
Nutrients, Phytochemicals, and In Vitro Antioxidant and Antimicrobial Activities of Lulo (Solanum quitoense Lam.) Fruit Pulp, Peel, and Seeds

 ,
,  , ,
, ,  , ,
, ,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals, Standards, and Biological Materials
2.2. Plant Material
2.3. Compositional Analysis
2.3.1. Centesimal Composition and Energy
2.3.2. Soluble Sugars and Organic Acids
2.3.3. Fatty Acids and Tocopherols
2.3.4. Mineral Composition
2.4. Preparation of Hydroethanolic Extracts
2.5. HPLC-DAD-ESI/MS Analysis of Phenolic Compounds
2.6. Evaluation of Bioactive Properties
2.6.1. In Vitro Cell-Based Antioxidant Activity
2.6.2. Antimicrobial Activity Against Foodborne Pathogens
2.7. Statistical Analysis
3. Results and Discussion
3.1. Nutritional Composition
3.1.1. Centesimal Composition and Energy
3.1.2. Soluble Sugars and Organic Acids
3.1.3. Tocopherols and Fatty Acids
3.1.4. Mineral Composition
3.2. Phenolic Composition
3.3. Bioactive Properties
3.3.1. In Vitro Antioxidant Activity
3.3.2. Antimicrobial Activity
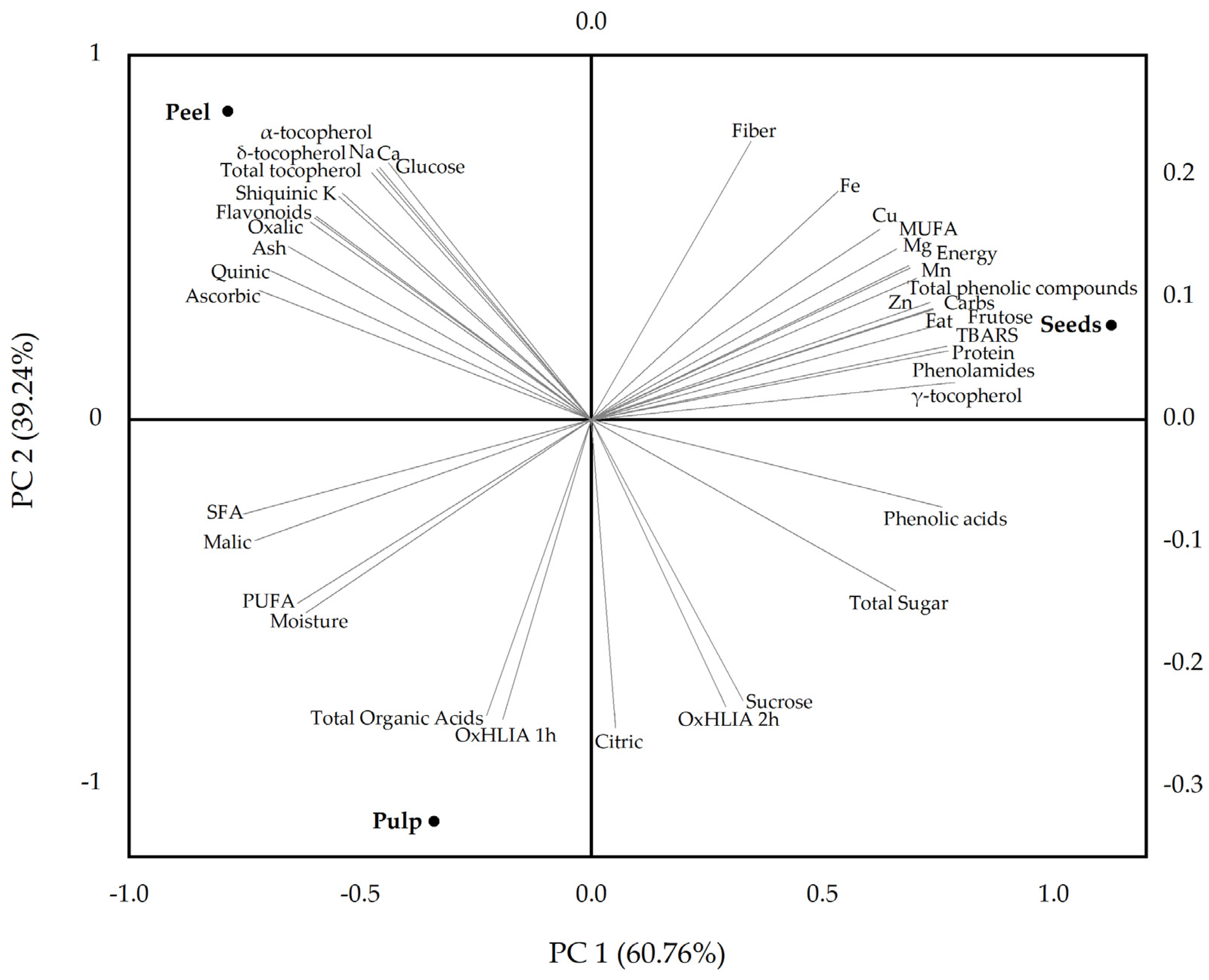
3.4. Multivariate Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Obregón, A.J.; López, M.D.; Angeles, D. Nutritional and Bioactive Properties of Solanum quitoense Lam.: Native Fruit from the South American Andes. J. Microbiol. Biotechnol. Food Sci. 2024, 13, e10386. [Google Scholar] [CrossRef]
- Andrade-Cuvi, M.J.; Guijarro-Fuertes, M.; Concellón, A.; Vernaza, M.G.; Bravo-Vásquez, J. Quality and Bioaccessibility of Antioxidants of Bread Enriched with Naranjilla (Solanum quitoense) Fruit. Nutr. Food Sci. 2021, 51, 1282–1298. [Google Scholar] [CrossRef]
- Igual, M.; Ramires, S.; Mosquera, L.H.; Martínez-Navarrete, N. Optimization of Spray Drying Conditions for Lulo (Solanum quitoense L.) Pulp. Powder Technol. 2014, 256, 233–238. [Google Scholar] [CrossRef]
- Obregón La Rosa, A.; Lozano Zanelly, G.A. Compuestos Nutricionales y Bioactivos de Tres Frutas Provenientes de La Sierra y La Selva de Perú Como Fuente Potencial de Nutrientes Para La Alimentación Humana. Cienc. Tecnol. Agropecu. 2021, 22, e1835. [Google Scholar] [CrossRef]
