
Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress

 ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material and Treatments
2.2. Determination of Biochemical Parameters
2.3. Determination of Growth Parameters and Cd Content
2.4. Statistical Analysis
3. Results
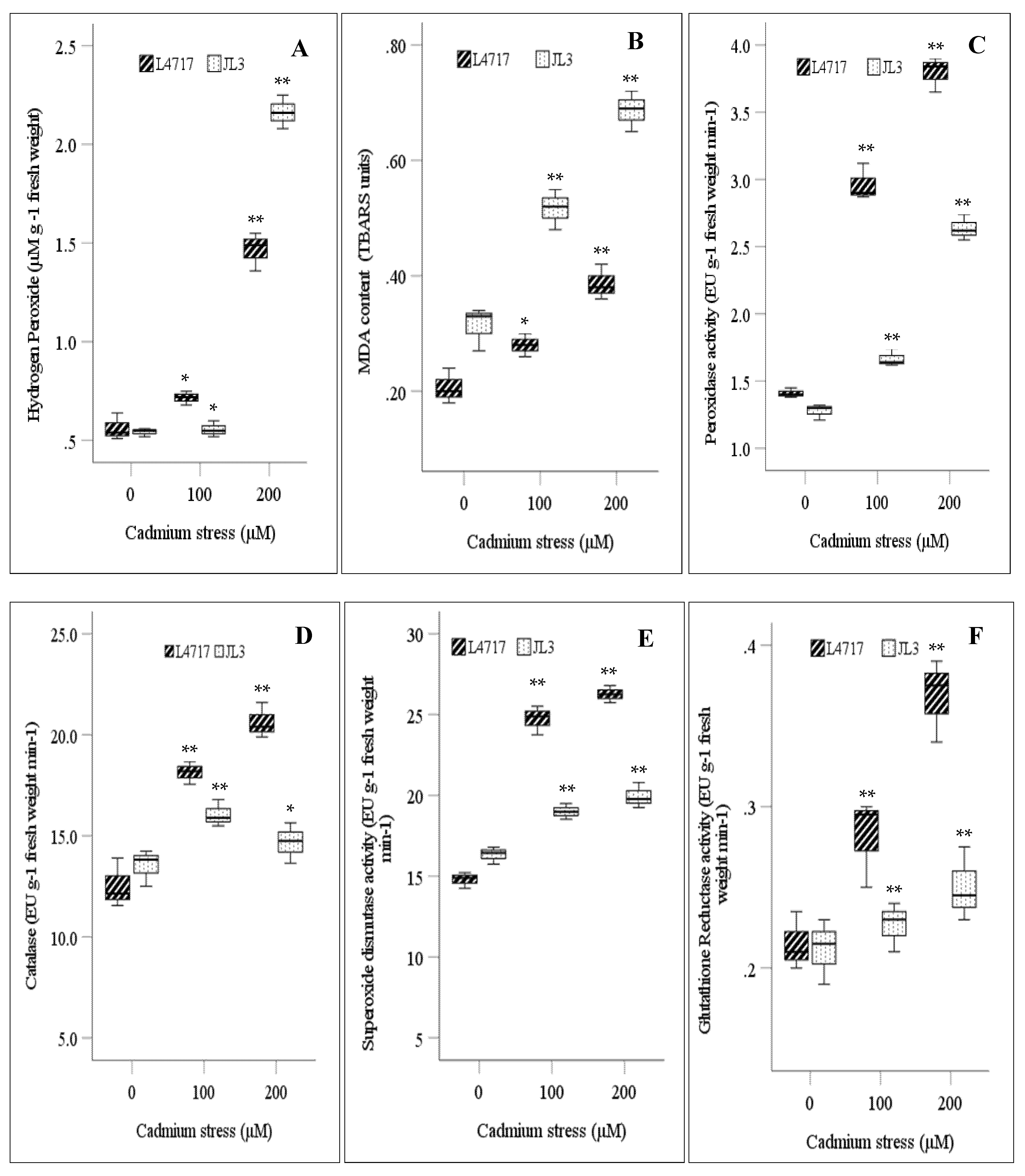
3.1. Effect of Cd on Biochemical Parameters
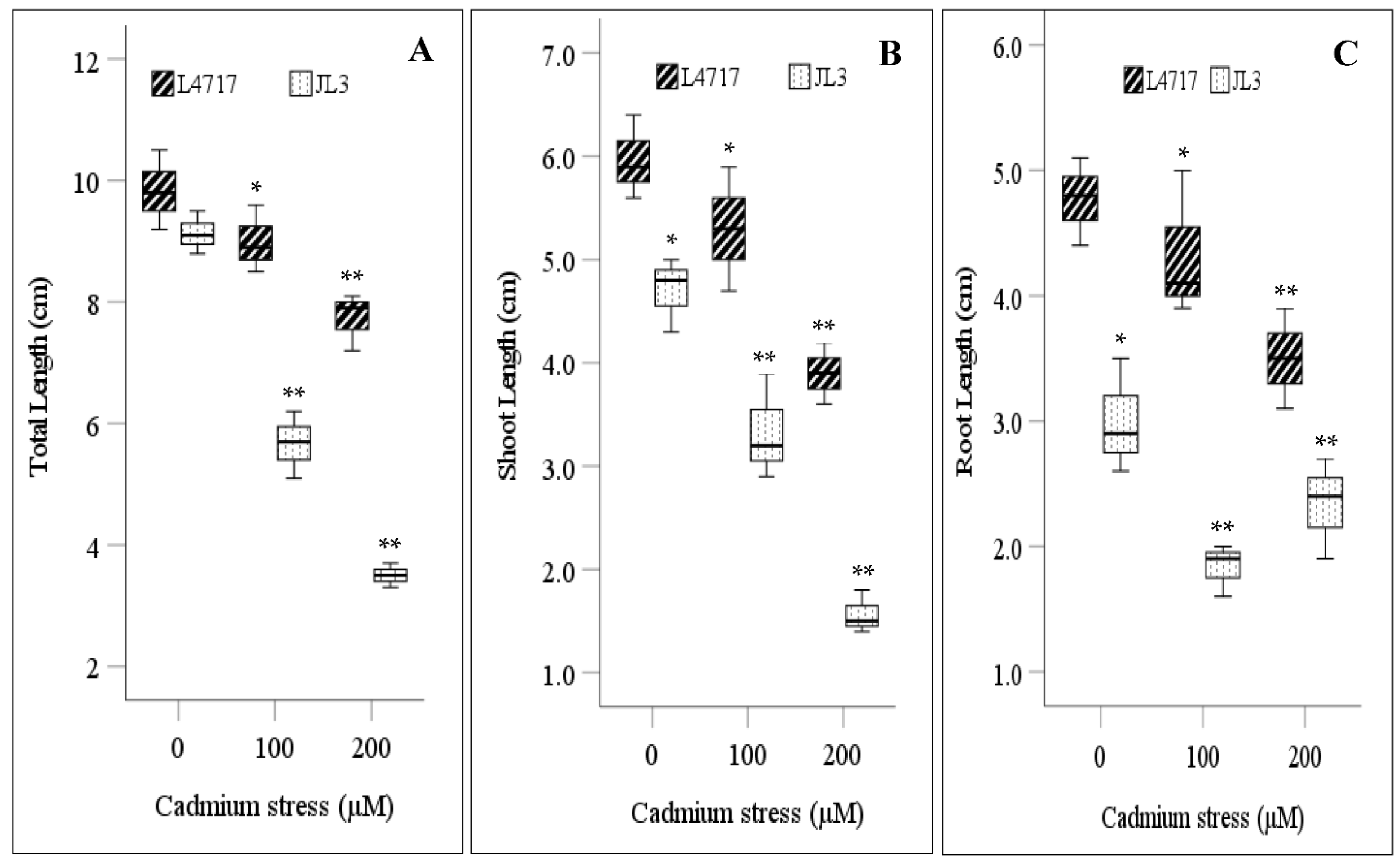
3.2. Effect of Cd Stress on Seedling Growth
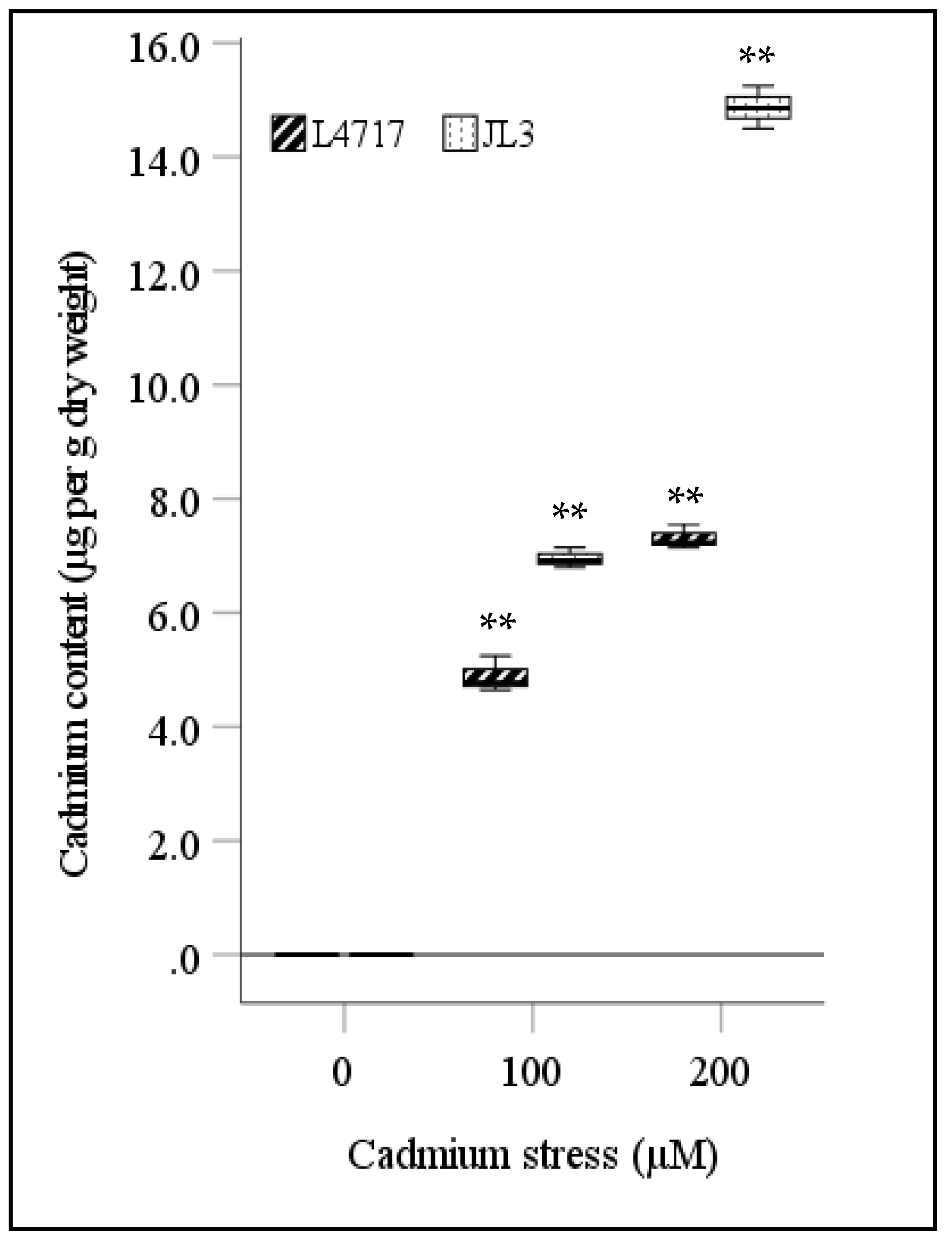
3.3. Cd Uptake
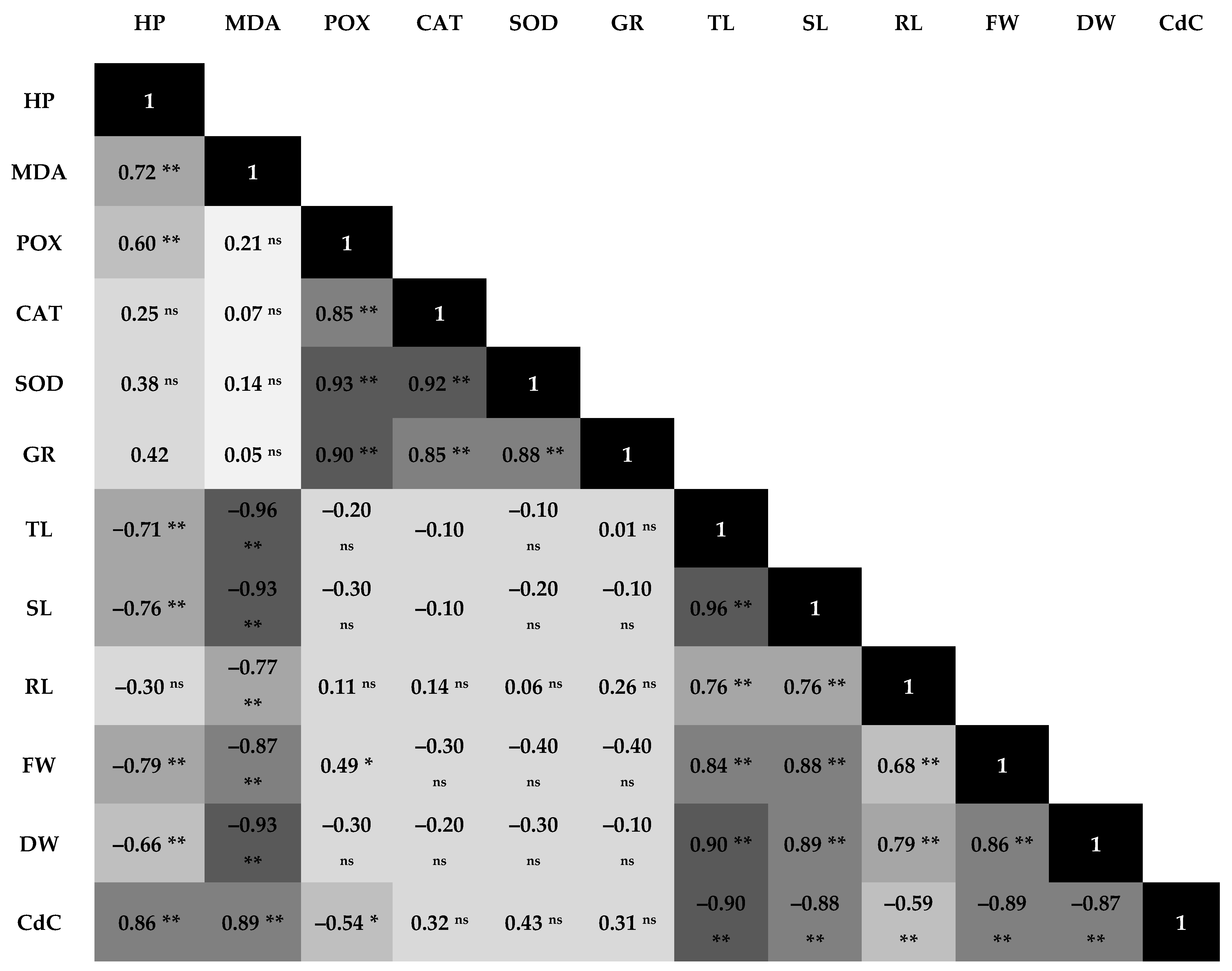
3.4. Pearson’s Correlations Analysis to Cd Stress
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bolan, N.S.; Adriano, D.C.; Mani, P.; Duraisamy, A.; Arulmozhiselvan, S. Immobilization and phytoavailability of Cd in variable charge soils: I. Effect of phosphate addition. Plant Soil 2003, 250, 83–94. [Google Scholar] [CrossRef]
- Choppala, G.; Ullah, S.; Bolan, N.; Bib, S.; Iqbal, M.; Rengel, Z.; Kunhikrishnan, A.; Ashwath, N.; Ok, Y.S. Cellular mechanisms in higher plants governing tolerance to Cd toxicity. Crit. Rev. Plant. Sci. 2004, 33, 374–391. [Google Scholar] [CrossRef]
- Paunov, M.; Koleva, L.; Vassilev, A.; Vangronsveld, J.I.D.; Goltsev, V. Effects of different metals on photosynthesis: Cd and zinc affect chlorophyll fluorescence in durum wheat. Int. J. Mol. Sci. 2018, 19, 787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biyani, K.; Tripathi, D.K.; Lee, J.H.; Muneer, S. Dynamic role of iron supply in amelioration of Cd stress by modulating antioxidative pathways and peroxidase enzymes in mungbean. AoB Plants 2019, 11, plz005. [Google Scholar] [CrossRef] [Green Version]
