
Heat Shock Protein 27 Affects Myeloid Cell Activation and Interaction with Prostate Cancer Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Oxidant Exposure and Hsp27 Quantification
2.3. Metabolic Activity and Viability
2.4. Analysis of Oxidation and Mitochondria
2.5. Chemokine, Cytokine, and Growth Factor Quantification
2.6. Co-Cultures
2.7. Cell Surface Marker Expression Analysis
2.8. Live-Cell High-Content Imaging
2.9. Statistical and Software Analysis
3. Results
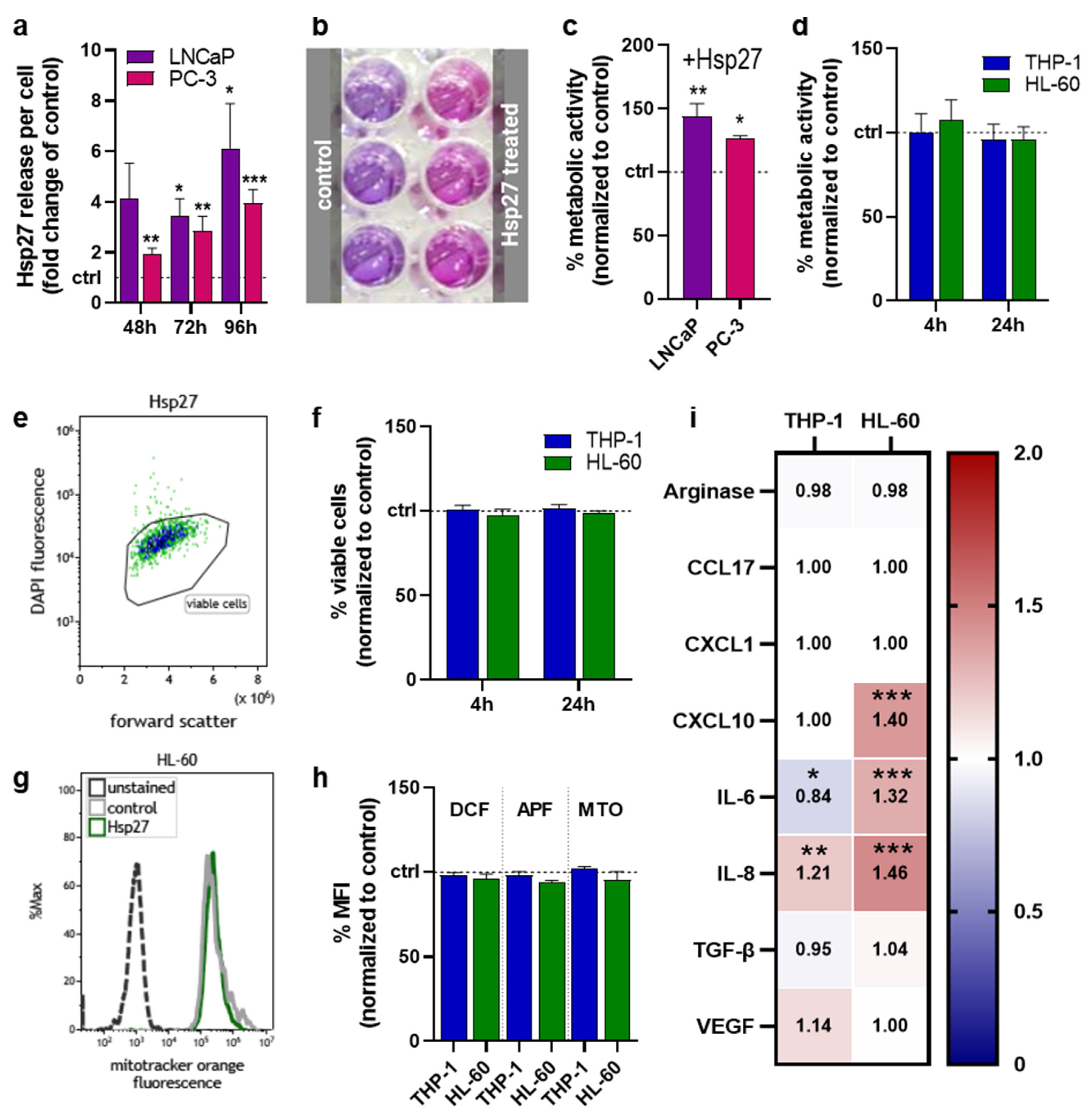
3.1. Hsp27 Is Induced in Prostate Cancer Cells upon Oxidative Stress Exposure
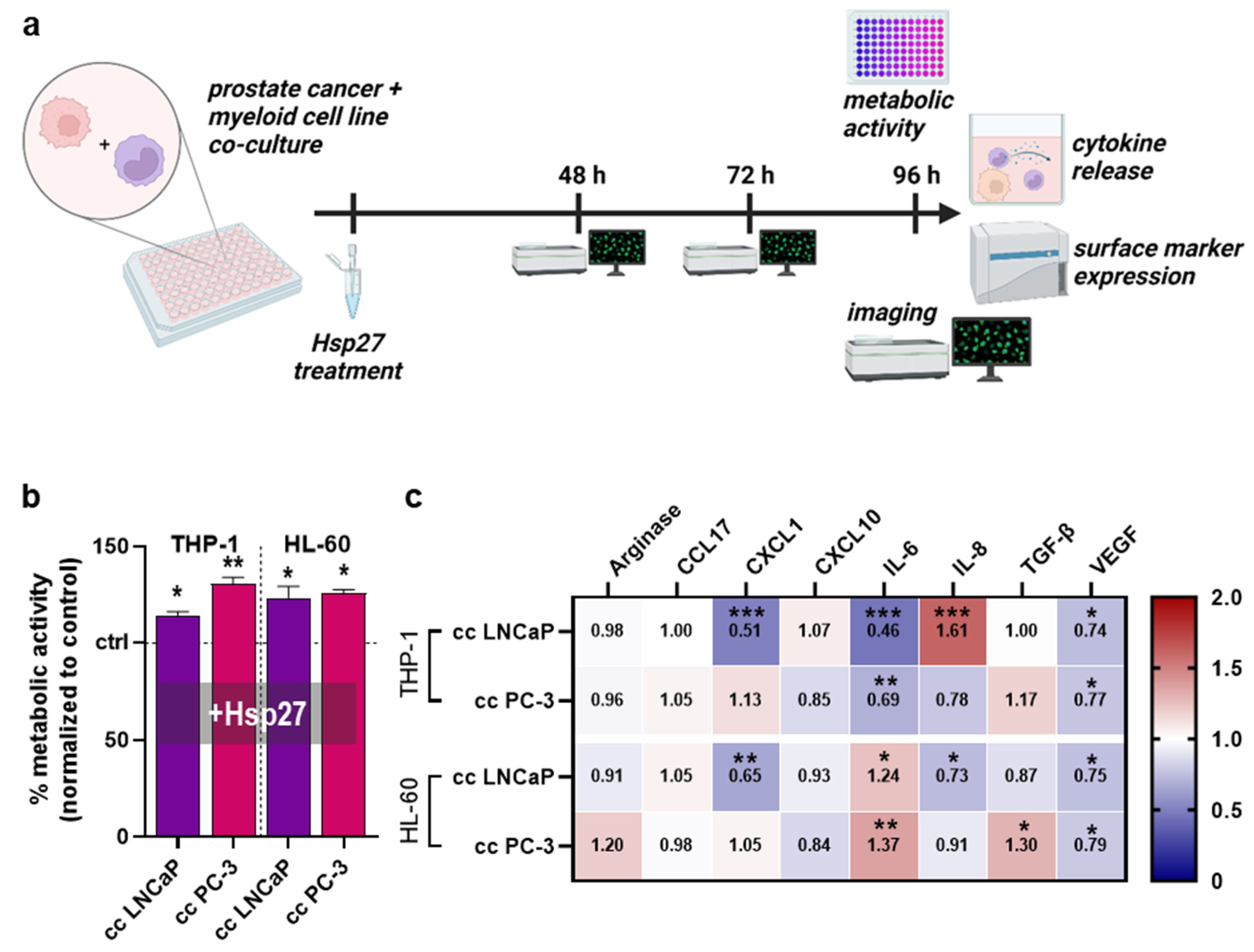
3.2. Hsp27 Altered Secretion Profiles of Prostate Cancer Cell–Myeloid Cell Co-Cultures
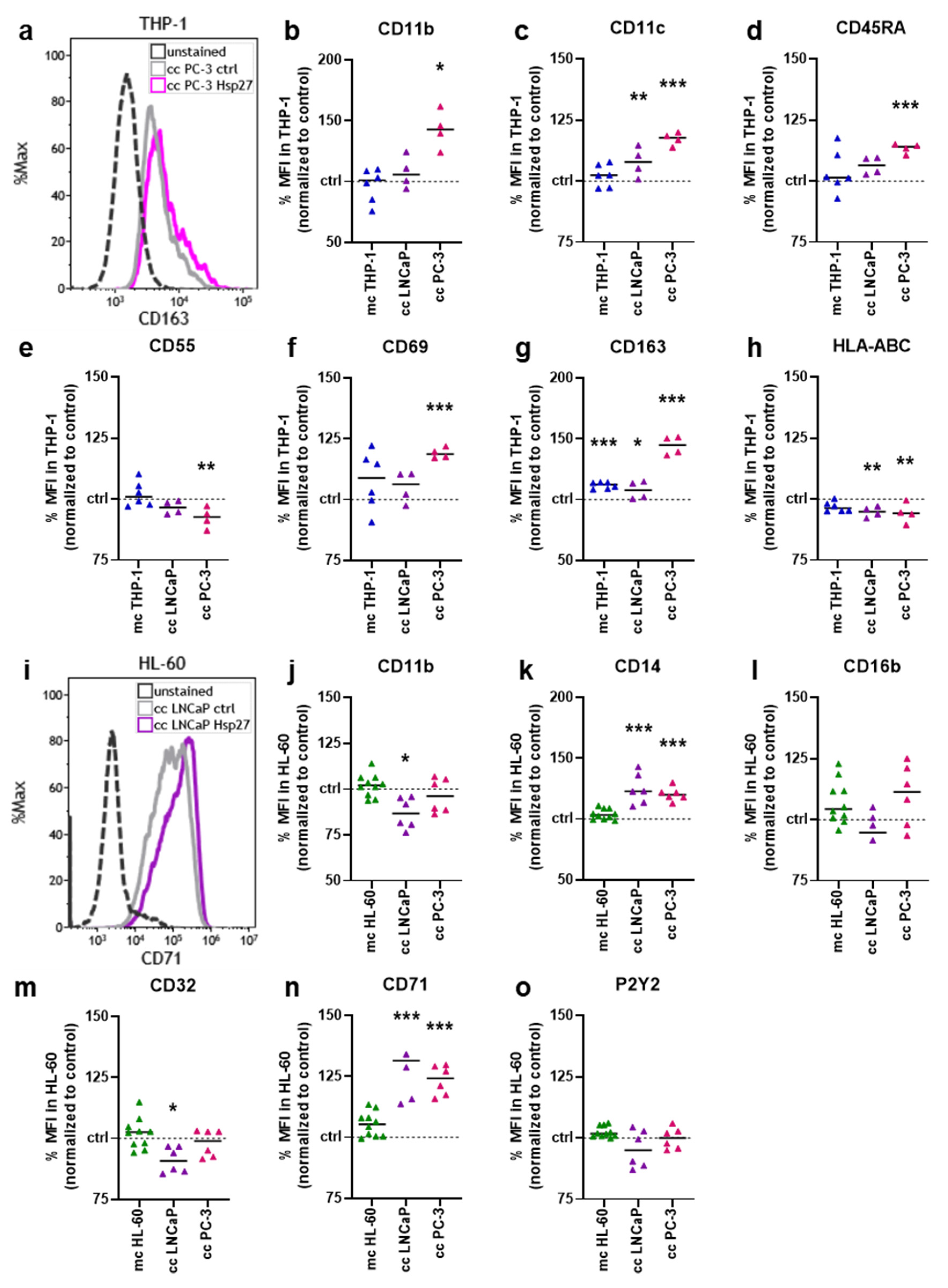
3.3. Hsp27 Altered Surface Marker Profiles of Prostate Cancer–Myeloid Cell Co-Cultures
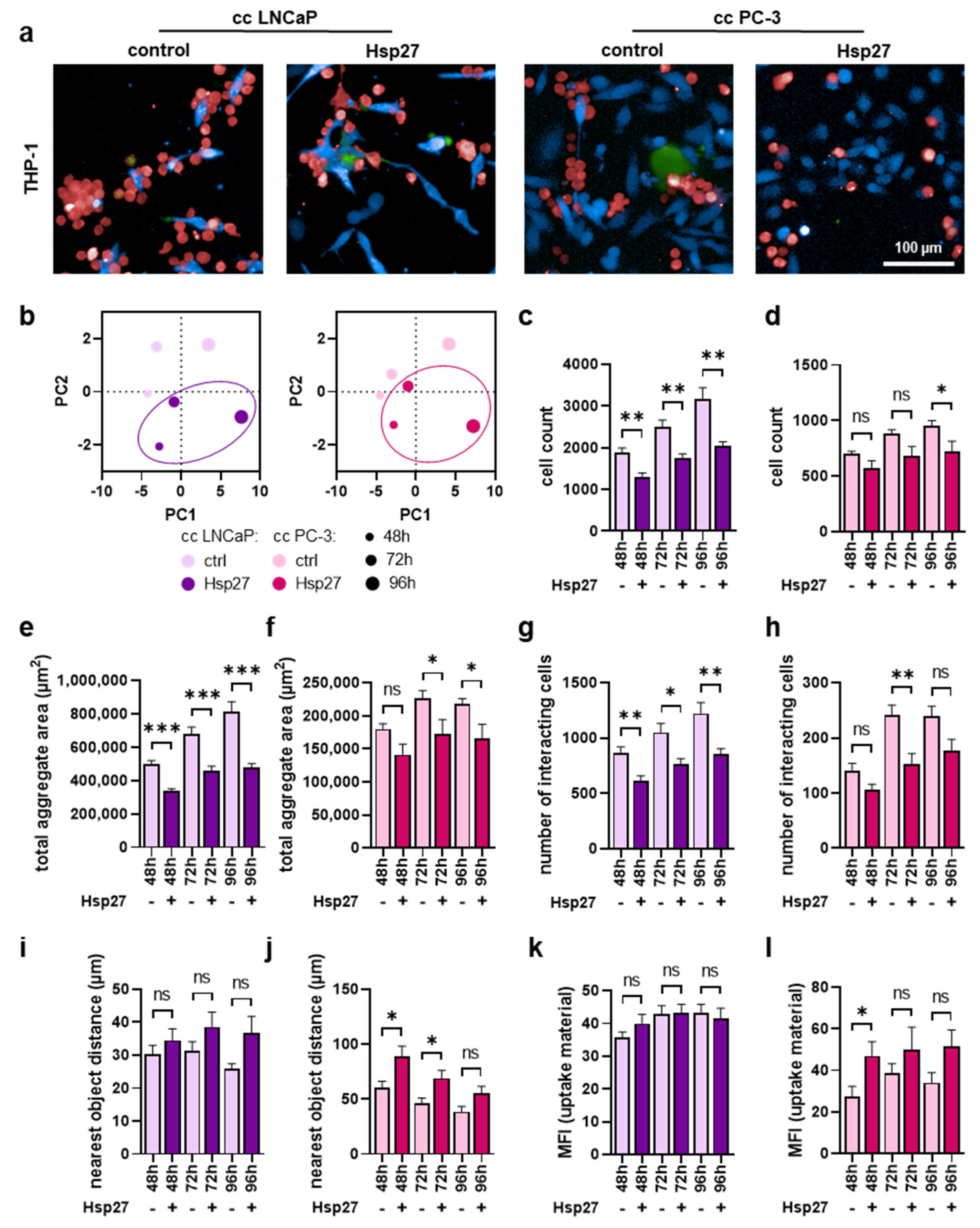
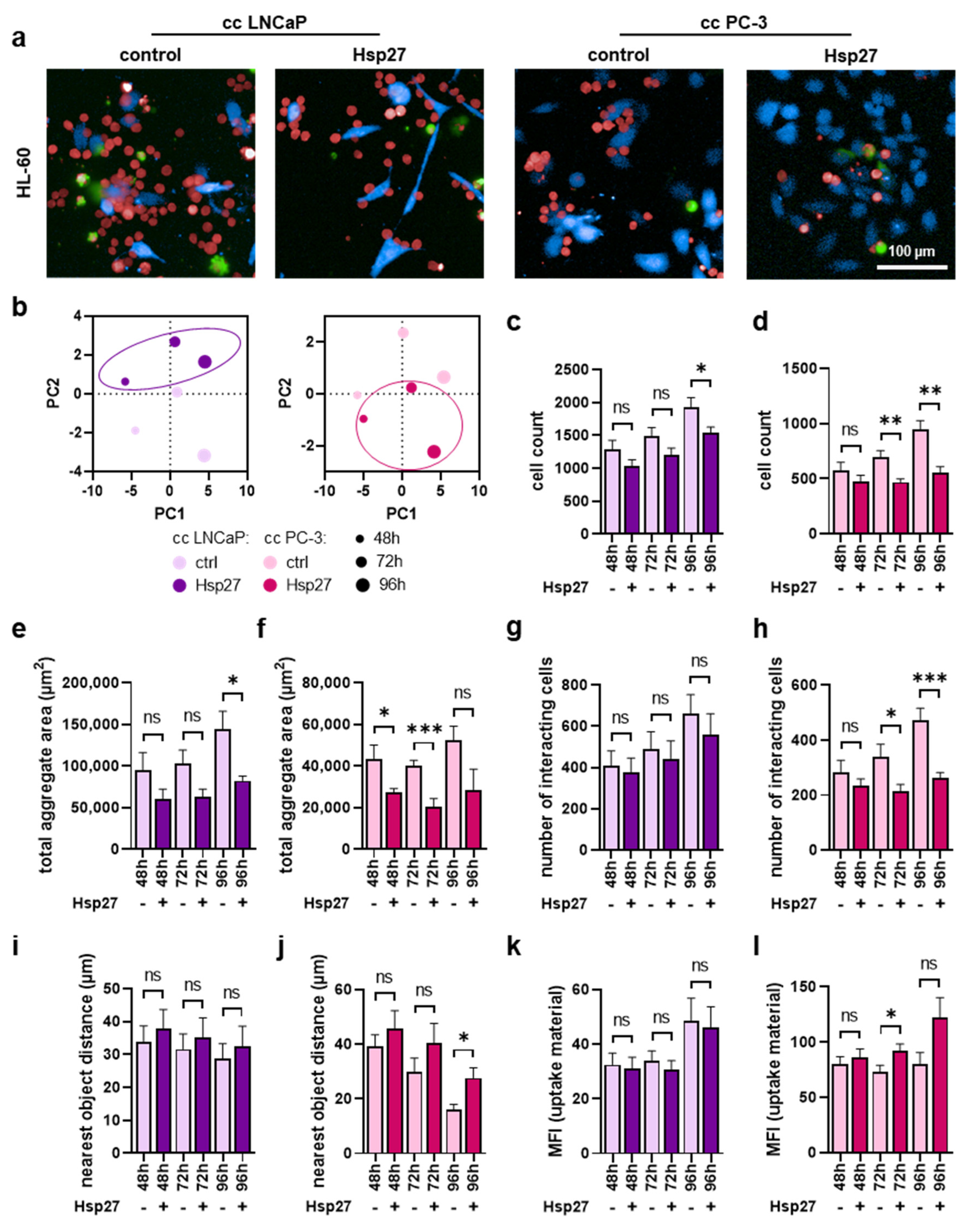
3.4. Myeloid Cell Proliferation and Interaction with Prostate Cancer Cells Were Mitigated upon Hsp27 Treatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lanneau, D.; Brunet, M.; Frisan, E.; Solary, E.; Fontenay, M.; Garrido, C. Heat shock proteins: Essential proteins for apoptosis regulation. J. Cell Mol. Med. 2008, 12, 743–761. [Google Scholar] [CrossRef]
- Kattaia, A.A.A.A.; Abd El-Baset, S.A.; Mohamed, E.M. Heat shock proteins in oxidative and nitrosative stress. In Heat Shock Proteins and Stress; Asea, A.A.A., Kaur, P., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 127–138. [Google Scholar]
- Federico, A.; Tuccillo, C.; Terracciano, F.; D’Alessio, C.; Galdiero, M.; Finamore, E.; D’Isanto, M.; Peluso, L.; Del Vecchio Blanco, C.; Loguercio, C. Heat shock protein 27 expression in patients with chronic liver damage. Immunobiology 2005, 209, 729–735. [Google Scholar] [CrossRef]