- Gancel, A.L.; Alter, P.; Dhuique-Mayer, C.; Ruales, J.; Vaillant, F. Identifying Carotenoids and Phenolic Compounds in Naranjilla (Solanum quitoense Lam. var. Puyo Hybrid), an Andean Fruit. J. Agric. Food Chem. 2008, 56, 11892–11899. [Google Scholar] [CrossRef]
- Jiang, D.; Guo, Y.; Niu, C.; Long, S.; Jiang, Y.; Wang, Z.; Wang, X.; Sun, Q.; Ling, W.; An, X.; et al. Exploration of the Antioxidant Effect of Spermidine on the Ovary and Screening and Identification of Differentially Expressed Proteins. Int. J. Mol. Sci. 2023, 24, 5793. [Google Scholar] [CrossRef]
- Niechcial, A.; Schwarzfischer, M.; Wawrzyniak, M.; Atrott, K.; Laimbacher, A.; Morsy, Y.; Katkeviciute, E.; Häfliger, J.; Westermann, P.; Akdis, C.A.; et al. Spermidine Ameliorates Colitis via Induction of Anti-Inflammatory Macrophages and Prevention of Intestinal Dysbiosis. J. Crohns Colitis 2023, 17, 1489–1503. [Google Scholar] [CrossRef]
- Walters, D.; Meurer-Grimes, B.; Rovira, I. Antifungal Activity of Three Spermidine Conjugates. FEMS Microbiol. Lett. 2006, 201, 255–258. [Google Scholar] [CrossRef]
- Büttner, S.; Broeskamp, F.; Sommer, C.; Markaki, M.; Habernig, L.; Alavian-Ghavanini, A.; Carmona-Gutierrez, D.; Eisenberg, T.; Michael, E.; Kroemer, G.; et al. Spermidine Protects against α-Synuclein Neurotoxicity. Cell Cycle 2014, 13, 3903–3908. [Google Scholar] [CrossRef]
- Wang, I.F.; Tsai, K.J.; Shen, C.K.J. Spermidine on neurodegenerative diseases. Cell Cycle 2015, 14, 697–698. [Google Scholar] [CrossRef]
- Forero, D.P.; Masatani, C.; Fujimoto, Y.; Coy-Barrera, E.; Peterson, D.G.; Osorio, C. Spermidine Derivatives in Lulo (Solanum quitoense Lam.) Fruit: Sensory (Taste) versus Biofunctional (ACE-Inhibition) Properties. J. Agric. Food Chem. 2016, 64, 5375–5383. [Google Scholar] [CrossRef]
- Gęgotek, A.; Skrzydlewska, E. Chapter Nine—Ascorbic Acid as Antioxidant. In Antioxidants; Litwack, G., Ed.; Vitamins and Hormones; Academic Press: New York, NY, USA, 2023; Volume 121, pp. 247–270. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 20th ed.; Latimer, G.W., Ed.; AOAC International: Gaithersburg, MD, USA, 2016. [Google Scholar]
- Regulation (EU) No 1169/2011 of the European Parliament and of the Council. 2011. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2011:304:0018:0063:en:PDF (accessed on 28 November 2024).
