
The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
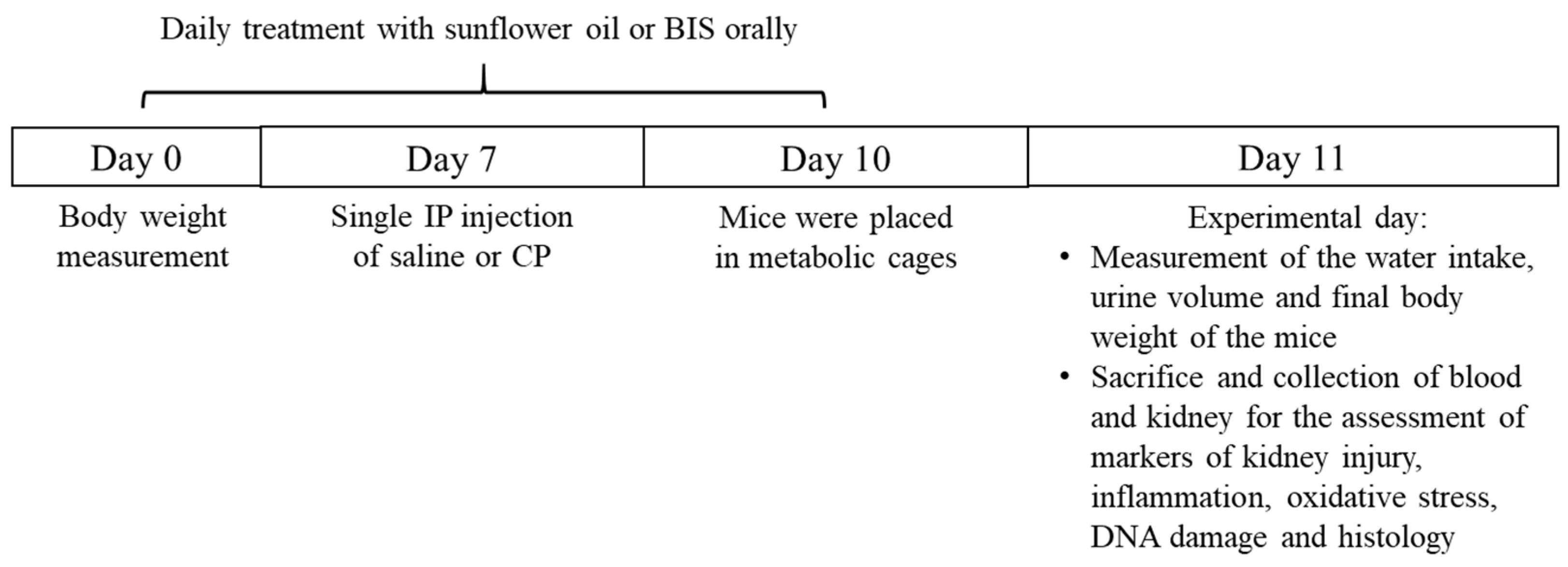
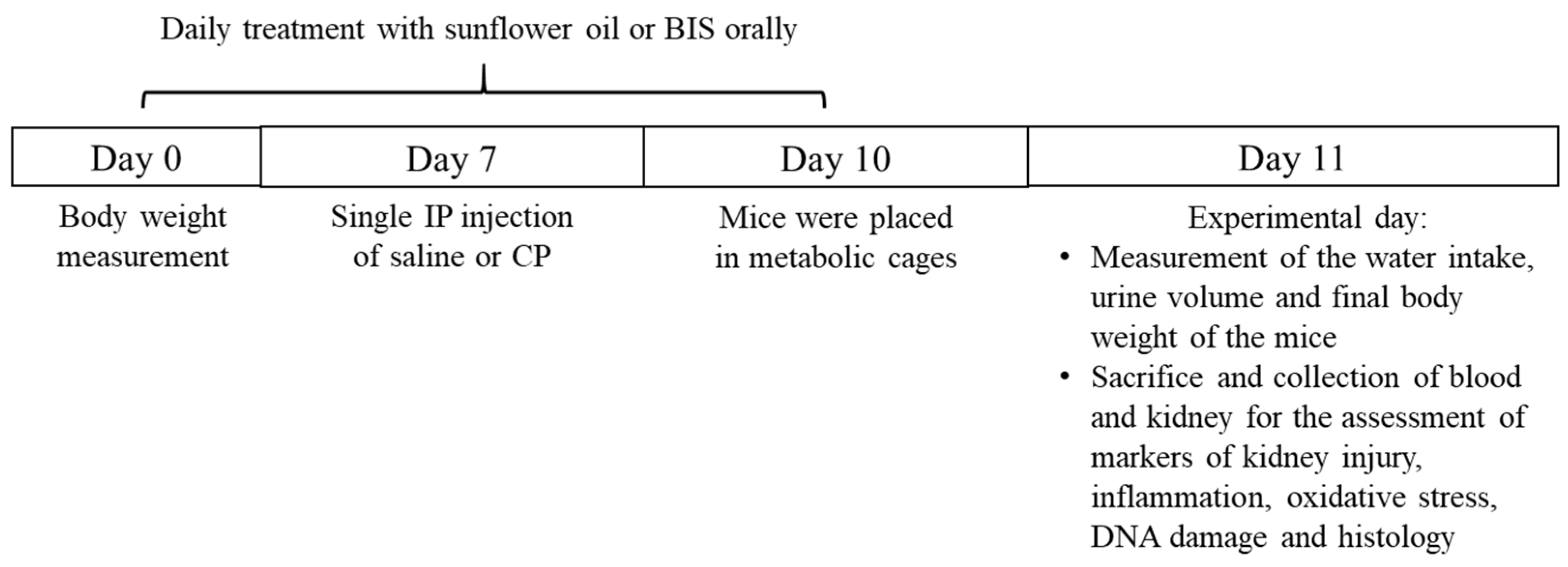
2.2. Experimental Design
2.3. Homogenization of Kidney
2.4. Biochemical Analysis
2.5. Measurement of Markers of Inflammation, Kidney Injury and Oxidative Stress in Kidney Homogenates, Plasma, and Urine
2.6. Measurement of Thiobarbituric Acid Reactive Substances (TBARS) and Reduced Glutathione (GSH) in Kidney Homogenates
2.7. Assessment of Pteridine in Urine
2.8. DNA Damage Assessment by Comet Assay
2.9. Measurement of Phosphorylated NF-κB in Kidney Homogenates
2.10. Histopathology
2.11. Statistics
3. Results
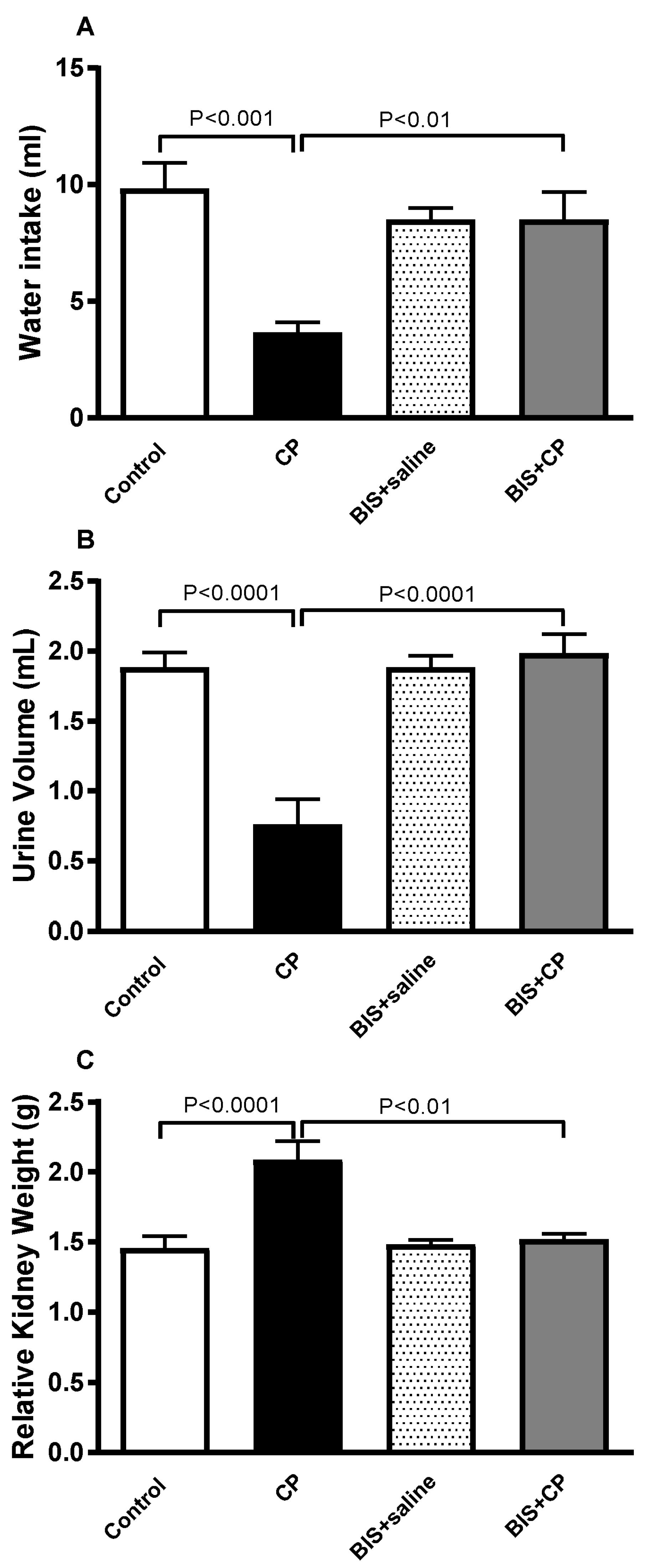
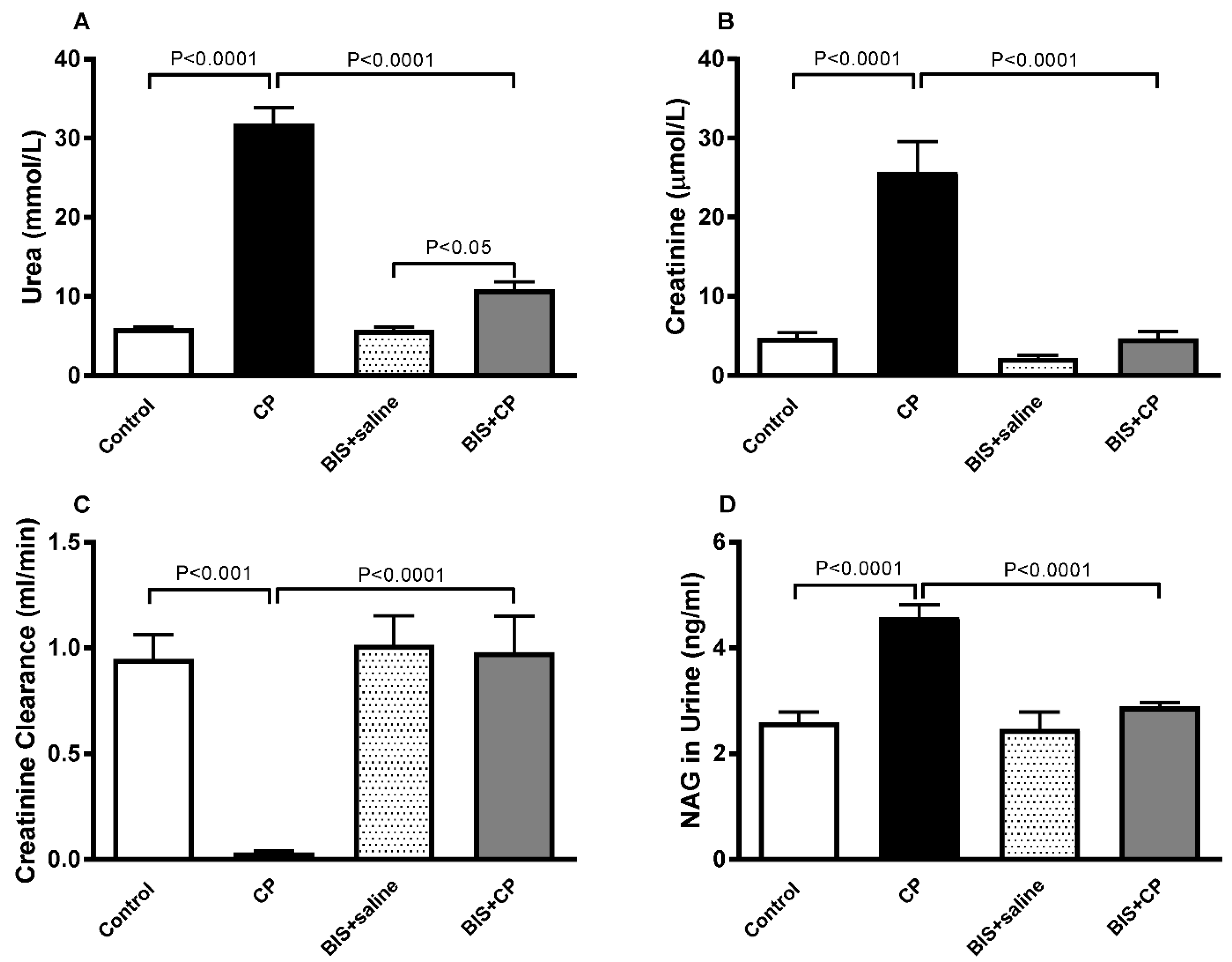
3.1. The Effects of BIS on Physiological and Biochemical Parameters
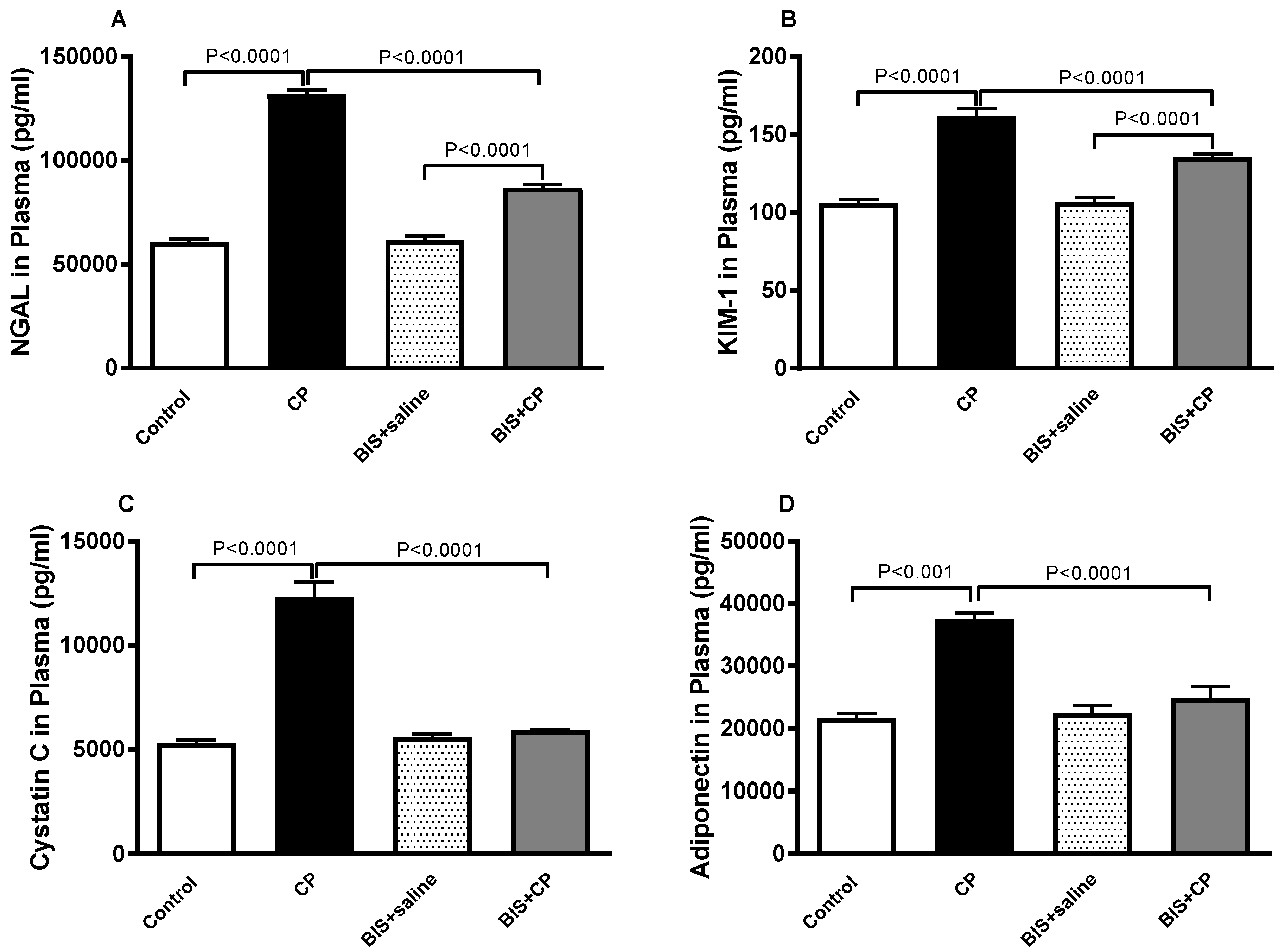
3.2. Effects of BIS on NGAL, KIM-1, Cystatin C and Adiponectin in Plasma
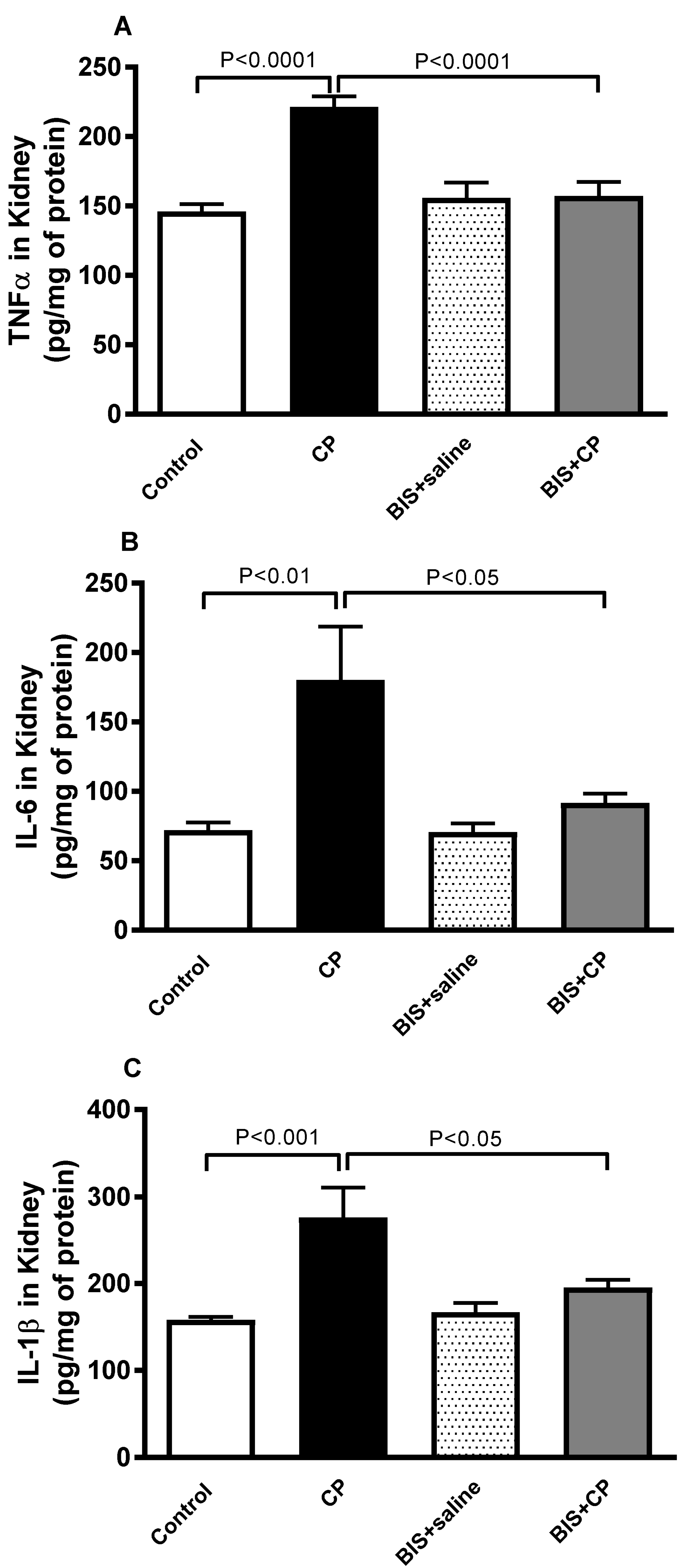
3.3. Effects of BIS on the Concentrations of Pro-Inflammatory Cytokines in Kidney Homogenate
3.4. Antioxidant Effects of BIS in Kidney
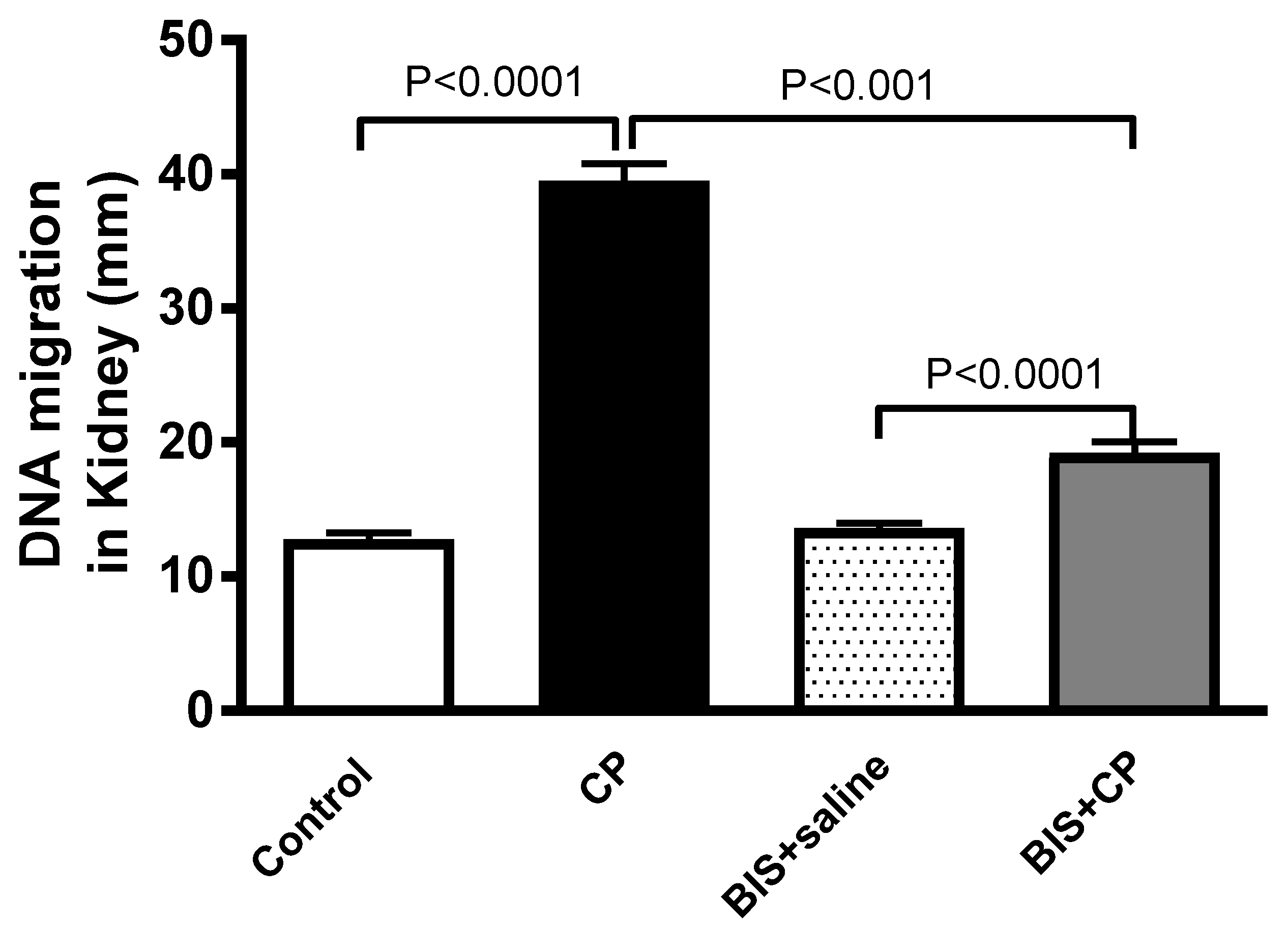
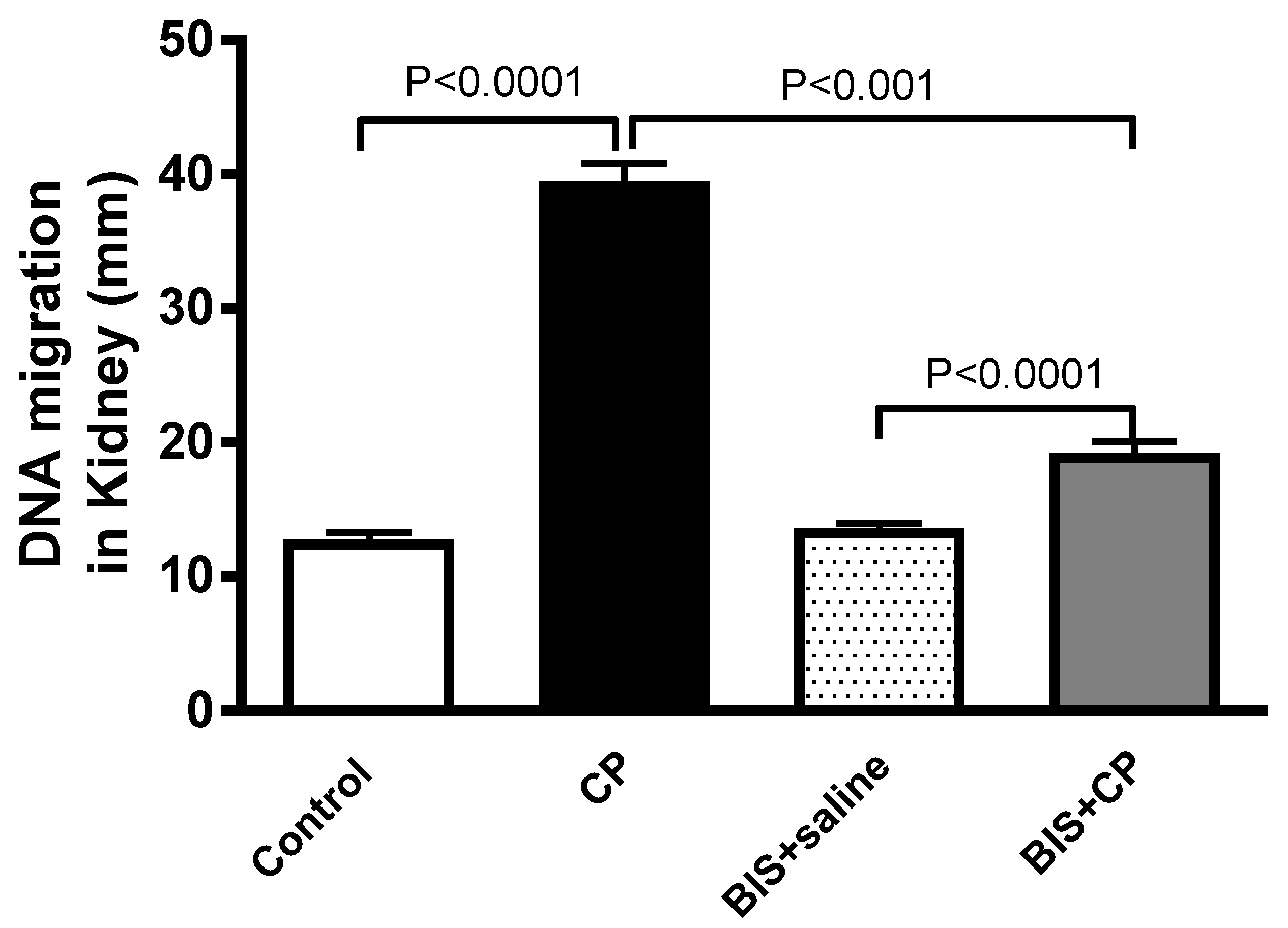
3.5. Effects of BIS on Kidney DNA Damage
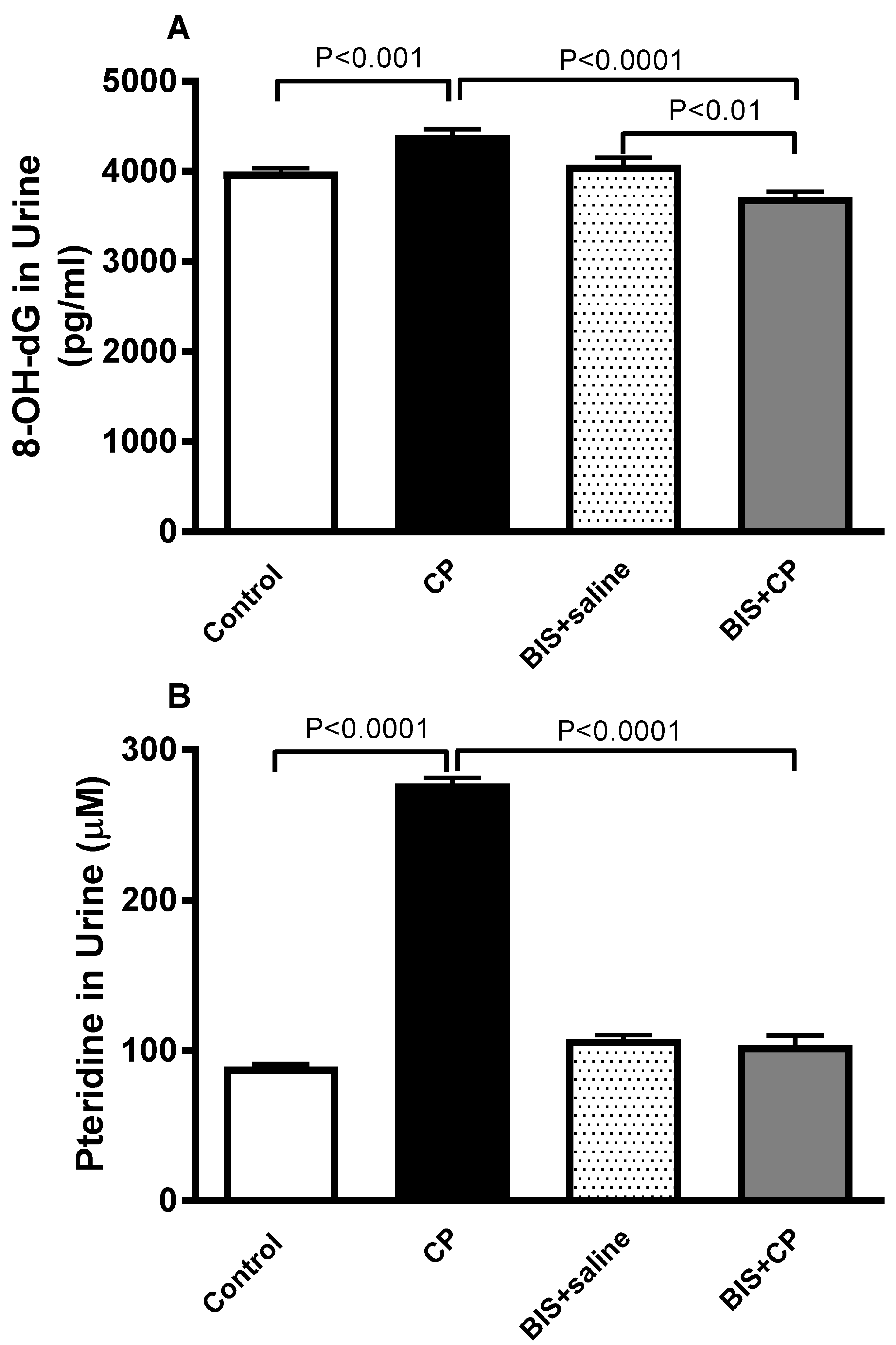
3.6. Effects of BIS on 8-OH-dG and Pteridine Concentrations in Urine
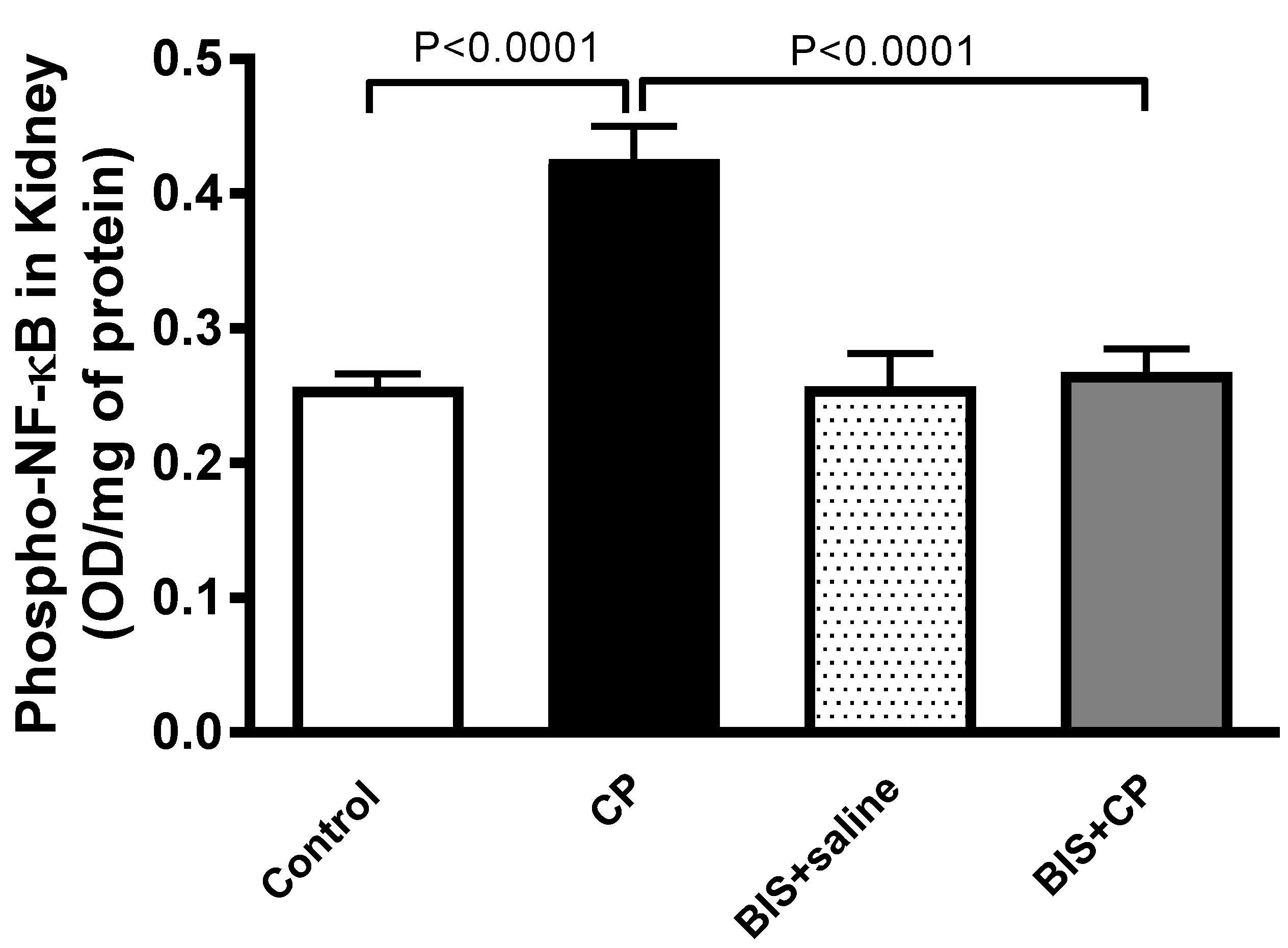
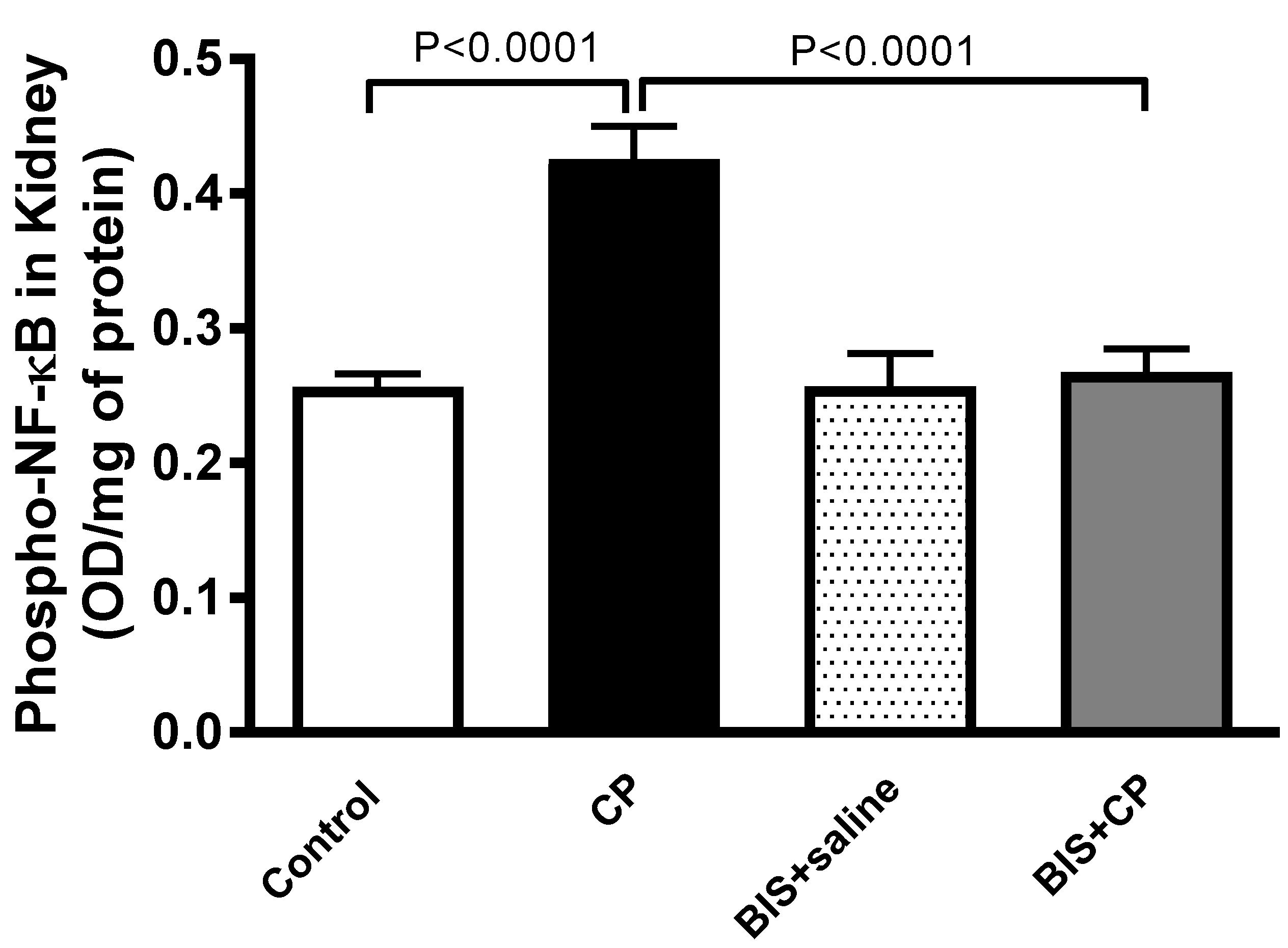
3.7. Effects of BIS of the Expression of Phopho-NF-κB in Kidney Homogenate
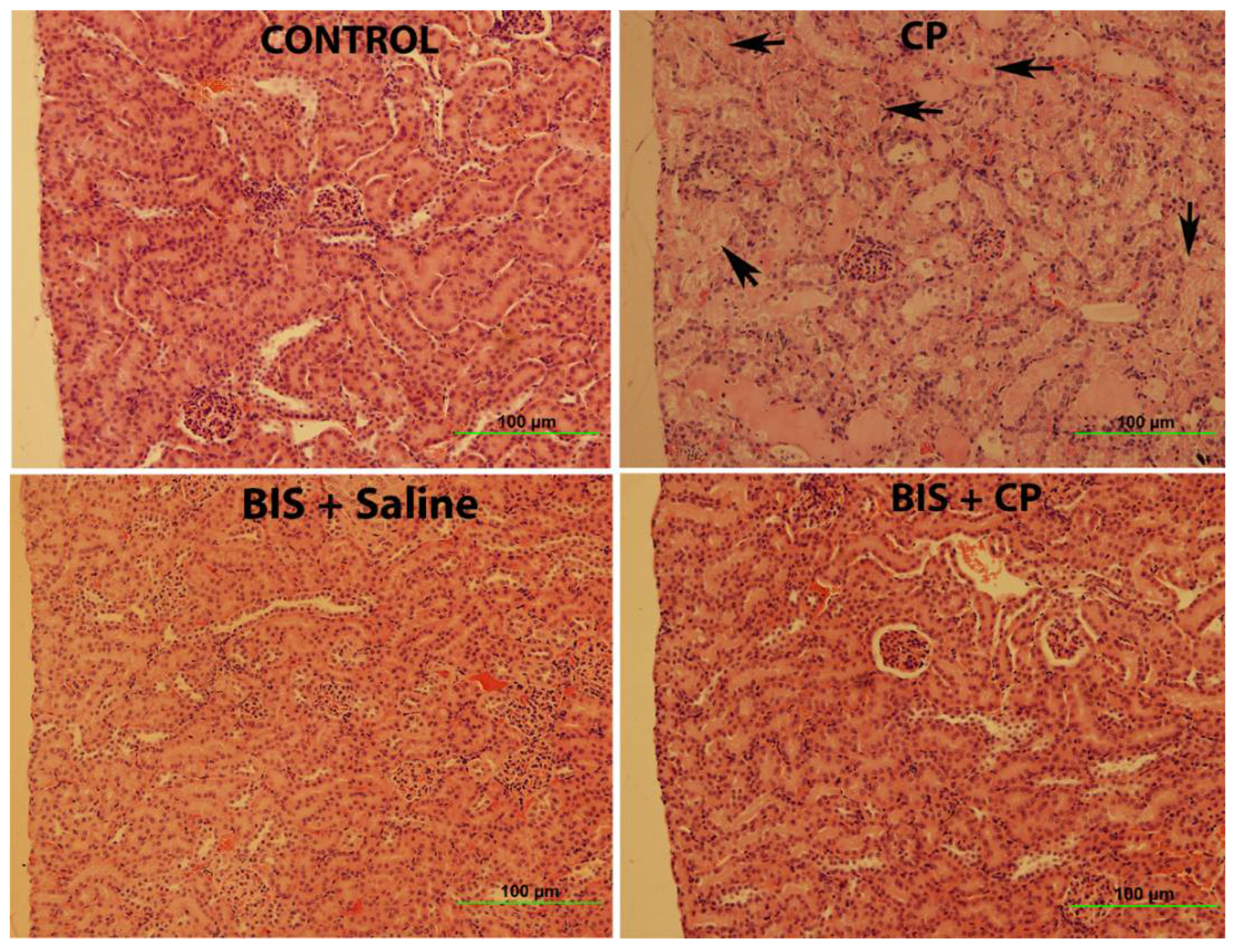
3.8. Effects of BIS on Kidney Histology Assessed by Light Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perše, M.; Večerić-Haler, Ž. Cisplatin-Induced Rodent Model of Kidney Injury: Characteristics and Challenges. BioMed Res. Int. 2018, 2018, 1462802. [Google Scholar] [CrossRef]
- Pabla, N.; Dong, Z. Cisplatin nephrotoxicity: Mechanisms and renoprotective strategies. Kidney Int. 2008, 73, 994–1007. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.L.C.; de Brito, B.B.; da Silva, F.A.F.; Botelho, A.C.D.S.; de Melo, F.F. Nephrotoxicity in cancer treatment: An overview. World J. Clin. Oncol. 2020, 11, 190–204. [Google Scholar] [CrossRef]