- Nguyen, A.; Chen, P.; Cai, H. Role of camkii in hydrogen peroxide activation of erk1/2, p38 mapk, hsp27 and actin reorganization in endothelial cells. FEBS Lett. 2004, 572, 307–313. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. Ros function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Sies, H. Biochemistry of oxidative stress. Angew. Chem. Int. Ed. Engl. 1986, 25, 1058–1071. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Garrido, C.; Bruey, J.M.; Fromentin, A.; Hammann, A.; Arrigo, A.P.; Solary, E. Hsp27 inhibits cytochrome c-dependent activation of procaspase-9. FASEB J. 1999, 13, 2061–2070. [Google Scholar] [CrossRef]
- Arrigo, A.P. Hsp27: Novel regulator of intracellular redox state. IUBMB Life 2001, 52, 303–307. [Google Scholar] [CrossRef]
- Vidyasagar, A.; Wilson, N.A.; Djamali, A. Heat shock protein 27 (hsp27): Biomarker of disease and therapeutic target. Fibrogenesis Tissue Repair 2012, 5, 7. [Google Scholar] [CrossRef]
- Mehlen, P.; Kretz-Remy, C.; Préville, X.; Arrigo, A.P. Human hsp27, drosophila hsp27 and human alphab-crystallin expression-mediated increase in glutathione is essential for the protective activity of these proteins against tnfalpha-induced cell death. EMBO J. 1996, 15, 2695–2706. [Google Scholar] [CrossRef]
- Arrigo, A.P.; Virot, S.; Chaufour, S.; Firdaus, W.; Kretz-Remy, C.; Diaz-Latoud, C. Hsp27 consolidates intracellular redox homeostasis by upholding glutathione in its reduced form and by decreasing iron intracellular levels. Antioxid. Redox Signal. 2005, 7, 414–422. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Du, J.; Gu, Z.; Zhao, Y. Reactive oxygen species-regulating strategies based on nanomaterials for disease treatment. Adv. Sci. 2021, 8, 2002797. [Google Scholar] [CrossRef]