- Barros, L.; Pereira, E.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Bioactivity and Chemical Characterization in Hydrophilic and Lipophilic Compounds of Chenopodium ambrosioides L. J. Funct. Foods 2013, 5, 1732–1740. [Google Scholar] [CrossRef]
- Añibarro-Ortega, M.; Pinela, P.; Barros, L.; Ćirić, A.; Silva, S.P.; Coelho, E.; Mocan, A.; Calhelha, R.C.; Soković, M.; Coimbra, M.A.; et al. Compositional Features and Bioactive Properties of Aloe vera Leaf (Fillet, Mucilage, and Rind) and Flower. Antioxidants 2019, 8, 444. [Google Scholar] [CrossRef]
- Soković, M.; Glamočlija, J.; Marin, P.D.; Brkić, D.; van Griensven, L.J.L.D. Antibacterial Effects of the Essential Oils of Commonly Consumed Medicinal Herbs Using an In Vitro Model. Molecules 2010, 15, 7532–7546. [Google Scholar] [CrossRef]
- Soković, M.; van Griensven, L.J.L.D. Antimicrobial Activity of Essential Oils and Their Components against the Three Major Pathogens of the Cultivated Button Mushroom. Agaricus Bisporus. Eur. J. Plant Pathol. 2006, 116, 211–224. [Google Scholar] [CrossRef]
- USDA FoodData Central. Available online: https://fdc.nal.usda.gov/ (accessed on 23 January 2022).
- Acosta, O.; Mercedes Pérez, A. Chemical Characterization, Antioxidant Properties, and Volatile Constituents of Naranjilla (Solanum quitoense Lam.) Cultivated in Costa Rica. Arch. Latinoam. Nutr. 2009, 59, 88–94. [Google Scholar]
- The European Parliament and The Council Regulation (EC). No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on Nutrition and Health Claims Made on Foods. Off. J. Eur. Union. 2006, 12, 3–18. [Google Scholar]
- Elleuch, M.; Bedigian, D.; Roiseux, O.; Besbes, S.; Blecker, C.; Attia, H. Dietary Fibre and Fibre-Rich By-Products of Food Processing: Characterisation, Technological Functionality and Commercial Applications: A Review. Food Chem. 2011, 124, 411–421. [Google Scholar] [CrossRef]
- Forero, D.P.; Carriazo, J.G.; Osorio, C. Effect of Different Drying Methods on Morphological, Thermal, and Biofunctional Properties of Lulo (Solanum quitoense Lam.) Fruit Powders. Dry. Technol. 2016, 34, 1085–1094. [Google Scholar] [CrossRef]
- Li, J.; Zhang, C.; Liu, H.; Liu, J.; Jiao, Z. Profiles of Sugar and Organic Acid of Fruit Juices: A Comparative Study and Implication for Authentication. J. Food Qual. 2020, 2020, 7236534. [Google Scholar] [CrossRef]
- Sánchez-Capa, M.; Corell González, M.; Mestanza-Ramón, C. Edible Fruits from the Ecuadorian Amazon: Ethnobotany, Physicochemical Characteristics, and Bioactive Components. Plants 2023, 12, 3635. [Google Scholar] [CrossRef]
- European Food Safety Authority. Dietary Reference Values for Vitamin C. EFSA J. 2013, 11, 3418. [Google Scholar] [CrossRef]
- Añibarro-Ortega, M.; Dias, M.I.; Petrović, J.; Mandim, F.; Núñez, S.; Soković, M.; López, V.; Barros, L.; Pinela, J. Nutrients, Phytochemicals, and In Vitro Biological Activities of Goldenberry (Physalis peruviana L.) Fruit and Calyx. Plants 2025, 14, 327. [Google Scholar] [CrossRef]
- Petkova, N.T.; Popova, V.T.; Ivanova, T.A.; Mazova, N.N.; Panayotov, N.D.; Stoyanova, A. Nutritional Composition of Different Cape Gooseberry Genotypes (Physalis peruviana L.)—A Comparative Study. Food Res. 2021, 5, 191–202. [Google Scholar] [CrossRef]
- Ohkatsu, Y.; Kajiyama, T.; Arai, Y. Antioxidant Activities of Tocopherols. Polym. Degrad. Stab. 2001, 72, 303–311. [Google Scholar] [CrossRef]