- Yao, X.; Panichpisal, K.; Kurtzman, N.; Nugent, K. Cisplatin nephrotoxicity: A review. Am. J. Med. Sci. 2007, 334, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Humanes, B.; Camaño, S.; Lara, J.M.; Sabbisetti, V.; González-Nicolás, M.; Bonventre, J.V.; Tejedor, A.; Lázaro, A. Cisplatin-induced renal inflammation is ameliorated by cilastatin nephroprotection. Nephrol. Dial. Transplant. 2017, 32, 1645–1655. [Google Scholar] [CrossRef] [Green Version]
- Makris, K.; Spanou, L. Acute Kidney Injury: Definition, Pathophysiology and Clinical Phenotypes. Clin. Biochem. Rev. 2016, 37, 85–98. [Google Scholar]
- Waikar, S.S.; Bonventre, J.V. Creatinine Kinetics and the Definition of Acute Kidney Injury. J. Am. Soc. Nephrol. 2009, 20, 672–679. [Google Scholar] [CrossRef] [Green Version]
- Wainford, R.D.; Weaver, R.J.; Stewart, K.N.; Brown, P.; Hawksworth, G.M. Cisplatin nephrotoxicity is mediated by gamma glutamyltranspeptidase, not via a C-S lyase governed biotransformation pathway. Toxicology 2008, 249, 184–193. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; McMillan, K.L.; Wu, J.; Gillings, N.; Flores, B.; Moe, O.W.; Hu, M.C. Cisplatin nephrotoxicity as a model of chronic kidney disease. Lab. Investig. 2018, 98, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Chawla, L.S.; Eggers, P.W.; Star, R.A.; Kimmel, P.L. Acute Kidney Injury and Chronic Kidney Disease as Interconnected Syndromes. N. Engl. J. Med. 2014, 371, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghorbani, A.; Baradaran, A. Comment on: The effects of Vitamin E and Selenium on cisplatin-induced nephrotoxicity in cancer patients treated with cisplatin-based chemotherapy: A randomized, placebo-controlled study. J. Res. Med. Sci. 2013, 18, 922–923. [Google Scholar] [PubMed]
- Ali, B. Agents ameliorating or augmenting experimental gentamicin nephrotoxicity: Some recent research. Food Chem. Toxicol. 2003, 41, 1447–1452. [Google Scholar] [CrossRef]
- Santoso, J.T.; Lucci, J.A.; Coleman, R.L.; Schafer, I.; Hannigan, E.V. Saline, mannitol, and furosemide hydration in acute cisplatin nephrotoxicity: A randomized trial. Cancer Chemother. Pharmacol. 2003, 52, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Rjiba-Touati, K.; Ayed-Boussema, I.; Bouaziz, C.; Belarbia, A.; Azzabi, A.; Achour, A.; Hassen, W.; Bacha, H. Protective effect of erythropoietin against cisplatin-induced nephrotoxicity in rats: Antigenotoxic and antiapoptotic effect. Drug Chem. Toxicol. 2011, 35, 89–95. [Google Scholar] [CrossRef]