- Rahmatizadeh, R.; Arvin, S.M.J.; Jamei, R.; Mozaffari, H.; Nejhad, F.R. Response of tomato plants to interaction effects of magnetic (Fe3O4) nanoparticles and Cd stress. J. Plant Interact. 2019, 14, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Hussain, B.; Ashraf, M.N.; Rahman, S.U.; Abbas, A.; Li, J.; Farooq, M. Cd stress in paddy fields: Effects of soil conditions and remediation strategies. Sci. Total Environ. 2021, 754, 142188. [Google Scholar] [CrossRef] [PubMed]
- Adrees, M.; Khan, Z.; Ali, S.; Hafeez, M.; Khalid, S.; Zia-ur-Rehman, M.; Hussain, A.; Hussain, K.; Chatha, S.A.S.; Rizwan, M. Simultaneous mitigation of Cd and drought stress in wheat by soil application of iron nanoparticles. Chemosphere 2020, 238, 124681. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, R.; Khan, N. Cd toxicity in plants and role of mineral nutrients in its alleviation. Am. J. Plant Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef] [Green Version]
- Hermans, C.; Chen, J.; Coppens, F.; Inzé, D.; Verbruggen, N. Low magnesium status in plants enhances tolerance to Cd exposure. New Phytol. 2011, 192, 428–436. [Google Scholar] [CrossRef]
- Lešková, A.; Giehl, R.F.H.; Hartmann, A.; Fargašová, A.; Wirén, N.V. Heavy metals induce iron deficiency responses at different hierarchic and regulatory levels. Plant Physiol. 2017, 174, 1648–1668. [Google Scholar] [CrossRef] [Green Version]
