
New Tricks with Old Dogs: Computational Identification and Experimental Validation of New miRNA–mRNA Regulation in hiPSC-CMs

 ,
,
Abstract
:1. Introduction
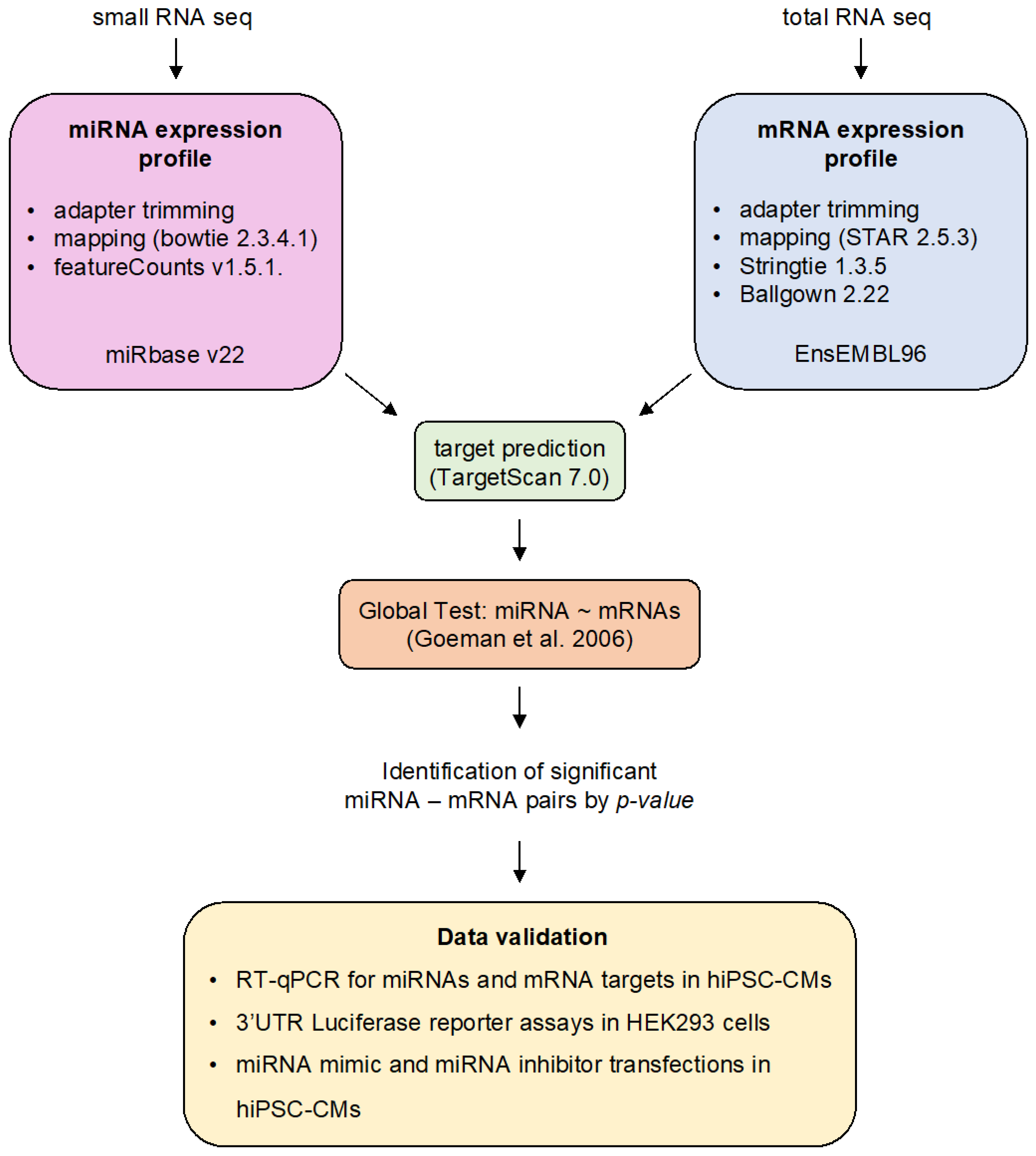
2. Materials and Methods
2.1. Human Induced Pluripotent Stem Cell-Derived Cardiomyocyte (hiPSC-CM) Small RNA and Gene Expression Data
2.2. Bioinformatic Analyses