- Conklin, K.A. Chemotherapy-associated oxidative stress: Impact on chemotherapeutic effectiveness. Integr. Cancer Ther. 2004, 3, 294–300. [Google Scholar] [CrossRef]
- Lampros, M.; Vlachos, N.; Voulgaris, S.; Alexiou, G.A. The role of hsp27 in chemotherapy resistance. Biomedicines 2022, 10, 897. [Google Scholar] [CrossRef]
- Brany, D.; Dvorska, D.; Halasova, E.; Skovierova, H. Cold atmospheric plasma: A powerful tool for modern medicine. Int. J. Mol. Sci. 2020, 21, 2932. [Google Scholar] [CrossRef]
- Wende, K.; von Woedtke, T.; Weltmann, K.D.; Bekeschus, S. Chemistry and biochemistry of cold physical plasma derived reactive species in liquids. Biol. Chem. 2018, 400, 19–38. [Google Scholar] [CrossRef]
- Schutz, C.S.; Stope, M.B.; Bekeschus, S. H2a.X phosphorylation in oxidative stress and risk assessment in plasma medicine. Oxid. Med. Cell. Longev. 2021, 2021, 2060986. [Google Scholar] [CrossRef]
- Bekeschus, S.; Moritz, J.; Helfrich, I.; Boeckmann, L.; Weltmann, K.-D.; Emmert, S.; Metelmann, H.-R.; Stoffels, I.; von Woedtke, T. Ex vivo exposure of human melanoma tissue to cold physical plasma elicits apoptosis and modulates inflammation. Appl. Sci. 2020, 10, 1971. [Google Scholar] [CrossRef]
- Miebach, L.; Freund, E.; Horn, S.; Niessner, F.; Sagwal, S.K.; von Woedtke, T.; Emmert, S.; Weltmann, K.D.; Clemen, R.; Schmidt, A.; et al. Tumor cytotoxicity and immunogenicity of a novel v-jet neon plasma source compared to the kinpen. Sci. Rep. 2021, 11, 136. [Google Scholar] [CrossRef]
- Weiss, M.; Gumbel, D.; Hanschmann, E.M.; Mandelkow, R.; Gelbrich, N.; Zimmermann, U.; Walther, R.; Ekkernkamp, A.; Sckell, A.; Kramer, A.; et al. Cold atmospheric plasma treatment induces anti-proliferative effects in prostate cancer cells by redox and apoptotic signaling pathways. PLoS ONE 2015, 10, e0130350. [Google Scholar] [CrossRef]