- Hasan, S.A.; Fariduddin, Q.; Ali, B.; Hayat, S.; Ahmad, A. Cd: Toxicity and tolerance in plants. J. Environ. Biol. 2009, 30, 165–174. [Google Scholar] [PubMed]
- Muneer, S.; Kim, T.H.; Qureshi, M.I. Fe modulates Cd-induced oxidative stress and the expression of stress response proteins in the nodules of mungbean. Plant Growth Regul. 2012, 68, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, P.; Abdel Latef, A.A.; Abid Allah, E.F.; Hashem, A.; Sarwat, M.; Anjum, N.A.; Gucel, S. Calcium and potassium supplementation enhanced growth, osmolyte secondary metabolite production, and enzymatic antioxidant machinery in Cd-exposed chickpea (Cicer arietinum L.). Front. Plant Sci. 2016, 7, 513–520. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, L.; Yang, L.; Li, H. Dynamics of rhizosphere properties and antioxidative responses in wheat (Triticum aestivum L.) under Cd stress. Ecotoxicol. Environ. Saf. 2014, 102, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, R.K.; Pandey, P.; Rajpoot, R.; Rani, A.; Dubey, R.S. Cd and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings. Protoplasma 2014, 251, 1047–1065. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M.N.V. Iron-and manganese-assisted Cd tolerance in Oryza sativa L.: Lowering of rhizotoxicity next to functional photosynthesis. Planta 2015, 241, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Yadava, D.K.; Hussain, F.; Mohapatra, T. Nutritional security through crop biofortification in India: Status and future prospects. Indian J. Med. Res. 2018, 148, 621–631. [Google Scholar]
