
iPSC Therapy for Myocardial Infarction in Large Animal Models: Land of Hope and Dreams



Abstract

1. Introduction
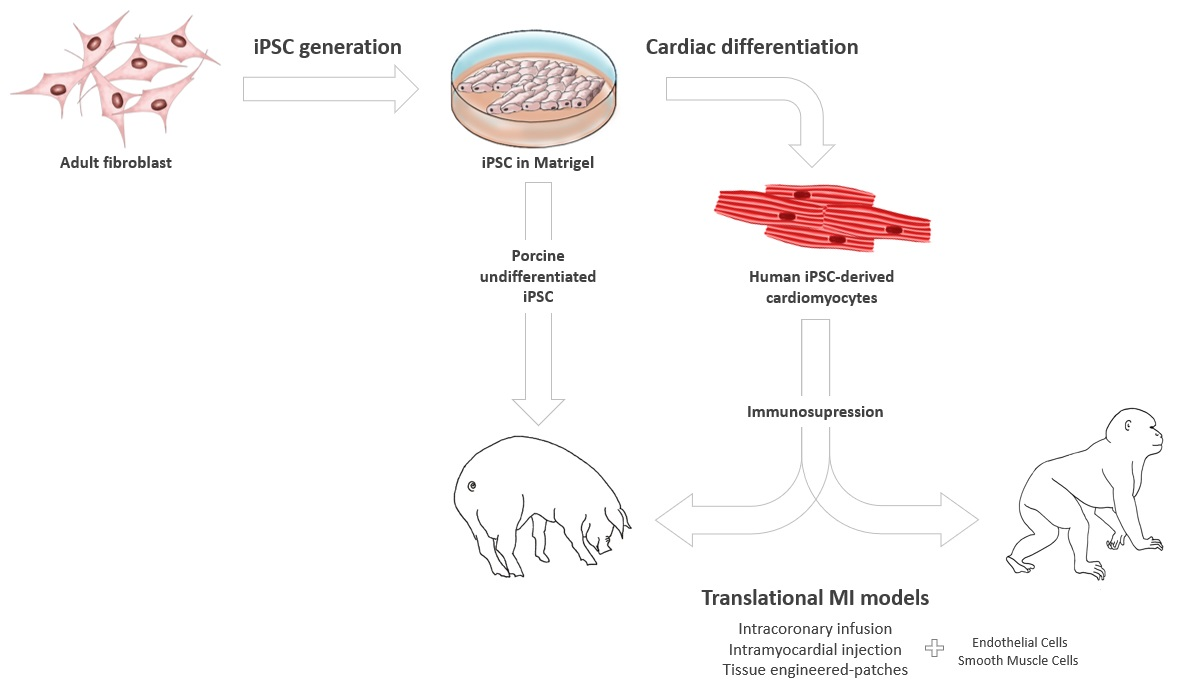
2. iPSC Generation and Cardiac Differentiation
2.1. Obtaining iPSCs from Large Animals
2.2. Cardiac Lineage iPSC Differentiation in Large Animal Models
3. Large Animal Models for Translational Cardiovascular Studies
3.1. The Importance of Large Animal Models in Cardiac Stem Cell Therapies
3.2. Critical Differences between Large and Small Animal Models in Pre-Clinical Studies
3.3. Pre-Clinical Stem Cell Research for Cardiovascular Diseases
4. Applications of iPSC-Based Therapies in Large Animal Models of MI
4.1. Undifferentiated iPSCs
4.2. Differentiated iPSCs
4.2.1. Allogeneic iPSC-CMs: Non-Primate Models
4.2.2. Xenogeneic hiPSC-CMs
4.2.3. Combination of Multiple hiPSC-Cardiovascular Lineage Cell Populations: Cardiomyocytes, Endothelial, and Smooth Muscle Cells
4.2.4. iPSC-Derived Mesenchymal Stem Cells
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Khan, M.A.; Hashim, M.J.; Mustafa, H.; Baniyas, M.Y.; Al Suwaidi, S.K.B.M.; AlKatheeri, R.; Alblooshi, F.M.K.; Almatrooshi, M.E.A.H.; Alzaabi, M.E.H.; Al Darmaki, R.S.; et al. Global Epidemiology of Ischemic Heart Disease: Results from the Global Burden of Disease Study. Cureus 2020, 12, e9349. [Google Scholar] [CrossRef]
- McMurray, J.; Pfeffer, M. Heart failure. Lancet 2005, 365, 1877–1889. [Google Scholar] [CrossRef]
- Koitabashi, N.; Kass, D. Reverse remodeling in heart failure: Mechanisms and therapeutic opportunities. Nat. Rev. Cardio 2011, 9, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Towbin, J.A.; Bowles, N.E. The failing heart. Nature 2002, 415, 227–233. [Google Scholar] [CrossRef]
- Yamashita, J.; Itoh, H.; Hirashima, M.; Ogawa, M.; Nishikawa, S.; Yurugi, T.; Naito, M.; Nakao, K.; Nishikawa, S. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000, 408, 92–96. [Google Scholar] [CrossRef]
- Yamashita, J.K.; Takano, M.; Hiraoka-Kanie, M.; Shimazu, C.; Peishi, Y.; Yanagi, K.; Nakano, A.; Inoue, E.; Kita, F.; Nishikawa, S.-I. Prospective identification of cardiac progenitors by a novel single cell-based cardiomyocyte induction. FASEB J. 2005, 19, 1534–1536. [Google Scholar] [CrossRef] [PubMed]
- Masumoto, H.; Ikuno, T.; Takeda, M.; Fukushima, H.; Marui, A.; Katayama, S.; Shimizu, T.; Ikeda, T.; Okano, T.; Sakata, R.; et al. Human iPS cell-engineered cardiac tissue sheets with cardiomyocytes and vascular cells for cardiac regeneration. Sci. Rep. 2014, 4, 6716. [Google Scholar] [CrossRef]
- Yeung, E.; Fukunishi, T.; Bai, Y.; Bedja, D.; Pitaktong, I.; Mattson, G.; Jeyaram, A.; Lui, C.; Ong, C.S.; Inoue, T.; et al. Cardiac regeneration using human-induced pluripotent stem cell-derived biomaterial-free 3D-bioprinted cardiac patch in vivo. J. Tissue Eng. Regen. Med. 2019, 13, 2031–2039. [Google Scholar] [CrossRef]
- Wendel, J.S.; Ye, L.; Tao, R.; Zhang, J.; Zhang, J.; Kamp, T.J.; Tranquillo, R.T. Functional Effects of a Tissue-Engineered Cardiac Patch From Human Induced Pluripotent Stem Cell-Derived Cardiomyocytes in a Rat Infarct Model. Stem. Cells Transl. Med. 2015, 4, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Xuan, W.; Wang, Y.; Tang, Y.; Ali, A.; Hu, H.; Maienschein-Cline, M.; Ashraf, M. Cardiac Progenitors Induced from Human Induced Pluripotent Stem Cells with Cardiogenic Small Molecule Effectively Regenerate Infarcted Hearts and Attenuate Fibrosis. Shock 2018, 50, 627–639. [Google Scholar] [CrossRef]
- Gao, L.; Kupfer, M.; Jung, J.; Yang, L.; Zhang, P.; da Sie, Y.; Tran, Q.; Ajeti, V.; Freeman, B.; Fast, V.; et al. Myocardial Tissue Engineering With Cells Derived From Human-Induced Pluripotent Stem Cells and a Native-Like, High-Resolution, 3-Dimensionally Printed Scaffold. Circ. Res. 2017, 120, 1318–1325. [Google Scholar] [CrossRef]
- Tsang, H.G.; Rashdan, N.A.; Whitelaw, C.B.A.; Corcoran, B.M.; Summers, K.M.; MacRae, V.E. Large animal models of cardiovascular disease. Cell Biochem. Funct. 2016, 34, 113–132. [Google Scholar] [CrossRef] [PubMed]
- Biswas, A.; Hutchins, R. Embryonic stem cells. Stem Cells Dev. 2007, 16, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Vazin, T.; Freed, W.J. Human embryonic stem cells: Derivation, culture, and differentiation: A review. Restor. Neurol. Neurosci. 2010, 28, 589–603. [Google Scholar] [CrossRef] [PubMed]
- Pourquie, O. Human embryonic stem cells get organized. Nature 2018, 558, 35–36. [Google Scholar] [CrossRef]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Ware, C.B. Concise Review: Lessons from Naïve Human Pluripotent Cells. Stem Cells 2017, 35, 35–41. [Google Scholar] [CrossRef]
- Stirparo, G.G.; Boroviak, T.; Guo, G.; Nichols, J.; Smith, A.; Bertone, P. Integrated analysis of single-cell embryo data yields a unified transcriptome signature for the human pre-implantation epiblast. Development 2018, 145, dev169672. [Google Scholar] [CrossRef]
- Nichols, J.; Smith, A. Naive and primed pluripotent states. Cell Stem Cell 2009, 4, 487–492. [Google Scholar] [CrossRef]
- Rossant, J. Mouse and human blastocyst-derived stem cells: Vive les differences. Development 2015, 142, 9–12. [Google Scholar] [CrossRef]
- Samanta, M.; Kalantry, S. Generating primed pluripotent epiblast stem cells: A methodology chapter. Curr. Top. Dev. Biol. 2020, 138, 139–174. [Google Scholar]
- Nakamura, T.; Yabuta, Y.; Okamoto, I.; Sasaki, K.; Iwatani, C.; Tsuchiya, H.; Saitou, M. Single-cell transcriptome of early embryos and cultured embryonic stem cells of cynomolgus monkeys. Sci. Data 2017, 4, 170067. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, S.; Edsgärd, D.; Reinius, B.; Deng, Q.; Panula, S.P.; Codeluppi, S.; Reyes, A.P.; Linnarsson, S.; Sandberg, R.; Lanner, F. Single-Cell RNA-Seq Reveals Lineage and X Chromosome Dynamics in Human Preimplantation Embryos. Cell 2016, 165, 1012–1026. [Google Scholar] [CrossRef]
- Boroviak, T.; Nichols, J. Primate embryogenesis predicts the hallmarks of human naïve pluripotency. Development 2017, 144, 175–186. [Google Scholar] [CrossRef]
- Boroviak, T.; Loos, R.; Lombard, P.; Okahara, J.; Behr, R.; Sasaki, E.; Nichols, J.; Smith, A.; Bertone, P. Lineage-Specific Profiling Delineates the Emergence and Progression of Naive Pluripotency in Mammalian Embryogenesis. Dev. Cell 2015, 35, 366–382. [Google Scholar] [CrossRef]
- Piliszek, A.; Madeja, Z.E. Pre-implantation Development of Domestic Animals. Curr. Top. Dev. Biol. 2018, 128, 267–294. [Google Scholar]
- Haridhasapavalan, K.K.; Borgohain, M.P.; Dey, C.; Saha, B.; Narayan, G.; Kumar, S.; Thummer, R.P. An insight into non-integrative gene delivery approaches to generate transgene-free induced pluripotent stem cells. Gene 2019, 686, 146–159. [Google Scholar] [CrossRef]
- Cherkashova, E.A.; Leonov, G.E.; Namestnikova, D.D.; Solov’eva, A.A.; Gubskii, I.L.; Bukharova, T.B.; Gubskii, L.V.; Goldstein, D.V.; Yarygin, K.N. Methods of Generation of Induced Pluripotent Stem Cells and Their Application for the Therapy of Central Nervous System Diseases. Bull. Exp. Biol. Med. 2020, 168, 566–573. [Google Scholar] [CrossRef]
- Schlaeger, T.M.; Daheron, L.; Brickler, T.R.; Entwisle, S.; Chan, K.; Cianci, A.; DeVine, A.; Ettenger, A.; Fitzgerald, K.; Godfrey, M.; et al. A comparison of non-integrating reprogramming methods. Nat. Biotechnol. 2015, 33, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, K.; Smuga-Otto, K.; Tian, S.; Stewart, R.; Slukvin, I.I.; Thomson, J.A. Human induced pluripotent stem cells free of vector and transgene sequences. Science 2009, 324, 797–801. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Y.L.; Loh, C.Y.Y. Episomal Induced Pluripotent Stem Cells: Functional and Potential Therapeutic Applica-tions. Cell Transplant 2019, 28, 112S–131S. [Google Scholar] [CrossRef]
- Skowron, K.; Tomsia, M.; Czekaj, P. An experimental approach to the generation of human embryonic stem cells equivalents. Mol. Biotechnol. 2014, 56, 12–37. [Google Scholar] [CrossRef]