- Semmler, M.L.; Bekeschus, S.; Schafer, M.; Bernhardt, T.; Fischer, T.; Witzke, K.; Seebauer, C.; Rebl, H.; Grambow, E.; Vollmar, B.; et al. Molecular mechanisms of the efficacy of cold atmospheric pressure plasma (cap) in cancer treatment. Cancers 2020, 12, 269. [Google Scholar] [CrossRef]
- Bekeschus, S.; Clemen, R.; Niessner, F.; Sagwal, S.K.; Freund, E.; Schmidt, A. Medical gas plasma jet technology targets murine melanoma in an immunogenic fashion. Adv. Sci. 2020, 7, 1903438. [Google Scholar] [CrossRef]
- Privat-Maldonado, A.; Schmidt, A.; Lin, A.; Weltmann, K.D.; Wende, K.; Bogaerts, A.; Bekeschus, S. Ros from physical plasmas: Redox chemistry for biomedical therapy. Oxid. Med. Cell. Longev. 2019, 2019, 9062098. [Google Scholar] [CrossRef]
- Pinthus, J.H.; Bryskin, I.; Trachtenberg, J.; Lu, J.P.; Singh, G.; Fridman, E.; Wilson, B.C. Androgen induces adaptation to oxidative stress in prostate cancer: Implications for treatment with radiation therapy. Neoplasia 2007, 9, 68–80. [Google Scholar] [CrossRef]
- Aloy, M.T.; Hadchity, E.; Bionda, C.; Diaz-Latoud, C.; Claude, L.; Rousson, R.; Arrigo, A.P.; Rodriguez-Lafrasse, C. Protective role of hsp27 protein against gamma radiation-induced apoptosis and radiosensitization effects of hsp27 gene silencing in different human tumor cells. Int. J. Radiat. Oncol. Biol. Phys. 2008, 70, 543–553. [Google Scholar] [CrossRef]
- Rocchi, P.; So, A.; Kojima, S.; Signaevsky, M.; Beraldi, E.; Fazli, L.; Hurtado-Coll, A.; Yamanaka, K.; Gleave, M. Heat shock protein 27 increases after androgen ablation and plays a cytoprotective role in hormone-refractory prostate cancer. Cancer Res. 2004, 64, 6595–6602. [Google Scholar] [CrossRef]