- Sakurai, K.; Shen, C.; Shiraishi, I.; Inamura, N.; Hisatsune, T. Consumption of Oleic Acid on the Preservation of Cognitive Functions in Japanese Elderly Individuals. Nutrients 2021, 13, 284. [Google Scholar] [CrossRef]
- Song, W.; Zhang, K.; Xue, T.; Han, J.; Peng, F.; Ding, C.; Lin, F.; Li, J.; Sze, F.T.A.; Gan, J.; et al. Cognitive Improvement Effect of Nervonic Acid and Essential Fatty Acids on Rats Ingesting Acer truncatum Bunge Seed Oil Revealed by Lipidomics Approach. Food Funct. 2022, 13, 2475–2490. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Lucci, P.; Núñez, O.; Tundis, R.; Balzano, M.; Frega, N.G.; Conte, L.; Moret, S.; Filatova, D.; Moyano, E.; et al. Native Colombian Fruits and Their By-Products: Phenolic Profile, Antioxidant Activity and Hypoglycaemic Potential. Foods 2019, 8, 89. [Google Scholar] [CrossRef]
- Cerón, A.F.; Osorio, M.O.; Hurtado, B.A. Identification of Fatty Acids Contained in the Oils Extracted from Seeds of Three Different Species of Fruit. Acta Agron. 2012, 61, 126–132. [Google Scholar]
- Swain, J.H.; Minisola, S.; Razzaque, M.S.; Wimalawansa, S.J. Minerals and Human Health: From Deficiency to Toxicity. Nutrients 2025, 17, 454. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Dietary Reference Values for Potassium. EFSA J. 2016, 14, e04592. [Google Scholar] [CrossRef]
- Wu, S.B.; Meyer, R.S.; Whitaker, B.D.; Litt, A.; Kennelly, E.J. A New Liquid Chromatography-Mass Spectrometry-Based Strategy to Integrate Chemistry, Morphology, and Evolution of Eggplant (Solanum) Species. J. Chromatogr. A 2013, 1314, 154–172. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Zheng, W.; Kuhnert, N. Profiling the Chlorogenic Acids of Aster by HPLC-MSn. Phytochem. Anal. 2006, 17, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Johnston, K.L.; Knight, S.; Kuhnert, N. Hierarchical Scheme for LC-MSn Identification of Chlorogenic Acids. J. Agric. Food Chem. 2003, 51, 2900–2911. [Google Scholar] [CrossRef]
- Caleja, C.; Barros, L.; Antonio, A.L.; Ciric, A.; Soković, M.; Oliveira, M.B.P.P.; Santos-Buelga, C.; Ferreira, I.C.F.R. Foeniculum vulgare Mill. As Natural Conservation Enhancer and Health Promoter by Incorporation in Cottage Cheese. J. Funct. Foods 2015, 12, 428–438. [Google Scholar] [CrossRef]
- Ferreres, F.; Figueiredo, R.; Bettencourt, S.; Carqueijeiro, I.; Oliveira, J.; Gil-Izquierdo, A.; Pereira, D.M.; Valentão, P.; Andrade, P.B.; Duarte, P.; et al. Identification of Phenolic Compounds in Isolated Vacuoles of the Medicinal Plant Catharanthus roseus and Their Interaction with Vacuolar Class III Peroxidase: An H2O2 Affair? J. Exp. Bot. 2011, 62, 2841–2854. [Google Scholar] [CrossRef]
- Miguel, M.; Barros, L.; Pereira, C.; Calhelha, R.C.; Garcia, P.A.; Castro, M.Á.; Santos-Buelga, C.; Ferreira, I.C.F.R. Chemical Characterization and Bioactive Properties of Two Aromatic Plants: Calendula officinalis L. (Flowers) and Mentha cervina L. (Leaves). Food Funct. 2016, 7, 2223–2232. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Pietta, P. Flavonoids as Antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Chen, C. Sinapic Acid and Its Derivatives as Medicine in Oxidative Stress-Induced Diseases and Aging. Oxid. Med. Cell. Longev. 2015, 2016, 3571614. [Google Scholar] [CrossRef]