2.2.1. Differential miRNA Expression Analysis
2.2.2. miRNA–mRNA Prediction Analysis
2.3. HiPSC Differentiation into Cardiomyocytes
2.4. RNA Extraction
2.5. Real Time Quantitative PCR (RT-qPCR)
2.6. Real Time Quantitative PCR for miRNA Expression Levels
2.7. miRNA Mimic and Inhibitor Transfections
2.8. Dual-Luciferase Reporter Assays
2.9. HiPSC-CM β-Adrenergic Stimulation
3. Results
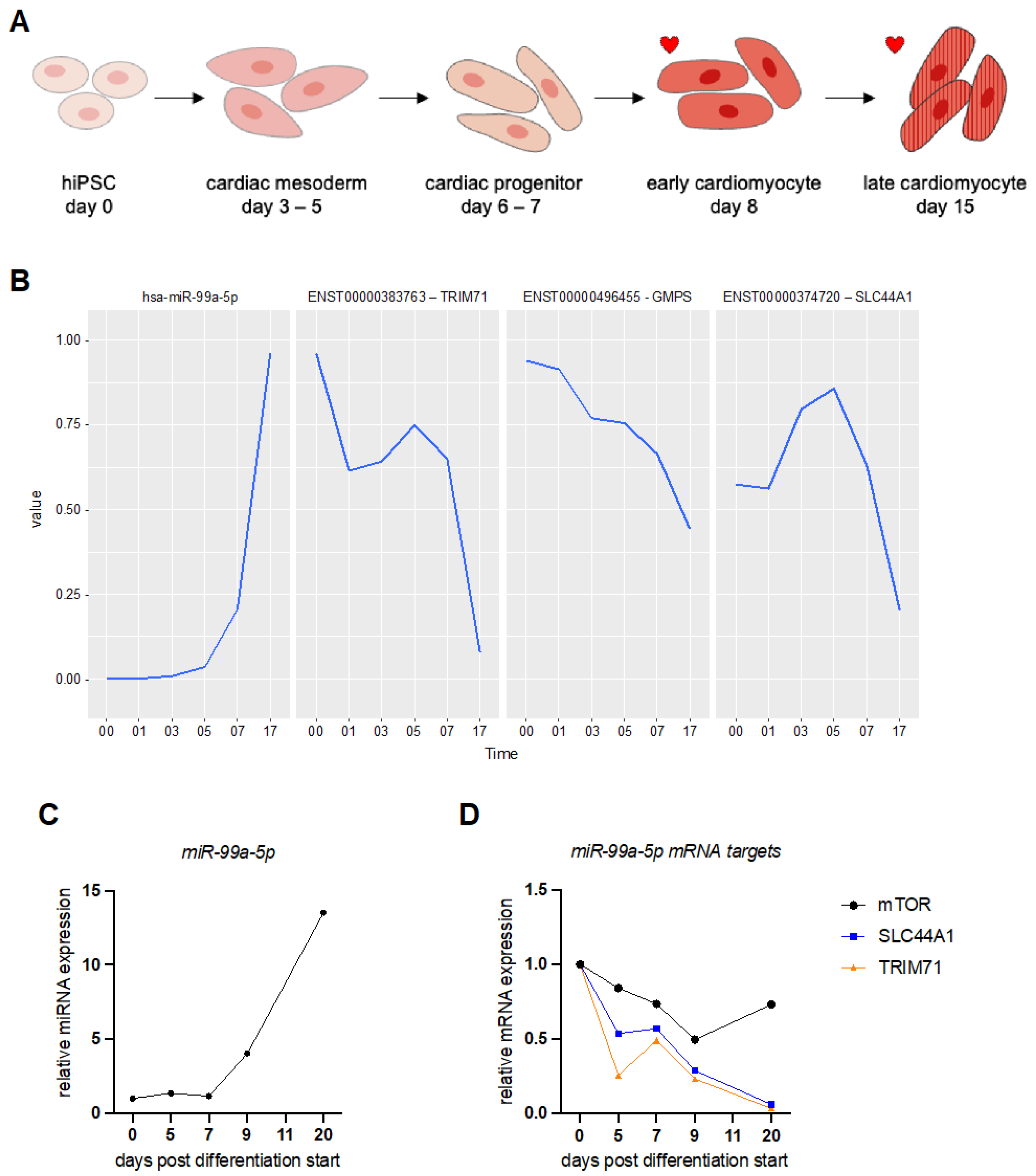
3.1. MicroRNA Expression Profiling during Human Induced Pluripotent Stem Cell (hiPSC) Differentiation into Cardiomyocytes
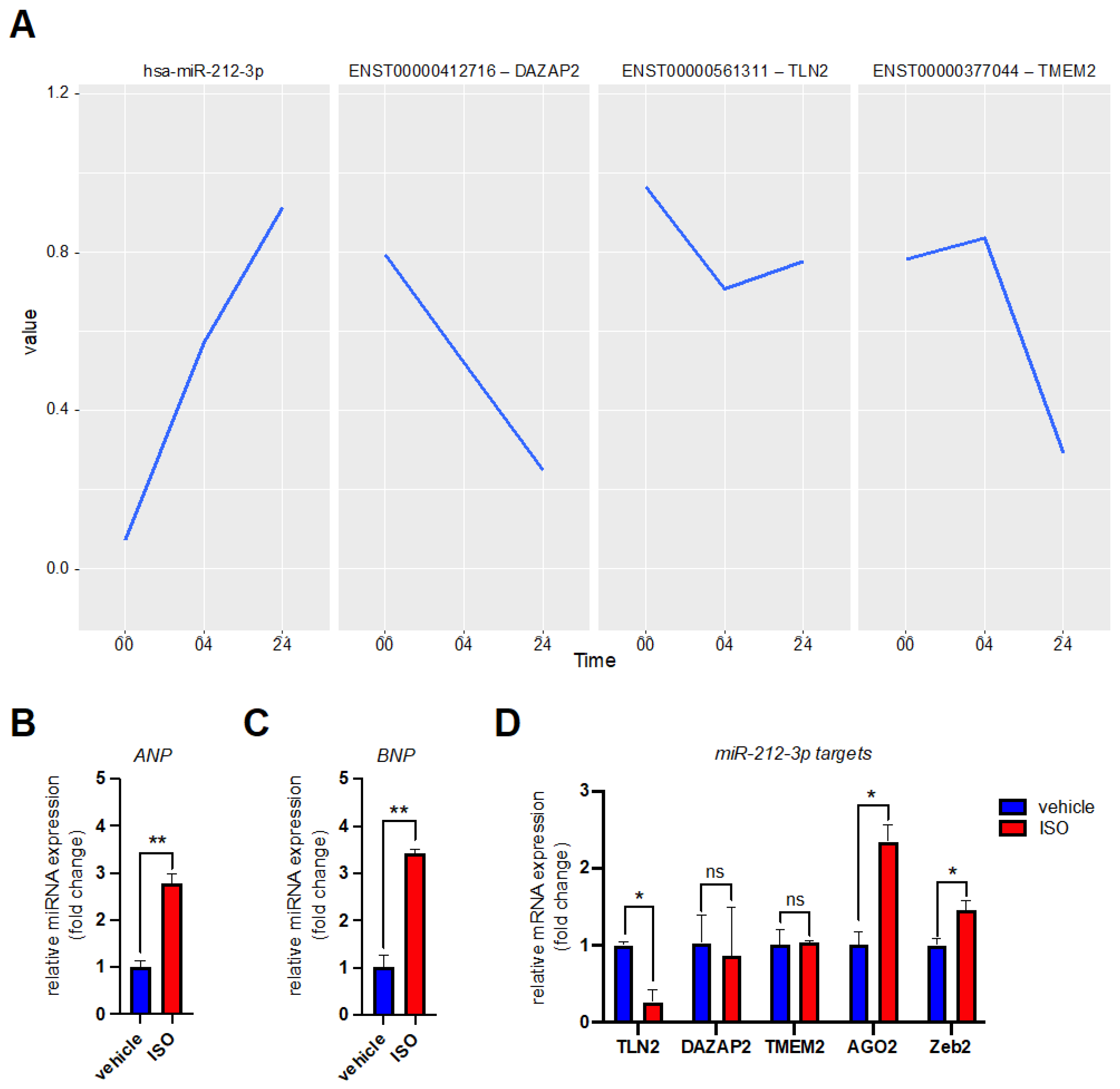
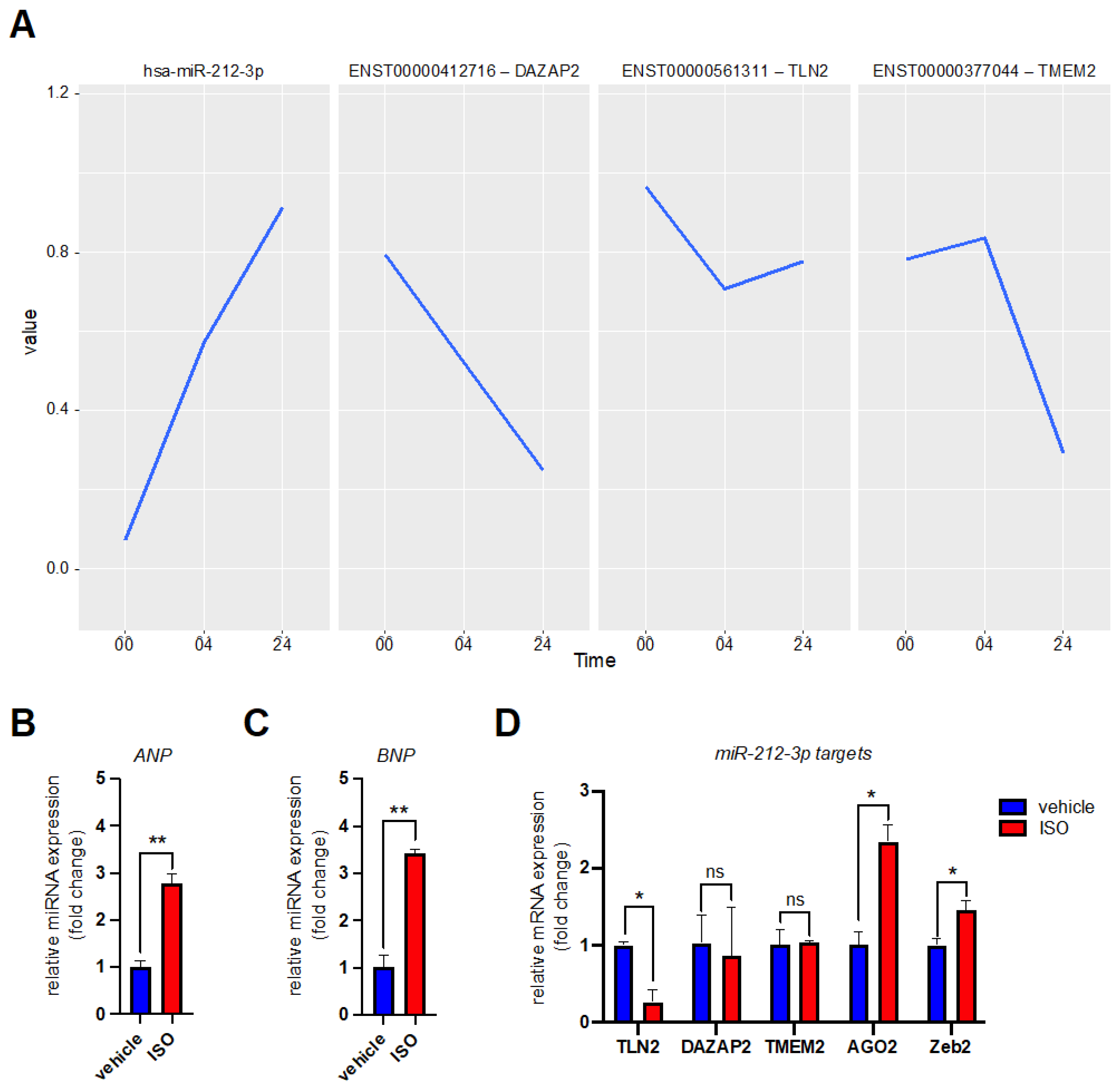
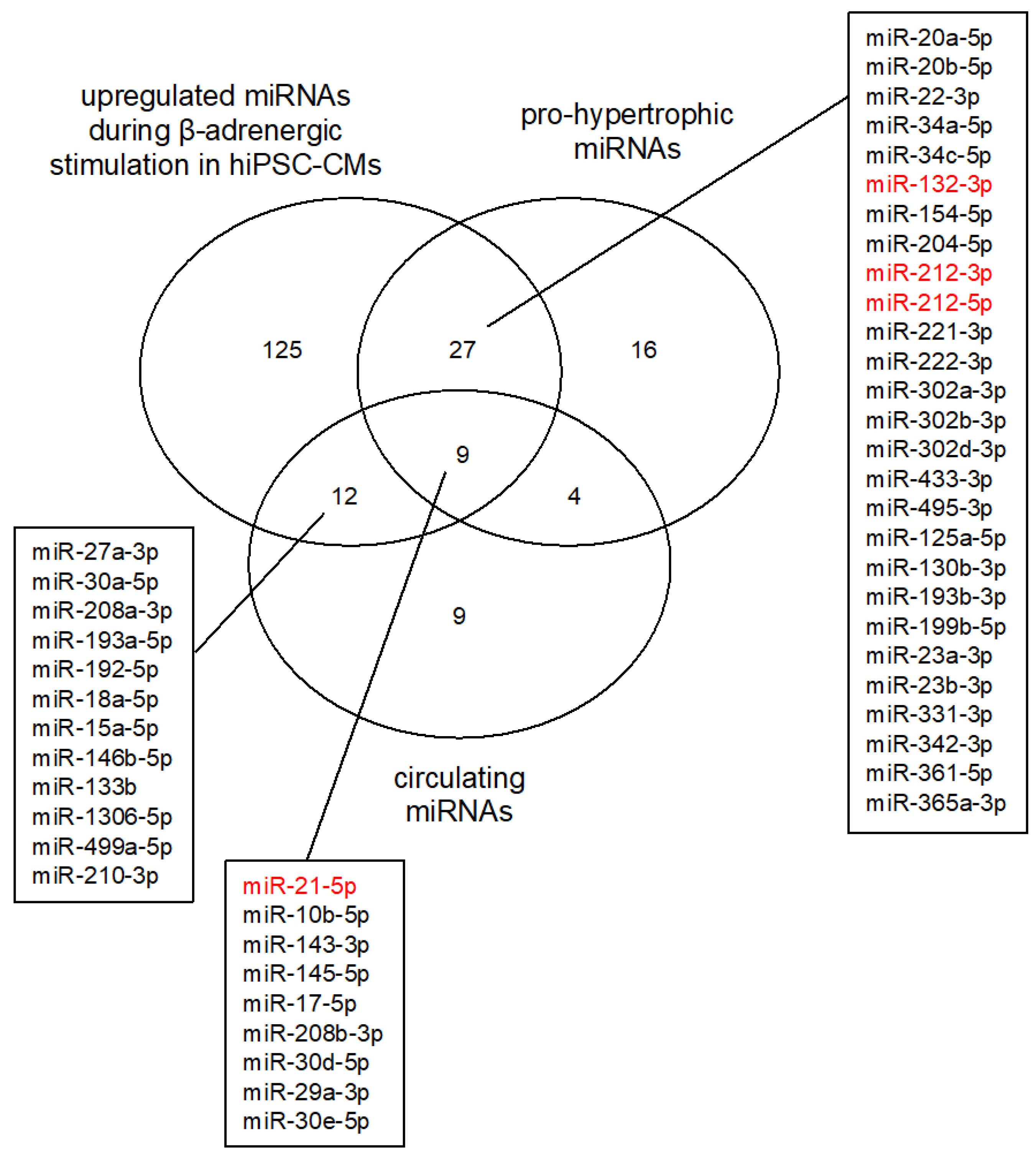
3.2. MicroRNA Expression Profiling after β-Adrenergic Stimulation
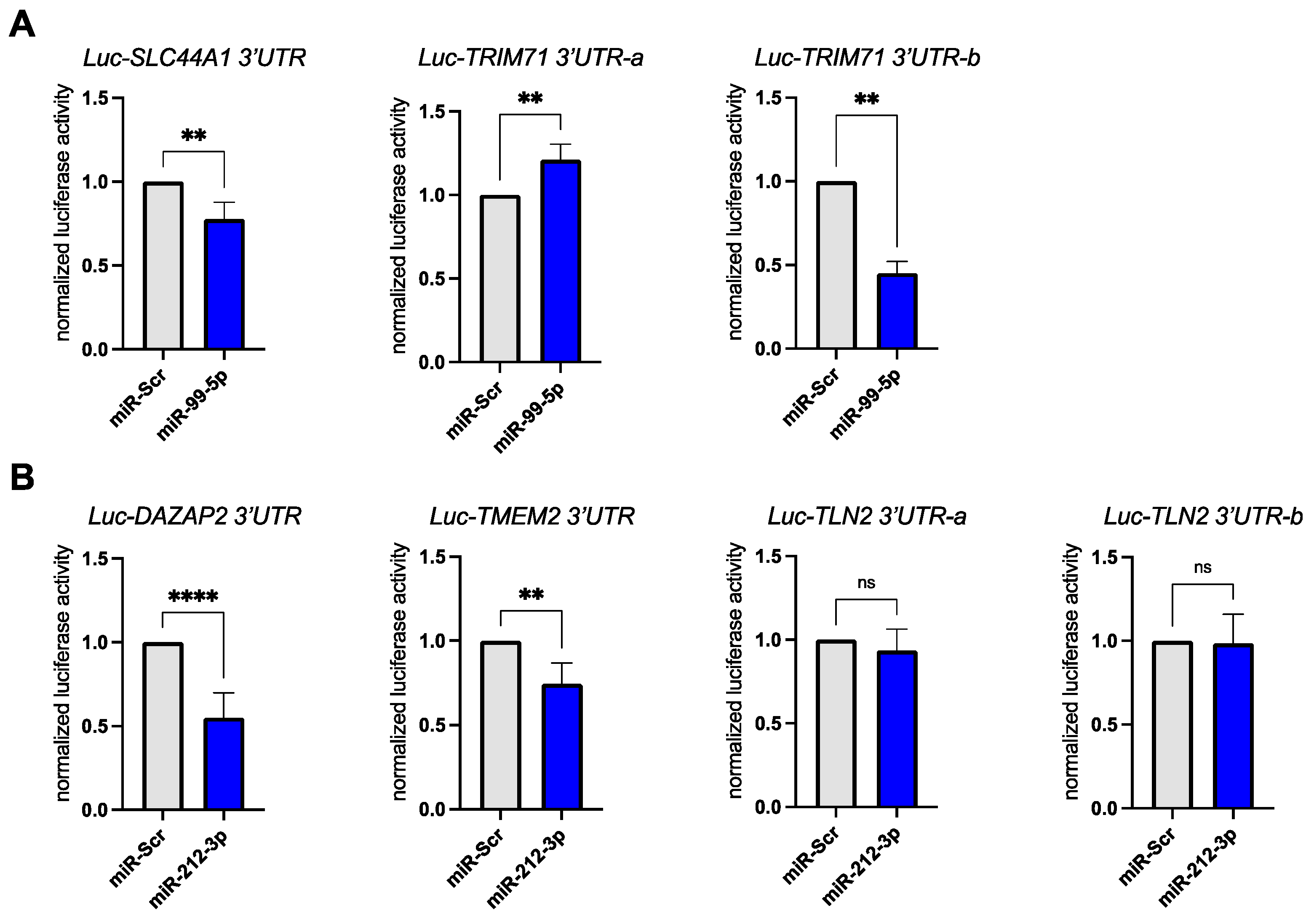