- Velikova, V.; Yordanova, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, peroxidase and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [Green Version]
- Aebi, H.E. Catalase. In Methods in Enzymatic Analysis; Bergmeyer, U.S., Ed.; Verlag Chemie: Weinheim, Germany, 1983; Volume 1, pp. 273–286. [Google Scholar]
- Dhindsa, R.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence correlated with increased level of membrane permeability, lipid peroxidation and decreased level of SOD and CAT. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Rao, M.V. Cellular detoxification mechanisms to determine age dependent injury in tropical plant exposed to SO2. J. Plant Physiol. 1992, 140, 733–740. [Google Scholar] [CrossRef]
- Alexander, P.D.; Alloway, B.J.; Dourado, A.M. Genotypic variations in the accumulation of Cd, Cu, Pb and Zn exhibited by six commonly grown vegetables. Environ. Pollut. 2006, 144, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to Cd stress. J. Plant Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.H.; Al-Wahabi, M.; Sakran, A.; Basalah, M.; Ali, H. Effect of calcium and potassium on antioxidant system of Vicia faba L. under Cd stress. Int. J. Mol. Sci. 2012, 13, 6604–6619. [Google Scholar] [CrossRef] [Green Version]
- Muneer, S.; Kim, T.H.; Choi, B.C.; Lee, B.S.; Lee, J.H. Effect of CO, NOx and SO2 on ROS production, photosynthesis and ascorbate-glutathione pathway to induce fragaria × annasa as a hyperaccumulator. Redox Biol. 2014, 2, 91–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero-Puertas, M.C.; Corpas, F.J.; Rodriguez-Serrano, M.; Gomez, M.; Del Rio, L.A.; Sandalio, L.M. Differential expression and regulation of antioxidative enzymes by Cd in pea plants. J. Plant Physiol. 2007, 164, 1346–1357. [Google Scholar] [CrossRef]