- Miller, R.P.; Tadagavadi, R.K.; Ramesh, G.; Reeves, W.B. Mechanisms of Cisplatin Nephrotoxicity. Toxins 2010, 2, 2490–2518. [Google Scholar] [CrossRef] [Green Version]
- Liang, Y.; Liu, H.-H.; Chen, Y.-J.; Sun, H. Antitumor Activity of the Protein and Small Molecule Component Fractions from Agrocybe aegerita Through Enhancement of Cytokine Production. J. Med. Food 2014, 17, 439–446. [Google Scholar] [CrossRef]
- De Siqueira, R.J.; Freire, W.B.; Vasconcelos-Silva, A.A.; Fonseca-Magalhães, P.A.; Lima, F.J.; Brito, T.S.; Mourao, L.T.C.; Ribeiro, R.A.; Lahlou, S.; Magalhaes, P.J.C. In-vitro characterization of the pharmacological effects induced by (−)-α-bisabolol in rat smooth muscle preparations. Can. J. Physiol. Pharmacol. 2012, 90, 23–35. [Google Scholar] [CrossRef]
- Seki, T.; Kokuryo, T.; Yokoyama, Y.; Suzuki, H.; Itatsu, K.; Nakagawa, A.; Mizutani, T.; Miyake, T.; Uno, M.; Yamauchi, K.; et al. Antitumor effects of α-bisabolol against pancreatic cancer. Cancer Sci. 2011, 102, 2199–2205. [Google Scholar] [CrossRef]
- Fernandes, M.Y.D.; Carmo, M.R.S.D.; Fonteles, A.A.; Neves, J.C.D.S.; da Silva, A.T.A.; Pereira, J.F.; Ferreira, E.D.O.; de Lima, N.M.R.; Neves, K.R.T.; de Andrade, G.M. (−)-α-bisabolol prevents neuronal damage and memory deficits through reduction of proinflammatory markers induced by permanent focal cerebral ischemia in mice. Eur. J. Pharmacol. 2019, 842, 270–280. [Google Scholar] [CrossRef]
- Braga, P.C.; Sasso, M.D.; Fonti, E.; Culici, M. Antioxidant Activity of Bisabolol: Inhibitory Effects on Chemiluminescence of Human Neutrophil Bursts and Cell-Free Systems. Pharmacology 2008, 83, 110–115. [Google Scholar] [CrossRef]
- Sampaio, T.L.; de Menezes, R.R.P.P.B.; da Costa, M.F.B.; Meneses, G.C.; Arrieta, M.C.V.; Chaves-Filho, A.J.M.; de Morais, G.B.; Libório, A.B.; Alves, R.S.; Evangelista, J.S.A.M.; et al. Nephroprotective effects of (−)-α-bisabolol against ischemic-reperfusion acute kidney injury. Phytomedicine 2016, 23, 1843–1852. [Google Scholar] [CrossRef] [PubMed]
