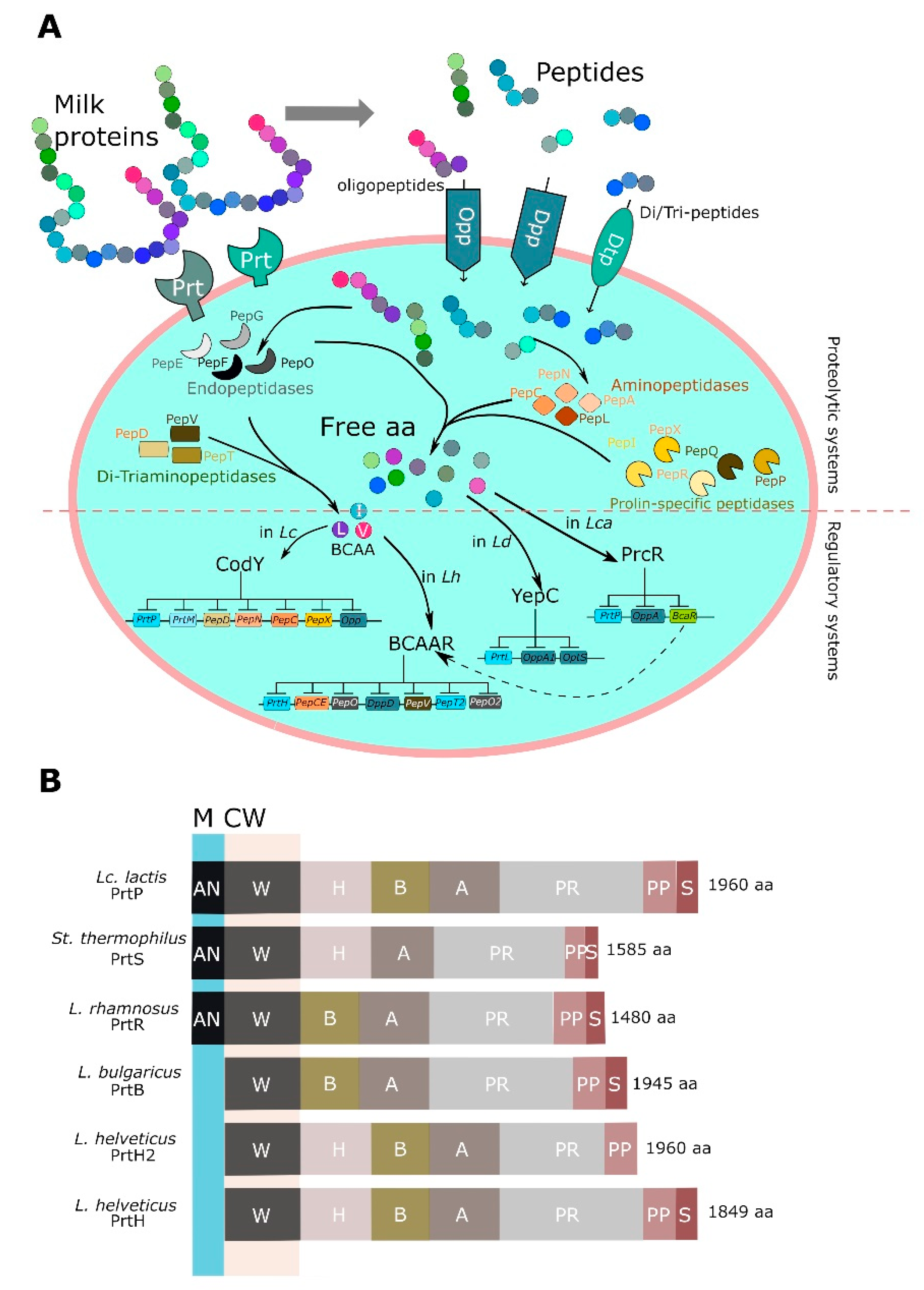
4. LAB Proteolytic System
4.1. Proteases
4.2. Transport System and Peptidases
4.3. Regulation of Proteolytic Systems
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. WHO Action Plan for Implementation of the European Strategy for the Prevention and Control of Noncommunicable Diseases, 2012–2016. World Health Organization, Regional Office for Europe, 2012. Available online: http://www.euro.who.int/__data/assets/pdf_file/0019/170155/e96638.pdf (accessed on 1 September 2019).
- Carruba, G.; Cocciadiferro, L.; Di Cristina, A.; Granata, O.M.; Dolcemascolo, C.; Campisi, I.; Zarcone, M.; Cinquegrani, M.; Traina, A. Nutrition, aging and cancer: Lessons from dietary intervention studies. Immun. Ageing 2016, 13, 13. [Google Scholar] [CrossRef]
- Tilman, D.; Clark, M. Global diets link environmental sustainability and human health. Nature 2014, 515, 518–522. [Google Scholar] [CrossRef]
- Martirosyan, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Food Health Dis. 2015, 5, 209–223. [Google Scholar]
- Tuomilehto, J.; Tiss, A.; Tsang, C.; Khadir, A.; Bahorun, T.; Alkhatib, A.; Arefanian, H.; Barake, R. Functional foods and lifestyle approaches for diabetes prevention and management. Nutrients 2017, 9, 1310. [Google Scholar]
- Battino, M.; Forbes-Hernández, T.Y.; Gasparrini, M.; Afrin, S.; Cianciosi, D.; Zhang, J.; Manna, P.P.; Reboredo-Rodríguez, P.; Varela Lopez, A.; Quiles, J.L.; et al. Relevance of functional foods in the Mediterranean diet: The role of olive oil, berries and honey in the prevention of cancer and cardiovascular diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 893–920. [Google Scholar] [CrossRef] [PubMed]
- Veselkov, K.; Gonzalez, G.; Aljifri, S.; Galea, D.; Mirnezami, R.; Youssef, J.; Bronstein, M.; Laponogov, I. HyperFoods: Machine intelligent mapping of cancer-beating molecules in foods. Sci. Rep. 2019, 9, 9237. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Douglas, G.L.; Klaenhammer, T.R. Genomic evolution of domesticated microorganisms. Annu. Rev. Food Sci. Technol. 2010, 1, 397–414. [Google Scholar] [CrossRef] [PubMed]
- Campbell-Platt, G. Fermented foods—A world perspective. Food Res. Int. 1994, 27, 253–257. [Google Scholar] [CrossRef]
- Metchnikoff, E. The Prolongation of Life; Putnam’s Sons: New York, NY, USA; London, UK, 1908. [Google Scholar]
- van Hylckama, V.J.E.; Veiga, P.; Zhang, C.; Derrien, M.; Zhao, L. Impact of microbial transformation of food on health—From fermented foods to fermentation in the gastro-intestinal tract. Curr. Opin. Biotechnol. 2011, 22, 211–219. [Google Scholar] [CrossRef]
- Shiby, V.K.; Mishra, H.N. Fermented milks and milk products as functional foods—A Review. Crit. Rev. Food Sci. Nutr. 2013, 53, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D. Changes in diet and lifestyle and long-term weight gain in women and men. N. Engl. J. Med. 2011, 364, 2392–2404. [Google Scholar] [CrossRef] [PubMed]
- Keast, D.R.; Hill Gallant, K.M.; Albertson, A.M.; Gugger, C.K.; Holschuh, N.M. Associations between yogurt, dairy, calcium, and vitamin D intake and obesity among U.S. children aged 8–18 years: NHANES, 2005–2008. Nutrients 2015, 7, 1577–1593. [Google Scholar] [CrossRef] [PubMed]
- Pothuraju, R.; Sharma, R.K.; Chagalamarri, J.; Kavadi, P.K.; Jangra, S. Influence of milk fermented with Lactobacillus rhamnosus NCDC 17 alone and in combination with herbal ingredients on diet induced adiposity and related gene expression in C57BL/6J mice. Food Funct. 2015, 6, 3576–3584. [Google Scholar] [CrossRef] [PubMed]
- Panahi, S.; Tremblay, A. The potential role of yogurt in weight management and prevention of type 2 diabetes. J. Am. Coll. Nutr. 2016, 35, 717–731. [Google Scholar] [CrossRef]
- Chen, G.C.; Szeto, I.M.Y.; Chen, L.H.; Han, S.F.; Li, Y.J.; van Hekezen, R.; Qin, L.Q. Dairy products consumption and metabolic syndrome in adults: Systematic review and meta-analysis of observational studies. Sci. Rep. 2015, 5, 14606. [Google Scholar] [CrossRef]
- Capel, F.; Bongard, V.; Malpuech-Brugère, C.; Karoly, E.; Michelotti, G.A.; Rigaudière, J.P.; Jouve, C.; Ferrières, J.; Marmonier, C.; Sébédio, J.L. Metabolomics reveals plausible interactive effects between dairy product consumption and metabolic syndrome in humans. Clin. Nutr. 2019. [Google Scholar] [CrossRef]
- Engberink, M.F.; Hendriksen, M.A.H.; Schouten, E.G.; van Rooij, F.J.; Hofman, A.; Witteman, J.C.; Geleijnse, J.M. Inverse association between dairy intake and hypertension: The Rotterdam study. Am. J. Clin. Nutr. 2009, 89, 1877–1883. [Google Scholar] [CrossRef]
- Huang, W.Y.; Davidge, S.T.; Wu, J. Bioactive natural constituents from food sources-potential use in hypertension prevention and treatment. Crit. Rev. Food Sci. Nutr. 2013, 53, 615–630. [Google Scholar] [CrossRef]
- Fekete, A.A.; Givens, D.I.; Lovegrove, J.A. Casein-derived lactotripeptides reduce systolic and diastolic blood pressure in a meta-analysis of randomised clinical trials. Nutrients 2015, 7, 659–681. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Sun, Q.; Giovannucci, E.; Mozaffarian, D.; Manson, J.E.; Willett, W.C.; Hu, F.B. Dairy consumption and risk of type 2 diabetes: 3 cohorts of US adults and an updated meta-analysis. BMC Med. 2014, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Díaz-López, A.; Bulló, M.; Martínez-González, M.A.; Corella, D.; Estruch, R.; Fitó, M.; Gómez-Gracia, E.; Fiol, M.; García de la Corte, F.J.; Ros, E.; et al. Dairy product consumption and risk of type 2 diabetes in an elderly Spanish Mediterranean population at high cardiovascular risk. Eur. J. Nutr. 2016, 55, 349–360. [Google Scholar]
- Barengolts, E.; Smith, E.D.; Reutrakul, S.; Tonucci, L.; Anothaisintawee, T. The effect of probiotic yogurt on glycemic control in type 2 diabetes or obesity: A meta-analysis of nine randomized controlled trials. Nutrients 2019, 11, E671. [Google Scholar] [CrossRef]
- Cavero-Redondo, I.; Alvarez-Bueno, C.; Sotos-Prieto, M.; Gil, A.; Martinez-Vizcaino, V.; Ruiz, J.R. Milk and Dairy Product consumption and risk of mortality: An overview of systematic reviews and meta-analyses. Adv. Nutr. 2019, 10 (Suppl. 2), S97–S104. [Google Scholar] [CrossRef]
- Thorning, T.K.; Bertram, H.C.; Bonjour, J.P.; de Groot, L.; Dupont, D.; Feeney, E.; Ipsen, R.; Lecerf, J.M.; Mackie, A.; McKinley, M.C.; et al. Whole dairy matrix or single nutrients in assessment of health effects: Current evidence and knowledge gaps. Am. J. Clin. Nutr. 2017, 105, 1033–1045. [Google Scholar] [CrossRef]
- Lebeer, S.; Bron, P.A.; Marco, M.L.; Van Pijkeren, J.P.; O’Connell Motherway, M.; Hill, C.; Pot, B.; Roos, S.; Klaenhammer, T. Identification of probiotic effector molecules: Present state and future perspectives. Curr. Opin. Biotechnol. 2018, 49, 217–223. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Binda, S.; Bron, P.A.; Gross, G.; Hill, C.; van Hylckama Vlieg, J.E.T.; Lebeer, S.; Satokari, R.; Ouwehand, A.C. Understanding mode of action can drive the translational pipeline towards more reliable health benefits for probiotics. Curr. Opin. Biotechnol. 2019, 56, 55–60. [Google Scholar] [CrossRef]
- Bu, G.; Luo, Y.; Chen, F.; Liu, K.; Zhu, T. Milk processing as a tool to reduce cow’s milk allergenicity: A mini-review. Dairy Sci. Technol. 2013, 93, 211–223. [Google Scholar] [CrossRef]
- Gobbetti, M.; Minervini, F.; Rizzello, C.G. Bioactive peptides in dairy products. In Handbook of Food Products Manufacturing; Hui, Y.H., Ed.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2007; pp. 489–517. [Google Scholar]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef]
- Castro, J.M.; Tornadijo, M.E.; Fresno, J.M.; Sandoval, H. Biocheese: A food probiotic carrier. Biomed Res. Int. 2015, 2015, 723056. [Google Scholar] [CrossRef] [PubMed]
- Şanlier, N.; GÖkcen, B.B.; Sezgİn, A.C. Health Benefits of Fermented Foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, F.; Pellegrino, L.; Verduci, E.; Ghiselli, A.; Bernabei, R.; Calvani, R.; Cetin, I.; Giampietro, M.; Perticone, F.; Piretta, L.; et al. Cow’s Milk Consumption and Health: A Health Professional’s Guide. J. Am. Coll. Nutr. 2019, 38, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef] [PubMed]
- Yahfoufi, N.; Mallet, J.F.; Graham, E.; Matar, C. Role of probiotics and prebiotics in immunomodulation. Curr. Opin. Food Sci. 2018, 20, 82–91. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharm. 2017, 174, 1378–1394. [Google Scholar] [CrossRef]
- Vermeirssen, V.; Van, C.J.; Verstraete, W. Bioavailability of angiotensin I converting enzyme inhibitory peptides. Br. J. Nutr. 2004, 92, 357–366. [Google Scholar] [CrossRef]
- Xu, Q.; Hong, H.; Wu, J.; Yan, X. Bioavailability of bioactive peptides derived from food proteins across the intestinal epithelial membrane: A review. Trends Food Sci. Technol. 2019, 86, 399–411. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Fogliano, V. Food matrix interaction and bioavailability of bioactive peptides: Two faces of the same coin? J. Funct. Foods 2017, 35, 9–12. [Google Scholar] [CrossRef]
- Caira, S.; Pinto, G.; Vitaglione, P.; Dal Piaz, F.; Ferranti, P.; Addeo, F. Identification of casein peptides in plasma of subjects after a cheese-enriched diet. Food Res. Int. 2016, 84, 108–112. [Google Scholar] [CrossRef]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Diminiczak, A.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef] [PubMed]
- Borghi, C.; Cicero, A.F.G. Nutraceuticals with clinically detectable blood pressure lowering effect: A review of available randomized clinical trials and their meta-analyses. Br. J. Clin. Pharm. 2017, 83, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Fekete, A.A.; Givens, D.I.; Lovegrove, J.A. The impact of milk proteins and peptides on blood pressure and function: A review of evidence from human intervention studies. Nutr. Res. Rev. 2013, 26, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, R.J.; Murray, B.A. Bioactive peptides and lactic fermentations. Int. J. Dairy Technol. 2006, 59, 118–125. [Google Scholar] [CrossRef]