- Rocchi, P.; Beraldi, E.; Ettinger, S.; Fazli, L.; Vessella, R.L.; Nelson, C.; Gleave, M. Increased hsp27 after androgen ablation facilitates androgen-independent progression in prostate cancer via signal transducers and activators of transcription 3–mediated suppression of apoptosis. Cancer Res. 2005, 65, 11083–11093. [Google Scholar] [CrossRef]
- Lin, L.; Liu, A.; Peng, Z.; Lin, H.-J.; Li, P.-K.; Li, C.; Lin, J. Stat3 is necessary for proliferation and survival in colon cancer–initiating cells. Cancer Res. 2011, 71, 7226–7237. [Google Scholar] [CrossRef]
- Schwager, J.; Bompard, A.; Raederstorff, D.; Hug, H.; Bendik, I. Resveratrol and omega-3 pufas promote human macrophage differentiation and function. Biomedicines 2022, 10, 1524. [Google Scholar] [CrossRef]
- Tamassia, N.; Le Moigne, V.; Calzetti, F.; Donini, M.; Gasperini, S.; Ear, T.; Cloutier, A.; Martinez, F.O.; Fabbri, M.; Locati, M.; et al. The myd88-independent pathway is not mobilized in human neutrophils stimulated via tlr4. J. Immunol. 2007, 178, 7344–7356. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Guo, S.; Stiles, J.K. The emerging role of cxcl10 in cancer (review). Oncol. Lett. 2011, 2, 583–589. [Google Scholar] [CrossRef]
- Ha, H.; Debnath, B.; Neamati, N. Role of the cxcl8-cxcr1/2 axis in cancer and inflammatory diseases. Theranostics 2017, 7, 1543–1588. [Google Scholar] [CrossRef]
- Ferrara, N.; Gerber, H.-P.; LeCouter, J. The biology of vegf and its receptors. Nat. Med. 2003, 9, 669–676. [Google Scholar] [CrossRef]