- Maurya, A.K.; Singh, M.; Dubey, V.; Srivastava, S.; Luqman, S.; Bawankule, D.U. α-()-bisabolol reduces pro-inflammatory cytokine production and ameliorates skin inflammation. Curr. Pharm. Biotechnol. 2014, 15, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Meeran, M.F.N.; Azimullah, S.; Eddin, L.B.; Dwivedi, V.D.; Jha, N.K.; Ojha, S. α-Bisabolol, a Dietary Bioactive Phytochemical Attenuates Dopaminergic Neurodegeneration through Modulation of Oxidative Stress, Neuroinflammation and Apoptosis in Rotenone-Induced Rat Model of Parkinson’s disease. Biomolecules 2020, 10, 1421. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, G.F.D.; Da Costa, F.N.; Campos, A.R. Corneal antinociceptive effect of (−)-α-bisabolol. Pharm. Biol. 2017, 55, 1089–1092. [Google Scholar] [CrossRef]
- Janero, D.R. Malondialdehyde and thiobarbituric acid-reactivity as diagnostic indices of lipid peroxidation and peroxidative tissue injury. Free Radic. Biol. Med. 1990, 9, 515–540. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Aguilar Diaz De Leon, J.; Borges, C.R. Evaluation of Oxidative Stress in Biological Samples Using the Thiobarbituric Acid Reactive Substances Assay. J. Vis. Exp. 2020, 159, e61122. [Google Scholar] [CrossRef]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Wakabayashi, I.; Nakanishi, M.; Ohki, M.; Suehiro, A.; Uchida, K. A simple and useful method for evaluation of oxidative stress in vivo by spectrofluorometric estimation of urinary pteridines. Sci. Rep. 2020, 10, 11223. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Ali, B.H. Chronic exposure to water-pipe smoke induces cardiovascular dysfunction in mice. Am. J. Physiol. Heart Circ. Physiol. 2017, 312, H329–H339. [Google Scholar] [CrossRef]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Ali, B.H. Aortic Oxidative Stress, Inflammation and DNA Damage Following Pulmonary Exposure to Cerium Oxide Nanoparticles in a Rat Model of Vascular Injury. Biomolecules 2019, 9, 376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Suleimani, Y.M.; Al Mahruqi, A.S.; Al Za’abi, M.; Shalaby, A.; Ashique, M.; Nemmar, A.; Ali, B.H. Effect of diesel exhaust particles on renal vascular responses in rats with chronic kidney disease. Environ. Toxicol. 2017, 32, 541–549. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Raza, H.; Subramaniyan, D.; John, A.; Elwasila, M.; Ali, B.H.; Adeghate, E. Evaluation of the pulmonary effects of short-term nose-only cigarette smoke exposure in mice. Exp. Biol. Med. 2012, 237, 1449–1456. [Google Scholar] [CrossRef] [PubMed]