- Acharya, K.R.; Sturrock, E.D.; Riordan, J.F.; Ehlers, M.R.W. ACE revisited a new target for structure-based drug design. Nat. Rev. 2013, 2, 891–902. [Google Scholar] [CrossRef]
- Coates, D. The angiotensin converting enzyme (ACE). Int. J. Biochem. Cell. Biol. 2003, 35, 769–773. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Gerocarni, B.; Laghi, L.; Borghi, C. Blood pressure lowering effect of lactotripeptides assumed as functional foods: A meta-analysis of current available clinical trials. J. Hum. Hypertens. 2011, 25, 425–436. [Google Scholar] [CrossRef]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T. Anti-hypertensive effect of sour milk and peptides isolated from it that are inhibitors to angiotensin I-converting enzyme. J. Dairy. Sci. 1995, 78, 1253–1257. [Google Scholar] [CrossRef]
- Basiricò, L.; Catalani, E.; Morera, P.; Cattaneo, S.; Stuknyte, M.; Bernabucci, U.; De Noni, I.; Nardone, A. Release of angiotensin converting enzyme-inhibitor peptides during in vitro gastrointestinal digestion of Parmigiano Reggiano PDO cheese and their absorption through an in vitro model of intestinal epithelium. J. Dairy Sci. 2015, 98, 7595–7601. [Google Scholar] [CrossRef]
- Stuknyte, M.; Cattaneo, S.; Masotti, F.; De Noni, I. Occurrence and fate of ACE-inhibitor peptides in cheeses and in their digestates following in vitro static gastrointestinal digestion. Food Chem. 2015, 168, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Kunda, P.B.; Benavente, F.; Catalá-Clariana, S.; Giménez, E.; Barbosa, J.; Sanz-Nebot, V. Identification of bioactive peptides in a functional yogurt by micro liquid chromatography time-of-flight mass spectrometry assisted by retention time prediction. J. Chromatogr. A 2012, 1229, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Chanson-Rolle, A.; Aubin, F.; Braesco, V.; Hamasaki, T.; Kitakaze, M. Influence of the lactotripeptides Isoleucine–Proline–Proline and Valine–Proline–Proline on systolic blood pressure in Japanese subjects. PLoS ONE 2015, 10, e0142235. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.G.; Aubin, F.; Braesco, V.; Borghi, C. Do the lactotripeptides Isoleucine–Proline–Proline andValine–Proline–Proline reduce systolic blood pressure in European subjects? Am. J. Hypertens. 2013, 26, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Sieber, R.; Bütikofer, U.; Egger, C.; Portmann, R.; Walther, B.; Wechsler, D. ACE-inhibitory activity and ACE-inhibiting peptides in different cheese varieties. Dairy Sci. Tech. 2010, 90, 47–73. [Google Scholar] [CrossRef]
- Lu, Y.; Govindasamy-Lucey, S.; Lucey, J.A. Angiotensin-I-converting enzyme-inhibitory peptides in commercial Wisconsin Cheddar cheeses of different ages. J. Dairy Sci. 2016, 99, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, N.; Maeno, M.; Takano, T. Purification and characterization of an anti-hypertensive peptide from a yogurt-like product fermented by Lactobacillus helveticus CPN4. J. Dairy Sci. 1999, 82, 1388–1393. [Google Scholar] [CrossRef]
- Maeno, M.; Yamamoto, N.; Takano, T. ldentification of an anti-hypertensive peptide from casein hydrolysate produced by a proteinase from Lactobacillus helveticus CP790. J. Dairy Sci. 1996, 79, 1316–1321. [Google Scholar] [CrossRef]
- Miguel, M.; Recio, I.; Ramos, M.; Delgado, M.A.; Aleixandre, M.A. Antihypertensive effect of peptides obtained from Enterococcus faecalis-fermented milk in rats. J. Dairy Sci. 2006, 89, 3352–3359. [Google Scholar] [CrossRef]
- Contreras, M.M.; Carrón, R.; Montero, M.J.; Ramos, M.; Recio, I. Novel casein-derived peptides with antihypertensive activity. Int. Dairy J. 2009, 19, 566–573. [Google Scholar] [CrossRef]
- Contreras, M.M.; Sanchez, D.; Sevilla, M.A.; Recio, I.; Amigo, L. Resistance of casein-derived bioactive peptides to simulated gastrointestinal digestion. Int. Dairy J. 2013, 32, 71–78. [Google Scholar] [CrossRef]
- Rival, S.G.; Boeriu, C.G.; Wichers, H.J. Caseins and casein hydrolysates. 2. Antioxidative properties and relevance to lipoxygenase inhibition. J. Agr. Food Chem. 2001, 49, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.; Gómez-Ruiz, J.A.; Recio, I.; Aleixandre, A. Changes in arterial blood pressure after single oral administration of milk-casein-derived peptides in spontaneously hypertensive rats. Mol. Nutr. Food Res. 2010, 54, 1422–1427. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rivera, L.; Ferreira Santos, P.; Miralles, B.; Carrón, R.; Montero, M.J.; Recio, I. Peptide fragments from β-casein f(134–138), HLPLP, generated by the action of rat blood plasma peptidases show potent antihypertensive activity. Food Res. Int. 2016, 88, 348–353. [Google Scholar] [CrossRef]
- Abubakar, A.; Saito, T.; Kitazawa, H.; Kawai, Y.; Itoh, T. Structural analysis of new antihypertensive peptides derived from cheese whey protein by proteinase K digestion. J. Dairy Sci. 1998, 81, 3131–3138. [Google Scholar] [CrossRef]
- Quirós, A.; Hernández-Ledesma, B.; Ramos, M.; Amigo, L.; Recio, I. Angiotensin-converting enzyme inhibitory activity of peptides derived from caprine kefir. J. Dairy Sci. 2005, 88, 3480–3487. [Google Scholar]
- Dallas, D.C.; Citerne, F.; Tian, T.; Silva, V.L.; Kalanetra, K.M.; Frese, S.A.; Robinson, R.C.; Mills, D.A.; Barile, D. Peptidomic analysis reveals proteolytic activity of kefir microorganisms on bovine milk proteins. Food Chem. 2016, 15, 273–284. [Google Scholar] [CrossRef]
- Wakai, T.; Yamamoto, N. Antihypertensive Peptides Specific to Lactobacillus helveticus Fermented Milk. In Biotechnology—Molecular Studies and Novel Applications for Improved Quality of Human Life; Sammour, R., Ed.; InTech: Rijeka, Croatian, 2012; ISBN 978-953-51-0151-2. [Google Scholar]
- Saito, T.; Nakamura, T.; Kitazawa, H.; Kawai, Y.; Itoh, T. Isolation and structural analysis of antihypertensive peptides that exist naturally in Gouda cheese. J. Dairy Sci. 2000, 83, 1434–1440. [Google Scholar] [CrossRef]
- Amorim, F.G.; Coitinho, L.B.; Dias, A.T.; Friques, A.G.F.; Monteiro, B.L.; Rezende, L.C.D.; Pereira, T.M.C.; Campagnaro, B.P.; De Pauw, E.; Vasquez, E.C.; et al. Identification of new bioactive peptides from Kefir milk through proteopeptidomics: Bioprospection of antihypertensive molecules. Food Chem. 2019, 1, 109–119. [Google Scholar] [CrossRef]
- Aluko, R.E. Anti-hypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef]
- Li, S.; Bu, T.; Zheng, J.; Liu, L.; He, G.; Wu, J. Preparation, bioavailability, and mechanism of emerging activities of Ile-Pro-Pro and Val-Pro-Pro. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1097–1110. [Google Scholar] [CrossRef]
- Beltrán-Barrientos, L.M.; Hernández-Mendoza, A.; Torres-Llanez, M.J.; González-Córdova, A.F.; Vallejo-Córdoba, B. Fermented milk as anti-hypertensive functional food. J. Dairy Sci. 2016, 99, 4099–4110. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and characterization of angiotensin I-converting enzyme inhibitors from sour milk. J. Dairy. Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Hebert, E.M.; Mamone, G.; Picariello, G.; Raya, R.R.; Savoy, G.; Ferranti, P.; Addeo, F. Characterization of the pattern of alpha(s1)- and beta-casein breakdown and release of a bioactive peptide by a cell envelope proteinase from Lactobacillus delbrueckii subsp. lactis CRL 581. Appl. Environ. Microbiol. 2008, 74, 3682–3689. [Google Scholar] [CrossRef] [PubMed]
- Solieri, L.; Rutella, G.S.; Tagliazucchi, D. Impact of non-starter lactobacilli on release of peptides with angiotensin-converting enzyme inhibitory and antioxidant activities during bovine milk fermentation. Food Microbiol. 2015, 51, 108–116. [Google Scholar] [CrossRef]
- Rutella, G.S.; Tagliazucchi, D.; Solieri, L. Survival and bioactivities of selected probiotic lactobacilli in yogurt fermentation and cold storage: New insights for developing a bi-functional dairy food. Food Microbiol. 2016, 60, 54–61. [Google Scholar] [CrossRef]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Wechsler, D. Quantification of the angiotensin-converting enzyme inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in hard, semi-hard and soft cheeses. Int. Dairy J. 2007, 17, 968–975. [Google Scholar] [CrossRef]
- Bütikofer, U.; Meyer, J.; Sieber, R.; Walther, B.; Wechsler, D. Occurrence of the angiotensin-converting enzyme–inhibiting tripeptides Val-Pro-Pro and Ile-Pro-Pro in different cheese varieties of Swiss origin. J. Dairy Sci. 2008, 91, 29–38. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Shamsia, S.; Conte, A. Release of angiotensin converting enzyme-inhibitory peptides during in vitro gastro-intestinal digestion of camel milk. Int. Dairy J. 2016, 56, 119–128. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Shamsia, S.; Helal, A.; Conte, A. Angiotensin-converting enzyme inhibitory peptides from goats’ milk released by in vitro gastro-intestinal digestion. Int. Dairy J. 2017, 71, 6–16. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Martini, S.; Shamsia, S.; Helal, A.; Conte, A. Biological activities and peptidomic profile of in vitro-digested cow, camel, goat and sheep milk. Int. Dairy J. 2018, 81, 19–27. [Google Scholar] [CrossRef]
- Rutella, S.G.; Solieri, L.; Martini, S.; Tagliazucchi, D. Release of the antihypertensive tripeptides Valine-Proline-Proline and Isoleucine-Proline-Proline from bovine milk caseins during in vitro gastrointestinal digestion. J. Agric. Food Chem. 2016, 64, 8509–8516. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Y.; Qin, L.Q.; Wang, P.Y.; Li, W.; Chang, C. Effect of milk tripeptides on blood pressure: A meta-analysis of randomized controlled trials. Nutrition 2008, 24, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Miralles, B.; Amigo, L.; Recio, I. Critical review and perspectives on food-derived antihypertensive peptides. J. Agric. Food Chem. 2018, 66, 9384–9390. [Google Scholar] [CrossRef] [PubMed]