- Mansouri, M.; Bellon-Echeverria, I.; Rizk, A.; Ehsaei, Z.; Cianciolo-Cosentino, C.; Silva, C.S.; Xie, Y.; Boyce, F.M.; Davis, M.W.; Neuhauss, S.C.; et al. Highly efficient baculovirus-mediated multigene delivery in primary cells. Nat. Commun. 2016, 7, 11529. [Google Scholar] [CrossRef]
- Ye, H.; Wang, Q. Efficient Generation of Non-Integration and Feeder-Free Induced Pluripotent Stem Cells from Human Peripheral Blood Cells by Sendai Virus. Cell Physiol. Biochem. 2018, 50, 1318–1331. [Google Scholar] [CrossRef] [PubMed]
- Fusaki, N.; Ban, H.; Nishiyama, A.; Saeki, K.; Hasegawa, M. Efficient induction of transgene-free human pluripotent stem cells using a vector based on Sendai virus, an RNA virus that does not integrate into the host genome. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 348–362. [Google Scholar] [CrossRef]
- Ban, H.; Nishishita, N.; Fusaki, N.; Tabata, T.; Saeki, K.; Shikamura, M.; Takada, N.; Inoue, M.; Hasegawa, M.; Kawamata, S.; et al. Efficient generation of transgene-free human induced pluripotent stem cells (iPSCs) by temperature-sensitive Sendai virus vectors. Proc. Natl. Acad. Sci. USA 2011, 108, 14234–14239. [Google Scholar] [CrossRef]
- Okumura, T.; Horie, Y.; Lai, C.Y.; Lin, H.T.; Shoda, H.; Natsumoto, B.; Fujio, K.; Kumaki, E.; Okano, T.; Ono, S.; et al. Robust and highly efficient hiPSC generation from patient non-mobilized peripheral blood-derived CD34+ cells using the auto-erasable Sendai virus vector. Stem Cell Res. Ther. 2019, 10, 185. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Yuasa, S.; Oda, M.; Egashira, T.; Yae, K.; Kusumoto, D.; Nakata, H.; Tohyama, S.; Hashimoto, H.; Kodaira, M.; et al. Generation of induced pluripotent stem cells from human terminally differentiated circulating T cells. Cell Stem Cell. 2010, 7, 11–14. [Google Scholar] [CrossRef]
- Woltjen, K.; Michael, I.P.; Mohseni, P.; Desai, R.; Mileikovsky, M.; Hämäläinen, R.; Cowling, R.; Wang, W.; Liu, P.; Gertsenstein, M.; et al. PiggyBac transposition reprograms fibroblasts to induced pluripotent stem cells. Nature 2009, 458, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Brightwell, S.; Kaji, K. PiggyBac Transposon Mediated Reprogramming and Flow Cytometry Analysis of CD44 and ICAM1 Cell-Surface Marker Changes. Methods Mol. Biol. 2016, 1357, 285–923. [Google Scholar]
- Behringer, R.; Gertsenstein, M.; Nagy, K.V.; Nagy, A. Reprogramming Mouse Fibroblasts with piggyBac Transposons. Cold Spring Harb. Protoc. 2017, 2017, pdb.prot092627. [Google Scholar] [CrossRef] [PubMed]
- Bernal, J.A. RNA-based tools for nuclear reprogramming and lineage-conversion: Towards clinical applications. J. Cardiovasc. Transl. Res. 2013, 6, 956–968. [Google Scholar] [CrossRef]
- Warren, L.; Manos, P.D.; Ahfeldt, T.; Loh, Y.H.; Li, H.; Lau, F.; Ebina, W.; Mandal, P.K.; Smith, Z.D.; Meissner, A.; et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell Stem Cell 2010, 7, 618–630. [Google Scholar] [CrossRef]
- Warren, L.; Lin, C. mRNA-Based Genetic Reprogramming. Mol. Ther. 2019, 27, 729–734. [Google Scholar] [CrossRef]
- Seo, B.J.; Hong, Y.J.; Do, J.T. Cellular Reprogramming Using Protein and Cell-Penetrating Peptides. Int. J. Mol. Sci. 2017, 18, 552. [Google Scholar] [CrossRef]
- Ezashi, T.; Telugu, B.P.; Roberts, R.M. Induced pluripotent stem cells from pigs and other ungulate species: An alternative to embryonic stem cells? Reprod. Domest. Anim. 2012, 47, 92–97. [Google Scholar] [CrossRef]
- Su, Y.; Zhu, J.; Salman, S.; Tang, Y. Induced pluripotent stem cells from farm animals. J. Anim. Sci. 2020, 98, skaa343. [Google Scholar] [CrossRef] [PubMed]
- Ezashi, T.; Telugu, B.P.; Alexenko, A.P.; Sachdev, S.; Sinha, S.; Roberts, R.M. Derivation of induced pluripotent stem cells from pig somatic cells. Proc. Natl. Acad. Sci. USA 2009, 106, 10993–10998. [Google Scholar] [CrossRef] [PubMed]
- Iborra-Egea, O.; Martínez-Falguera, D.; Roura, S.; Bayes-Genis, A.; Raya, Á.; Gálvez-Montón, C. Porcine iPSC generation: Testing different protocols to a successful application. Methods Mol. Biol. 2021. [Google Scholar] [CrossRef]
- Gao, X.; Nowak-Imialek, M.; Chen, X.; Chen, D.; Herrmann, D.; Ruan, D.; Chen, A.C.H.; Eckersley-Maslin, M.A.; Ahmad, S.; Lee, Y.L.; et al. Establishment of porcine and human expanded potential stem cells. Nat. Cell Biol. 2019, 21, 687–699. [Google Scholar] [CrossRef]
- Xu, J.; Yu, L.; Guo, J.; Xiang, J.; Zheng, Z.; Gao, D.; Shi, B.; Hao, H.; Jiao, D.; Zhong, L.; et al. Generation of pig induced pluripotent stem cells using an extended pluripotent stem cell culture system. Stem Cell Res. Ther. 2019, 10, 193. [Google Scholar] [CrossRef]
- Zhong, C.; Wu, J.; Izpisua-Belmonte, J.C. Pig Chimeric Model with Human Pluripotent Stem Cells. Methods Mol. Biol. 2019, 2005, 101–124. [Google Scholar]
- Ezashi, T.; Yuan, Y.; Roberts, R.M. Pluripotent Stem Cells from Domesticated Mammals. Annu. Rev. Anim. Biosci. 2016, 4, 223–253. [Google Scholar] [CrossRef]
- Guo, R.; Morimatsu, M.; Feng, T.; Lan, F.; Chang, D.; Wan, F.; Ling, Y. Stem cell-derived cell sheet transplantation for heart tissue repair in myocardial infarction. Stem Cell Res. Ther. 2020, 11, 19. [Google Scholar] [CrossRef]
- Esteban, M.A.; Xu, J.; Yang, J.; Peng, M.; Qin, D.; Li, W.; Jiang, Z.; Chen, J.; Deng, K.; Zhong, M.; et al. Generation of induced pluripotent stem cell lines from Tibetan miniature pig. J. Biol. Chem. 2009, 284, 17634–17640. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Chen, J.; Ren, J.; Bao, L.; Liao, J.; Cui, C.; Rao, L.; Li, H.; Gu, Y.; Dai, H.; et al. Generation of pig induced pluripotent stem cells with a drug-inducible system. J. Mol. Cell Biol. 2009, 1, 46–54. [Google Scholar] [CrossRef]
- Zhang, Y.; Wei, C.; Zhang, P.; Li, X.; Liu, T.; Pu, Y.; Li, Y.; Cao, Z.; Cao, H.; Liu, Y.; et al. Efficient reprogramming of naïve-like induced pluripotent stem cells from porcine adipose-derived stem cells with a feeder-independent and serum-free system. PLoS ONE 2014, 9, e85089. [Google Scholar]
- Ruan, W.; Han, J.; Li, P.; Cao, S.; An, Y.; Lim, B.; Li, N. A novel strategy to derive iPS cells from porcine fibroblasts. Sci. China Life Sci. 2011, 54, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Guo, Y.; Li, Z.; Liu, Y.; Gao, X.; Gao, Y.; Cheng, X.; Hu, J.; Wang, H. Porcine induced pluripotent stem cells require LIF and maintain their developmental potential in early stage of embryos. PLoS ONE 2012, 7, e51778. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Song, G.; An, X.; Fan, A.; Tan, W.; Tang, B.; Zhang, X.; Li, Z. miRNAs promote generation of porcine-induced pluripotent stem cells. Mol. Cell Biochem. 2014, 389, 209–218. [Google Scholar] [CrossRef]
- Zhang, S.; Guo, Y.; Cui, Y.; Liu, Y.; Yu, T.; Wang, H. Generation of intermediate porcine iPS cells under culture condition favorable for mesenchymal to epithelial transition. Stem. Cell Rev. Rep. 2015, 11, 24–38. [Google Scholar] [CrossRef]
- Kues, W.A.; Herrmann, D.; Barg-Kues, B.; Haridoss, S.; Nowak-Imialek, M.; Buchholz, T.; Streeck, M.; Grebe, A.; Grabundzija, I.; Merkert, S.; et al. Derivation and characterization of sleeping beauty transposon-mediated porcine induced pluripotent stem cells. Stem Cells Dev. 2013, 22, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, A.; Allegrucci, C.; Alberio, R. Modulation of pluripotency in the porcine embryo and iPS cells. PLoS ONE 2012, 7, e49079. [Google Scholar] [CrossRef]
- Chakritbudsabong, W.; Sariya, L.; Pamonsupornvichit, S.; Pronarkngver, R.; Chaiwattanarungruengpaisan, S.; Ferreira, J.N.; Setthawong, P.; Phakdeedindan, P.; Techakumphu, M.; Tharasanit, T.; et al. Generation of a pig induced pluripotent stem cell (piPSC) line from embryonic fibroblasts by incorporating LIN28 to the four transcriptional factor-mediated reprogramming: VSMUi001-D. Stem Cell Res. 2017, 24, 21–24. [Google Scholar] [CrossRef]