- Mi, X.-J.; Hou, J.-G.; Wang, Z.; Han, Y.; Ren, S.; Hu, J.-N.; Chen, C.; Li, W. The protective effects of maltol on cisplatin-induced nephrotoxicity through the AMPK-mediated PI3K/Akt and p53 signaling pathways. Sci. Rep. 2018, 8, 15922. [Google Scholar] [CrossRef] [Green Version]
- Bhatia, S.P.; McGinty, D.; Letizia, C.S.; Api, A.M. Fragrance material review on α-bisabolol. Food Chem. Toxicol. 2008, 46 (Suppl. S11), S72–S76. [Google Scholar] [CrossRef]
- Cavalieri, E.; Mariotto, S.; Fabrizi, C.; de Prati, A.C.; Gottardo, R.; Leone, S.; Berra, L.V.; Lauro, G.M.; Ciampa, A.R.; Suzuki, H. α-Bisabolol, a nontoxic natural compound, strongly induces apoptosis in glioma cells. Biochem. Biophys. Res. Commun. 2004, 315, 589–594. [Google Scholar] [CrossRef]
- Rocha, N.F.M.; Rios, E.R.V.; Carvalho, A.M.R.; Cerqueira, G.S.; de Araújo Lopes, A.; Leal, L.K.A.M.; Dias, M.L.; de Sousa, D.P.; de Sousa, F.C.F. Anti-nociceptive and anti-inflammatory activities of (−)-α-bisabolol in rodents. Naunyn-Schmiedebergs Arch. Exp. Pathol. Pharmakol. 2011, 384, 525–533. [Google Scholar] [CrossRef]
- Leite, G.e.O.; Fernandes, C.N.; de Menezes, I.R.; da Costa, J.G.; Campos, A.R. Attenuation of visceral nociception by α-bisabolol in mice: Investigation of mechanisms. Org. Med. Chem. Lett. 2012, 2, 18. [Google Scholar] [CrossRef] [Green Version]
- Meeran, M.F.N.; Laham, F.; Al-Taee, H.; Azimullah, S.; Ojha, S. Protective effects of α-bisabolol on altered hemodynamics, lipid peroxidation, and nonenzymatic antioxidants in isoproterenol-induced myocardial infarction: In vivo and in vitro evidences. J. Biochem. Mol. Toxicol. 2018, 32, e22200. [Google Scholar] [CrossRef]
- Abdelrahman, A.M.; Al Suleimani, Y.; Shalaby, A.; Ashique, M.; Manoj, P.; Nemmar, A.; Ali, B.H. Effect of canagliflozin, a sodium glucose co-transporter 2 inhibitor, on cisplatin-induced nephrotoxicity in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2019, 392, 45–53. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Malik, N.; Sanger, G.J.; Friedman, M.I.; Andrews, P.L. Pica—A model of nausea? Species differences in response to cisplatin. Physiol. Behav. 2005, 85, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Shahid, F.; Farooqui, Z.; Khan, F. Cisplatin-induced gastrointestinal toxicity: An update on possible mechanisms and on available gastroprotective strategies. Eur. J. Pharmacol. 2018, 827, 49–57. [Google Scholar] [CrossRef]