- Quirós, A.; Dávalos, A.; Lasunción, M.A.; Ramos, M.; Recio, I. Bioavailability of the antihypertensive peptide LHLPLP: Transepithelial flux of HLPLP. Int. Dairy J. 2008, 18, 279–286. [Google Scholar] [CrossRef]
- Kaiser, S.; Martin, M.; Lunow, D.; Rudolph, S.; Mertten, S.; Möckel, U.; Deußen, A.; Henle, T. Tryptophan-containing dipeptides are bioavailable and inhibit plasma human angiotensin-converting enzyme in vivo. Int. Dairy J. 2016, 52, 107–114. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Technological options for the production of health-promoting proteins and peptides derived from milk and colostrum. Curr. Pharm. Des. 2007, 13, 829–843. [Google Scholar] [CrossRef]
- Halliwell, B. Role of free radicals in the neurodegenerative diseases: Therapeutic implications for antioxidant treatment. Drugs Aging 2001, 18, 479–484. [Google Scholar] [CrossRef]
- Liu, Q.; Raina, A.; Smith, M.; Sayre, L.; Perry, G. Hydroxynonenal, toxic carbonyls, and Alzheimer disease. Mol. Asp. Med. 2003, 24, 305–313. [Google Scholar] [CrossRef]
- Abuja, P.; Albertini, R. Methods for monitoring oxidative stress, lipid peroxidation and oxidation resistance of lipoproteins. Clin. Chim. Acta 2001, 306, 1–17. [Google Scholar] [CrossRef]
- Halliwell, B. Lipid peroxidation, antioxidants and cardiovascular disease: How should we move forward? Cardiov. Res. 2000, 47, 410–418. [Google Scholar] [CrossRef]
- Halliwell, B.; Whiteman, M. Measuring reactive species and oxidative damage in vivo and in cell culture: How should you do it and what do the results mean? Br. J. Pharm. 2004, 142, 231–255. [Google Scholar] [CrossRef] [PubMed]
- Hoelzl, C.; Bichler, J.; Ferk, F.; Simic, T.; Nersesyan, A.; Elbling, L.; Ehrlich, V.; Chakraborty, A.; Knasmüller, S. Methods for the detection of antioxidants which prevent age related diseases: A critical review with particular emphasis on human intervention studies. J. Phys. Pharm. 2005, 56, 49–64. [Google Scholar]
- Collins, A. Antioxidant intervention as a route to cancer prevention. Eur. J. Cancer 2005, 41, 1923–1930. [Google Scholar] [CrossRef] [PubMed]
- Packard, R.R.S.; Libby, P. Inflammation in atherosclerosis: From vascular biology to biomarker discovery and risk prediction. Clin. Chem. 2008, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Inflammation in atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Cichosz, G.; Czeczot, H.; Ambroziak, A.; Bielecka, M.M. Natural antioxidants in milk and dairy products. Int. J. Dairy Technol. 2017, 70, 165–178. [Google Scholar]
- Lindmark-Månsson, H.; Åkesson, B. Antioxidative factors in milk. Br. J. Nutr. 2000, 84, 103–110. [Google Scholar] [CrossRef]
- Park, Y.W. Overview of bioactive components in milk and dairy products. In Bioactive Components in Milk and Dairy Products; Park, Y.W., Ed.; Wiley-Blackwell Publishers: Ames, IA, USA; Oxford, UK, 2009; pp. 3–14. [Google Scholar]
- Pihlanto, A. Lactic fermentation and bioactive peptides. In Lactic Acid Bacteria R & R for Food, Health and Livestock Purposes; IntechOpen: Marcelino, Kongo, 2013. [Google Scholar]
- Hernández-Ledesma, B.; Amigo, L.; Ramos, M.; Recio, I. Application of high-performance liquid chromatography-tandem mass spectrometry to the identification of biologically active peptides produced by milk fermentation and simulated gastrointestinal digestion. J. Chromatogr. 2004, 1049, 107–114. [Google Scholar] [CrossRef]
- Jin, Y.; Yub, Y.; Qi, Y.; Wang, F.; Yan, J.; Zou, H. Peptide profiling and the bioactivity character of yogurt in the simulated gastrointestinal digestion. J. Prot. 2016, 141, 24–46. [Google Scholar] [CrossRef]
- Korhonen, H. Milk-derived bioactive peptides: From science to applications. J. Fun. Foods 2009, 1, 177–187. [Google Scholar] [CrossRef]
- Parrella, A.; Caterino, E.; Cangiano, M.; Criscuolo, E.; Russo, E.; Lavorgna, M.; Isidori, M. Antioxidant properties of different milk fermented with lactic acid bacteria and yeast. Int. J. Food Sci. Tech. 2012, 47, 2493–2502. [Google Scholar] [CrossRef]
- Pattom, S.; Hongsprabhas, P. Effect of coagulants on antioxidant capacity of milk protein curds and their tryptic hydrolysates. J. Food Biochem. 2013, 37, 203–211. [Google Scholar]
- Abadia-Garcia, L.; Cardador, A.; del Campo, M.S.T.; Arvizu, S.M.; Castano-Tostado, E.; Regalado-Gonzalez, C. Influence of probiotic strains added to cottage cheese on generation of potentially antioxidant peptides, antilisterial activity, and survival of probiotic microorganisms in simulated gastrointestinal conditions. Int. Dairy J. 2013, 33, 191–197. [Google Scholar] [CrossRef]
- Kumar, S.; Teotia, U.V.S.; Sanghi, A. Antioxidative property of cow milk caseinates hydrolyzed with different proteases. Int. J. Pharm. Pharm. Sci. 2013, 5, 418–422. [Google Scholar]
- Mushtaq, M.; Gani, A.; Masoodi, F.A.; Ahmad, M. Himalayan cheese (Kalari/Kradi)-Effect of different probiotic strains on oxidative stability, microbiological, sensory and nutraceutical properties during storage. Lwt-Food Sci. Technol. 2016, 67, 74–81. [Google Scholar] [CrossRef]
- Pihlanto, A. Whey proteins and peptides: Emerging properties to promote health. Nutrafoods 2011, 10, 29–42. [Google Scholar] [CrossRef]
- Chang, O.K.; Seol, K.H.; Jeong, S.G.; Oh, M.H.; Park, B.Y.; Perrin, C.; Ham, J.S. Casein hydrolysis by Bifidobacterium longum KACC91563 and antioxidant activities of peptides derived therefrom. J. Dairy Sci. 2013, 96, 5544–5555. [Google Scholar] [CrossRef]
- Power, O.; Jakeman, P.; FitzGerald, R.J. Antioxidative peptides: Enzymatic production, in vitro and in vivo antioxidant activity and potential applications of milk-derived antioxidative peptides. Amino Acids 2013, 44, 797–820. [Google Scholar] [CrossRef]
- Ajibola, C.F.; Fashakin, J.B.; Fagbemi, T.N.; Aluko, R.E. Effect of Peptide Size on Antioxidant Properties of African Yam Bean Seed (Sphenostylis stenocarpa) Protein Hydrolysate Fractions. Int. J. Mol. Sci. 2011, 12, 6685–6702. [Google Scholar] [CrossRef]
- Pihlanto, A. Antioxidative peptides derived from milk proteins. Int. Dairy J. 2006, 16, 1306–1314. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Helal, A.; Verzelloni, E.; Conte, A. Bovine milk antioxidant properties: Effect of in vitro digestion and identification of antioxidant compounds. Dairy Sci. Technol. 2016, 96, 657–676. [Google Scholar] [CrossRef]
- Hernandez-Ledesma, B.; Miralles, B.; Amigo, L.; Ramos, M.; Recio, I. Identification of antioxidant and ACE-inhibitory peptides in fermented milk. J. Sci. Food Agric. 2005, 85, 1041–1048. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Yokoo, Y.; Fujimaki, M. Studies on antioxidative activities of amino compounds on fats and oils. Part II. Antioxidative activities of dipeptides and their synergistic effects on tocopherol. Nippon Shokuhin Kogyo Gakkaishi 1975, 22, 425–430. [Google Scholar] [CrossRef]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Kitts, D.D.; Weiler, K. Bioactive proteins and peptides from food sources. Applications of bioprocesses used in isolation and recovery. Curr. Pharm. Des. 2003, 9, 1309–1323. [Google Scholar] [CrossRef]
- Xiong, Y.L. Antioxidant peptides. In Bioactive Proteins and Peptides as Functional Foods and Neutraceuticals; Mine, Y., Jiang, B., Li-Chan, E., Eds.; Wiley-Blackwell: Ames, IA, USA, 2010; pp. 29–39. [Google Scholar]
- Eisele, T.; Stressler, T.; Kranz, B.; Fischer, L. Bioactive peptides generated in an enzyme membrane reactor using Bacillus lentus alkaline peptidase. Eur. Food Res. Tech. 2013, 236, 483–490. [Google Scholar] [CrossRef]
- Gupta, A.; Mann, B.; Kumar, R.; Sangwan, R.B. Identification of antioxidant peptides in cheddar cheese made with adjunct culture Lactobacillus casei ssp. casei 300. Milchwissenschaft 2010, 65, 396–399. [Google Scholar]
- Rival, S.G.; Fornaroli, S.; Boeriu, C.G.; Wichers, H.J. Caseins and casein hydrolysates. 1. Lipoxygenase inhibitory properties. J. Agric. Food Chem. 2001, 49, 287–294. [Google Scholar] [CrossRef]
- Farvin, K.H.S.; Baron, C.P.; Nielsen, N.S.; Otte, J.; Jacobsen, C. Antioxidant activity of yoghurt peptides: Part 2-characterisation of peptide fractions. Food Chem. 2010, 123, 1090–1097. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Amigo, L.; Ramos, M.; Recio, I. Angiotensin converting enzyme inhibitory activity in commercial fermented products. Formation of peptides under simulated gastrointestinal digestion. J. Agric. Food Chem. 2004, 52, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Taha, S.; El Abd, M.; De Gobba, C.; Abdel-Hamid, M.; Khalil, E.; Hassan, D. Antioxidant and antibacterial activities of bioactive peptides in buffalo’s yoghurt fermented with different starter cultures. Food. Sci. Biotechnol. 2017, 26, 1325–1332. [Google Scholar] [CrossRef] [PubMed]
- Pisanu, S.; Pagnozzi, D.; Pes, M.; Pirisi, A.; Roggio, T.; Uzzau, S.; Addis, M.F. Differences in the peptide profile of raw and pasteurised ovine milk cheese and implications for its bioactive potential. Int. Dairy J. 2015, 42, 26–33. [Google Scholar] [CrossRef]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides-opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef]
- López-Expósito, I.; Quiros, A.; Amigo, L.; Recio, I. Casein hydrolysates as a source of antimicrobial, antioxidant and antihypertensive peptides. Lait 2007, 87, 241–249. [Google Scholar] [CrossRef]
- Kohmura, M.; Nio, N.; Ariyoshi, Y. Inhibition of angiotensin-converting enzyme by synthetic peptide fragments of various beta-caseins. Agric. Biol. Chem. 1990, 54, 1101–1102. [Google Scholar] [CrossRef]