- Mao, J.; Zhang, Q.; Deng, W.; Wang, H.; Liu, K.; Fu, H.; Zhao, Q.; Wang, X.; Liu, L. Epigenetic Modifiers Facilitate Induction and Pluripotency of Porcine iPSCs. Stem Cell Rep. 2017, 8, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Nagy, K.; Sung, H.K.; Zhang, P.; Laflamme, S.; Vincent, P.; Agha-Mohammadi, S.; Woltjen, K.; Monetti, C.; Michael, I.P.; Smith, L.C. Induced pluripotent stem cell lines derived from equine fibroblasts. Stem Cell Rev. Rep. 2011, 7, 693–702. [Google Scholar] [CrossRef]
- Khodadadi, K.; Sumer, H.; Pashaiasl, M.; Lim, S.; Williamson, M.; Verma, P.J. Induction of pluripotency in adult equine fibroblasts without c-MYC. Stem Cells Int. 2012, 2012, 29160. [Google Scholar] [CrossRef]
- Breton, A.; Sharma, R.; Diaz, A.C.; Parham, A.G.; Graham, A.; Neil, C.; Whitelaw, C.B.; Milne, E.; Donadeu, F.X. Derivation and characterization of induced pluripotentstem cells from equine fibroblasts. Stem Cells Dev. 2013, 22, 611–621. [Google Scholar] [CrossRef]
- Whitworth, D.J.; Ovchinnikov, D.A.; Sun, J.; Fortuna, P.R.; Wolvetang, E.J. Generation and characterization of leukemia inhibitory factor-dependent equineinduced pluripotent stem cells from adult dermal fibroblasts. Stem Cells Dev. 2014, 23, 1515–1523. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.M.; Kim, A.Y.; Lee, E.J.; Park, J.K.; Park, S.I.; Cho, S.G.; Kim, H.K.; Kim, S.Y.; Jeong, K.S. Generation of Equine-Induced Pluripotent Stem Cellsand Analysis of Their Therapeutic Potential for Muscle Injuries. Cell Transpl. 2016, 25, 2003–2016. [Google Scholar] [CrossRef]
- Quattrocelli, M.; Giacomazzi, G.; Broeckx, S.Y.; Ceelen, L.; Bolca, S.; Spaas, J.H.; Sampaolesi, M. Equine-Induced Pluripotent Stem Cells Retain Lineage Commitment Tward Myogenic and Chondrogenic Fates. Stem Cell Rep. 2016, 6, 55–63. [Google Scholar] [CrossRef]
- Moro, L.N.; Amin, G.; Furmento, V.; Waisman, A.; Garate, X.; Neiman, G.; La Greca, A.; Santín-Velazque, N.L.; Luzzani, C.; Sevlever, G.E.; et al. MicroRNA characterization in equine induced pluripotent stem cells. PLoS ONE 2018, 13, e0207074. [Google Scholar] [CrossRef] [PubMed]
- Pessôa, L.V.F.; Pires, P.R.L.; Del Collado, M.; Pieri, N.C.G.; Recchia, K.; Souza, A.F.; Perecin, F.; da Silveira, J.C.; de Andrade, A.F.C.; Ambrosio, C.E.; et al. Generation and miRNA Characterization of Equine Induced Pluripotent Stem Cells Derived from Fetal and Adult Multipotent Tissues. Stem Cells Int. 2019, 2019, 1393791. [Google Scholar] [CrossRef]
- Han, X.; Han, J.; Ding, F.; Cao, S.; Lim, S.S.; Dai, Y.; Zhang, R.; Zhang, Y.; Lim, B.; Li, N. Generation of induced pluripotent stem cells from bovine embryonic fibroblast cells. Cell Res. 2011, 21, 1509–1512. [Google Scholar] [CrossRef]
- Sumer, H.; Liu, J.; Malaver-Ortega, L.F.; Lim, M.L.; Khodadadi, K.; Verma, P.J. NANOG is a key factor for induction of pluripotency in bovine adult fibroblasts. J. Anim. Sci. 2011, 89, 708–716. [Google Scholar] [CrossRef]
- Cao, H.; Yang, P.; Pu, Y.; Sun, X.; Yin, H.; Zhang, Y.; Zhang, Y.; Li, Y.; Liu, Y.; Fang, F.; et al. Characterization of bovine induced plu-ripotent stem cells by lentiviral transduction of reprogramming factor fusion proteins. Int. J. Biol. Sci. 2012, 8, 498–511. [Google Scholar] [CrossRef]
- Talluri, T.R.; Kumar, D.; Glage, S.; Garrels, W.; Ivics, Z.; Debowski, K.; Behr, R.; Niemann, H.; Kues, W.A. Derivation and characterization of bovine induced plu-ripotent stem cells by transposon-mediated reprogramming. Cell Reprogram. 2015, 17, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Malaver-Ortega, L.F.; Sumer, H.; Liu, J.; Verma, P.J. Inhibition of JAK-STAT ERK/MAPK and Glycogen Synthase Kinase-3 Induces a Change in Gene Expression Profile of Bovine Induced Pluripotent Stem Cells. Stem Cells Int. 2016, 2016, 5127984. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Z.; Zhang, J.; Yang, J.; Gao, X.; Wu, B.; Zhao, G.; Bao, S.; Hu, S.; Liu, P.; et al. Characterization of the single-cell derived bovine induced pluripotent stem cells. Tissue Cell 2017, 49, 521–527. [Google Scholar] [CrossRef]
- Pillai, V.V.; Kei, T.G.; Reddy, S.E.; Das, M.; Abratte, C.; Cheong, S.H.; Selvaraj, V. Induced pluripotent stem cell generation from bovine somatic cells indicates unmet needs for pluripotency sustenance. Anim. Sci. J. 2019, 90, 1149–1160. [Google Scholar] [CrossRef]
- Shimada, H.; Nakada, A.; Hashimoto, Y.; Shigeno, K.; Shionoya, Y.; Nakamura, T. Generation of canine induced pluripotent stem cells by retroviral transduc-tion and chemical inhibitors. Mol. Reprod. Dev. 2010, 77, 2. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Suhr, S.T.; Chang, E.A.; Wang, K.; Ross, P.J.; Nelson, L.L.; Venta, P.J.; Knott, J.G.; Cibelli, J.B. Generation of leukemia inhibitory factor and basic fi-broblast growth factor-dependent induced pluripotent stem cells from canine adult somatic cells. Stem Cells Dev. 2011, 20, 1669–1678. [Google Scholar] [CrossRef]
- Lee, A.S.; Xu, D.; Plews, J.R.; Nguyen, P.K.; Nag, D.; Lyons, J.K.; Han, L.; Hu, S.; Lan, F.; Liu, J.; et al. Preclinical derivation and imaging of autologously transplanted canine induced pluripotent stem cells. J. Biol. Chem. 2011, 286, 32697–32704. [Google Scholar] [CrossRef] [PubMed]
- Koh, S.; Thomas, R.; Tsai, S.; Bischoff, S.; Lim, J.H.; Breen, M.; Olby, N.J.; Piedrahita, J.A. Growth requirements and chromosomal instability of induced pluripotent stem cells generated from adult canine fibroblasts. Stem Cells Dev. 2013, 22, 951–963. [Google Scholar] [CrossRef]
- Whitworth, D.J.; Ovchinnikov, D.A.; Wolvetang, E.J. Generation and characterization of LIF-dependent canine induced pluripotent stem cells from adult dermal fibroblasts. Stem Cells Dev. 2012, 21, 2288–2297. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, T.; Hatoya, S.; Kanegi, R.; Sugiura, K.; Wijewardana, V.; Kuwamura, M.; Tanaka, M.; Yamate, J.; Izawa, T.; Takahashi, M.; et al. Generation of functional platelets from canine induced pluripotent stem cells. Stem Cells Dev. 2013, 22, 2026–2035. [Google Scholar] [CrossRef]
- Bao, L.; He, L.; Chen, J.; Wu, Z.; Liao, J.; Rao, L.; Ren, J.; Li, H.; Zhu, H.; Qian, L.; et al. Reprogramming of ovine adult fibroblasts to pluripotency via drug-inducible expression of defined factors. Cell Res. 2011, 21, 600–608. [Google Scholar] [CrossRef]
- Li, Y.; Cang, M.; Lee, A.S.; Zhang, K.; Liu, D. Reprogramming of sheep fibroblasts into pluripotency under a drug-inducible expression of mouse-derived defined factors. PLoS ONE 2011, 6, e15947. [Google Scholar] [CrossRef]
- Liu, J.; Balehosur, D.; Murray, B.; Kelly, J.M.; Sumer, H.; Verma, P.J. Generation and characterization of reprogrammed sheep induced pluripo-tent stem cells. Theriogenology 2012, 77, 338–346.e1. [Google Scholar] [CrossRef]
- Shi, H.; Fu, Q.; Li, G.; Ren, Y.; Hu, S.; Ni, W.; Guo, F.; Shi, M.; Meng, L.; Zhang, H.; et al. Roles of p53 and ASF1A in the Repro-gramming of Sheep Kidney Cells to Pluripotent Cells. Cell Reprogram. 2015, 17, 441–452. [Google Scholar] [CrossRef] [PubMed]
- Tai, D.; Liu, P.; Gao, J.; Jin, M.; Xu, T.; Zuo, Y.; Liang, H.; Liu, D. Generation of Arbas Cashmere Goat Induced Pluripotent Stem Cells Through Fibroblast Reprogramming. Cell Reprogram. 2015, 17, 297–305. [Google Scholar] [CrossRef]
- Song, H.; Li, H.; Huang, M.; Xu, D.; Gu, C.; Wang, Z.; Dong, F.; Wang, F. Induced pluripotent stem cells from goat fibroblasts. Mol. Reprod Dev. 2013, 80, 1009–1017. [Google Scholar] [CrossRef]
- Sandmaier, S.E.; Nandal, A.; Powell, A.; Garrett, W.; Blomberg, L.; Donovan, D.M.; Talbot, N.; Telugu, B.P. Generation of induced pluripotent stem cells from domestic goats. Mol. Reprod. Dev. 2015, 82, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zuo, Q.; Wang, Y.; Song, J.; Yang, H.; Zhang, Y.; Li, B. Inducing goat pluripotent stem cells with four transcription factor mRNAs that activate endogenous promoters. BMC Biotechnol. 2017, 17, 11. [Google Scholar] [CrossRef]
- Chung, M.J.; Park, S.; Son, J.Y.; Lee, J.Y.; Yun, H.H.; Lee, E.J.; Lee, E.M.; Cho, G.J.; Lee, S.; Park, H.S.; et al. Differentiation of equine induced pluripotent stem cells into mesenchymal lineage for therapeutic use. Cell Cycle 2019, 18, 2954–2971. [Google Scholar] [CrossRef] [PubMed]
