
LL-37 and Double-Stranded RNA Synergistically Upregulate Bronchial Epithelial TLR3 Involving Enhanced Import of Double-Stranded RNA and Downstream TLR3 Signaling


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Cell Culture
2.2. Real-Time RT-qPCR
2.3. Western Blot
2.4. Assessment of Cell Viability
2.5. Measurement of Poly I:C Import Using Fluorescence Imaging
2.6. Agents
2.7. Statistics
3. Results
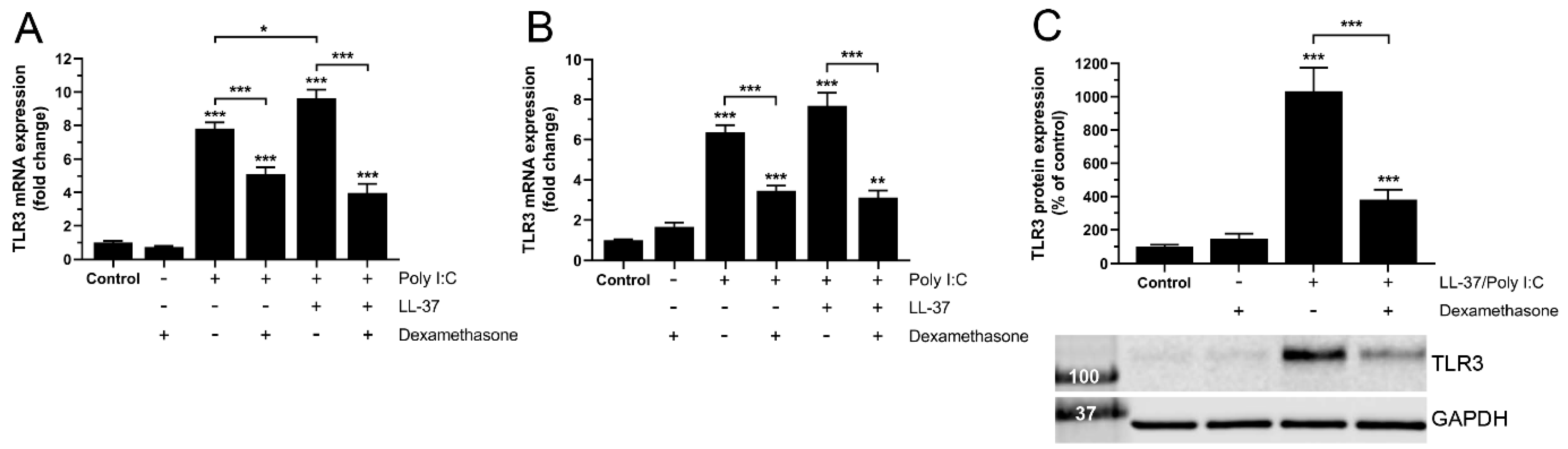
3.1. Dexamethasone Reduces TLR3 Expression Induced by Combined Treatment with LL-37 and Poly I:C in BEAS-2B Cells
3.2. Dexamethasone-Induced Down-Regulation of TLR3 Expression Is Associated with Reduced NF-κB Activity in BEAS-2B Cells
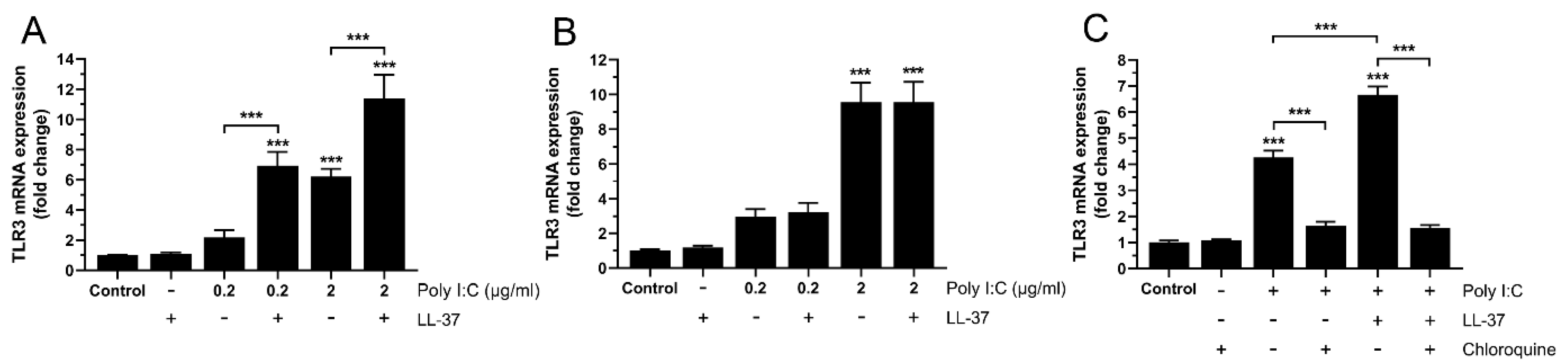
3.3. Upregulation of TLR3 Expression by Poly I:C and LL-37/poly I:C Involves Downstream TLR3 Signaling in BEAS-2B Cells
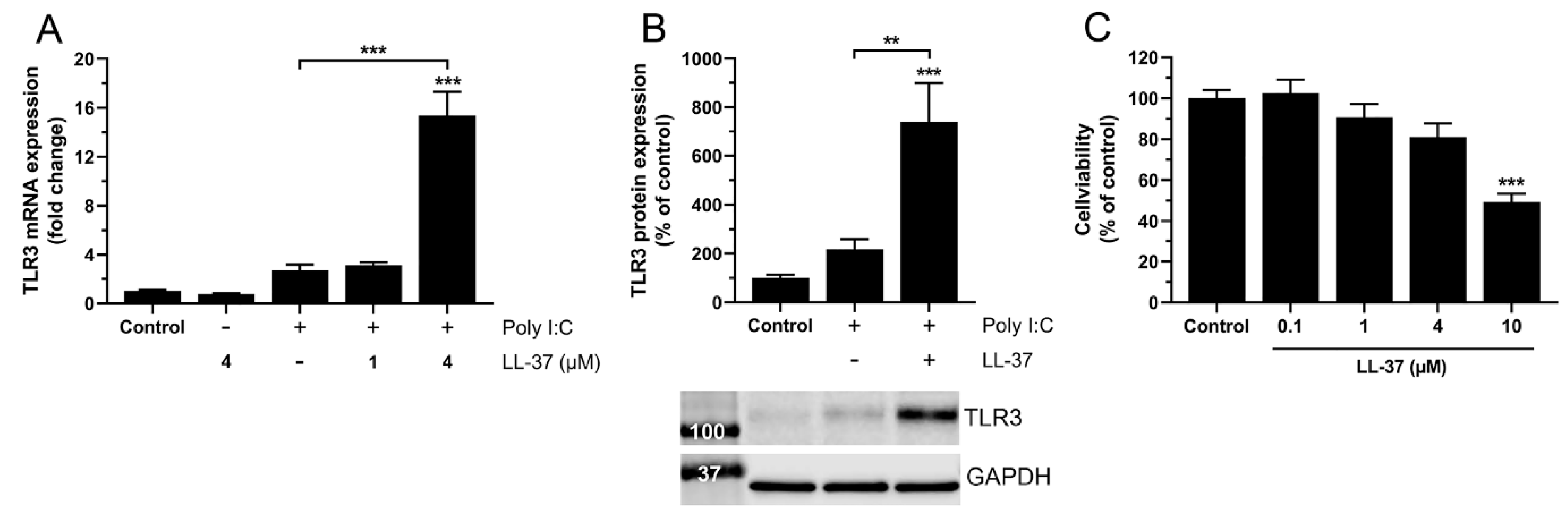
3.4. High but Not Low Concentrations of LL-37 Enhance Poly I:C-Induced TLR3 Expression at 24 h, Indicating Rapid Turnover of LL-37 in BEAS-2B Cells
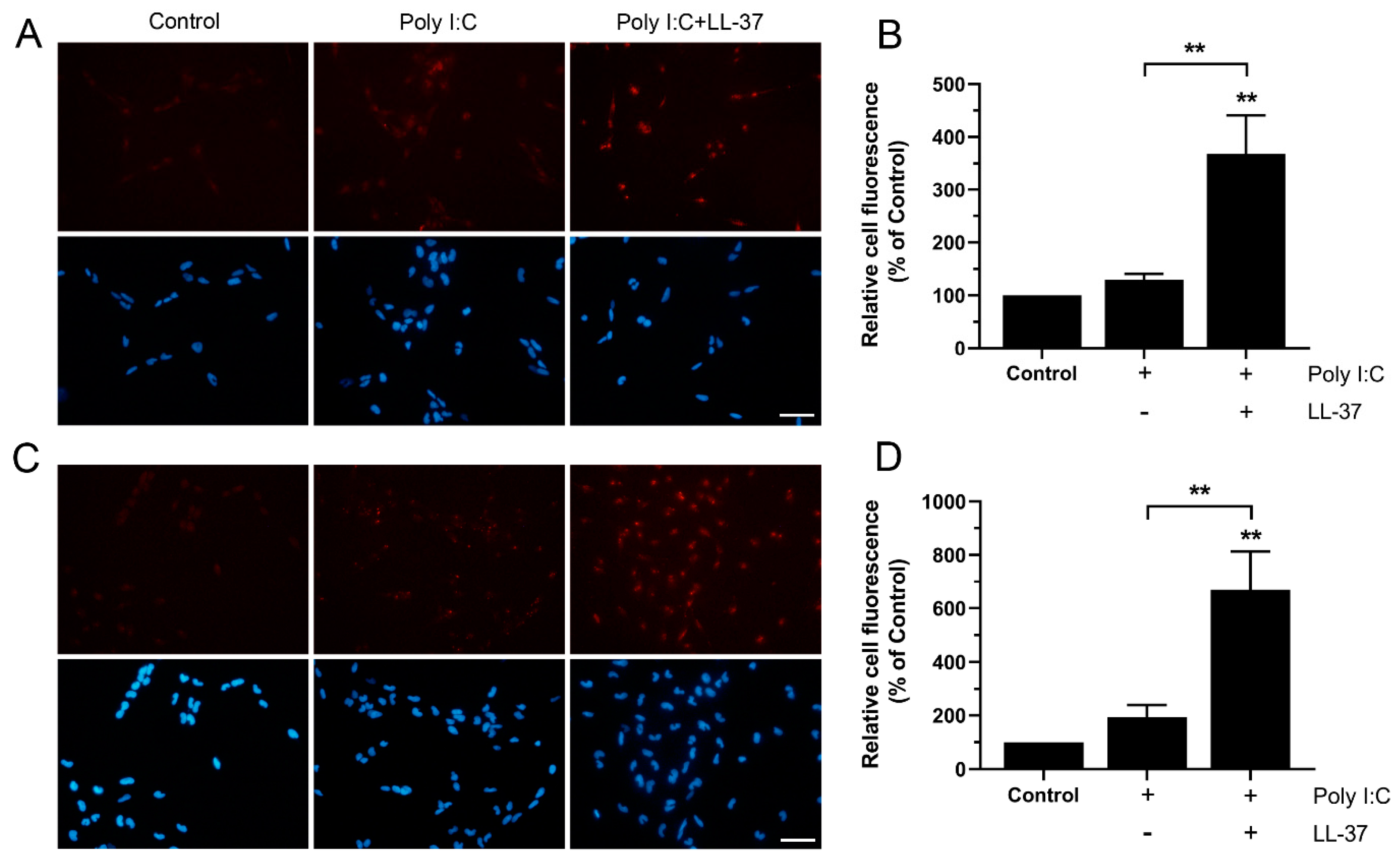
3.5. LL-37 Increases Import of Poly I:C in BEAS-2B Cells
4. Discussion
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Lancaster, G.I.; Khan, Q.; Drysdale, P.; Wallace, F.; Jeukendrup, A.E.; Drayson, M.T.; Gleeson, M. The physiological regulation of toll-like receptor expression and function in humans. J. Physiol. 2005, 563, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Kariko, K.; Ni, H.; Capodici, J.; Lamphier, M.; Weissman, D. mRNA is an endogenous ligand for toll-like receptor 3. J. Biol. Chem. 2003, 279, 12542–12550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawai, T.; Akira, S. Antiviral signaling through pattern recognition receptor. J. Biochem. 2007, 141, 137–145. [Google Scholar] [CrossRef]
- Alexopoulou, L.; Holt, A.C.; Medzhitov, R.; Flavell, R.A. Recognition of double-stranded RNA and activation of NF-κB by toll-like receptor 3. Nature 2001, 413, 732–738. [Google Scholar] [CrossRef]