- Raj, N.D.; Singh, D. A Critical Appraisal on Ferulic Acid: Biological Profile, Biopharmaceutical Challenges and Nano Formulations. Health Sci. Rev. 2022, 5, 100063. [Google Scholar] [CrossRef]
- Shackebaei, D.; Hesari, M.; Ramezani-Aliakbari, S.; Pashaei, M.; Yarmohammadi, F.; Ramezani-Aliakbari, F. Cardioprotective Effect of Naringin against the Ischemia/Reperfusion Injury of Aged Rats. Naunyn Schmiedebergs Arch. Pharmacol. 2024, 397, 1209–1218. [Google Scholar] [CrossRef]
- Bugianesi, R.; Catasta, G.; Spigno, P.; D’Uva, A.; Maiani, G. Naringenin from Cooked Tomato Paste Is Bioavailable in Men. J. Nutr. 2002, 132, 3349–3352. [Google Scholar] [CrossRef]
- Rodrigues, C.A.; Nicácio, A.E.; Jardim, I.C.S.F.; Visentainer, J.V.; Maldaner, L. Determination of Phenolic Compounds in Red Sweet Pepper (Capsicum annuum L.) Using a Modified QuEChERS Method and UHPLC-MS/MS Analysis and Its Relation to Antioxidant Activity. J. Braz. Chem. Soc. 2019, 30, 1229–1240. [Google Scholar] [CrossRef]
- Shahidi, F.; Pinaffi-Langley, A.C.C.; Fuentes, J.; Speisky, H.; de Camargo, A.C. Vitamin E as an Essential Micronutrient for Human Health: Common, Novel, and Unexplored Dietary Sources. Free Radic. Biol. Med. 2021, 176, 312–321. [Google Scholar] [CrossRef]
- Panzella, L. Natural Phenolic Compounds for Health, Food and Cosmetic Applications. Antioxidants 2020, 9, 427. [Google Scholar] [CrossRef] [PubMed]
- De Souza Zanchet, M.Z.; Nardi, G.M.; De Oliveira Souza Bratti, L.; Filippin-Monteiro, F.B.; Locatelli, C. Lycium barbarum Reduces Abdominal Fat and Improves Lipid Profile and Antioxidant Status in Patients with Metabolic Syndrome. Oxid. Med. Cell. Longev. 2017, 2017, 9763210. [Google Scholar] [CrossRef]
- Hinestroza-Córdoba, L.I.; Serna, S.D.; Seguí, L.; Barrera, C.; Betoret, N. Characterization of Powdered Lulo (Solanum quitoense) Bagasse as a Functional Food Ingredient. Foods 2020, 9, 723. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, X.; Jiang, L.; Chu, Y.; Gao, S.; Jiang, X.; Zhang, Y.; Chen, Y.; Luo, S.; Peng, C. The Biological Activity Mechanism of Chlorogenic Acid and Its Applications in Food Industry: A Review. Front. Nutr. 2022, 9, 943911. [Google Scholar] [CrossRef]
- Roumani, M.; Duval, R.E.; Ropars, A.; Risler, A.; Robin, C.; Larbat, R. Phenolamides: Plant Specialized Metabolites with a Wide Range of Promising Pharmacological and Health-Promoting Interests. Biomed. Pharmacother. 2020, 131, 110762. [Google Scholar] [CrossRef]
- Tenea, G.N.; Hurtado, P.; Ortega, C. A Novel Weissella cibaria Strain UTNGt21O Isolated from Wild Solanum quitoense Fruit: Genome Sequence and Characterization of a Peptide with Highly Inhibitory Potential toward Gram-Negative Bacteria. Foods 2020, 9, 1242. [Google Scholar] [CrossRef] [PubMed]
- Jain, P.; Kumari, S.; Malik, A. Inhibition of Pyocyanin Production in Pseudomonas Aeruginosa by Natural Antimicrobial Compounds from Herbal Extracts. Asian J. Pharm. Clin. Res. 2017, 10, 389–392. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Constituent | Content (Per 100 g fw) | |||
|---|---|---|---|---|
| Peel | Pulp | Seeds | Edible Part * | |
| Moisture (g) | 71 ± 3 b | 87 ± 1 a | 51 ± 3 c | 82 |
| Proteins (g) | 2.06 ± 0.03 b | 1.22 ± 0.03 c | 4.37 ± 0.06 a | 1.64 |
| Ash (g) | 1.87 ± 0.03 a | 1.22 ± 0.06 b | 1.03 ± 0.06 c | 1.19 |