- Aragon-Ching, J.B.; Dahut, W.L. Vegf inhibitors and prostate cancer therapy. Curr. Mol. Pharmacol. 2009, 2, 161–168. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the il-6/jak/stat3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Cao, L.; Yang, M.; Tang, D.; Kang, R.; Min, X.; Zhu, S.; Yu, Y. Hsp27: A novel therapeutic target for pediatric m4/m5 acute myeloid leukemia. Oncol. Rep. 2013, 29, 1459–1466. [Google Scholar] [CrossRef] [PubMed]
- Schepers, H.; Geugien, M.; van der Toorn, M.; Bryantsev, A.L.; Kampinga, H.H.; Eggen, B.J.; Vellenga, E. Hsp27 protects aml cells against vp-16-induced apoptosis through modulation of p38 and c-jun. Exp. Hematol. 2005, 33, 660–670. [Google Scholar] [CrossRef]
- Zeng, C.; Wang, W.; Yu, X.; Yang, L.; Chen, S.; Li, Y. Pathways related to pma-differentiated thp1 human monocytic leukemia cells revealed by rna-seq. Sci. China Life Sci. 2015, 58, 1282–1287. [Google Scholar] [CrossRef]
- Corbi, A.L.; Lopez-Rodriguez, C. Cd11c integrin gene promoter activity during myeloid differentiation. Leuk Lymphoma 1997, 25, 415–425. [Google Scholar] [CrossRef]
- Sancho, D.; Gomez, M.; Sanchez-Madrid, F. Cd69 is an immunoregulatory molecule induced following activation. Trends Immunol. 2005, 26, 136–140. [Google Scholar] [CrossRef]
- Baxter, E.W.; Graham, A.E.; Re, N.A.; Carr, I.M.; Robinson, J.I.; Mackie, S.L.; Morgan, A.W. Standardized protocols for differentiation of thp-1 cells to macrophages with distinct m(ifnγ+lps), m(il-4) and m(il-10) phenotypes. J. Immunol. Methods 2020, 478, 112721. [Google Scholar] [CrossRef]
- Choo, S.Y. The hla system: Genetics, immunology, clinical testing, and clinical implications. Yonsei Med. J. 2007, 48, 11–23. [Google Scholar] [CrossRef]
- Buxadé, M.; Huerga Encabo, H.; Riera-Borrull, M.; Quintana-Gallardo, L.; López-Cotarelo, P.; Tellechea, M.; Martínez-Martínez, S.; Redondo, J.M.; Martín-Caballero, J.; Flores, J.M.; et al. Macrophage-specific mhcii expression is regulated by a remote ciita enhancer controlled by nfat5. J. Exp. Med. 2018, 215, 2901–2918. [Google Scholar] [CrossRef]