- Ali, B.H.; Al Salam, S.; Al Suleimani, Y.; Al Za’Abi, M.; Ashique, M.; Manoj, P.; Sudhadevi, M.; Al Tobi, M.; Nemmar, A. Ameliorative Effect of Sesamin in Cisplatin-Induced Nephrotoxicity in Rats by Suppressing Inflammation, Oxidative/Nitrosative Stress, and Cellular Damage. Physiol. Res. 2020, 69, 61–72. [Google Scholar] [CrossRef] [PubMed]
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Ali, B.A.; Zulli, A.; Apostolopoulos, V. Mechanisms of Cisplatin-Induced Acute Kidney Injury: Pathological Mechanisms, Pharmacological Interventions, and Genetic Mitigations. Cancers 2021, 13, 1572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ramesh, G.; Norbury, C.C.; Reeves, W.B. Cisplatin-induced nephrotoxicity is mediated by tumor necrosis factor-α produced by renal parenchymal cells. Kidney Int. 2007, 72, 37–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramesh, G.; Reeves, W.B. TNF-alpha mediates chemokine and cytokine expression and renal injury in cisplatin nephrotoxicity. J. Clin. Investig. 2002, 110, 835–842. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [Green Version]
- Nemmar, A.; Beegam, S.; Yuvaraju, P.; Yasin, J.; Shahin, A.; Ali, B.H. Interaction of amorphous silica nanoparticles with erythrocytes in vitro: Role of oxidative stress. Cell Physiol. Biochem. 2014, 34, 255–265. [Google Scholar] [CrossRef]
- Lefèvre, G.; Beljean-Leymarie, M.; Beyerle, F.; Bonnefont-Rousselot, D.; Cristol, J.P.; Thérond, P.; Torreilles, J. Evaluation of lipid peroxidation by measuring thiobarbituric acid reactive substances. Ann. Biol. Clin. 1998, 56, 305–319. [Google Scholar]
- Meng, H.; Fu, G.; Shen, J.; Shen, K.; Xu, Z.; Wang, Y.; Jin, B.; Pan, H. Ameliorative Effect of Daidzein on Cisplatin-Induced Nephrotoxicity in Mice via Modulation of Inflammation, Oxidative Stress, and Cell Death. Oxidative Med. Cell. Longev. 2017, 2017, 3140680. [Google Scholar] [CrossRef]
- Wang, S.; Xie, J.; Li, J.; Liu, F.; Wu, X.; Wang, Z. Cisplatin suppresses the growth and proliferation of breast and cervical cancer cell lines by inhibiting integrin β5-mediated glycolysis. Am. J. Cancer Res. 2016, 6, 1108–1117. [Google Scholar] [PubMed]