| Crude fat (g) | 0.49 ± 0.01 b | 0.098 ± 0.002 c | 2.90 ± 0.07 a | 0.48 |
| Dietary fiber | 16.5 ± 0.8 b | 4.5 ± 0.3 c | 19.4 ± 0.6 a | 6.5 |
| Carbohydrates (g) | 7.3 ± 0.1 b | 5.9 ± 0.4 c | 20.9 ± 0.4 a | 7.9 |
| Energy (kcal) | 77 ± 2 b | 39 ± 3 c | 167 ± 3 a | 56 |
| Fructose (g) | 2.2 ± 0.1 b | 2.11 ± 0.04 b | 3.0 ± 0.2 a | 2.23 |
| Glucose (g) | 1.0 ± 0.1 a | 0.97 ± 0.02 a | 0.98 ± 0.03 a | 0.97 |
| Sucrose (g) | 1.33 ± 0.04 c | 2.7 ± 0.1 a | 2.26 ± 0.04 b | 2.64 |
| Total sugar (g) | 4.5 ± 0.2 c | 5.8 ± 0.2 b | 6.2 ± 0.2 a | 5.9 |
| Oxalic acid (mg) | 134 ± 11 a | 60 ± 3 b | 49 ± 2 c | 68 |
| Quinic acid (mg) | 559 ± 43 a | 180 ± 4 b | nd | 156 |
| Malic acid (mg) | 220 ± 16 b | 272 ± 13 a | nd | 235 |
| Shikimic acid (mg) | 40 ± 2 | nd | nd | - |
| Citric acid (g) | 1.57 ± 0.02 c | 4.2 ± 0.3 a | 2.53 ± 0.06 b | 3.99 |
| Ascorbic acid (mg) | 25.2 ± 0.3 a | 9.7 ± 0.2 b | nd | 8.4 |
| Total organic acids (g) | 2.56 ± 0.09 b | 4.7 ± 0.3 a | 2.58 ± 0.06 b | 4.45 |
| α-Tocopherol (mg) | 7.9 ± 0.1 a | 0.37 ± 0.01 b | 0.25 ± 0.02 c | 0.35 |
| β-Tocopherol (mg) | nd | nd | nd | nd |
| γ-Tocopherol (mg) | nd | 0.197 ± 0.004 b | 1.7 ± 0.1 a | 0.4 |
| δ-Tocopherol (mg) | 5.12 ± 0.03 | nd | nd | nd |
| Total tocopherols (mg) | 13.0 ± 0.1 a | 0.57 ± 0.01 c | 2.0 ± 0.1 b | 0.76 |
| Fatty Acid | Peel (Relative %) | Pulp (Relative %) | Seeds (Relative %) |
|---|---|---|---|
| C13:0 | 0.51 ± 0.03 | nd | nd |
| C14:0 | 1.07 ± 0.07 a | 0.45 ± 0.01 b | 0.30 ± 0.01 c |
| C14:1 | 0.20 ± 0.01 | nd | nd |
| C15:0 | 0.46 ± 0.04 a | 0.35 ± 0.01 b | 0.11 ± 0.01 c |
| C16:0 | 25.7 ± 0.2 b | 24.0 ± 0.6 c | 28.0 ± 0.4 a |
| C16:1 | 1.8 ± 0.1 b | 0.23 ± 0.01 c | 4.85 ± 0.05 a |
| C17:0 | 0.52 ± 0.03 b | 0.70 ± 0.06 a | 0.55 ± 0.02 b |
| C17:1 | 0.08 ± 0.01 b | nd | 0.25 ± 0.02 a |
| C18:0 | 6.2 ± 0.2 b | 7.0 ± 0.4 a | 0.52 ± 0.03 c |
| C18:1n-9c | 19.9 ± 0.4 b | 6.6 ± 0.2 c | 44.9 ± 0.9 a |
| C18:1n-9t | nd | nd | 10.3 ± 0.5 |
| C18:2n-6c | 9.0 ± 0.1 b | 23.9 ± 0.6 a | nd |
| C18:2n-6t | nd | 0.16 ± 0.01 | nd |
| C18:3n-6 | nd | nd | 0.48 ± 0.04 |
| C18:3n-3 | 23.6 ± 0.3 b | 28.1 ± 0.4 a | 5.6 ± 0.1 c |
| C20:0 | 2.08 ± 0.04 b | 2.49 ± 0.08 a | 1.08 ± 0.05 c |
| C20:1 | 0.59 ± 0.02 a | nd | 0.37 ± 0.03 b |
| C20:2 | 0.50 ± 0.02 b | nd | 0.30 ± 0.01 a |
| C21:0 | 0.34 ± 0.01 b | 0.18 ± 0.01 a | nd |
| C20:3n-3 | 0.42 ± 0.03 | nd | nd |
| C22:0 | 1.46 ± 0.04 b | 2.20 ± 0.03 a | 0.95 ± 0.04 c |
| C20:5n-3 | 0.18 ± 0.01 | nd | nd |
| C23:0 | 0.90 ± 0.06 a | 0.52 ± 0.01 b | 0.34 ± 0.01 c |
| C24:0 | 0.74 ± 0.01 c | 2.92 ± 0.06 a | 1.07 ± 0.08 b |
| C24:1 | 3.5 ± 0.2 | nd | nd |
| Fatty acid class | |||
| SFA | 40.2 ± 0.8 a | 40.98 ± 0.06 a | 32.9 ± 0.5 b |
| MUFA | 26.1 ± 0.4 b | 6.9 ± 0.2 c | 60.7 ± 0.4 a |
| PUFA | 33.7 ± 0.4 b | 52.2 ± 0.2 a | 6.42 ± 0.09 c |
| PUFA/SFA | 0.84 ± 0.02 b | 1.27 ± 0.01 a | 0.2 ± 0.01 c |
| Element | Content (Per 100 g fw) | |||
|---|---|---|---|---|
| Peel | Pulp | Seeds | Edible Part * | |
| K (mg) | 899 ± 30 a | 459 ± 11 b | 403 ± 20 c | 451 |
| Na (mg) | 10.8 ± 0.7 a | 4.0 ± 0.2 c | 4.9 ± 0.3 b | 4.1 |
| Ca (mg) | 35 ± 2 a | 4.8 ± 0.2 c | 9.1 ± 0.2 b | 5.4 |
| Mg (mg) | 42 ± 2 b | 18.8 ± 0.4 c | 96 ± 5 a | 29 |
| Fe (mg) | 1.30 ± 0.02 b | 0.26 ± 0.01 c | 2.08 ± 0.07 a | 0.51 |
| Mn (mg) | 0.38 ± 0.01 b | 0.079 ± 0.003 c | 1.25 ± 0.06 a | 0.24 |