- Oshimumi, H.; Matsumoto, M.; Funami, K.; Akazawa, T.; Seya, T. TICAM-1, an adaptor molecule that participates in toll-like receptor 3-mediated interferon-β induction. Nat. Immunol. 2003, 4, 161–167. [Google Scholar] [CrossRef]
- Mansour, S.C.; Pena, O.M.; Hancock, R.E. Host defense peptides: Front-line immunomodulators. Trends Immunol. 2014, 35, 443–450. [Google Scholar] [CrossRef]
- Sørensen, O.E.; Follin, P.; Johnsen, A.H.; Calafat, J.; Tjabringa, G.S.; Hiemstra, P.S.; Borregaard, N. Human cathelicidin, hCAP-18, is processed to the antimicrobial peptide LL-37 by extracellular cleavage with proteinase 3. Blood 2001, 97, 3951–3959. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, K.; Schauber, J.; Coda, A.; Lin, H.; Dorschner, R.A.; Schechter, N.M.; Bonnart, C.; Descargues, P.; Hovnanian, A.; Gallo, R.L. Kallikrein-mediated proteolysis regulates the antimicrobial effects of cathelicidins in skin. FASEB 2006, 20, 2068–2080. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Cho, Y.; Dinh, N.-N.; Waring, A.J.; Lehrer, R.I. Activities of LL-37, a cathelin-associated antimicrobial peptide of human neutrophils. Antimicrob Agents Chemother 1998, 42, 2206–2214. [Google Scholar] [CrossRef] [Green Version]
- Xhindoli, D.; Pacor, S.; Benincasa, M.; Scocchi, M.; Gennaro, R.; Tossi, A. The human cathelicidin LL-37—A pore-forming antibacterial peptide and host-cell modulator. Biochim. Biophys. Acta (BBA) 2016, 1858, 546–566. [Google Scholar] [CrossRef] [PubMed]
- Larrick, J.W.; Hirata, M.; Balint, R.F.; Lee, J.; Zhong, J.; Wright, S.C. Human CAP18: A novel antimicrobial lipopolysaccharide-binding protein. Infect. Immun. 1995, 63, 1291–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agier, J.; Efenberger, M.; Brzezinska-Blaszczyk, E. Cathelicidin impact on inflammatory cells. Cent. Eur. J. Immunol. 2015, 40, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Cederlund, A.; Gudmundsson, G.H.; Agerberth, B. Antimicrobial peptides important in innate immunity. FEBS J. 2011, 278, 3942–3951. [Google Scholar] [CrossRef] [PubMed]