- Nemmar, A.; Al-Salam, S.; Yuvaraju, P.; Beegam, S.; Ali, B.H. Emodin mitigates diesel exhaust particles-induced increase in airway resistance, inflammation and oxidative stress in mice. Respir. Physiol. Neurobiol. 2015, 215, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Saad, A.A.; Youssef, M.I.; El-Shennawy, L.K. Cisplatin induced damage in kidney genomic DNA and nephrotoxicity in male rats: The protective effect of grape seed proanthocyanidin extract. Food Chem. Toxicol. 2009, 47, 1499–1506. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Al-Salam, S.; Beegam, S.; Yuvaraju, P.; Ali, B.H. Waterpipe Smoke Exposure Triggers Lung Injury and Functional Decline in Mice: Protective Effect of Gum Arabic. Oxidative Med. Cell. Longev. 2019, 2019, 8526083. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, H.; Unoda, K.; Ito, T.; Kitaoka, H.; Kimura, F.; Hanafusa, T. The Relation of Urinary 8-OHdG, A Marker of Oxidative Stress to DNA, and Clinical Outcomes for Ischemic Stroke. Open Neurol. J. 2012, 6, 51–57. [Google Scholar] [CrossRef]
- Cooke, M.S.; Evans, M.D.; Herbert, K.E.; Lunec, J. Urinary 8-oxo-2′-deoxyguanosine—Source, significance and supplements. Free Radic. Res. 2000, 32, 381–397. [Google Scholar] [CrossRef]
- Vaidya, V.S.; Waikar, S.S.; Ferguson, M.A.; Collings, F.B.; Sunderland, K.; Gioules, C.; Bradwin, G.; Matsouaka, R.; Betensky, R.A.; Curhan, G.C.; et al. Urinary Biomarkers for Sensitive and Specific Detection of Acute Kidney Injury in Humans. Clin. Transl. Sci. 2008, 1, 200–208. [Google Scholar] [CrossRef]
- Oettl, K.; Reibnegger, G. Pteridine derivatives as modulators of oxidative stress. Curr. Drug Metab. 2002, 3, 203–209. [Google Scholar] [CrossRef]
- Murr, C.; Fuith, L.C.; Widner, B.; Wirleitner, B.; Baier-Bitterlich, G.; Fuchs, D. Increased neopterin concentrations in patients with cancer: Indicator of oxidative stress? Anticancer Res. 1999, 19, 1721–1728. [Google Scholar]
- Fuchs, D.; Avanzas, P.; Arroyo-Espliguero, R.; Jenny, M.; Consuegra-Sanchez, L.; Kaski, J.C. The role of neopterin in atherogenesis and cardiovascular risk assessment. Curr. Med. Chem. 2009, 16, 4644–4653. [Google Scholar] [CrossRef]