| Cu (mg) | 0.25 ± 0.01 b | 0.124 ± 0.004 c | 0.42 ± 0.03 a | 0.16 |
| Zn (mg) | 0.68 ± 0.03 b | 0.18 ± 0.01 a | 3.5 ± 0.2 c | 0.63 |
| Peak | Rt (min) | λmax (nm) | [M-H]− (m/z) | MS2 (m/z) | Tentative Identification | Reference | Content (mg/g Extract) | ||
|---|---|---|---|---|---|---|---|---|---|
| Peel | Pulp | Seed | |||||||
| 1 | 5.44 | 328 | 341 | 179(100), 135(12) | Caffeic acid hexoside | [37,38] | - | 0.35 ± 0.01 | - |
| 2 | 6.76 | 326 | 353 | 191(100), 179(39) | 3-O-Caffeoylquinic acid | [5,37,38] | 1.57 ± 0.05 a | 0.67 ± 0.01 b | 0.53 ± 0.01 c |
| 3 | 7.03 | 326 | 353 | 191(100), 179(7) | cis-5-O-Caffeoylquinic acid | [5,37,38] | 3.7 ± 0.2 a | 2.47 ± 0.04 b | 1.47 ± 0.02 c |
| 4 | 7.34 | 326 | 353 | 191(100), 179(12) | trans-5-O-Caffeoylquinic acid | [5,37,38] | 8.60 ± 0.09 b | 12.1 ± 0.4 a | 1.15 ± 0.02 c |
| 5 | 9.02 | 276 | 289 | 159(100), 175(38), 131(20) | (-)-Epicatechin | [39] | - | 1.00 ± 0.01 a | 0.202 ± 0.004 b |
| 6 | 9.20 | 328 | 355 | 193(100) | Ferulic acid hexoside | MS/DAD | - | 1.69 ± 0.06 b | 7.1 ± 0.1 a |
| 7 | 9.73 | 331 | 385 | 223(100), 207(100), 161(23) | Sinapic acid hexoside | MS/DAD | - | 1.70 ± 0.05 b | 7.0 ± 0.1 a |
| 8 | 10.38 | 325 | 517 | 193(100), 134(70), 178(40) | Ferulic acid dihexoside | MS/DAD | - | - | 7.1 ± 0.1 |
| 9 | 11.19 | 349 | 639 | 477(100), 315(20) | Isorhamnetin-O-dihexoside | [5] | 0.30 ± 0.01 | - | - |
| 10 | 13.68 | 343 | 771 | 609(100), 301(32) | Quercetin-O-hexosyl-deoxyhexosyl-hexoside | [5] | 1.01 ± 0.01 | - | - |
| 11 | 14.23 | 342 | 625 | 463(100), 301(23) | Quercetin-O-dihexoside | MS/DAD | 0.97 ± 0.01 | - | - |
| 12 | 14.56 | 354 | 755 | 591(100), 301(93) | Quercetin-O-dideoxyhexosyl-hexoside | [40] | 2.19 ± 0.07 a | 0.88 ± 0.01 b | - |
| 13 | 15.43 | 356 | 639 | 477(56), 315(100) | Isorhamnetin-O-dihexoside | [5] | 1.10 ± 0.01 | - | - |
| 14 | 16.18 | 353 | 609 | 301(100) | Quercetin-O-deoxyhexosyl-hexoside | [5] | 1.45 ± 0.04 | - | - |
| 15 | 16.56 | 341 | 739 | 575(100), 285(40), 593(38) | Kaempferol-O-dideoxyhexosyl-hexoside | [40] | 0.97 ± 0.01 | - | - |
| 16 | 16.97 | 351 | 769 | 315(100) | Isorhamnetin-O-deoxyhexosyl-deoxyhexosyl-hexoside | [41] | 1.01 ± 0.02 | - | - |
| 17 | 17.17 | 284 | 595 | 433(100), 271(14) | Naringerin dihexoside | MS/DAD | 1.20 ± 0.01 | - | - |
| 18 | 17.42 | 326 | 799 | 637(100) | N1,N4,N8-tris(dihydrocaffeoyl) spermidine hexoside | [5] | - | - | 9.9 ± 0.2 |
| 19 | 17.66 | 327 | 637 | 473(100) | N1,N4,N8-tris(dihydrocaffeoyl) spermidine | [5] | - | - | 6.66 ± 0.08 |
| 20 | 17.79 | 351 | 609 | 301(100) | Quercetin-3-O-rutinoside | [5] | 1.28 ± 0.04 | - | - |
| 21 | 18.35 | 326 | 799 | 637(100) | N1,N4,N8-tris(dihydrocaffeoyl) spermidine hexoside | [5] | - | - | 6.2 ± 0.1 |
| 22 | 18.57 | 321 | 593 | 285(100), 429(54) | Kaempferol-O-deoxyhexosyl-hexoside | [5] | 1.33 ± 0.02 | - | - |
| 23 | 18.58 | 325 | 637 | 473(100) | N1,N4,N8-tris(dihydrocaffeoyl) spermidine | [5] | - | - | 4.5 ± 0.1 |
| 24 | 19.03 | 351 | 623 | 315(100) | Isorhamnetin-O-deoxyhexosil-hexoside | [5] | 1.41 ± 0.05 | - | - |
| 25 | 21.11 | 328 | 473 | 351(100), 308(88) | N1,N4 or N4,N8-bis(dihydrocaffeoyl) spermidine | [4] | - | - | 10.7 ± 0.3 |
| Σ Phenolic acids | 13.9 ± 0.3 c | 19.0 ± 0.4 b | 24.3 ± 0.5 a | ||||||
| Σ Flavonoids | 14.2 ± 0.4 a | 1.88 ± 0.01 b | 0.20 ± 0.01 c | ||||||
| Σ Phenolamides/HCAA/Spermidine derivatives | - | - | 37.8 ± 0.1 | ||||||