- Telugu, B.P.; Ezashi, T.; Roberts, R.M. Porcine induced pluripotent stem cells analogous to naïve and primed embryonic stem cells of the mouse. Int. J. Dev. Biol. 2010, 54, 1703–1711. [Google Scholar] [CrossRef]
- Wu, R.; Liu, Y.; Zhao, Y.; Bi, Z.; Yao, Y.; Liu, Q.; Wang, F.; Wang, Y.; Wang, X. m 6 A methylation controls pluripotency of porcine induced pluripotent stem cells by targeting SOCS3/JAK2/STAT3 pathway in a YTHDF1/YTHDF2-orchestrated manner. Cell Death Dis. 2019, 10, 171. [Google Scholar] [CrossRef]
- Wang, G.; Weng, R.; Lan, Y.; Guo, X.; Liu, Q.; Liu, X.; Lu, C.; Kang, J. Synergetic effects of DNA methylation and histone modification during mouse induced pluripotent stem cell generation. Sci. Rep. 2017, 7, 39527. [Google Scholar] [CrossRef] [PubMed]
- Habekost, M.; Jørgensen, A.L.; Qvist, P.; Denham, M. Transcriptomic profiling of porcine pluripotency identifies species-specific reprogramming requirements for culturing iPSCs. Stem Cell Res. 2019, 41, 101645. [Google Scholar] [CrossRef]
- Shi, B.; Gao, D.; Zhong, L.; Zhi, M.; Weng, X.; Xu, J.; Li, J.; Du, X.; Xin, Y.; Gao, J.; et al. IRF-1 expressed in the inner cell mass of the porcine early blastocyst enhances the pluripotency of induced pluripotent stem cells. Stem Cell Res. Ther. 2020, 11, 505. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.H.; Loo, S.J.; Gao, Y.; Tao, Z.H.; Su, L.P.; Wang, C.X.; Zhang, S.L.; Mu, Y.H.; Cui, Y.H.; Abdurrachim, D.; et al. Thymosin β4 increases cardiac cell proliferation, cell engraftment, and the reparative potency of human induced-pluripotent stem cell-derived cardiomyocytes in a porcine model of acute myocardial infarction. Theranostics 2021, 11, 7879–7895. [Google Scholar] [CrossRef] [PubMed]
- Biagi, D.; Fantozzi, E.T.; Campos-Oliveira, J.C.; Naghetini, M.V.; Ribeiro, A.F., Jr.; Rodrigues, S.; Ogusuku, I.; Vanderlinde, R.; Christie, M.L.A.; Mello, D.B.; et al. In Situ Maturated Early-Stage Human-Induced Pluripotent Stem Cell-Derived Cardiomyocytes Improve Cardiac Function by Enhancing Segmental Contraction in Infarcted Rats. J. Pers Med. 2021, 11, 374. [Google Scholar] [CrossRef]
- Kawaguchi, S.; Soma, Y.; Nakajima, K.; Kanazawa, H.; Tohyama, S.; Tabei, R.; Hirano, A.; Handa, N.; Yamada, Y.; Okuda, S.; et al. Intramyocardial Transplantation of Human iPS Cell-Derived Cardiac Spheroids Improves Cardiac Function in Heart Failure Animals. JACC Basic Transl. Sci. 2021, 6, 239–254. [Google Scholar] [CrossRef]
- Nishida, M.; Tanaka, Y.; Tanaka, Y.; Amaya, S.; Tanaka, N.; Uyama, H.; Masuda, T.; Onishi, A.; Sho, J.; Yokota, S.; et al. Human iPS cell derived RPE strips for secure delivery of graft cells at a target place with minimal surgical invasion. Sci. Rep. 2021, 11, 21421. [Google Scholar] [CrossRef]
- Duarri, A.; Rodríguez-Bocanegra, E.; Martínez-Navarrete, G.; Biarnés, M.; García, M.; Ferraro, L.L.; Kuebler, B.; Aran, B.; Izquierdo, E.; Aguilera-Xiol, E.; et al. Transplantation of Human Induced Pluripotent Stem Cell-Derived Retinal Pigment Epithelium in a Swine Model of Ge-ographic Atrophy. Int. J. Mol. Sci. 2021, 22, 10497. [Google Scholar] [CrossRef] [PubMed]
- Thavapalachandran, S.; Le, T.Y.L.; Romanazzo, S.; Rashid, F.N.; Ogawa, M.; Kilian, K.A.; Brown, P.; Pouliopoulos, J.; Barry, A.M.; Fahmy, P.; et al. Pluripotent stem cell-derived mesenchymal stromal cells improve cardiac function and vascularity after myocardial infarction. Cytotherapy 2021, 23, 1074–1084. [Google Scholar] [CrossRef] [PubMed]
- Lingam, S.; Liu, Z.; Yang, B.; Wong, W.; Parikh, B.H.; Ong, J.Y.; Goh, D.; Wong, D.S.L.; Tan, Q.S.W.; Tan, G.S.W.; et al. cGMP-grade human iPSC-derived retinal photoreceptor precursor cells rescue cone photoreceptor damage in non-human primates. Stem Cell Res. Ther. 2021, 12, 464. [Google Scholar] [CrossRef]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and Safety Issues of Stem Cell-Based Therapy. Int. J. Med. Sci. 2018, 15, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Tavakol, D.N.; Fleischer, S.; Vunjak-Novakovic, G. Harnessing organs-on-a-chip to model tissue regeneration. Cell Stem Cell 2021, 28, 993–1015. [Google Scholar] [CrossRef]
- Ronaldson-Bouchard, K.; Ma, S.P.; Yeager, K.; Chen, T.; Song, L.; Sirabella, D.; Morikawa, K.; Teles, D.; Yazawa, M.; Vunjak-Novakovic, G. Advanced maturation of human cardiac tissue grown from pluripotent stem cells. Nature 2018, 556, 239–243. [Google Scholar] [CrossRef]
- Zhao, Y.; Rafatian, N.; Feric, N.T.; Cox, B.J.; Aschar-Sobbi, R.; Wang, E.Y.; Aggarwal, P.; Zhang, B.; Conant, G.; Ronaldson-Bouchard, K.; et al. A Platform for Generation of Chamber-Specific Cardiac Tissues and Disease Modeling. Cell 2019, 176, 913–927. [Google Scholar] [CrossRef]
- Tang, B.L. Maturing iPSC-Derived Cardiomyocytes. Cells 2020, 9, 213. [Google Scholar] [CrossRef] [PubMed]
- Breckwoldt, K.; Letuffe-Brenière, D.; Mannhardt, I.; Schulze, T.; Ulmer, B.; Werner, T.; Benzin, A.; Klampe, B.; Reinsch, M.C.; Laufer, S.; et al. Differentiation of cardiomyocytes and generation of human engineered heart tissue. Nat. Protoc. 2017, 12, 1177–1197. [Google Scholar] [CrossRef] [PubMed]
- Shiba, Y.; Gomibuchi, T.; Seto, T.; Wada, Y.; Ichimura, H.; Tanaka, Y.; Ogasawara, T.; Okada, K.; Shiba, N.; Sakamoto, K.; et al. Allogeneic transplantation of iPS cell-derived cardiomyocytes regenerates primate hearts. Nature 2016, 538, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Bizy, A.; Klos, M. Optimizing the Use of iPSC-CMs for Cardiac Regeneration in Animal Models. Animals 2020, 10, 1561. [Google Scholar] [CrossRef]
- Zhang, J.; Wilson, G.F.; Soerens, A.G.; Koonce, C.H.; Yu, J.; Palecek, S.P.; Thomson, J.A.; Kamp, T.J. Functional cardiomyocytes derived from human induced pluripotent stem cells. Circ. Res. 2009, 104, e30–e41. [Google Scholar] [CrossRef]
- Cyganek, L.; Tiburcy, M.; Sekeres, K.; Gerstenberg, K.; Bohnenberger, H.; Lenz, C.; Henze, S.; Stauske, M.; Salinas, G.; Zimmermann, W.H.; et al. Deep phenotyping of human induced pluripotent stem cell-derived atrial and ventricular cardiomyocytes. JCI Insight 2018, 3, e99941. [Google Scholar] [CrossRef] [PubMed]
- Fujita, J. Development of Cardiac Regenerative Medicine Using Human iPS Cell-derived Cardiomyocytes. Keio J. Med. 2021, 70, 53–59. [Google Scholar] [CrossRef]
- Masumoto, H.; Yamashita, J.K. Human iPS Cell-Derived Cardiac Tissue Sheets: A Platform for Cardiac Regeneration. Curr. Treat Options Cardiovasc. Med. 2016, 18, 65. [Google Scholar] [CrossRef]
- van den Berg, C.W.; Elliott, D.A.; Braam, S.R.; Mummery, C.L.; Davis, R.P. Differentiation of Human Pluripotent Stem Cells to Cardiomyocytes under Defined Conditions. Methods Mol. Biol. 2016, 1353, 163–180. [Google Scholar] [PubMed]
- Paige, S.L.; Osugi, T.; Afanasiev, O.K.; Pabon, L.; Reinecke, H.; Murry, C.E. Endogenous Wnt/beta-catenin signaling is required for cardiac differentiation in human embryonic stem cells. PLoS ONE 2010, 5, e11134. [Google Scholar] [CrossRef]
- Laflamme, M.A.; Chen, K.Y.; Naumova, A.V.; Muskheli, V.; Fugate, J.A.; Dupras, S.K.; Reinecke, H.; Xu, C.; Hassanipour, M.; Police, S.; et al. Cardiomyocytes derived from human embryonic stem cells in pro-survivalfactors enhance function of infarcted rat hearts. Nat. Biotechnol. 2007, 25, 1015–1024. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Zweigerdt, R.; Soo, S.Y.; Ngoh, Z.X.; Tham, S.C.; Wang, S.T.; Graichen, R.; Davidson, B.; Colman, A.; Sun, W. Highly enriched cardiomyocytes from human embryonic stem cells. Cytotherapy 2008, 10, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Kempf, H.; Kropp, C.; Olmer, R.; Martin, U.; Zweigerdt, R. Cardiac differentiation of human pluripotent stem cells in scalable suspension culture. Nat. Protoc. 2015, 10, 1345–1361. [Google Scholar] [CrossRef] [PubMed]