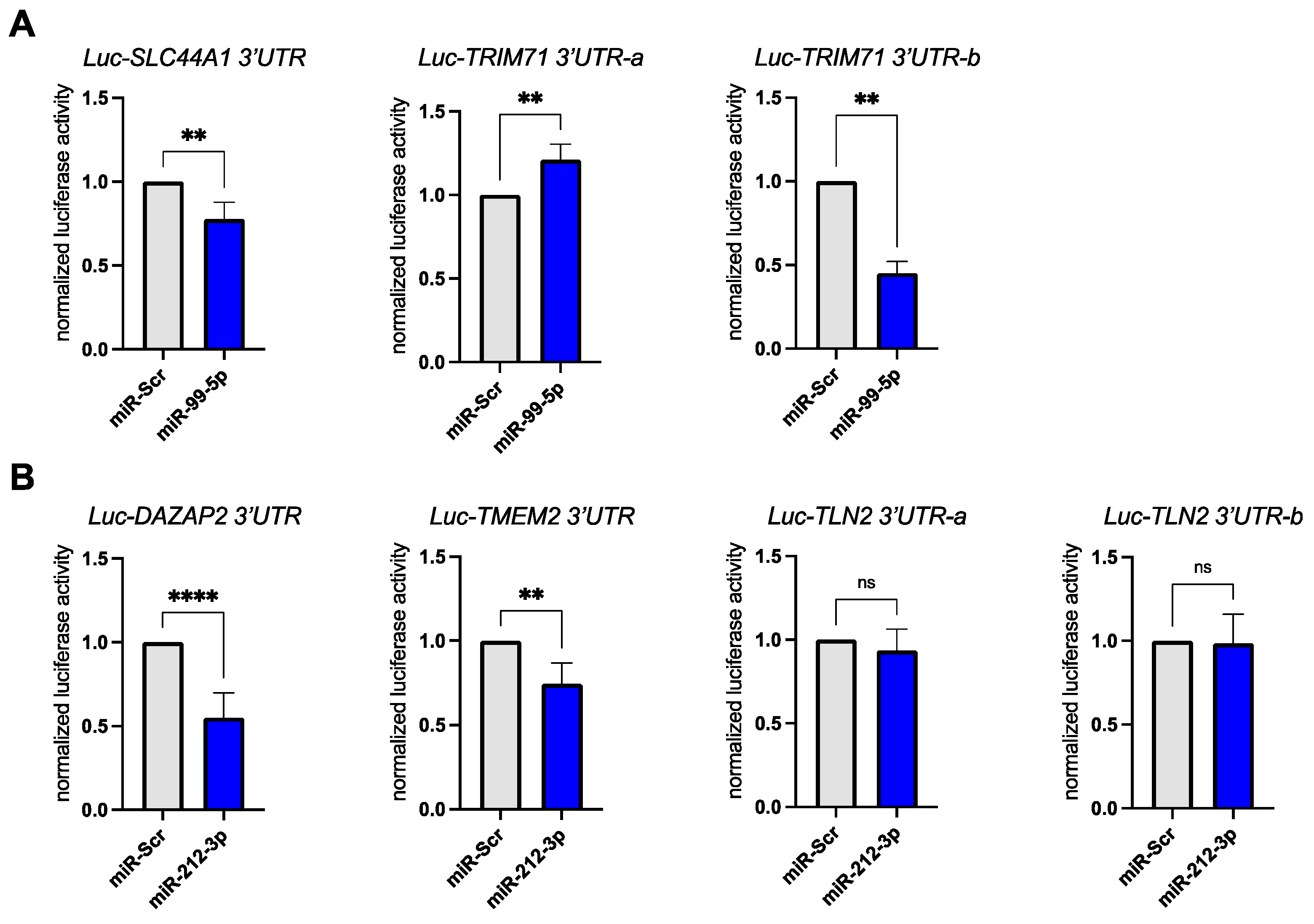
3.3. Validation of miR-99a-5p and miR-212-3p Target Genes in Reporter Assays
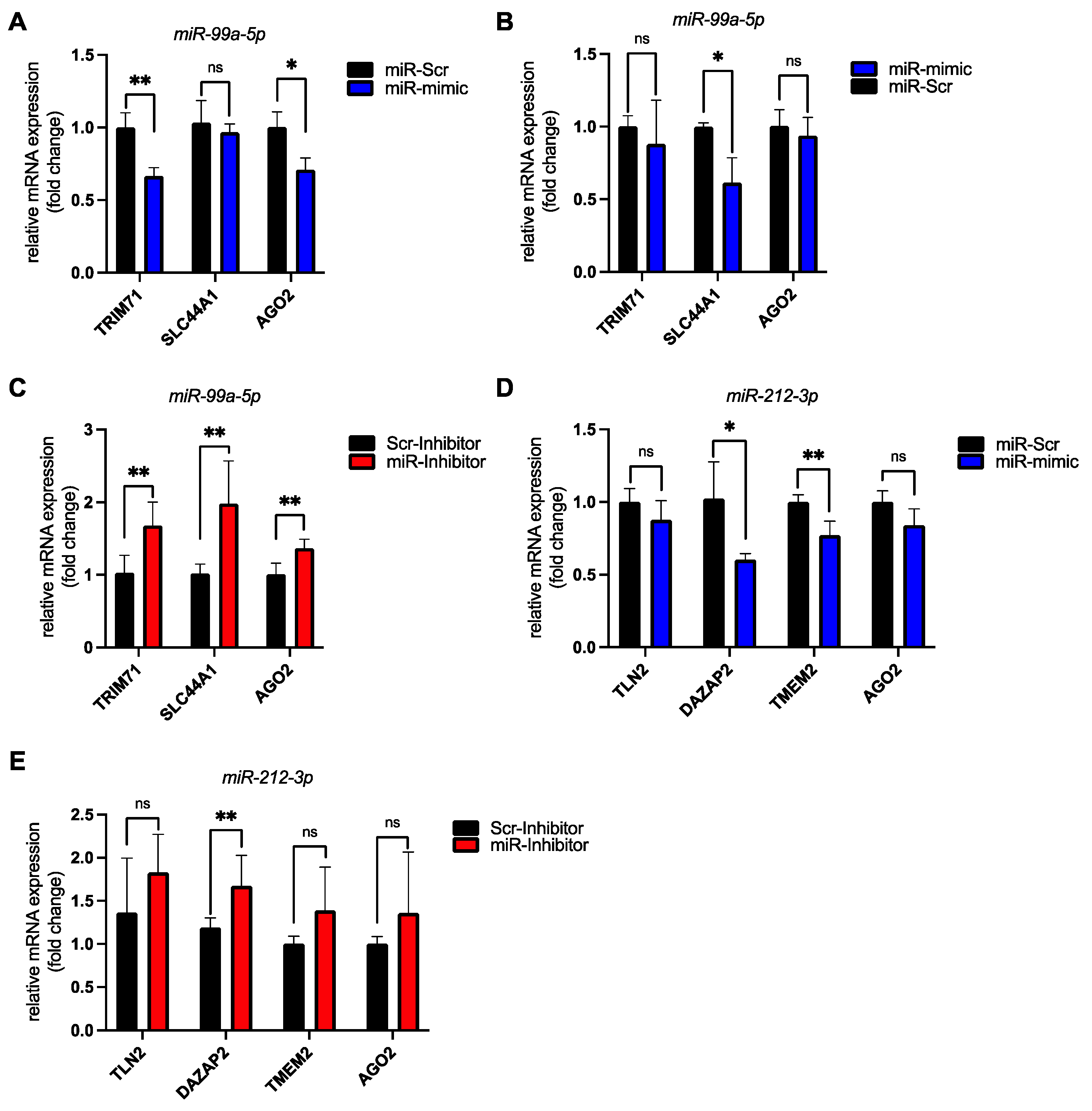
3.4. Validation of miR-99a-5p and miR-212-3p Target Genes in hiPSC-CMs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Johnson, C.O.; Addolorato, G.; Ammirati, E.; Baddour, L.M.; Barengo, N.C.; Beaton, A.Z.; Benjamin, E.J.; Benziger, C.P.; et