| Σ Phenolic compounds | 28.1 ± 0.7 b | 20.9 ± 0.4 c | 62.3 ± 0.4 a | ||||||
| Bioactivity | Peel Extract | Pulp Extract | Seed Extract | Positive Control | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Antioxidant activity | Trolox | ||||||||||
| OxHLIA (IC50, μg/mL) | Δt 60 min | 202 ± 4 b | 229 ± 4 c | 204 ± 4 b | 19.6 ± 0.7 a | ||||||
| Δt 120 min | 325 ± 4 b | 381 ± 8 c | 360 ± 12 c | 41 ± 1 a | |||||||
| TBARS formation inhibition (EC50, μg/mL) | 23.8 ± 0.8 b | 22.4 ± 0.6 b | 107 ± 4 c | 5.4 ± 0.3 a | |||||||
| Antimicrobial activity | E211 | E224 | |||||||||
| Bacterial strains | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| Bacillus cereus | 12.0 | 18.0 | 6.00 | 12.0 | 6.00 | 12.0 | 0.50 | 0.50 | 2.00 | 4.00 | |
| Staphylococcus aureus | 12.0 | 24.0 | 6.00 | 12.0 | 12.0 | 18.0 | 4.00 | 4.00 | 1.00 | 1.00 | |
| Listeria monocytogenes | 6.00 | 12.0 | 3.00 | 6.00 | 1.50 | 3.00 | 1.00 | 2.00 | 0.50 | 1.00 | |
| Escherichia coli | 12.0 | 24.0 | 6.00 | 12.0 | 12.0 | 18.0 | 1.00 | 2.00 | 0.50 | 1.00 | |
| Enterobacter cloacae | 12.0 | 18.0 | 3.00 | 6.00 | 6.00 | 12.0 | 2.00 | 4.00 | 0.50 | 0.50 | |
| Salmonella enterica subsp. enterica ser. Typhimurium | 12.0 | 18.0 | 6.00 | 12.0 | 12.0 | 18.0 | 1.00 | 2.00 | 1.00 | 1.00 | |
| Fungal strains | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | |
| Aspergillus fumigatus | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | 1.00 | 2.00 | 1.00 | 1.00 | |
| Aspergillus versicolor | 3.00 | 6.00 | 6.00 | 12.0 | 1.50 | 3.00 | 2.00 | 4.00 | 1.00 | 1.00 | |
| Aspergillus niger | 12.0 | 18.0 | 12.0 | 18.0 | 3.00 | 6.00 | 1.00 | 2.00 | 1.00 | 1.00 | |
| Trichoderma viride | 12.0 | 18.0 | 12.0 | 18.0 | 6.00 | 12.0 | 1.00 | 2.00 | 0.50 | 0.50 | |
| Penicillium funiculosum | 12.0 | 24.0 | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | 1.00 | 2.00 | 0.50 | 0.50 | |
| Penicillium verrucosum var. cyclopium | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | ≥24.0 | 2.00 | 4.00 | 1.00 | 1.00 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Añibarro-Ortega, M.; Dias, M.I.; Petrović, J.; Pereira, A.; Soković, M.; Barros, L.; Pinela, J. Nutrients, Phytochemicals, and In Vitro Antioxidant and Antimicrobial Activities of Lulo (Solanum quitoense Lam.) Fruit Pulp, Peel, and Seeds. Foods 2025, 14, 2083. https://doi.org/10.3390/foods14122083
Añibarro-Ortega M, Dias MI, Petrović J, Pereira A, Soković M, Barros L, Pinela J. Nutrients, Phytochemicals, and In Vitro Antioxidant and Antimicrobial Activities of Lulo (Solanum quitoense Lam.) Fruit Pulp, Peel, and Seeds. Foods. 2025; 14(12):2083. https://doi.org/10.3390/foods14122083
Chicago/Turabian StyleAñibarro-Ortega, Mikel, Maria Inês Dias, Jovana Petrović, Alexis Pereira, Marina Soković, Lillian Barros, and José Pinela. 2025. "Nutrients, Phytochemicals, and In Vitro Antioxidant and Antimicrobial Activities of Lulo (Solanum quitoense Lam.) Fruit Pulp, Peel, and Seeds" Foods 14, no. 12: 2083. https://doi.org/10.3390/foods14122083
APA StyleAñibarro-Ortega, M., Dias, M. I., Petrović, J., Pereira, A., Soković, M., Barros, L., & Pinela, J. (2025). Nutrients, Phytochemicals, and In Vitro Antioxidant and Antimicrobial Activities of Lulo (Solanum quitoense Lam.) Fruit Pulp, Peel, and Seeds. Foods, 14(12), 2083. https://doi.org/10.3390/foods14122083