- De Gobba, C.; Tompa, G.; Otte, J. Bioactive peptides from caseins released by cold active proteolytic enzymes from Arsukibacterium ikkense. Food Chem. 2014, 165, 205–215. [Google Scholar] [CrossRef]
- Clare, D.A.; Catignani, G.L.; Swaisgood, H.E. Biodefense properties of milk: The role of antimicrobial proteins and peptides. Curr. Pharm. Des. 2003, 9, 1239–1255. [Google Scholar] [CrossRef]
- FitzGerald, R.J.; Meisel, H. Milk protein hydrolysates and bioactive peptides. In Advanced Dairy Chemistry—1 Proteins; Fox, P.F., Mc Sweeney, P.L.H., Eds.; Springer: Boston, MA, USA, 2003; pp. 675–698. [Google Scholar]
- Li, G.; Le, G.; Shi, Y.; Shrestha, S. Angiotensin I-converting enzyme inhibitory peptides derived from food proteins and their physiological and pharmacological effects. Nutr. Res. 2004, 24, 469–486. [Google Scholar] [CrossRef]
- López-Expósito, I.; Recio, I. Antibacterial activity of peptides and folding variants from milk proteins. Int. Dairy J. 2006, 16, 1294–1305. [Google Scholar] [CrossRef]
- López-Expósito, I.; Amigo, L.; Recio, I. Identification of the initial binding sites of αs2-casein f(183–207) and effect on bacterial membranes and cell morphology. Biochim. Biophys. Acta Biomembr. 2008, 1778, 2444–2449. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- Fontenele, M.A.; Bastos, M.S.R.; Dos Santos, K.M.O.; Bemquerer, M.P.; do Egito, A.S. Peptide profile of Coalho cheese: A contribution for Protected Designation of Origin (PDO). Food Chem. 2017, 15, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Lahov, E.; Regelson, W. Antibacterial and inmunostimulating casein-derived substances from milk: Casecidin, isracidin peptides. Fed. Chem. Toxic. 1996, 34, 131–145. [Google Scholar] [CrossRef]
- López-Expósito, I.; Gómez-Ruiz, J.A.; Amigo, L.; Recio, I. Identification of antibacterial peptides from ovine as2-casein. Int. Dairy J. 2006, 16, 1072–1080. [Google Scholar] [CrossRef]
- López-Expósito, I.; Minervini, F.; Amigo, L.; Recio, I. Identification of antibacterial peptides from bovine kappa-casein. J. Food Prot. 2006, 69, 2992–2997. [Google Scholar] [CrossRef]
- Barbé, F.; Le Feunteun, S.; Rémond, D.; Ménard, O.; Jardin, J.; Henry, G.; Laroche, B.; Dupont, D. Tracking the in vivo release of bioactive peptides in the gut during digestion: Mass spectrometry peptidomic characterization of effluents collected in the gut of dairy matrices fed mini-pigs. Food Res. Int. 2014, 63, 147–156. [Google Scholar]
- Yvon, M.; Pelissier, J.P. Characterization and kinetics of evacuation of peptides resulting from casein hydrolysis in the stomach of the calf. J. Agric. Food Chem. 1987, 35, 148–156. [Google Scholar] [CrossRef]
- Boutrou, R.; Gaudichon, C.; Dupont, D.; Jardin, J.; Airinei, G.; Marsset-Baglieri, A.; Benamouzig, R.; Tomé, D.; Leonil, J. Sequential release of milk protein-derived bioactive peptides in the jejunum in healthy humans. Am. J. Clin. Nutr. 2013, 97, 1314–1323. [Google Scholar] [CrossRef]
- World Health Organization, Diabetes Mellitus Fact Sheet. 2018. Available online: http://www.who.int/news-room/fact-sheets/detail/diabetes (accessed on 1 September 2019).
- Taylor, S.I. Deconstructing type 2 diabetes. Cell 1999, 97, 9–12. [Google Scholar] [CrossRef]
- Nauck, M. Incretin therapies: Highlighting common features and differences in the modes of action of glucagon-like peptide-1 receptor agonists and dipeptidyl peptidase-4 inhibitors. Diabetes Obes. Metab. 2016, 18, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, H.B.; Branner, S.; Wiberg, F.C.; Wagtmann, N. Crystal structure of human dipeptidyl peptidase IV/CD26 in complex with a substrate analog. Nat. Struct. Biol. 2003, 10, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Mentlein, R. Dipeptidyl-peptidase IV (CD26)—Role in the inactivation of regulatory peptides. Regul. Pept. 1999, 85, 9–24. [Google Scholar] [CrossRef]
- Langley, A.K.; Suffoletta, T.J.; Jennings, H.R. Dipeptidyl peptidase IV inhibitors and the incretin system in type 2 diabetes mellitus. Pharmacotherapy 2007, 27, 1163–1180. [Google Scholar] [CrossRef]
- Vilsbøll, T.; Krarup, T.; Madsbad, S.; Holst, J.J. No reactive hypoglycaemia in Type 2 diabetic patients after subcutaneous administration of GLP-1 and intravenous glucose. Diabet. Med. 2001, 18, 144–149. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Evaluation of the potential of dietary Proteins as precursors of dipeptidyl peptidase (DPP)-IV inhibitors by an in silico approach. J. Funct. Foods 2012, 4, 403–422. [Google Scholar] [CrossRef]
- Uenishi, H.; Kabuki, T.; Seto, Y.; Serizawa, A.; Nakajima, H. Isolation and identification of casein-derived dipeptidyl-peptidase 4 (DPP-4)-inhibitory peptide LPQNIPPL from gouda-type cheese and its effect on plasma glucose in rats. Int. Dairy J. 2012, 22, 24–30. [Google Scholar] [CrossRef]
- Li, C.; Kwok, L.-Y.; Mi, Z.; Bala, J.; Xue, J.; Yang, J. Characterization of the angiotensin-converting enzyme inhibitory activity of fermented milks produced with Lactobacillus casei. J. Dairy Sci. 2017, 100, 9495–9507. [Google Scholar] [CrossRef]
- Jakubowicz, D.; Froy, O.; Ahren, B.; Boaz, M.; Landau, Z.; Bar-Dayan, Y.; Ganz, T.; Barnea, M.; Wainstein, J. Incretin, insulinotropic and glucose-lowering effects of whey protein pre-load in type 2 diabetes: A randomized clinical trial. Diabetologia 2014, 57, 1807–1811. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; FitzGerald, R.J. The scientific evidence for the role of milk protein-derived bioactive peptides in humans: A review. J. Funct. Foods 2015, 640, 640–656. [Google Scholar] [CrossRef]
- Montel, M.C.; Buchin, S.; Mallet, A.; Delbés-Paus, C.; Vuitton, D.A.; Desmasures, N.; Berthier, F. Traditional cheeses: Rich and diverse microbiota with associated benefits. Int. J. Food Microbiol. 2014, 177, 136–154. [Google Scholar] [CrossRef] [PubMed]
- EFSA—European Food Safety Authority. Update of the list of QPS-recommended biological agents intentionally added to food or feed as notified to EFSA 4: Suitability of taxonomic units notified to EFSA until March 2016. EFSA J. 2016, 14, 4522. [Google Scholar]
- Carminati, D.; Giraffa, G.; Zago, M.; Marcó, M.B.; Guglielmotti, D.; Binetti, A.; Reinheimer, J. Lactic acid bacteria for dairy fermentations. In Biotechnology of Lactic Acid Bacteria: Novel Applications, 2nd ed.; Mozzi, F., Raya, R.R., Vignolo, G.M., Eds.; John Wiley & Sons, Ltd.: New York, NY, USA, 2015; pp. 191–208. [Google Scholar]
- Cogan, T.M.; Beresford, T.P.; Steele, J.; Broadbent, J.; Shah, N.P.; Ustunol, Z. Invited review: Advances in starter cultures and cultured foods. J. Dairy Sci. 2007, 90, 4005–4021. [Google Scholar] [CrossRef] [PubMed]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 1–19. [Google Scholar] [CrossRef]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial succession and flavour production in the fermented dairy beverage kefir. mSystems 2016, 1, e00052-16. [Google Scholar] [CrossRef]
- Macori, G.; Cotter, P.D. Novel insights into the microbiology of fermented dairy foods. Curr. Opin. Biotechnol. 2018, 49, 172–178. [Google Scholar] [CrossRef]
- Solieri, L.; Dakal, T.C.; Giudici, P. Next-generation sequencing and its potential impact on food microbial genomics. Ann. Microbiol. 2013, 63, 21–37. [Google Scholar] [CrossRef]
- De Filippis, F.; Parente, E.; Ercolini, D. Metagenomics insights into food fermentations. Microb. Biotechnol. 2017, 10, 91–102. [Google Scholar] [CrossRef]
- Yeluri Jonnala, B.R.; McSweeney, P.L.H.; Sheehan, J.J.; Cotter, P.D. Sequencing of the cheese microbiome and its relevance to industry. Front Microbiol. 2018, 9, 1020. [Google Scholar] [CrossRef]
- Gobbetti, M.; Neviani, E.; Calasso, M.; De Angelis, M.; Fox, F.P.; Di Cagno, R. Drivers that establish and assembly the lactic acid bacteria biota in cheeses. Trends Food Sci. Technol. 2018, 78, 244–254. [Google Scholar] [CrossRef]
- Makarova, K.; Slesarev, A.; Wolf, Y.; Sorokin, A.; Mirkin, B.; Koonin, E.; Pavlov, A.; Pavlova, N.; Karamychev, V.; Polouchine, N.; et al. Comparative genomics of the lactic acid bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 15611–15616. [Google Scholar] [CrossRef] [PubMed]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Abushelaibi, A.; Al-Mahadin, S.; El-Tarabily, K.; Shah, N.P.; Ayyash, M. Characterization of potential probiotic lactic acid bacteria isolated from camel milk. Lebensm. Wiss. Technol. 2017, 79, 316–325. [Google Scholar] [CrossRef]
- Makete, G.; Aiyegoro, O.A.; Thantsha, M.S. Isolation, identification and screening of potential probiotic bacteria in milk from South African Saanen goats. Probiotics Antimicrob. Proteins 2017, 9, 246–254. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Kianirad, M. Antioxidant activity of camel and bovine milk fermented by lactic acid bacteria isolated from traditional fermented camel milk (Chal). Dairy Sci. Technol. 2016, 96, 443–457. [Google Scholar] [CrossRef]
- Solieri, L.; Bianchi, A.; Giudici, P. Inventory of non starter lactic acid bacteria from ripened Parmigiano Reggiano cheese as assessed by a culture dependent multiphasic approach. Syst. Appl. Microbiol. 2012, 35, 270–277. [Google Scholar] [CrossRef]
- Pino, A.; Russo, N.; Van Hoorde, K.; De Angelis, M.; Sferrazzo, G.; Randazzo, C.L.; Caggia, C. Piacentinu Ennese PDO Cheese as reservoir of promising probiotic bacteria. Microorganisms 2019, 7, 254. [Google Scholar] [CrossRef]
- Pino, A.; Van Hoorde, K.; Pitino, I.; Russo, N.; Carpino, S.; Caggia, C.; Randazzo, C.L. Survival of potential probiotic lactobacilli used as adjunct cultures on Pecorino Siciliano cheese ripening and passage through the gastrointestinal tract of healthy volunteers. Int. J. Food Microbiol. 2017, 252, 42–52. [Google Scholar] [CrossRef]