- Laco, F.; Woo, T.L.; Zhong, Q.; Szmyd, R.; Ting, S.; Khan, F.J.; Chai, C.L.L.; Reuveny, S.; Chen, A.; Oh, S. Unraveling the Inconsistencies of Cardiac Differentiation Efficiency Induced by the GSK3β Inhibitor CHIR99021 in Human Pluripotent Stem Cells. Stem Cell Rep. 2018, 10, 1851–1866. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Hsiao, C.; Wilson, G.; Zhu, K.; Hazeltine, L.B.; Azarin, S.M.; Raval, K.K.; Zhang, J.; Kamp, T.J.; Palecek, S.P. Robust cardiomyocyte differentiation from human pluripotent stem cells via temporal modulation of canonical Wnt signaling. Proc. Natl. Acad. Sci. USA 2012, 109, E1848–E1857. [Google Scholar] [CrossRef] [PubMed]
- Lian, X.; Zhang, J.; Azarin, S.M.; Zhu, K.; Hazeltine, L.B.; Bao, X.; Hsiao, C.; Kamp, T.J.; Palecek, S.P. Directed cardiomyocyte differentiation from human pluripotent stem cells by modulating Wnt/β-catenin signaling under fully defined conditions. Nat. Prot. 2013, 8, 162–175. [Google Scholar] [CrossRef]
- Mummery, C.; Zhang, J.; Ng, E.S.; Elliott, D.; Elefanty, A.; Kamp, T. Differentiation of Human Embryonic Stem Cells and Induced Pluripotent Stem Cells to Cardiomyocytes. Circ. Res. 2012, 111, 344–358. [Google Scholar] [CrossRef]
- Burridge, P.W.; Thompson, S.; Millrod, M.A.; Weinberg, S.; Yuan, X.; Peters, A.; Mahairaki, V.; Koliatsos, V.E.; Tung, L.; Zambidis, E.T. A Universal System for Highly Efficient Cardiac Differentiation of Human Induced Pluripotent Stem Cells That Eliminates Interline Variability. PLoS ONE 2011, 6, e18293. [Google Scholar] [CrossRef]
- Freund, C.; Oostwaard, D.W.; Monshouwer-Kloots, J.; Brink, S.V.D.; van Rooijen, M.; Xu, X.; Zweigerdt, R.; Mummery, C.; Passier, R. Insulin Redirects Differentiation from Cardiogenic Mesoderm and Endoderm to Neuroectoderm in Differentiating Human Embryonic Stem Cells. Stem Cells 2008, 26, 724–733. [Google Scholar] [CrossRef]
- Huang, C.Y.; Maia-Joca, R.P.M.; Ong, C.S.; Wilson, I.; DiSilvestre, D.; Tomaselli, G.F.; Reich, D.H. Enhancement of human iPSC-derived cardiomyocyte maturation by chemical conditioning in a 3D environment. J. Mol. Cell. Cardiol. 2020, 138, 1–11. [Google Scholar] [CrossRef]
- Li, P.; Cavallero, S.; Gu, Y.; Chen, T.H.P.; Hughes, J.; Hassan, A.B.; Brüning, J.C.; Pashmforoush, M.; Sucov, H.M. IGF signaling directs ventricular cardiomyocyte proliferation during embryonic heart development. Development 2011, 138, 1795–1805. [Google Scholar] [CrossRef]
- Lian, X.; Zhang, J.; Zhu, K.; Kamp, T.J.; Palecek, S.P. Insulin Inhibits Cardiac Mesoderm, Not Mesendoderm, Formation During Cardiac Differentiation of Human Pluripotent Stem Cells and Modulation of Canonical Wnt Signaling Can Rescue This Inhibition. Stem Cells 2012, 31, 447–457. [Google Scholar] [CrossRef] [PubMed]
- McDevitt, T.C.; Laflamme, M.; Murry, C.E. Proliferation of cardiomyocytes derived from human embryonic stem cells is mediated via the IGF/PI 3-kinase/Akt signaling pathway. J. Mol. Cell. Cardiol. 2005, 39, 865–873. [Google Scholar] [CrossRef] [PubMed]
- Kaichi, S.; Hasegawa, K.; Takaya, T.; Yokoo, N.; Mima, T.; Kawamura, T.; Morimoto, T.; Ono, K.; Baba, S.; Doi, H.; et al. Cell line-dependent differentiation of induced pluripotent stem cells into cardiomyocytes in mice. Cardiovasc. Res. 2010, 88, 314–323. [Google Scholar] [CrossRef][Green Version]
- Cao, N.; Liu, Z.; Chen, Z.; Wang, J.; Chen, T.; Zhao, X.; Ma, Y.; Qin, L.; Kang, J.; Wei, B.; et al. Ascorbic acid enhances the cardiac differentiation of induced pluripotent stem cells through promoting the proliferation of cardiac progenitor cells. Cell Res. 2012, 22, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef]
- Xavier-Neto, J.; Neville, C.M.; Shapiro, M.D.; Houghton, L.; Wang, G.F.; Nikovits, W., Jr.; Stockdale, F.E.; Rosenthal, N. A retinoic acid-inducible transgenic marker of sino-atrial development in the mouse heart. Development 1999, 126, 2677–2687. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.Z.; Xie, Y.; Moyes, K.W.; Gold, J.D.; Askari, B.; Laflamme, M.A. Neuregulin/ErbB signaling regulates cardiac subtype specification in differentiating human embryonic stem cells. Circ. Res. 2010, 107, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Gassanov, N.; Er, F.; Zagidullin, N.; Hoppe, U.C. Endothelin induces differentiation of ANP-EGFP expressing embryonic stem cells towards a pacemaker phenotype. FASEB J. 2004, 18, 1710–1712. [Google Scholar] [CrossRef]
- Yoshida, Y.; Yamanaka, S. Induced Pluripotent Stem Cells 10 Years Later: For Cardiac Applications. Circ. Res. 2017, 120, 1958–1968. [Google Scholar] [CrossRef]
- Yang, X.; Pabon, L.; Murry, C.E. Engineering adolescence: Maturation of human pluripotent stem cell-derived cardiomyocytes. Circ. Res. 2014, 114, 511–523. [Google Scholar] [CrossRef]
- Zhang, Z.; Bédard, E.; Luo, Y.; Wang, H.; Deng, S.; Kelvin, D.; Zhong, R. Animal models in xenotransplantation. Expert Opin. Investig. Drugs 2000, 9, 2051–2068. [Google Scholar] [CrossRef]
- Milani-Nejad, N.; Janssen, P.M. Small and large animal models in cardiac contraction research: Advantages and disadvantages. Pharmacol. Ther. 2014, 141, 235–249. [Google Scholar] [CrossRef]
- Fliegner, D.; Gerdes, C.; Meding, J.; Stasch, J.P. Translational In Vivo Models for Cardiovascular Diseases. Handb. Exp. Pharmacol. 2016, 232, 223–234. [Google Scholar]
- Recchia, F.A.; Lionetti, V. Animal models of dilated cardiomyopathy for translational research. Vet. Res. Commun. 2007, 31, 35–41. [Google Scholar] [CrossRef]
- Bolli, R.; Ghafghazi, S. Cell Therapy Needs Rigorous Translational Studies in Large Animal Models. J. Am. Coll. Cardiol. 2015, 66, 2000–2004. [Google Scholar] [CrossRef]
- Harding, J.; Roberts, R.M.; Mirochnitchenko, O. Large animal models for stem cell therapy. Stem Cell Res. Ther. 2013, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- van der Spoel, T.I.; Jansen of Lorkeers, S.J.; Agostoni, P.; van Belle, E.; Gyöngyösi, M.; Sluijter, J.P.; Cramer, M.J.; Doevendans, P.A.; Chamuleau, S.A. Human relevance of pre-clinical studies in stem cell therapy: Systematic review and meta-analysis of large animal models of ischaemic heart disease. Cardiovasc Res. 2011, 91, 649–658. [Google Scholar] [CrossRef]
- Wang, H. Small animal models of xenotransplantation. Methods Mol. Biol. 2012, 885, 125–153. [Google Scholar] [PubMed]
- da Silva Morais, A.; Oliveira, J.M.; Reis, R.L. Small Animal Models. Adv. Exp. Med. Biol. 2018, 1059, 423–439. [Google Scholar] [PubMed]
- Russell, J.C.; Proctor, S.D. Small animal models of cardiovascular disease: Tools for the study of the roles of metabolic syn-drome, dyslipidemia, and atherosclerosis. Cardiovasc. Pathol. 2006, 15, 318–330. [Google Scholar] [CrossRef]
- Bracken, M.B. Why animal studies are often poor predictors of human reactions to exposure. J. R. Soc. Med. 2009, 102, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Martić-Kehl, M.I.; Schibli, R.; Schubiger, P.A. Can animal data predict human outcome? Problems and pitfalls of translational animal research. Eur. J. Nucl. Med. Mol. Imaging 2012, 39, 1492–1496. [Google Scholar] [CrossRef]
- Veening-Griffioen, D.H.; Ferreira, G.S.; van Meer, P.J.K.; Boon, W.P.C.; Gispen-de Wied, C.C.; Moors, E.H.M.; Schellekens, H. Are some animal models more equal than others? A case study on the translational value of animal models of efficacy for Alzheimer’s disease. Eur. J. Pharmacol. 2019, 859, 172524. [Google Scholar] [CrossRef]
- Li, X.J.; Li, S. Large Animal Models of Huntington’s Disease. Curr. Top. Behav. Neurosci. 2015, 22, 149–160. [Google Scholar]
- Shim, J.; Al-Mashhadi, R.H.; Sørensen, C.B.; Bentzon, J.F. Large animal models of atherosclerosis—New tools for persistent problems in car-diovascular medicine. J. Pathol. 2016, 238, 257–266. [Google Scholar] [CrossRef]
- Hotham, W.E.; Henson, F.M.D. The use of large animals to facilitate the process of MSC going from laboratory to pa-tient-‘bench to bedside’. Cell Biol. Toxicol. 2020, 36, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Van der Velden, J.; Snibson, K.J. Airway disease: The use of large animal models for drug discovery. Pulm. Pharmacol. Ther. 2011, 24, 525–532. [Google Scholar] [CrossRef]