- Nijnik, A.; Pistolic, J.; Filewood, N.C.J.; Hancock, R.E.W. Signaling pathways mediating chemokine induction in keratinocytes by cathelicidin LL-37 and flagellin. J. Innate Immun. 2012, 4, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chen, Q.; Schmidt, A.P.; Anderson, G.M.; Wang, J.M.; Wooters, J.; Oppenheim, J.J.; Chertov, O. LL-37, the neutrophil granule– and epithelial cell–derived cathelicidin, utilizes formyl peptide receptor–like 1 (FPRL1) as a receptor to chemoattract human peripheral blood neutrophils, monocytes, and T cells. J. Exp. Med. 2000, 2000, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Türkoglu, O.; Emingil, G.; Eren, G.; Atmaca, H.; Kutukculer, N.; Atilla, G. Gingival crevicular fluid and serum hCAP18/LL-37 levels in generalized aggressive periodontitis. Clin. Oral Investig. 2017, 21, 763–769. [Google Scholar] [CrossRef]
- Morizane, S.; Gallo, R.L. Antimicrobial peptides in pathogenesis of psoriasis. J. Dermatol. 2012, 39, 225–230. [Google Scholar] [CrossRef]
- Liu, M.C.; Xiao, H.Q.; Brown, A.J.; Ritter, C.S.; Schroeder, J. Association of vitamin D and antimicrobial peptide production during late-phase allergic responses in the lung. Clin. Exp. Allergy 2012, 42, 383–391. [Google Scholar] [CrossRef]
- Bals, R.; Wang, X.; Zasloff, M.; Wilson, J.M. The peptide antibiotic LL-37/hCAP-18 is expressed in epithelia of the human lung where it has broad antimicrobial activity at the airway surface. Proc. Natl. Acad. Sci. USA 1998, 95, 9541–9546. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Zhang, J.; Yu, F.-S.X. Toll-like receptor 3 agonist poly(I:C)-induced antiviral response in human corneal epithelial cells. Immunology 2005, 117, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Adhikarakunnathu, S.; Bhardwaj, K.; Ranjith-Kumar, C.T.; Wen, Y.; Jordan, J.L.; Wu, L.H.; Dragnea, B.; Mateo, L.S.; Kao, C.C. LL37 