- Hmama, Z.; Nandan, D.; Sly, L.; Knutson, K.L.; Herrera-Velit, P.; Reiner, N.E. 1α,25-dihydroxyvitamin d3–induced myeloid cell differentiation is regulated by a vitamin d receptor–phosphatidylinositol 3-kinase signaling complex. J. Exp. Med. 1999, 190, 1583–1594. [Google Scholar] [CrossRef]
- Watanabe, M.; Shimamoto, Y.; Yoshida, S.; Suga, K.; Mizuguchi, Y.; Kohashi, O.; Yamaguchi, M. Intracellular multiplication of legionella pneumophila in hl-60 cells differentiated by 1,25-dihydroxyvitamin d3 and the effect of interferon gamma. J. Leukoc. Biol. 1993, 54, 40–46. [Google Scholar] [CrossRef]
- Benito, A.; Grillot, D.; Nuñez, G.; Fernández-Luna, J.L. Regulation and function of bcl-2 during differentiation-induced cell death in hl-60 promyelocytic cells. Am. J. Pathol. 1995, 146, 481–490. [Google Scholar]
- Wynn, T.A. Myeloid-cell differentiation redefined in cancer. Nat. Immunol. 2013, 14, 197–199. [Google Scholar] [CrossRef]
- Aldo, P.B.; Craveiro, V.; Guller, S.; Mor, G. Effect of culture conditions on the phenotype of thp-1 monocyte cell line. Am. J. Reprod. Immunol. 2013, 70, 80–86. [Google Scholar] [CrossRef]
- Salari, S.; Seibert, T.; Chen, Y.X.; Hu, T.; Shi, C.; Zhao, X.; Cuerrier, C.M.; Raizman, J.E.; O’Brien, E.R. Extracellular hsp27 acts as a signaling molecule to activate nf-kappab in macrophages. Cell Stress Chaperones 2013, 18, 53–63. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Fluorochrome | Clone | Vendor | Cat# |
|---|---|---|---|---|
| CD11b | APC-Cy7 | ICRF44 | BD Biosciences, Heidelberg, Germany | 557754 |
| CD11c | eFluor450 | 3.9 | Thermo Fisher Scientific, Dreieich, Germany | 48-0116-42 |