- Głowacka, K.; Źróbek-Sokolnik, A.; Okorski, A.; Najdzion, J. The effect of cadmium on the activity of stress-related enzymes and the ultrastructure of pea roots. Plants 2019, 8, 413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, M.I.; Qadir, S.; Zolla, L. Proteomics-based dissection of stress-responsive pathways in plants. J. Plant Physiol. 2007, 164, 1239–1260. [Google Scholar] [CrossRef]
- Astolfi, S.; Zuchi, S.; Hubberten, H.M.; Pinton, R.; Hoefgen, R. Supply of sulphur to S-deficient young barley seedlings restores their capability to cope with iron shortage. J. Exp. Bot. 2010, 61, 799–806. [Google Scholar] [CrossRef] [Green Version]
- Shao, G.; Chen, M.; Wang, W.; Mou, R.; Zhang, G. Iron nutrition affects Cd accumulation and toxicity in rice plants. Plant Growth Regul. 2007, 53, 33–42. [Google Scholar] [CrossRef]
- Sharma, S.S.; Kaul, S.; Metwally, A.; Goyal, K.C.; Finkemeier, I.; Dietz, K.J. Cd toxicity to barley (Hordeum vulgare) as affected by varying Fe nutritional status. Plant Sci. 2004, 166, 1287–1295. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | HP | MDA | POX | CAT | SOD | GR |
|---|---|---|---|---|---|---|
| Treatment | 728.59 ** | 111.71 ** | 587.71 ** | 47.55 ** | 238.04 ** | 28.63 ** |
| Genotype | 32.55 ** | 201.35 ** | 374.29 ** | 30.75 ** | 133.79 ** | 32.19 ** |