- Plews, J.R.; Gu, M.; Longaker, M.T.; Wu, J.C. Large animal induced pluripotent stem cells as pre-clinical models for studying human disease. J. Cell. Mol. Med. 2012, 16, 1196–1202. [Google Scholar] [CrossRef] [PubMed]
- Alstrup, K.O.; Winterdahl, M. Imaging techniques in large animals. Scand. J. Lab. Anim. Sci. 2009, 36, 55–66. [Google Scholar] [CrossRef]
- Yang, P.C. Is Reliable In Vivo Detection of Stem Cell Viability Possible in a Large Animal Model of Myocardial Injury? Circulation 2012, 126, 388–390. [Google Scholar] [CrossRef][Green Version]
- Gandolfi, F.; Vanelli, A.; Pennarossa, G.; Rahaman, M.; Acocella, F.; Brevini, T. Large animal models for cardiac stem cell therapies. Theriogenology 2011, 75, 1416–1425. [Google Scholar] [CrossRef] [PubMed]
- Amado, L.C.; Schuleri, K.H.; Saliaris, A.P.; Boyle, A.J.; Helm, R.; Oskouei, B.; Centola, M.; Eneboe, V.; Young, R.; Lima, J.A.; et al. Multimodality Noninvasive Imaging Demonstrates In Vivo Cardiac Regeneration After Mesenchymal Stem Cell Therapy. J. Am. Coll. Cardiol. 2006, 48, 2116–2124. [Google Scholar] [CrossRef]
- Mazhari, R.; Hare, J.M. Translational Findings from Cardiovascular Stem Cell Research. Trends Cardiovasc. Med. 2012, 22, 1–6. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, X.; Zhang, F.; Song, G.; Gu, W.; Chen, M.; Yang, B.; Li, D.; Wang, D.; Cao, K. Intramyocardial Injection of Pig Pluripotent Stem Cells Improves Left Ventricular Function and Perfusion: A Study in a Porcine Model of Acute Myocardial Infarction. PLoS ONE 2013, 8, e66688. [Google Scholar] [CrossRef]
- Zhang, F.; Song, G.; Li, X.; Gu, W.; Shen, Y.; Chen, M.; Yang, B.; Qian, L.; Cao, K. Transplantation of iPSc Ameliorates Neural Remodeling and Reduces Ventricular Arrhythmias in a Post-Infarcted Swine Model. J. Cell. Biochem. 2014, 115, 531–539. [Google Scholar] [CrossRef]
- Song, G.; Lingmei, Q.; Shen, Y.; Qian, L.; Kong, X.; Chen, M.; Cao, K.; Zhang, F. Transplantation of iPSc Restores Cardiac Function by Promoting Angiogenesis and Ameliorating Cardiac Remodeling in a Post-infarcted Swine Model. Cell Biophys. 2014, 71, 1463–1473. [Google Scholar] [CrossRef] [PubMed]
- Gálvez-Montón, C.; Soler-Botija, C.; Iborra-Egea, O.; Díaz-Güemes, I.; Martí, M.; Iglesias-García, O.; Prat-Vidal, C.; Crisóstomo, V.; Llucià-Valldeperas, A.; Perea-Gil, I.; et al. Preclinical Safety Evaluation of Allogeneic Induced Pluripotent Stem Cell-Based Therapy in a Swine Model of Myocardial Infarction. Tissue Eng. Part C Methods 2017, 23, 736–744. [Google Scholar] [CrossRef]
- Kawamura, T.; Miyagawa, S.; Fukushima, S.; Maeda, A.; Kashiyama, N.; Kawamura, A.; Miki, K.; Okita, K.; Yoshida, Y.; Shiina, T.; et al. Cardiomyocytes Derived from MHC-Homozygous Induced Pluripotent Stem Cells Exhibit Reduced Allogeneic Immunogenicity in MHC-Matched Non-human Primates. Stem Cell Rep. 2016, 6, 312–320. [Google Scholar] [CrossRef]
- Kashiyama, N.; Miyagawa, S.; Fukushima, S.; Kawamura, T.; Kawamura, A.; Yoshida, S.; Eiraku, S.; Harada, A.; Matsunaga, K.; Watabe, T.; et al. MHC-mismatched Allotransplantation of Induced Pluripotent Stem Cell-derived Cardiomyocyte Sheets to Improve Cardiac Function in a Primate Ischemic Cardiomyopathy Model. Transplantation 2019, 103, 1582–1590. [Google Scholar] [CrossRef]
- Ye, L.; Chang, Y.-H.; Xiong, Q.; Zhang, P.; Zhang, L.; Somasundaram, P.; Lepley, M.; Swingen, C.; Su, L.; Wendel, J.S.; et al. Cardiac Repair in a Porcine Model of Acute Myocardial Infarction with Human Induced Pluripotent Stem Cell-Derived Cardiovascular Cells. Cell Stem Cell 2014, 15, 750–761. [Google Scholar] [CrossRef]
- Ishigami, M.; Masumoto, H.; Ikuno, T.; Aoki, T.; Kawatou, M.; Minakata, K.; Ikeda, T.; Sakata, R.; Yamashita, J.K.; Minatoya, K. Human iPS cell-derived cardiac tissue sheets for functional restoration of infarcted porcine hearts. PLoS ONE 2018, 13, e0201650. [Google Scholar] [CrossRef]
- Gao, L.; Gregorich, Z.R.; Zhu, W.; Mattapally, S.; Oduk, Y.; Lou, X.; Kannappan, R.; Borovjagin, A.V.; Walcott, G.P.; Pollard, A.E.; et al. Large Cardiac Muscle Patches Engineered From Human Induced-Pluripotent Stem Cell–Derived Cardiac Cells Improve Recovery From Myocardial Infarction in Swine. Circulation 2018, 137, 1712–1730. [Google Scholar] [CrossRef]
- Liao, S.; Zhang, Y.; Ting, S.; Zhen, Z.; Luo, F.; Zhu, Z.; Jiang, Y.; Sun, S.; Lai, W.-H.; Lian, Q.; et al. Potent immunomodulation and angiogenic effects of mesenchymal stem cells versus cardiomyocytes derived from pluripotent stem cells for treatment of heart failure. Stem Cell Res. Ther. 2019, 10, 78. [Google Scholar] [CrossRef]
- Kawamura, M.; Miyagawa, S.; Miki, K.; Saito, A.; Fukushima, S.; Higuchi, T.; Kuratani, T.; Daimon, T.; Shimizu, T.; Okano, T.; et al. Feasibility, Safety, and Therapeutic Efficacy of Human Induced Pluripotent Stem Cell-Derived Cardiomyocyte Sheets in a Porcine Ischemic Cardiomyopathy Model. Circulation 2012, 126, S29–S37. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, M.; Miyagawa, S.; Fukushima, S.; Saito, A.; Miki, K.; Ito, E.; Sougawa, N.; Kawamura, T.; Daimon, T.; Shimizu, T.; et al. Enhanced Survival of Transplanted Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes by the Combination of Cell Sheets With the Pedicled Omental Flap Technique in a Porcine Heart. Circulation 2013, 128, S87–S94. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, M.; Miyagawa, S.; Fukushima, S.; Saito, A.; Miki, K.; Funakoshi, S.; Yoshida, Y.; Yamanaka, S.; Shimizu, T.; Okano, T.; et al. Enhanced Therapeutic Effects of Human iPS Cell Derived-Cardiomyocyte by Combined Cell-Sheets with Omental Flap Technique in Porcine Ischemic Cardiomyopathy Model. Sci. Rep. 2017, 7, 8824. [Google Scholar] [CrossRef]
- Zhao, M.; Nakada, Y.; Wei, Y.; Bian, W.; Chu, Y.; Borovjagin, A.V.; Xie, M.; Zhu, W.; Nguyen, T.; Zhou, Y.; et al. Cyclin D2 Overexpression Enhances the Efficacy of Human Induced Pluripotent Stem Cell–Derived Cardiomyocytes for Myocardial Repair in a Swine Model of Myocardial Infarction. Circulation 2021, 144, 210–228. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Fujita, J.; Matsui, M.; Tohyama, S.; Tamura, N.; Kanazawa, H.; Seki, T.; Kishino, Y.; Hirano, A.; Okada, M.; et al. Gelatin Hydrogel Enhances the Engraftment of Transplanted Cardiomyocytes and Angiogenesis to Ameliorate Cardiac Function after Myocardial Infarction. PLoS ONE 2015, 10, e0133308. [Google Scholar] [CrossRef]
- Montserrat, N.; Bahima, E.G.; Batlle, L.; Häfner, S.; Rodrigues, A.; González, F.; Belmonte, J.C.I. Generation of Pig iPS Cells: A Model for Cell Therapy. J. Cardiovasc. Transl. Res. 2011, 4, 121–130. [Google Scholar] [CrossRef]
- Nelson, T.J.; Martinez-Fernandez, A.; Terzic, A. Induced pluripotent stem cells: Developmental biology to regenerative medicine. Nat. Rev. Cardiol. 2010, 7, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Gálvez-Montón, C.; Prat-Vidal, C.; Roura, S.; Farré, J.; Soler-Botija, C.; Llucià-Valldeperas, A.; Díaz-Güemes, I.; Sánchez-Margallo, F.M.; Arís, A.; Bayes-Genis, A. Transposition of a pericardial-derived vascular adipose flap for myocardial salvage after infarct. Cardiovasc. Res. 2011, 91, 659–667. [Google Scholar] [CrossRef][Green Version]
- Petersdorf, E.W. The major histocompatibility complex: A model for understanding graft-versus-host disease. Blood 2013, 122, 1863–1872. [Google Scholar] [CrossRef] [PubMed]
- Bach, F.H.; Bach, M.L.; Sondel, P.M. Differential function of major histocompatibility complex antigens in T-lymphocyte activation. Nat. Cell Biol. 1976, 259, 273–281. [Google Scholar] [CrossRef]
- Janeway, C.A.; Travers, P.; Walport, M.; Shlomchik, M. Immunobiology: The Immune System in Health and Disease; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Flomenberg, N.; Baxter-Lowe, L.A.; Confer, D.; Fernandez-Vina, M.; Filipovich, A.; Horowitz, M.; Hurley, C.; Kollman, C.; Anasetti, C.; Noreen, H.; et al. Impact of HLA class I and class II high-resolution matching on outcomes of unrelated donor bone marrow transplantation: HLA-C mismatching is associated with a strong adverse effect on transplantation outcome. Blood 2004, 104, 1923–1930. [Google Scholar] [CrossRef]