| CD14 | AF700 | 61D3 | Thermo Fisher Scientific, Dreieich, Germany | 56-0149-42 |
| CD16b | PE | CLB-gran11.5 | BD Biosciences, Heidelberg, Germany | 550868 |
| CD32 | PE/Cy7 | FUN-2 | BioLegend, Amsterdam, The Netherlands | 303214 |
| CD45RA | AF700 | HI100 | BD Biosciences, Heidelberg, Germany | 560673 |
| CD55 | PE-Cy5.5 | IA10 | BD Biosciences, Heidelberg, Germany | 555695 |
| CD69 | BV650 | FN50 | BD Biosciences, Heidelberg, Germany | 563835 |
| CD71 | APC | M-A712 | BD Biosciences, Heidelberg, Germany | 551374 |
| CD163 | PE | GHI/61 | BD Biosciences, Heidelberg, Germany | 556018 |
| HLA-ABC | APC | G46-2.6 | BD Biosciences, Heidelberg, Germany | 555555 |
| P2Y2 | AF488 | E-3 | Santa Cruz Biotechnology, Heidelberg, Germany | sc-518019 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singer, D.; Ressel, V.; Stope, M.B.; Bekeschus, S. Heat Shock Protein 27 Affects Myeloid Cell Activation and Interaction with Prostate Cancer Cells. Biomedicines 2022, 10, 2192. https://doi.org/10.3390/biomedicines10092192
Singer D, Ressel V, Stope MB, Bekeschus S. Heat Shock Protein 27 Affects Myeloid Cell Activation and Interaction with Prostate Cancer Cells. Biomedicines. 2022; 10(9):2192. https://doi.org/10.3390/biomedicines10092192
Chicago/Turabian StyleSinger, Debora, Verena Ressel, Matthias B. Stope, and Sander Bekeschus. 2022. "Heat Shock Protein 27 Affects Myeloid Cell Activation and Interaction with Prostate Cancer Cells" Biomedicines 10, no. 9: 2192. https://doi.org/10.3390/biomedicines10092192
APA StyleSinger, D., Ressel, V., Stope, M. B., & Bekeschus, S. (2022). Heat Shock Protein 27 Affects Myeloid Cell Activation and Interaction with Prostate Cancer Cells. Biomedicines, 10(9), 2192. https://doi.org/10.3390/biomedicines10092192