| Treatment × Genotype | 77.26 ** | 14.18 ** | 67.68 ** | 22.47 ** | 69.76 ** | 10.27 ** |
| Source of Variation | TL | RL | SL | FW | DW | CdC |
|---|---|---|---|---|---|---|
| Treatment | 94.93 ** | 9.21 ** | 58.86 ** | 368.26 ** | 45.80 ** | 3483.91 ** |
| Genotype | 144.04 ** | 84.88 ** | 88.54 ** | 98.37 ** | 64.80 ** | 871.36 ** |
| Treatment × Genotype | 21.32 ** | 3.85 * | 2.52 * | 3.86 * | 2.90 * | 430.83 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bansal, R.; Priya, S.; Dikshit, H.K.; Jacob, S.R.; Rao, M.; Bana, R.S.; Kumari, J.; Tripathi, K.; Kumar, A.; Kumar, S.; et al. Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress. Toxics 2021, 9, 182. https://doi.org/10.3390/toxics9080182
Bansal R, Priya S, Dikshit HK, Jacob SR, Rao M, Bana RS, Kumari J, Tripathi K, Kumar A, Kumar S, et al. Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress. Toxics. 2021; 9(8):182. https://doi.org/10.3390/toxics9080182
Chicago/Turabian StyleBansal, Ruchi, Swati Priya, Harsh Kumar Dikshit, Sherry Rachel Jacob, Mahesh Rao, Ram Swaroop Bana, Jyoti Kumari, Kuldeep Tripathi, Ashok Kumar, Shiv Kumar, and et al. 2021. "Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress" Toxics 9, no. 8: 182. https://doi.org/10.3390/toxics9080182
APA StyleBansal, R., Priya, S., Dikshit, H. K., Jacob, S. R., Rao, M., Bana, R. S., Kumari, J., Tripathi, K., Kumar, A., Kumar, S., & H. M. Siddique, K. (2021). Growth and Antioxidant Responses in Iron-Biofortified Lentil under Cadmium Stress. Toxics, 9(8), 182. https://doi.org/10.3390/toxics9080182