- Zoumpopoulou, G.; Tzouvanou, A.; Mavrogonatou, E.; Alexandraki, V.; Georgalaki, M.; Anastasiou, R.; Papadelli, M.; Manolopoulou, E.; Kazou, M.; Kletsas, D.; et al. Probiotic features of lactic acid bacteria isolated from a diverse pool of traditional Greek dairy products regarding specific strain-host interactions. Prob. Antimicrob. Proteins. 2018, 10, 313–322. [Google Scholar] [CrossRef]
- Ołdak, A.; Zielińska, D.; Łepecka, A.; Długosz, E.; Kołożyn-Krajewska, D. Lactobacillus plantarum strains isolated from Polish regional cheeses exhibit anti-staphylococcal activity and selected probiotic properties. Probiotics Antimicrob. Proteins. 2019. [Epub ahead of print]. [Google Scholar] [CrossRef] [PubMed]
- Moyano, S.; Gonçalves dos Santos, M.T.; Galván, A.I.; Merchán, A.; González, E.; de Guía Córdoba, M.; Benito, M.J. Screening of autochthonous lactic acid bacteria strains from artisanal soft cheese: Probiotic characteristics and prebiotic metabolism. LWT-Food Sci. Technol. 2019, 114, 108388. [Google Scholar] [CrossRef]
- Reid, G.; Gadir, A.A.; Dhir, R. Probiotics: Reiterating what they are and what they are not. Front. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Benson, A.; Lebeer, S.; Merenstein, D.J.; Klaenhammer, T.R. Shared mechanisms among probiotic taxa: Implications for general probiotic claims. Curr. Opin. Biotechnol. 2018, 49, 207–216. [Google Scholar] [CrossRef]
- Linares, D.M.; Gómez, C.; Renes, E.; Fresno, J.M.; Tornadijo, M.E.; Ross, R.P.; Stanton, C. Lactic acid bacteria and bifidobacteria with potential to design natural biofunctional health-promoting dairy foods. Front. Microbiol. 2018, 8, 846. [Google Scholar] [CrossRef]
- Quan, S.; Tsuda, H.; Miyamoto, T. Angiotensin I-converting enzyme inhibitory peptides in skim milk fermented with Lactobacillus helveticus 130B4 from camel milk in inner Mongolia, China. J. Sci. Food Agric. 2018, 88, 2688–2692. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, C.R.; Machado, J.; Correia, S.; McCartney, A.L.; Elmore, J.S.; Jauregi, P. Highly proteolytic bacteria from semi-ripened Chiapas cheese elicit angiotensin-I converting enzyme inhibition and antioxidant activity. LWT-Food Sci. Technol. 2019, 111, 449–456. [Google Scholar] [CrossRef]
- Yamamoto, N.; Kawakami, N.; Ishida, Y.; Yada, H. Lactobacillus helveticus Bacterium Having High Capability of Producing Tripeptides, Fermented Milk Product, and Process for Preparing the Same. U.S. patent 09/508840, 18 March 2003. [Google Scholar]
- Chen, Y.; Liu, W.; Xue, J.; Yang, J.; Chen, X.; Shao, Y.; Kwok, L.Y.; Bilige, M.; Mang, L.; Zhang, H. Angiotensin-converting enzyme inhibitory activity of Lactobacillus helveticus strains from traditional fermented dairy foods and antihypertensive effect of fermented milk of strain H9. J. Dairy Sci. 2014, 97, 6680–6692. [Google Scholar] [CrossRef]
- Skrzypczak, K.; Gustaw, W.; Szwajgier, D.; Fornal, E.; Waśko, A. κ-Casein as a source of short-chain bioactive peptides generated by Lactobacillus helveticus. J. Food Sci. Technol. 2017, 54, 3679–3688. [Google Scholar] [CrossRef]
- Elfahri, K.; Donkor, O.; Vasiljevic, T. Potential of novel Lactobacillus helveticus strains and their cell wall bound proteases to release physiologically active peptides from milk proteins. Int. Dairy J. 2014, 38, 37–46. [Google Scholar] [CrossRef]
- Jensen, M.P.; Vogensen, F.K.; Ardö, Y. Variation in caseinolytic properties of six cheese related Lactobacillus helveticus strains. Int. Dairy J. 2009, 19, 661–668. [Google Scholar] [CrossRef]
- Hati, S.; Sakure, A.; Mandal, S. Impact of proteolytic Lactobacillus helveticus MTCC5463 on production of bioactive peptides derived from honey based fermented milk international. J. Pep. Res. Ther. 2017, 23, 297–303. [Google Scholar] [CrossRef]
- Atanasova, J.; Moncheva, P.; Ivanova, I. Proteolytic and antimicrobial activity of lactic acid bacteria grown in goat milk. Biotechnol. Equip. 2014, 28, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cui, L.; Chen, W.; Zhang, H. An application in Gouda cheese manufacture for a strain of Lactobacillus helveticus ND01. Int. J. Dairy Technol. 2011, 64, 386–393. [Google Scholar] [CrossRef]
- Minervini, F.; Algaron, F.; Rizzello, C.G.; Fox, P.F.; Monnet, V.; Gobbetti, M. Angiotensin I-converting-enzyme-inhibitory and antibacterial peptides from Lactobacillus helveticus PR4 proteinase-hydrolyzed caseins of milk from six species. Appl. Environ. Microbiol. 2003, 69, 5297–5305. [Google Scholar] [CrossRef]
- Beganović, J.; Kos, B.; Pavunc, A.L.; Uroić, K.; Džidara, P.; Šušković, J. Proteolytic activity of probiotic strain Lactobacillus helveticus M92. Anaerobe 2013, 20, 58–64. [Google Scholar]
- Wang, J.H.K.; Dong, C.; Chen, Y.F.; Cui, L.M.; Zhang, H. A new probiotic Cheddar cheese with high ACE-inhibitory activity and γ-aminobutyric acid content produced with koumiss-derived Lactobacillus casei Zhang. Food Technol. Biotechnol. 2010, 48, 62–70. [Google Scholar]
- Ahn, J.E.; Park, S.Y.; Atwal, A.; Gibbs, B.F.; Lee, B.H. Angiotensin i-converting enzyme (ACE) inhibitory peptides from whey fermented by Lactobacillus species. J. Food Biochem. 2009, 33, 587–602. [Google Scholar] [CrossRef]
- Abdel-Hamid, M.; Romeih, A.; Gamba, R.R.; Nagai, E.; Suzuki, T.; Koyanagi, T.; Enomoto, T. The biological activity of fermented milk produced by Lactobacillus casei ATCC 393 during cold storage. Int. Dairy J. 2019, 91, 1–8. [Google Scholar] [CrossRef]
- Sah, B.N.P.; Vasiljevic, T.; McKechnie, S.; Donkor, O.N. Effect of probiotics on antioxidant and antimutagenic activities of crude peptide extract from yogurt. Food Chem. 2014, 156, 264–270. [Google Scholar] [CrossRef]
- Didelot, S.; Bordenave-Juchereau, S.; Rosenfeld, E.; Piot, J.M.; Sannier, F. Peptides released from acid goat whey by a yeast-lactobacillus association isolated from cheese microflora. J. Dairy Res. 2006, 73, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Solanki, D.; Hati, S. Considering the potential of Lactobacillus rhamnosus for producing Angiotensin I-Converting Enzyme (ACE) inhibitory peptides in fermented camel milk (Indian breed). Food Biosci. 2018, 23, 16–22. [Google Scholar] [CrossRef]
- Moslehishad, M.; Ehsani, M.R.; Salami, M.; Mirdamadi, S.; Ezzatpanah, A.; Naslaji, A.N.; Moosavi-Movahedi, A.A. The comparative assessment of ACE-inhibitory and antioxidant activities of peptide fractions obtained from fermented camel and bovine milk by Lactobacillus rhamnosus PTCC 1637. Int. Dairy J. 2013, 29, 82–87. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Santiago-López, L.; Peres, C.M.; Peres, C.; Garcia, H.S.; Vallejo-Cordoba, B. Assessment of multifunctional activity of bioactive peptides derived from fermented milk by specific Lactobacillus plantarum strains. J. Dairy Sci. 2017, 100, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Al-Dhaheri, A.S.; Al Mahadin, S.; Kizhakkayil, J.; Abushelaibi, A. In vitro investigation of anticancer, antihypertensive, antidiabetic, and antioxidant activities of camel milk fermented with camel milk probiotic: A comparative study with fermented bovine milk. J. Dairy Sci. 2018, 101, 900–911. [Google Scholar] [CrossRef]
- Al-Dhaheri, A.S.; Al-Hemeiri, R.; Kizhakkayil, J.; Al-Nabulsi, A.; Abushelaibi, A.; Shah, N.P.; Ayyash, M. Health-promoting benefits of low-fat akawi cheese made by exopolysaccharide-producing probiotic Lactobacillus plantarum isolated from camel milk. J. Dairy Sci. 2017, 100, 7771–7779. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Q.; Ji, Z.; Shu, G.; Chen, H. Production and fermentation characteristics of angiotensin-converting enzyme inhibitory peptides of goat milk fermented by a novel wild Lactobacillus plantarum 69. LWT-Food Sci. Technol. 2018, 91, 532–540. [Google Scholar] [CrossRef]
- Vukotić, G.; Strahinić, I.; Begović, J.; Lukić, J.; Kojić, M.; Fira, D. Survey on proteolytic activity and diversity of proteinase genes in mesophilic lactobacilli. Microbiology 2016, 85, 33–41. [Google Scholar]
- Gobbetti, M.; Ferranti, P.; Smacchi, E.; Goffredi, F.; Addeo, F. Production of Angiotensin-I-Converting-enzyme-inhibitory peptides in fermented milks started by Lactobacillus delbrueckii subsp. bulgaricus SS1 and Lactococcus lactis subsp. cremoris FT4. Appl. Environ. Microbiol. 2000, 66, 3898–3904. [Google Scholar] [CrossRef]
- Georgalaki, M.; Zoumpopoulou, G.; Mavrogonatou, E.; Van Driessche, G.; Alexandraki, V.; Anastasiou, R. Evaluation of the antihypertensive angiotensin-converting enzyme inhibitory (ACE-I) activity and other probiotic properties of lactic acid bacteria isolated from traditional Greek dairy products. Int. Dairy J. 2017, 75, 10–21. [Google Scholar] [CrossRef]
- Villegas, J.M.; Picariello, G.; Mamone, G.; Espeche Turbay, M.B.; Savoy de Giori, G.; Hebert, E.M. Milk-derived angiotensin-I-converting enzyme inhibitory peptides generated by Lactobacillus delbrueckii subsp. lactis CRL 581. Peptidomics 2014, 1, 22–29. [Google Scholar] [CrossRef]
- Rodríguez-Figueroa, J.C.; González-Córdova, A.F.; Torres-Llanez, M.J.; Garciía, H.S.; Vallejo-Cordoba, B. Novel angiotensin I-converting enzyme inhibitory peptides produced in fermented milk by specific wild Lactococcus lactis strains. J. Dairy Sci. 2012, 95, 5536–5543. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Figueroa, J.C.; Reyes-Díaz, R.; González-Córdova, A.F.; Troncoso-Rojas, R.; Vargas-Arispuro, I.; Vallejo-Cordoba, B. Angiotensin-converting enzyme inhibitory activity of milk fermented by wild and industrial Lactococcus lactis strains. J. Dairy Sci. 2010, 93, 5032–5038. [Google Scholar] [CrossRef] [PubMed]
- Picon, A.; Garde, S.; Ávila, M.; Nuñez, M. Proteolytic activities, peptide utilization and oligopeptide transport systems of wild Lactococcus lactis strains. Int. Dairy J. 2010, 20, 156–162. [Google Scholar] [CrossRef]