and cationic peptides enhance TLR3 signaling by viral double-stranded RNAs. PLoS ONE 2011, 6, e26632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, D.; Vaughan, R.; Kao, C.C. LL-37 peptide enhancement of signal transduction by toll-like receptor 3 is regulated by pH. J. Biol. Chem. 2014, 289, 27614–27624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahl, S.; Cerps, S.; Rippe, C.; Swärd, K.; Uller, L.; Svensson, D.; Nilsson, B.O. Human host defense peptide LL-37 facilitates double-stranded RNA pro-inflammatory signaling through up-regulation of TLR3 expression in vascular smooth muscle cells. Inflamm. Res. 2020, 69, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, P.A. The role of endosomes in innate and adaptive immunity. Semin. Cell Dev. Biol. 2014, 31, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Svensson, D.; Wilk, L.; Morgelin, M.; Herwald, H.; Nilsson, B.O. LL-37-induced host cell cytotoxicity depends on cellular expression of the globular C1q receptor (p33). Biochem. J. 2016, 4731, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Basavarajappa, D.; Haeggström, J.Z.; Wan, M. P2X7 receptor regulates internalization of antimicrobial peptide LL-37 by human macrophages that promotes intracellular pathogen clearance. J. Immunol. 2015, 195, 1191–1201. [Google Scholar] [CrossRef] [Green Version]
- Anders, E.; Dahl, S.; Svensson, D.; Nilsson, B.O. LL-37-induced human osteoblast cytotoxicity and permeability occurs independently of cellular LL-37 uptake through clathrin-mediated endocytosis. BBRC 2018, 501, 280–285. [Google Scholar] [CrossRef]
- Burton, M.F.; Steel, P.G. The chemistry and biology of LL-37. Nat. Prod. Rep. 2009, 26, 1572–1584. [Google Scholar] [CrossRef]
- Aarbiou, J.; Tjabringa, G.S.; Verhoosel, R.M.; Ninaber, D.K.; White, S.R.; Peltenburg, L.T.C.; Rabe, K.F.; Hiemstra, P.S. Mechanisms of cell death induced by the neutrophil antimicrobial peptides a-defensins and LL-37. Inflamm. Res. 2006, 55, 119–127. [Google Scholar] [CrossRef]