- Robertson, J.; Gostner, J.M.; Nilsson, S.; Andersson, L.-M.; Fuchs, D.; Gisslen, M. Serum neopterin levels in relation to mild and severe COVID-19. BMC Infect. Dis. 2020, 20, 942. [Google Scholar] [CrossRef] [PubMed]
- Eisenhut, M. Neopterin in Diagnosis and Monitoring of Infectious Diseases. J. Biomark. 2013, 2013, 196432. [Google Scholar] [CrossRef] [Green Version]
- Song, N.; Thaiss, F.; Guo, L. NFκB and Kidney Injury. Front. Immunol. 2019, 10, 815. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Sheng, S.; Dou, H.; Chen, J.; Zhou, K.; Lin, Y.; Yang, H. α-Bisabolol suppresses the inflammatory response and ECM catabolism in advanced glycation end products-treated chondrocytes and attenuates murine osteoarthritis. Int. Immunopharmacol. 2020, 84, 106530. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, W.; Wang, L.; Li, W.; Sun, H.; He, X.; Zhang, J. The Protective Effects of Sika Deer Antler Protein on Cisplatin-Induced Nephrotoxicity. Cell. Physiol. Biochem. 2017, 43, 395–404. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | % of Necrotic Area | Necrosis Score |
|---|---|---|
| Control | 0 | 0 |
| CP-treated | 58 ± 3.0 **** | 3 |
| BIS-treated | 0 | 0 |
| BIS and CP-treated | 2 ± 0.6 ΔΔΔΔ | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaaba, N.E.; Beegam, S.; Elzaki, O.; Yasin, J.; Nemmar, B.M.; Ali, B.H.; Adeghate, E.; Nemmar, A. The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice. Biomedicines 2022, 10, 842. https://doi.org/10.3390/biomedicines10040842
Zaaba NE, Beegam S, Elzaki O, Yasin J, Nemmar BM, Ali BH, Adeghate E, Nemmar A. The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice. Biomedicines. 2022; 10(4):842. https://doi.org/10.3390/biomedicines10040842
Chicago/Turabian StyleZaaba, Nur Elena, Sumaya Beegam, Ozaz Elzaki, Javed Yasin, Bilal Mohamed Nemmar, Badreldin H. Ali, Ernest Adeghate, and Abderrahim Nemmar. 2022. "The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice" Biomedicines 10, no. 4: 842. https://doi.org/10.3390/biomedicines10040842
APA StyleZaaba, N. E., Beegam, S., Elzaki, O., Yasin, J., Nemmar, B. M., Ali, B. H., Adeghate, E., & Nemmar, A. (2022). The Nephroprotective Effects of α-Bisabolol in Cisplatin-Induced Acute Kidney Injury in Mice. Biomedicines, 10(4), 842. https://doi.org/10.3390/biomedicines10040842