- Nakatsuji, N.; Nakajima, F.; Tokunaga, K. HLA-haplotype banking and iPS cells. Nat. Biotechnol. 2008, 26, 739–740. [Google Scholar] [CrossRef]
- Taylor, C.J.; Peacock, S.; Chaudhry, A.N.; Bradley, J.A.; Bolton, E. Generating an iPSC Bank for HLA-Matched Tissue Transplantation Based on Known Donor and Recipient HLA Types. Cell Stem Cell 2012, 11, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Germanguz, I.; Sedan, O.; Zeevi-Levin, N.; Shtrichman, R.; Barak, E.; Ziskind, A.; Eliyahu, S.; Meiry, G.; Amit, M.; Itskovitz-Eldor, J.; et al. Molecular characterization and functional properties of cardiomyocytes derived from human inducible pluripotent stem cells. J. Cell. Mol. Med. 2009, 15, 38–51. [Google Scholar] [CrossRef]
- Ren, Y.; Lee, M.Y.; Schliffke, S.; Paavola, J.; Amos, P.J.; Ge, X.; Ye, M.; Zhu, S.; Senyei, G.; Lum, L.; et al. Small molecule Wnt inhibitors enhance the efficiency of BMP-4-directed cardiac differentiation of human pluripotent stem cells. J. Mol. Cell. Cardiol. 2011, 51, 280–287. [Google Scholar] [CrossRef]
- Beauchamp, J.; Morgan, J.E.; Pagel, C.; Partridge, T.A. Dynamics of Myoblast Transplantation Reveal a Discrete Minority of Precursors with Stem Cell–like Properties as the Myogenic Source. J. Cell Biol. 1999, 144, 1113–1122. [Google Scholar] [CrossRef]
- Qu, Z.; Balkir, L.; Van Deutekom, J.C.; Robbins, P.D.; Pruchnic, R.; Huard, J. Development of Approaches to Improve Cell Survival in Myoblast Transfer Therapy. J. Cell Biol. 1998, 142, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.-L.; Rokosh, G.; Sanganalmath, S.K.; Yuan, F.; Sato, H.; Mu, J.; Dai, S.; Li, C.; Chen, N.; Peng, Y.; et al. Intracoronary Administration of Cardiac Progenitor Cells Alleviates Left Ventricular Dysfunction in Rats With a 30-Day-Old Infarction. Circulation 2010, 121, 293–305. [Google Scholar] [CrossRef]
- Menasche, P. Cardiac cell therapy: Lessons from clinical trials. J. Mol. Cell. Cardiol. 2011, 50, 258–265. [Google Scholar] [CrossRef]
- Hsiao, L.-C.; Carr, C.; Chang, K.-C.; Lin, S.-Z.; Clarke, K. Stem Cell-Based Therapy for Ischemic Heart Disease. Cell Transplant. 2013, 22, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Templin, C.; Zweigerdt, R.; Schwanke, K.; Olmer, R.; Ghadri, J.-R.; Emmert, M.Y.; Müller, E.; Küest, S.M.; Cohrs, S.; Schibli, R.; et al. Transplantation and Tracking of Human-Induced Pluripotent Stem Cells in a Pig Model of Myocardial Infarction. Circulation 2012, 126, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Nunes, S.S. Bioengineering Approaches to Mature Human Pluripotent Stem Cell-Derived Cardiomyocytes. Front. Cell Dev. Biol. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Shadrin, I.; Lam, J.; Xian, H.-Q.; Snodgrass, H.R.; Bursac, N. Tissue-engineered cardiac patch for advanced functional maturation of human ESC-derived cardiomyocytes. Biomaterials 2013, 34, 5813–5820. [Google Scholar] [CrossRef]
- Nguyen, P.K.; Rhee, J.-W.; Wu, J.C. Adult stem cell therapy and heart failure, 2000 to 2016: A systematic review. JAMA Cardiol. 2016, 1, 831–841. [Google Scholar] [CrossRef]
- Poulin, M.-F.; Deka, A.; Mohamedali, B.; Schaer, G.L. Clinical Benefits of Stem Cells for Chronic Symptomatic Systolic Heart Failure: A Systematic Review of the Existing Data and Ongoing Trials. Cell Transplant. 2016, 25, 1911–1923. [Google Scholar] [CrossRef]
- Ishida, M.; Miyagawa, S.; Saito, A.; Fukushima, S.; Harada, A.; Ito, E.; Ohashi, F.; Watabe, T.; Hatazawa, J.; Matsuura, K.; et al. Transplantation of Human-induced Pluripotent Stem Cell-derived Cardiomyocytes Is Superior to Somatic Stem Cell Therapy for Restoring Cardiac Function and Oxygen Consumption in a Porcine Model of Myocardial Infarction. Transplantation 2019, 103, 291–298. [Google Scholar] [CrossRef]
- Chandra, A.; Srivastava, R.K.; Kashyap, M.P.; Kumar, R.; Srivastava, R.N.; Pant, A.B. The Anti-Inflammatory and Antibacterial Basis of Human Omental Defense: Selective Expression of Cytokines and Antimicrobial Peptides. PLoS ONE 2011, 6, e20446. [Google Scholar] [CrossRef]
- Beeres, S.L.M.A.; Atsma, D.E.; Van Ramshorst, J.; Schalij, M.J.; Bax, J.J. Cell therapy for ischaemic heart disease. Heart 2008, 94, 1214–1226. [Google Scholar] [CrossRef]
- Hou, D.; Youssef, E.A.-S.; Brinton, T.J.; Zhang, P.; Rogers, P.; Price, E.T.; Yeung, A.C.; Johnstone, B.H.; Yock, P.G.; March, K.L. Radiolabeled Cell Distribution After Intramyocardial, Intracoronary, and Interstitial Retrograde Coronary Venous Delivery. Circulation 2005, 112, I150–I156. [Google Scholar] [CrossRef]
- Hattori, F.; Chen, H.; Yamashita, H.; Tohyama, S.; Satoh, Y.-S.; Yuasa, S.; Li, W.; Yamakawa, H.; Tanaka, T.; Onitsuka, T.; et al. Nongenetic method for purifying stem cell–derived cardiomyocytes. Nat. Methods 2009, 7, 61–66. [Google Scholar] [CrossRef]
- Rane, A.A.; Christman, K.L. Biomaterials for the Treatment of Myocardial Infarction: A 5-Year Update. J. Am. Coll. Cardiol. 2011, 58, 2615–2629. [Google Scholar] [CrossRef]
- Tabei, R.; Kawaguchi, S.; Kanazawa, H.; Tohyama, S.; Hirano, A.; Handa, N.; Hishikawa, S.; Teratani, T.; Kunita, S.; Fukuda, J.; et al. Development of a transplant injection device for optimal distribution and retention of human induced pluripotent stem cell—derived cardiomyocytes. J. Heart Lung Transplant. 2019, 38, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Lan, C.-C.E.; Wu, C.-S.; Chiou, M.-H.; Hsieh, P.-C.; Yu, H.-S. Low-Energy Helium-Neon Laser Induces Locomotion of the Immature Melanoblasts and Promotes Melanogenesis of the More Differentiated Melanoblasts: Recapitulation of Vitiligo Repigmentation In Vitro. J. Investig. Dermatol. 2006, 126, 2119–2126. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Ye, L.; Zhang, P.; Lepley, M.; Swingen, C.; Zhang, L.; Kaufman, D.; Zhang, J. Bioenergetic and Functional Consequences of Cellular Therapy: Activation of Endogenous Cardiovascular Progenitor Cells. Circ. Res. 2012, 111, 455–468. [Google Scholar] [CrossRef]
- Hsieh, P.C.; Davis, M.E.; Lisowski, L.K.; Lee, R.T. Endothelial-cardiomyocyte interactions in cardiac development and repair. Annu. Rev. Physiol. 2006, 68, 51–66. [Google Scholar] [CrossRef]
- Brutsaert, D.L. Cardiac Endothelial-Myocardial Signaling: Its Role in Cardiac Growth, Contractile Performance, and Rhythmicity. Physiol. Rev. 2003, 83, 59–115. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.E.; Hsieh, P.C.; Takahashi, T.; Song, Q.; Zhang, S.; Kamm, R.D.; Grodzinsky, A.J.; Anversa, P.; Lee, R.T. Local myocardial insulin-like growth factor 1 (IGF-1) delivery with biotinylated peptide nanofibers improves cell therapy for myocardial infarction. Proc. Natl. Acad. Sci. USA 2006, 103, 8155–8160. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Hu, Q.; Wang, X.; Mansoor, A.; Lee, J.; Feygin, J.; Zhang, G.; Suntharalingam, P.; Boozer, S.; Mhashilkar, A.; et al. Bioenergetic and Functional Consequences of Bone Marrow–Derived Multipotent Progenitor Cell Transplantation in Hearts With Postinfarction Left Ventricular Remodeling. Circulation 2007, 115, 1866–1875. [Google Scholar] [CrossRef]
- Masumoto, H.; Matsuo, T.; Yamamizu, K.; Uosaki, H.; Narazaki, G.; Katayama, S.; Marui, A.; Shimizu, T.; Ikeda, T.; Okano, T.; et al. Pluripotent Stem Cell-Engineered Cell Sheets Reassembled with Defined Cardiovascular Populations Ameliorate Reduction in Infarct Heart Function Through Cardiomyocyte-Mediated Neovascularization. Stem Cells 2012, 30, 1196–1205. [Google Scholar] [CrossRef]