- Clark, A.R. Anti-inflammatory functions of glucocorticoid-induced genes. Mol. Cell Endocrinol. 2007, 275, 79–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Dexamethasone in Hospitalized Patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Bortolasci, C.C.; Puri, B.K.; Marx, W.; O’Neil, A.; Athan, E.; Walder, K.; Berk, M.; Olive, L.; Carvalho, A.F.; et al. The cytokine storms of COVID-19, H1N1 influenza, CRS and MAS compared. Can one sized treatment fit all? Cytokine 2021, 144, 155593. [Google Scholar] [CrossRef] [PubMed]
- Attiq, A.; Yao, L.J.; Afzal, S.; Khan, M.A. The triumvirate of NF-κB, inflammation and cytokine storm in COVID-19. Int. Immunopharmacol. 2021, 101, 108255. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodahl, S.; Cerps, S.; Uller, L.; Nilsson, B.-O. LL-37 and Double-Stranded RNA Synergistically Upregulate Bronchial Epithelial TLR3 Involving Enhanced Import of Double-Stranded RNA and Downstream TLR3 Signaling. Biomedicines 2022, 10, 492. https://doi.org/10.3390/biomedicines10020492
Bodahl S, Cerps S, Uller L, Nilsson B-O. LL-37 and Double-Stranded RNA Synergistically Upregulate Bronchial Epithelial TLR3 Involving Enhanced Import of Double-Stranded RNA and Downstream TLR3 Signaling. Biomedicines. 2022; 10(2):492. https://doi.org/10.3390/biomedicines10020492
Chicago/Turabian StyleBodahl, Sara, Samuel Cerps, Lena Uller, and Bengt-Olof Nilsson. 2022. "LL-37 and Double-Stranded RNA Synergistically Upregulate Bronchial Epithelial TLR3 Involving Enhanced Import of Double-Stranded RNA and Downstream TLR3 Signaling" Biomedicines 10, no. 2: 492. https://doi.org/10.3390/biomedicines10020492
APA StyleBodahl, S., Cerps, S., Uller, L., & Nilsson, B.-O. (2022). LL-37 and Double-Stranded RNA Synergistically Upregulate Bronchial Epithelial TLR3 Involving Enhanced Import of Double-Stranded RNA and Downstream TLR3 Signaling. Biomedicines, 10(2), 492. https://doi.org/10.3390/biomedicines10020492