- Soleymanzadeh, N.; Mirdamadi, S.; Mirzaei, M.; Kianirad, M. Novel β-casein derived antioxidant and ACE-inhibitory active peptide from camel milk fermented by Leuconostoc lactis PTCC1899: Identification and molecular docking. Int. Dairy J. 2019, 97, 201–208. [Google Scholar] [CrossRef]
- Miclo, L.; Roux, E.; Genay, M.; Brusseaux, E.; Poirson, C.; Jameh, N.; Perrin, C.; Dary, A. Variability of hydrolysis of β-, αs1-, and αs2-caseins by 10 strains of Streptococcus thermophilus and resulting bioactive peptides. J. Agric. Food Chem. 2012, 60, 554–565. [Google Scholar] [CrossRef]
- Balthazar, C.F.; Santillo, A.; Guimarães, G.T.; Capozzi, V.; Russo, P.; Caroprese, M.; Marino, R.; Esmerino, A.E.; Raices, R.S.L.; Silva, M.C.; et al. Novel milk-juice beverage with fermented sheep milk and strawberry (Fragaria × ananassa): Nutritional and functional characterization. J. Dairy Sci. 2019, in press. [Google Scholar] [CrossRef]
- Ong, L.; Henriksson, A.; Shah, N.P. Proteolytic pattern and organic acid profiles of probiotic Cheddar cheese as influenced by probiotic strains of Lactobacillus acidophilus, Lb. paracasei, Lb. casei or Bifidobacterium sp. Int. Dairy J. 2007, 17, 67–78. [Google Scholar] [CrossRef]
- Padghan, P.V.; Mann, B.; Sharma, R.; Bajaj, R.; Saini, P. Production of Angiotensin-I-Converting-Enzyme-Inhibitory Peptides in Fermented Milks (Lassi) Fermented by Lactobacillus acidophillus with Consideration of Incubation Period and Simmering Treatment. Int. J. Pept. Res Ther. 2016, 23, 69–79. [Google Scholar] [CrossRef]
- Muguerza, B.; Ramos, M.; Sánchez, E.; Manso, M.A.; Miguel, M.; Aleixandre, A.; Delgado, M.A.; Recio, I. Antihypertensive activity of milks fermented by Enterococcus faecalis strains isolated from raw milk. Int. Dairy J. 2006, 16, 61–69. [Google Scholar] [CrossRef]
- Biscola, V.; Choiset, Y.; Rabesona, H.; Chobert, J.M.; Haertlé, T.; Franco, B.D.G.M. Brazilian artisanal ripened cheeses as sources of proteolytic lactic acid bacteria capable of reducing cow milk allergy. J. Appl. Microbiol. 2018, 125, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Ebner, J.; Arslan, A.A.; Fedorova, M.; Hoffmann, R.; Küçükçetin, A.; Pischetsrieder, M. Peptide profiling of bovine kefir reveals 236 unique peptides released from caseins during its production by starter culture or kefir grains. J. Proteom. 2018, 117, 41–57. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, P.-L.; Gauthier, S.; Bachelard, H.; Santure, M.; Roy, D. Antihypertensive activity of casein-enriched milk fermented by Lactobacillus helveticus. Int. Dairy J. 2002, 12, 995–1004. [Google Scholar] [CrossRef]
- Raveschot, C.; Cudennec, B.; Deracinois, B.; Frémont, M.; Vaeremans, M.; Dugersuren, J.; Demberel, S.; Drider, D.; Dhulster, P.; Coutte, F.; et al. Proteolytic activity of Lactobacillus strains isolated from Mongolian traditional dairy products: A multiparametric analysis. Food Chem. 2020, 304, 125415. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, M.W.; Tellez, A.M. Lactobacillus helveticus: The proteolytic system. Front Microbiol. 2013, 4, 30. [Google Scholar] [CrossRef]
- O’Sullivan, O.; O’Callaghan, J.; Sangrador-Vegas, A. Comparative genomics of lactic acid bacteria reveals a niche-specific gene set. BMC Microbiol. 2009, 9, 50. [Google Scholar] [CrossRef]
- Alcántara, C.; Bäuerl, C.; Revilla-Guarinos, A.; Pérez-Martínez, G.; Monedero, V.; Zúñiga, M. Peptide and amino acid metabolism is controlled by an OmpR-family response regulator in Lactobacillus casei. Mol. Microbiol. 2016, 100, 25–41. [Google Scholar] [CrossRef]
- Kunji, E.R.S.; Mierau, I.; Hagting, A.; Poolman, B.; Konings, W.N. The proteolytic systems of lactic acid bacteria. Ant. Leeuw. Int. 1996, 70, 187–221. [Google Scholar] [CrossRef]
- Sadat-Mekmene, L.; Jardin, J.; Corre, C.; Mollé, D.; Richoux, R.; Delage, M.M.; Lortal, S.; Gagnaire, V. Simultaneous presence of PrtH and PrtH2 proteinases in Lactobacillus helveticus strains improves breakdown of the pure αs1-casein. Appl. Environ. Microbiol. 2011, 77, 179–186. [Google Scholar] [CrossRef]
- Liu, M.; Bayjanov, J.R.; Renckens, B.; Nauta, A.; Siezen, R.J. The proteolytic system of lactic acid bacteria revisited: A genomic comparison. BMC Genomics. 2010, 11, 36. [Google Scholar] [CrossRef]
- Sun, Z.; Harris, H.M.; McCann, A.; Guo, C.; Argimon, S.; Zhang, W.; Yang, X.; Jeffery, I.B.; Cooney, J.C.; Kagawa, T.F.; et al. Expanding the biotechnology potential of lactobacilli through comparative genomics of 213 strains and associated genera. Nat. Commun. 2015, 6, 8322. [Google Scholar] [CrossRef] [PubMed]
- Pederson, J.A.; Mileski, G.J.; Weimer, B.C.; Steele, J.L. Genetic characterization of a cell envelope-associated proteinase from Lactobacillus helveticus CNRZ32. J. Bacteriol. 1999, 181, 4592–4597. [Google Scholar] [PubMed]
- Genay, M.; Sadat, L.; Gagnaire, V.; Lortal, S. prtH2 and not prtH is the ubiquitous cell-wall proteinase gene in Lactobacillus helveticus. Appl. Environ. Microbiol. 2009, 19, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, J.R.; Hughes, J.E.; Welker, D.L.; Tompkins, T.A.; Steele, J.L. Complete genome sequence for Lactobacillus helveticus CNRZ 32, an industrial cheese starter and cheese flavour adjunct. Genome Announc. 2013, 1, e00590-13. [Google Scholar] [CrossRef]
- Savijoki, K.; Ingmer, H.; Varmanen, P. Proteolytic systems of lactic acid bacteria. Appl. Microbiol. Biotechnol. 2006, 71, 394–406. [Google Scholar] [CrossRef]
- Zhao, W.; Chen, Y.; Sun, Z.; Wang, J.; Zhou, Z.; Sun, T.; Wang, J.; Zhou, Z.; Sun, T.; Wang, L.; et al. Complete genome sequence of Lactobacillus helveticus H10. J. Bacteriol. 2011, 193, 2666–2667. [Google Scholar] [CrossRef]
- Tynkkynen, S.; Buist, G.; Kunji, E.; Kok, J.; Poolman, B.; Venema, G.; Haandrikman, A. Genetic and biochemical characterization of the oligopeptide transport system of Lactococcus lactis. J. Bacteriol. 1993, 175, 7523–7532. [Google Scholar] [CrossRef]
- Detmers, F.J.M.; Kunji, E.R.S.; Lanfermeijer, F.C.; Poolman, B.; Konings, W.N. Kinetics and specificity of peptide uptake by the oligopeptide transport system of Lactococcus lactis. Biochemistry 1998, 37, 16671–16679. [Google Scholar] [CrossRef]
- Doeven, M.K.; Abele, R.; Tampé, R.; Poolman, B. The binding specificity of OppA determines the selectivity of the oligopeptide ATP-binding cassette transporter. J. Biol. Chem. 2004, 279, 32301–32307. [Google Scholar] [CrossRef]
- Peltoniemi, K.; Vesanto, E.; Palva, A. Genetic characterization of an oligopeptide transport system from Lactobacillus delbrueckii subsp. Bulgaricus. Arch. Microbiol. 2002, 177, 457–467. [Google Scholar] [CrossRef]
- Hagting, A.; Kunji, E.R.S.; Leenhouts, K.J.; Poolman, B.; Konings, W.N. The di- and tripeptide transport protein of Lactococcus lactis: A new type of bacterial transporter. J. Biol. Chem. 1994, 269, 11391–11399. [Google Scholar] [PubMed]
- Nakajima, H.; Hagting, A.; Kunji, E.R.; Poolman, B.; Konings, W.N. Cloning and functional expression in Escherichia coli of the gene encoding the di- and tripeptide transport protein of Lactobacillus helveticus. Appl. Environ. Microbiol. 1997, 63, 2213–2217. [Google Scholar]
- Foucaud, C.; Kunji, E.R.S.; Hagting, A.; Richard, J.; Konings, W.N.; Desmazeaud, M.; Poolman, B. Specificity of peptide transport systems in Lactococcus lactis: Evidence for third system which transport hydrophobic di- and tripeptides. J. Bacteriol. 1995, 177, 4652–4657. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sanz, Y.; Lanfermeijer, F.C.; Renault, P.; Bolotin, A.; Konings, W.N.; Poolman, B. Genetic and functional characterization of dpp genes encoding a dipeptide transport system in Lactococcus lactis. Arch. Microbiol. 2001, 175, 334–343. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Doeven, M.K.; Kok, J.; Poolman, B. Specificity and selectivity determinants of peptide transport in Lactococcus lactis and other microorganisms. Mol. Microbiol. 2005, 57, 640–649. [Google Scholar] [CrossRef]
- Christensen, J.E.; Dudley, E.G.; Pederson, J.A.; Steele, J.L. Peptidases and amino acid catabolism in lactic acid bacteria. Antonie Van Leeuwenhoek 1999, 76, 217–246. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.R.; Schick, J.; Henrich, B.; Plapp, R. Lactobacillus delbrueckii subsp. lactis DSM7290 pepG gene encodes a novel cysteine aminopeptidase. Microbiology 1997, 143, 527–537. [Google Scholar] [CrossRef]
- Fenster, H.M.; Parkin, K.M.; Steele, J.L. Characterization of a Thiol-Dependent Endopeptidase from Lactobacillus helveticus CNRZ32. J. Bacteriol. 1997, 179, 2529–2533. [Google Scholar] [CrossRef]
- Shridar, V.M.; Hughes, J.E.; Welker, D.L.; Broadbent, J.R.; Steele, J.L. Identification of endopeptidase genes from the genomic sequence of Lactobacillus helveticus CNRZ32 and the role of these genes in hydrolysis of model bitter peptides. Appl. Environ. Microbiol. 2005, 71, 3025–3032. [Google Scholar] [CrossRef]
- Monnet, V.; Nardi, M.; Chopin, A.; Chopin, M.C.; Gripon, J.C. Biochemical and genetic characterization of PepF, an oligopeptidase from Lactococcus lactis. J. Biol. Chem. 1994, 269, 32070–32076. [Google Scholar]
- Pritchard, G.G.; Freebairn, A.D.; Coolbear, T. Purification and characterization of an endopeptidase from Lactococcus lactis subsp. cremoris SK11. Microbiology 1994, 140, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Christensen, J.E.; Broadbent, J.R.; Steele, J.L. Identification and characterization of Lactobacillus helveticus PepO2, an endopeptidase with post-proline specificity. Appl. Environ. Microbiol. 2003, 69, 1276–1282. [Google Scholar] [CrossRef] [PubMed]
- Fernández de Palencia, P.; Peláez, C.; Martín-Hernández, C. Purification and characterization of an aminopeptidase (Pep C-like) from Lactobacillus casei subsp. casei IFPL 731 isolated from raw goat’s milk cheese. Int. Dairy J. 1997, 7, 629–634. [Google Scholar]