- Olivetti, G.; Capasso, J.M.; Meggs, L.G.; Sonnenblick, E.H.; Anversa, P. Cellular basis of chronic ventricular remodeling after myocardial infarction in rats. Circ. Res. 1991, 68, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Riegler, J.; Tiburcy, M.; Ebert, A.D.; Tzatzalos, E.; Raaz, U.; Abilez, O.J.; Shen, Q.; Kooreman, N.G.; Neofytou, E.; Chen, V.C.; et al. Human Engineered Heart Muscles Engraft and Survive Long Term in a Rodent Myocardial Infarction Model. Circ. Res. 2015, 117, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, F.; Breckwoldt, K.; Pecha, S.; Kelly, A.; Geertz, B.; Starbatty, J.; Yorgan, T.; Cheng, K.-H.; Lessmann, K.; Stolen, T.; et al. Cardiac repair in guinea pigs with human engineered heart tissue from induced pluripotent stem cells. Sci. Transl. Med. 2016, 8, 363ra148. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, M.; Liao, J.; Chang, C.; Liu, Y.; Padhiar, A.A.; Zhou, Y.; Zhou, G. Induced Pluripotent Stem Cell-Derived Mesenchymal Stem Cells Hold Lower Heterogeneity and Great Promise in Biological Research and Clinical Applications. Front. Cell Dev. Biol. 2021, 9, 716907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, Z.; Jiang, D.; Liang, X.; Liao, S.; Zhang, Z.; Yue, W.; Li, X.; Chiu, S.-M.; Chai, Y.-H.; et al. iPSC-MSCs with High Intrinsic MIRO1 and Sensitivity to TNF-α Yield Efficacious Mitochondrial Transfer to Rescue Anthracycline-Induced Cardiomyopathy. Stem Cell Rep. 2016, 7, 749–763. [Google Scholar] [CrossRef]
- Lian, Q.; Zhang, Y.; Zhang, J.; Zhang, H.K.; Wu, X.; Zhang, Y.; Lam, F.F.-Y.; Kang, S.; Xia, J.C.; Lai, W.-H.; et al. Functional Mesenchymal Stem Cells Derived From Human Induced Pluripotent Stem Cells Attenuate Limb Ischemia in Mice. Circulation 2010, 121, 1113–1123. [Google Scholar] [CrossRef]
- Ambrosio, C.; Zomer, H.; Vidane, A.; Gonçalves, N. Mesenchymal and induced pluripotent stem cells: General insights and clinical perspectives. Stem Cells Cloning Adv. Appl. 2015, 8, 125–134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Specie | Similarities | Differences |
|---|---|---|
| Canine | Closed circulatory system composed by heart, veins, and arteries Physiological function of the heart Heart anatomy (4 chambers) Hemodynamic parameters | Anatomic variation in the thoracic cavity Collateral coronary circulation Smaller heart size Resting heart rate (60–160 bpm) Number of pulmonary veins (4–8) Presence of the left azygous vein Positioning of vena cava Size and shape of the atrial appendage Higher heart weight/body weight ratio Tricuspid valve with 2 leaflets Physiological respiratory arrhythmia |
| Swine | Closed circulatory system composed by heart, veins, and arteries Physiological function of the heart Heart anatomy (4 chambers) Heart size Coronary anatomy Analogous coronary pattern Lack of collateral coronary circulation Left coronary supplies the majority of the myocardium Hemodynamic parameters Heart weight/body weight ratio Valvular anatomy | Anatomic variation in the thoracic cavity Number of pulmonary veins (2) Higher content of Purkinje fibers Higher presence of numerous nerves Presence of the left azygous vein Faster sinus rhythm Shorter PR interval |
| Ovine | Closed circulatory system composed by heart, veins, and arteries Physiological function of the heart Heart anatomy (4 chambers) Lack of collateral coronary circulation Heart rate | Anatomic variation in the thoracic cavity Lower heart weight/body weight ratio Shorter and immobile ascending aorta Structure and composition of the valves Intervalvular or membranous septum absent |
| Non-human primates | Closed circulatory system composed by heart, veins, and arteries Physiological function of the heart Heart anatomy (4 chambers) Lack of collateral coronary circulation Genomic organization of the MHC region Genetic and metabolic parameters Ratio of heart weight to body weight | Faster heart rate |
| Species; Gender | Sample Size | Model | Delivery | Cell Type | Immuno-Suppressive Therapy | Benefits | Adverse Events | Ref |
|---|---|---|---|---|---|---|---|---|
| Swine; NS | Sham n = 6 PBS n = 6 piPSC n = 6 | MI | IM | piPSC | No | Cardiac autonomic nerve regeneration; less ventricular arrhythmia; myocardial perfusion; cardiac function. | No | [166,167] |
| Swine; NS | Sham n = 6 PBS n = 6 piPSC n = 6 | MI | IM | piPSC | CsA + METH-PSL | Reduction of scar size; angiogenesis; less apoptosis and fibrosis. | No | [168] |
| Swine; NS | AGTP n = 6 Scaffold n = 13 Scaffold + AGTP n = 7 piPSC + AGTP n = 9 piPSC + Scaffold n = 11 piPSC + Scaffold + AGTP n = 11 | MI | Scaffold + adipose pedicle (AGTP) | piPSC | No | None | No | [169] |
| Non-Human Primate; NS | MHC matched iPSC-CMs + TAC + MMF + PSL n = 2 MHC mismatched iPSC-CMs + TAC + MMF + PSL n = 3 MHC matched iPSC-CMs +TAC n = 1 MHC matched iPSC-CMs + No drug n = 1 | Healthy | Sheet (back) vs. IM (heart) | iPSC-CMs | Tac + MMF + PSL | No host immune response in MHC-matched group + TAC + MMF + PSL. | No | [170] |
| Non-Human Primate; NS | PSC vehicle n = 5 MHC matched iPSC-CMs n = 5 | MI | IM | iPSC-CMs | Tac+ METH-PSL | Cardiac function; less fibrosis; higher vascular density. | Ventricular arrhythmias | [114] |
| Non- Human Primate; NS | Sham n = 4 MHC matched iPSC-CMs n = 4 MHC mismatched iPSC-CMs n = 4 | MI | Sheet | iPSC-CMs | Tac + MMF + PSL | Cardiac function; less fibrosis; higher vascular density. | No | [171] |
| Swine; NS | Sham n = 17 MI n = 17 Cells n = 17 Patch n = 17 Cells + Patch n = 17 | MI | Sheet vs. IM | hiPSC-CMs hiPSC-ECs hiPSC-SMCs | CsA | Cardiac function; cardiac metabolism; higher vascular density; apoptosis reduction. | No | [172] |
| Swine; NS | Sham n = 5 Cells + Patch n = 5 | MI | Sheet | hiPSC-CMs hiPSC-ECs hiPSC-SMCs | Yes; NS | Cardiac function; less fibrosis; higher vascular density. | No | [173] |
| Swine; NS | Sham n = 8 MI n = 15 Cells + Patch n = 13 Patch n= 14 | MI | Sheet | hiPSC-CMs hiPSC-ECs hiPSC-SMCs | CsA + Methy-PSL | Cardiac function; less fibrosis; infarct size; less apoptosis. | No | [174] |
| Swine; Female | MI n = 9 hESC-CMs n = 10 hiPSC-MSCs n = 9 | HF | IM | hiPSC-MSCs vs. hESC-CMs | CsA + steroid | Cardiac function; higher vascular density; less inflammation. | No | [175] |
| Swine; Female | Sham n = 6 Cell + Patch n = 6 | MI | Sheet | hiPSC-CMs | Tac | Cardiac function; higher vascular density; less fibrosis. | No | [176] |
| Swine; Female | Cell + Patch n = 8 Cell + Patch + Omentum flap n = 8 Sham n = 5 Cell + Patch + Omentum flap n = 7 Cell + Patch n = 6 Omentum flap n = 5 | MI | Sheet + omentum flap | hiPSC-MSCs hMSCs | Tac | Cardiac function; higher vascular density. | No | [177,178] |
| Swine; Female | MI n = 8; Tb4 n = 9; Tb4 Cell n = 8; Cell n = 8. | MI | Sheet + Tb4 microspheres | hiPSC-CMs | CsA | Cardiac function; higher vascular density; less fibrosis; reduction of scar size | No | [101] |
| Swine; Female | Sham n = 7 PBS n = 7 CCND2WTCMs n = 7 CCND2OECM n = 7 | MI | IM | hiPSC-CCND2WTCMs vs. hiPSC-CCND2oeCMs | No | Host CM proliferation; angiogenesis in border zone; cardiac function; less fibrosis; reduced hypertrophy. | No | [179] |
| Swine; female | Healthy pigs n = 15 Healthy mini-pigs n = 20 | Healthy | 3D spheroids injection device | hiPSC-CMs | NS | Higher engraftment. | No | [180] |
| Swine; Female | Sham n = 5 Hydrogel n = 7 Cardiac spheroid n = 8 | HF | 3D spheroids injection device | hiPSC-CMs | Yes; NS | Cardiac function; infarct size reduction. | Ventricular arrhythmias | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Falguera, D.; Iborra-Egea, O.; Gálvez-Montón, C. iPSC Therapy for Myocardial Infarction in Large Animal Models: Land of Hope and Dreams. Biomedicines 2021, 9, 1836. https://doi.org/10.3390/biomedicines9121836
Martínez-Falguera D, Iborra-Egea O, Gálvez-Montón C. iPSC Therapy for Myocardial Infarction in Large Animal Models: Land of Hope and Dreams. Biomedicines. 2021; 9(12):1836. https://doi.org/10.3390/biomedicines9121836
Chicago/Turabian StyleMartínez-Falguera, Daina, Oriol Iborra-Egea, and Carolina Gálvez-Montón. 2021. "iPSC Therapy for Myocardial Infarction in Large Animal Models: Land of Hope and Dreams" Biomedicines 9, no. 12: 1836. https://doi.org/10.3390/biomedicines9121836
APA StyleMartínez-Falguera, D., Iborra-Egea, O., & Gálvez-Montón, C. (2021). iPSC Therapy for Myocardial Infarction in Large Animal Models: Land of Hope and Dreams. Biomedicines, 9(12), 1836. https://doi.org/10.3390/biomedicines9121836