- Fernández de Palencia, P.; Peláez, C.; Martín-Hernández, C. Characterization of the aminopeptidase system from Lactobacillus casei subsp. casei IFPL 731. J. Agric. Food Chem. 1997, 45, 3778–3781. [Google Scholar]
- Vesanto, E.; Vermanen, P.; Steele, J.L.; Palva, A. Characterization and expression of the Lactobacillus helveticus pepC gene encoding a general aminopeptidase. Eur. J. Biochem. 1994, 224, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Klein, J.R.; Klein, U.; Schad, M.; Plapp, R. Cloning, DNA sequence analysis and partial characterization of pepN, a lysyl aminopeptidase from Lactobacillus delbrueckii ssp. lactis DSM7290. Eur. J. Biochem. 1993, 217, 105–114. [Google Scholar] [CrossRef]
- Bouchier, P.L.; FitzGerald, R.J.; O’Cuinn, G. Hydrolysis of αs1- and β-casein-derived peptides with a broad specificity aminopeptidase and proline specific aminopeptidases from Lactococcus lactis subsp. cremoris AM2. FEBS Lett. 1999, 445, 321–324. [Google Scholar] [CrossRef]
- Mars, I.; Monnet, V. An aminopeptidase P from Lactococcus lactis with original specificity. Biochim. Biophys. Acta 1995, 1243, 209–215. [Google Scholar] [CrossRef]
- El Abboudi, M.; El Soda, M.; Pandian, S.; Simard, R.E.; Olson, N.F. Purification of X-prolyl dipeptidyl aminopeptidase from Lactobacillus casei subspecies. Int. J. Food Microbiol. 1992, 15, 57–98. [Google Scholar] [CrossRef]
- Stressler, T.; Eisele, T.; Kranz, B.; Fischer, L. PepX from Lactobacillus helveticus: Automated multi-step purification and determination of kinetic parameters with original tripeptide substrates. J. Mol. Catal. B-Enzym. 2017, 108, 103–110. [Google Scholar] [CrossRef]
- Fernández-Espla, M.D.; Martín-Hernández, M.C. Purification and characterization of a dipeptidase from Lactobacillus casei ssp. casei IFPL 731 isolated from goat cheese made from raw milk. J. Dairy Sci. 1997, 80, 1497–1504. [Google Scholar] [CrossRef]
- Savijoki, K.; Palva, A. Purification and molecular characterization of a tripeptidase (PepT) from Lactobacillus helveticus. Appl. Environ. Microbiol. 2000, 66, 794–800. [Google Scholar] [CrossRef] [PubMed]
- Varmanen, P.; Rantanen, T.; Palva, A.; Tynkkynen, S. Cloning and characterization of a prolinase gene (pepR) from Lactobacillus rhamnosus. Appl. Environ. Microbiol. 1998, 64, 1831–1836. [Google Scholar] [PubMed]
- Varmanen, P.; Rantanen, T.; Palva, A. An operon from Lactobacillus helveticus composed of a proline iminopeptidase gene (pepI) and two genes coding for putative members of the ABC transporter family of proteins. Microbiology 1996, 142, 3459–3468. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Fernández-Espla, M.D.; Martín-Hernández, M.C.; Fox, P.F. Purification and characterization of a prolidase from Lactobacillus casei subsp. casei IFPL 731. Appl. Environ. Microbiol. 1997, 63, 314–316. [Google Scholar]
- Cai, H.; Thompson, R.; Budinich, M.; Broadbent, J.R.; Steele, J.L. Genome sequence and comparative genome analysis of Lactobacillus casei: Insights into their niche-associated evolution. Genome Biol. Evol. 2009, 1, 239–257. [Google Scholar] [CrossRef]
- Pangallo, D.; Kraková, L.; Puškárová, A.; Šoltys, K.; Bučková, M.; Koreňová, J.; Budiš, J.; Kuchta, T. Transcription activity of lactic acid bacterial proteolysis-related genes during cheese maturation. Food Microbiol. 2019, 82, 416–425. [Google Scholar] [CrossRef]
- Soeryapranata, E.; Powers, J.R.; Ünlü, G. Cloning and characterization of debittering peptidases, PepE, PepO, PepO2, PepO3, and PepN, of Lactobacillus helveticus WSU19. Int. Dairy J. 2007, 17, 1096–1106. [Google Scholar] [CrossRef]
- Parra, L.; Fernández de Palencia, P.; Casal, V.; Requena, T.; Peláez, C. Hydrolysis of β-casein (193–209) fragment by whole cells and fractions of Lactobacillus casei and Lactococcus lactis. J. Food Sci. 1999, 64, 899–902. [Google Scholar] [CrossRef]
- Ueno, K.; Mizuno, S.; Yamamoto, N. Purification and characterization of an endopeptidase that has an important role in the carboxyl terminal processing of antihypertensive peptides in Lactobacillus helveticus CM4. Lett. Appl. Microbiol. 2004, 39, 313–318. [Google Scholar] [CrossRef]
- Cremonesi, P.; Chessa, S.; Castiglioni, B. Genome sequence and analysis of Lactobacillus helveticus. Front. Microbiol. 2013, 3, 435. [Google Scholar] [CrossRef] [PubMed]
- Wakai, T.; Yamaguchi, N.; Hatanaka, M.; Nakamura, Y.; Yamamoto, N. Repressive processing of antihypertensive peptides, Val-Pro-Pro and Ile-Pro-Pro, in Lactobacillus helveticus fermented milk by added peptides. J. Biosci. Bioeng. 2012, 114, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Guédon, E.; Renault, P.; Ehrlich, S.D.; Delorme, C. Transcriptional pattern of genes coding for the proteolytic system of Lactococcus lactis and evidence for coordinated regulation of key enzymes by peptide supply. J. Bacteriol. 2001, 183, 3614–3622. [Google Scholar] [CrossRef] [PubMed]
- Chambellon, E.; Yvon, M. CodY-Regulated Aminotransferases AraT and BcaT play a major role in the growth of Lactococcus lactis in milk by regulating the intracellular pool of amino acids. Appl. Environ. Microbiol. 2003, 69, 3061–3068. [Google Scholar] [CrossRef]
- Wakai, T.; Yamamoto, N. A novel branched chain amino acids responsive transcriptional regulator, BCARR, negatively acts on the proteolytic system in Lactobacillus helveticus. PLoS ONE 2013, 8, e75976. [Google Scholar] [CrossRef]
- Brown, L.; Villegas, J.M.; Elean, M.; Fadda, S.; Mozzi, F.; Saavedra, L.; Hebert, H.M. YebC, a putative transcriptional factor involved in the regulation of the proteolytic system of Lactobacillus. Sci. Rep. 2017, 7, 8579. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brand | Category | Producers | Molecules | Bioactivities |
|---|---|---|---|---|
| BioPURE-GMP | Whey protein isolate | Davisco, USA | κ-casein f(106–109) | Satiety regulation, anti-carcinogenic, antimicrobial, anti-thrombotic BPs |
| BioZate | Whey protein isolate | Davisco, USA | β-lactoglobulin fragments | Hypotensive BPs |
| C12 Peption | Ingredients | DMV, The Netherlands | Casein derived FFVAPFPEVFGK | Hypotensive BP |
| Capolac | Ingredients | Arla Food, Denmark | Casein phosphopeptide | Mineral absorption |
| Casein DP/Peptio Drink | Soft milk drink | Kracie Pharmaceuticals, Japan | Casein derived FFVAPFPEVFGK | Hypotensive |
| CE90CPP | Ingredients | DMV, The Netherlands | Casein phosphopeptide | Mineral absorption |
| PeptoPro | Flavored drink | DSM Food specialties, The Netherlands | Casein phosphopeptide | Improves muscle performance |
| Cysteine peptide | Ingredients/hydrolysate | DMV, The Netherlands | Casein peptides | hypotensive BPs |
| Praventin | Capsule | DMV, The Netherlands | Lactoferrin-enriched whey protein hydrolysate | Antimicrobial BPs (reduced skin infection) |
| Festivo | Low-fat cheese | MIT Agrifood Research, Finland | αS1-casein f(1–6), αS1-casein f(1–7), αS1-casein f(1–9) | Hypotensive BPs |
| Kotsu Kotsu calcium | Soft drink | Asahi Soft Drink | Casein phosphopeptide | Mineral absorption |
| Evolus | Fermented milk | Valio Ltd. Finland | VPP, IPP | Hypotensive BPs |
| Calpis/Ameal S 120 | Sour milk | Calpis Co Ltd, Japan | VPP, IPP | Hypotensive BPs |
| Ameal S | Tablet | Calpis Co Ltd, Japan | VPP, IPP | Hypotensive BPs |
| Immunel | Ingredient | Wild Co., Germany | Milk peptides | Anti-inflammatory BPs |
| Tegricel | Ingredient | Wild Co., Germany | Milk peptides | Anti-inflammatory BPs |
| Lactium/ProDiet F200 | Flavored milk drink/capsules | Ingredia, France | αS1-casein f(91–100) YLGYLEQLLR | Stress relief BPs |
| Vivinal Alpha | Ingredient | Borculo Domo Ingredients | α-lactalbumin-rich whey protein hydrolysate | Aids relaxation and sleep |
| Dermylex | Tablet | Advitech Inc., Canada | Whey protein extract XP-828L | Reduces symptoms of psoriasis |
| Capolac | Ingredient | Arla Food Ingredients | Casein phosphopeptide | Mineral absorption |
| Recaldent | Chewing gum | Cadbury Enterprises | Calcium casein peptone-calcium phosphate | Anticariogenic BPs |
| Peptide | Fragment | Source | In vivo Model | Dose | Systolic Blood Pressure Reduction | References |
|---|---|---|---|---|---|---|
| VPP 1 | β-casein f(84–86) | Fermented milk, cheese, yogurt | Huma SHR | 3–100 mg/day 5 mg/kg | −3.73 mmHg −20.1 mmHg | [23,51,52,53,54,55,56,57,58] |
| IPP 1 | β-casein f(74–76) κ-casein f(108–110) | Fermented milk, cheese, yogurt | Human SHR | 3–100 mg/day 5 mg/kg | −3.73 mmHg −18.3 mmHg | [23,51,52,53,54,55,56,57,58] |
| YP | Various fragments in β-casein, αS1-casein, κ-casein | Cheese, yogurt | SHR | 1 mg/kg | −27.4 mmHg | [55,59,60] |
| TKVIP | αS2-casein f(198–202) | Yogurt | SHR | 1 mg/kg | −9.2 mmHg | [55,61] |
| KVLPVPQ | β-casein A2 f(169–175) | Cheese, yogurt | SHR | 2 mg/kg | −31.5 mmHg | [55,61] |
| LHLPLP | β-casein A2 f(133–138) | Cheese, yogurt | SHR | 3 mg/kg | −25.3 mmHg | [53,54,55,62] |
| RYLGY | αS1-casein f(90–94) | Cheese | SHR | 5 mg/kg | −25 mmHg | [53,54,63,64] |
| AYFYPEL | αS1-casein f(143–149) | Cheese | SHR | 5 mg/kg | −20 mmHg | [53,54,63] |
| RYLG | αS1-casein f(90–93) | Cheese | SHR | 5 mg/kg | −18 mmHg | [53,54,64] |
| RY | Various fragments in αS1-casein, αS2-casein, κ-casein | Fermented milk | SHR | 5 mg/kg | −18 mmHg | [64,65] |
| HLPLP | β-casein A2 f(134–138) | Cheese | SHR | 7 mg/kg | −23.5 mmHg | [53,54,66] |
| LPLP | β-casein A2 f(135–138) | Yogurt | SHR | 7 mg/kg | −16 mmHg | [55,67] |
| PLP | β-casein A2 f(136–138) | Cheese | SHR | 7 mg/kg | −21.2 mmHg | [59,67] |
| FP | Various fragments in β-casein, αS1-casein, αS2-casein | Cheese, yogurt | SHR | 8 mg/kg | −27 mmHg | [55,58,59,68] |
| LVYPFTGPIPN | β-casein caprine f(58–68) | Kefir | SHR | 10 mg/kg | −28 mmHg | [66,69] |
| VRGPFPIIV | β-casein f(201–209) | Yogurt | SHR | 10 mg/kg | −16 mmHg | [55,62] |
| KKYNVPQL | αS1-casein caprine f(102–109) | Cheese | SHR | 10 mg/kg | −11.5 mmHg | [48,58,66] |
| AVPYPQR | β-casein f(177–183) | Kefir | SHR | 100 mg/kg | −10 mmHg | [70,71] |
| RPKHPIKHQ | αS1-casein f(1–9) | Cheese | SHR | n.a. | −9.3 mmHg | [48,58,72] |
| TPVVVPPFLQP | β-casein f(80–90) | Cheese, yogurt | SHR | n.a. | −8 mmHg | [55,59,68] |
| YPFPGPIPN | β-casein A2 f(60–68) | Cheese, kefir | SHR | n.a. | −7 mmHg | [48,58,72,73] |
| Peptide | Fragment | Source | References |
|---|---|---|---|
| ARHPHPHLSFM • | κ-casein f(96–106) | Yogurt | [55,129] |
| AVPYPQR * | β-casein f(177–183) | Kefir | [70,126] |
| AYFYPE • | αS1-casein f(143–148) | Yogurt, cheese | [55,64] |
| AYFYPEL • | αS1-casein f(143–149) | Cheese | [53,54,64] |
| EMPFPK ▪ | β-casein f(108–113) | Yogurt | [55] |
| EVFGKEKVN ▪ | αS1-casein f(30–38) | Kefir | [70,131] |
| FALPQYLK • | αs2-casein f(174–181) | Kefir | [70,132] |
| FSDKIAKYIPIQ ▪ | κ-casein f(18–29) | Yogurt | [55] |
| GPVRGPFPII • | β-casein f(199–208) | Fermented milk, yogurt | [119,127] |
| HLPLPL • | β-casein f(133–138) | Yogurt, fermented milk | [55,105] |
| IPIQY • | κ-casein f(26–32) | Yogurt | [127] |
| IPIQYVL • | κ-casein f(26–30) | Fermented milk | [119] |
| KAVPYPQ • | β-casein f(176–182) | Yogurt | [127] |
| KIHPFAQTQ • | β-casein f(48–56) | Yogurt | [133,134] |
| KVLPVPQ • | β-casein f(169–175) | Fermented milk | [119] |
| KVLPVPQK • | β-casein f(169–176) | Fermented milk | [105,126] |
| LQDKIHP • | β-casein f(45–51) | Yogurt | [134] |
| PYVRYL * | αs2-casein f(203–208) | Kefir | [132] |
| QEPVLGPVRGPFPII • | β-casein f(194–208) | Yogurt | [127] |
| QQPVLGPVRGPFPIIV • | β-casein f(194–209) | Yogurt | [127] |
| RDMPIQ • | β-casein f(183–188) | Fermented milk | [105,134] |
| RPKHPIK ▪ | αS1-casein f(1–7) | Cheese | [58] |
| RPKHPIKHQGLPQEVLNENLLRF ▪ | αS1-casein f(1–23) | Kefir | [70] |
| RY • | Various fragments in αS1-casein, αS2-casein, κ-casein | Fermented milk, cheese | [53,119] |
| RYLG • | αS1-casein f(90–93) | Cheese | [53,54,64] |
| RYLGY • | αS1-casein f(90–94) | Cheese | [53,54,64] |
| SDIPNPIGSENSE ▪ | αS1-casein f(180–192) | Kefir | [70] |
| SKVLPVPQ • | β-casein f(168–175) | Fermented milk drinks | [128] |
| STVATL ▪ | κ-casein f(141–146) | Yogurt | [55] |
| TVQVTSTAV ▪ | κ-casein f(161–169) | Yogurt | [55] |
| VKEAMAPK • | β-casein f(98–105) | Fermented milk, cheese | [105,125,126] |
| VLNENLLR ▪ | αS1-casein f(15–22) | Kefir | [70] |
| VLPVPQK * | β-casein f(170–176) | Fermented milk | [105,126] |
| VPYPQ • | β-casein f(178–182) | Yogurt | [127] |
| VYPFPGPIPN • | β-casein A2 f(59–68) | Kefir | [70,124] |
| YQEPVLGPVRGPFPI ▪ | β-casein f(191–205) | Kefir | [70] |
| YQEPVLGPVRGPFPIIV * | β-casein f(191–207) | Kefir | [124] |
| YVL ▪ | κ-casein f(30–32) | Yogurt | [55] |
| Peptide | Fragment | Source | IC50 μmol/L | References |
|---|---|---|---|---|
| WL | α-lactalbumin f(104–105), α-lactalbumin f(118–119) | Yogurt | 44 | [55] |
| LPQNIPPL | β-casein f(70–77) | Cheese | 46 | [157] |
| LPQ | β-casein f(70–72) | Cheese | 82 | [55] |
| VPITPTL | αs2-casein f(117–123) | Cheese | 110 | [157] |
| VPITPT | αs2-casein f(117–122) | Cheese | 130 | [157] |
| LPQNIPP | β-casein f(70–76) | Cheese | 160 | [157] |
| GPFPILV | β-casein caprine f(201–207) | Kefir | 163 | [69] |
| FPGPIPN | β-casein f(62–68) | Cheese | 260 | [157] |
| YP | Various fragments in β-casein, αS1-casein, κ-casein | Yogurt | 658 | [55] |
| YPFPGPIPN | β-casein f(60–68) | Cheese, kefir | 670 | [48,58,73,157,158] |
| PGPIHNS | β-casein f(63–69) | Cheese | 1000 | [157] |
| IPPLTQTPV | β-casein f(74–82) | Cheese | 1300 | [157] |
| PQNIPPL | β-casein f(71–77) | Cheese | 1500 | [157] |
| VPPFIQPE | β-casein f(84–91) | Cheese | 2500 | [157] |
| Species | Strain | Food Source | Novel Dairy Product | BPs-mediated Activities | References |
|---|---|---|---|---|---|
| L. helveticus | 130B4 | Mongolian camel milk | Fermented skim milk | ACE-i | [186] |
| s6-HTCH, s10-AVCH, s12-AVCH | Mexican Chiapas cheese | Fermented milk | ACE-i | [187] | |
| CM4 1, CP790 1 | Japanese sour milk | Fermented milk | ACE-i | [188] | |
| CPN4 1 | Japanese sour milk | Yogurt | ACE-i | [60] | |
| H9 1 | Traditional Tibetan kurut (fermented yak milk) | Different kinds of fermented milk, yogurt | ACE-i | [189] | |
| 141, T105 | Polish dairy products | Fermented milk | Antioxidative, opioid, stimulating, hypotensive, immunomodulating, antibacterial and antithrombotic | [190] | |
| ASCC474, ASCC1188, ASCC1315 | Australia dairy products | Fermented milk | ACE-i, immunomodulatory | [191] | |
| DPC4571 | Swiss Cheese whey | Fermented milk; adjunct in Cheddar cheese | ACE-i | [192] | |
| MTCC5463 | Indian dairy product | Honey- flavored fermented milk | Antihypertensive | [193] | |
| LBB BY 21 | Yogurt | Fermented milk | Antimicrobial | [194] | |
| ND01 | Chinese naturally fermented milk | Gouda cheese | ACE-i | [195] | |
| PR4 | Italian cheese | Fermented caseinate | ACE-i | [196] | |
| M92 | Spontaneously acidified milk | Yogurt | ACE-i | [197] | |
| L. casei | IMAU20411 | Chinese Dairy food | Fermented milk | ACE-i | [158] |
| Zhang | Koumiss | Cheddar cheese | ACE-i | [198] | |
| LLG | Cheese | Fermented whey drink | ACE-i | [199] | |
| PRA205 | Parmigiano Reggiano cheese | Fermented skim milk, yogurt | ACE-i | [80] | |
| ATCC393 | Dairy food | Fermented milk | ACE-i, antioxidant and anticancer | [200] | |
| ATCC393 | Dairy food | Yogurt with St. thermophilus and L. bulgaricus | [201] | ||
| L. paracasei | in co-culture with Candida | Comté cheese | Whey drink | ACE-i, reduction in α-lactalbumin and β-lactoglobulin | [202] |
| MTCC 5945 (NS4) | Fermented milk | Fermented camel milk | ACE-i | [203] | |
| PTCC 1637 | Camel sour milk | Fermented camel milk | ACE-i and antioxidant | [204] | |
| L. plantarum | 55 | Portuguese raw ewe milk semisoft cheeses | Fermented cow skim milk | anti-inflammatory, antihemolytic and antioxidant | [205] |
| KX881772, KX881779 | Camel milk | Fermented cow camel milk | ACE-i, antiproliferation | [206] | |
| KX881772, KX881780 | Camel milk | Low-fat akawi cheese | ACE-i, antioxidant | [207] | |
| 69 | Mongolian fermented cow milk | Fermented goat milk | ACE-i | [208] | |
| LMG18024 | Buffalo milk | Fermented skim milk | ACE-i | [209] | |
| BGPV2–45a, BGBUK 2–5, BGGA-8 | Homemade cheese | Fermented skim milk | ACE-i | [209] | |
| L. delbrueckii subsp. bulgaricus | SS1 | Dairy product | Fermented milk | ACE-i | [210] |
| ACA-DC 87 | Greek sheep milk yogurt | Greek yogurt and feta cheese | ACE-i | [211] | |
| b38 | Yogurt | Fermented milk | Antimicrobial | [194] | |
| CRL 581 | Argentinian hard cheese | Fermented milk | ACE-i, anti-inflammatory | [212] | |
| Lc. lactis subsp. lactis | NRRL B-50571, NRRL B-50572 | Cheese | Fermented cow milk | ACE-i | [213] |
| Q1, Q5 | Chihuahua cheese | Fermented milk | ACE-i | [214] | |
| ESI197, M21, P21 | Manchego cheese | Manchego cheese | ACE-i | [215] | |
| KX881782 | Camel milk | Fermented camel milk | ACE-i, antiproliferation | [206] | |
| Lc. lactis subsp. cremoris | FT4 | Dairy product | Fermented milk | ACE-i | [210] |
| Leu. lactis | PTCC1899 | Traditional fermented camel milk (Chal) | Fermented milk | ACE-i, antioxidant | [176,216] |
| St. thermophilus | ACA-DC 835 | Greek Cow milk yogurt | Greek yogurt and feta cheese | ACE-i | [211] |
| LMD-9, PB302, PB385, CNRZ404, ATE19PB8, Y4, HAD8a, CNRZ1066 | Yogurt | Fermented cow milk | ACE-i, immunomodulating and antimicrobial | [217] | |
| CNRZ445 | Cheese | Fermented cow milk | ACE-i | [217] | |
| ATCC19258 | Pasteurized milk | Fermented cow milk | ACE-i | [217] | |
| in co-culture with L. delbrueckii subsp. bulgaricus | Yogurt | milk–juice beverage with fermented sheep milk and strawberries | ACE-i | [218] | |
| L. acidophilus | L10 | Dairy food | Fermented milk | ACE-i | [219] |
| NCDC-15 in co-culture with St. thermophilus NCDC167 | Lassi | Fermented buffalo milk | ACE-i, immunomodulatory, antioxidant, opioid and cytomodulatory | [220] | |
| E. faecalis2 | CECT 5727, CECT 5728, CECT 5826, CECT 5827 | Raw milk | Fermented skim milk | ACE-i | [221] |
| 43 | Brazilian artisanal ripened cheese | Fermented cow milk | nr | [222] | |
| Kefir grains | Lactobacilli and yeast consortium | - | Fermented cow milk | antihypertensive, antimicrobial, immunomodulatory, opioid and anti-oxidative | [70] |
| Lactobacilli and yeast consortium | - | Kefir | ACE-i, antimicrobial | [223] | |
| L. reuteri | KX881777 | Camel milk | Fermented cow camel milk | ACE-i, antiproliferation | [206] |
| P. acidilactici | 90 | Brazilian artisanal ripened cheese | Fermented cow milk | nr | [222] |
| Undefined LAB species | s6-HTCH, s10-AVCH, s12-AVCH | Chiapas cream cheese | Fermented milk | ACE-i, antioxidant | [187] |
| W. viridescens | 111 | Brazilian artisanal ripened cheese | Fermented cow milk | nr | [222] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tagliazucchi, D.; Martini, S.; Solieri, L. Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food. Fermentation 2019, 5, 96. https://doi.org/10.3390/fermentation5040096
Tagliazucchi D, Martini S, Solieri L. Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food. Fermentation. 2019; 5(4):96. https://doi.org/10.3390/fermentation5040096
Chicago/Turabian StyleTagliazucchi, Davide, Serena Martini, and Lisa Solieri. 2019. "Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food" Fermentation 5, no. 4: 96. https://doi.org/10.3390/fermentation5040096
APA StyleTagliazucchi, D., Martini, S., & Solieri, L. (2019). Bioprospecting for Bioactive Peptide Production by Lactic Acid Bacteria Isolated from Fermented Dairy Food. Fermentation, 5(4), 96. https://doi.org/10.3390/fermentation5040096