al. Global burden of cardiovascular diseases and risk factors, 1990–2019: Update from the GBD 2019 study. J. Am. Coll. Cardiol. 2020, 76, 2982–3021. [Google Scholar] [CrossRef]
- Barry, S.P.; Townsend, P.A. What causes a broken heart—Molecular insights into heart failure. Int. Rev. Cell Mol. Biol. 2010, 284, 113–179. [Google Scholar] [CrossRef]
- Knight, W.E.; Cao, Y.; Lin, Y.-H.; Chi, C.; Bai, B.; Sparagna, G.C.; Zhao, Y.; Du, Y.; Londono, P.; Reisz, J.A.; et al. Maturation of pluripotent stem cell-derived cardiomyocytes enables modeling of human hypertrophic cardiomyopathy. Stem Cell Rep. 2021, 16, 519–533. [Google Scholar] [CrossRef]
- Musunuru, K.; Sheikh, F.; Gupta, R.M.; Houser, S.R.; Maher, K.O.; Milan, D.J.; Terzic, A.; Wu, J.C. Induced pluripotent stem cells for cardiovascular disease modeling and precision medicine: A scientific statement from the American heart association. Circ. Genom. Precis. Med. 2018, 11, e000043. [Google Scholar] [CrossRef] [Green Version]
- Gebert, L.F.R.; MacRae, I.J. Regulation of MicroRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Small, E.M.; Olson, E.N. Pervasive roles of MicroRNAs in cardiovascular biology. Nature 2011, 469, 336–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most mammalian MRNAs are conserved targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Pang, J.K.S.; Phua, Q.H.; Soh, B.-S. Applications of MiRNAs in cardiac development, disease progression and regeneration. Stem Cell Res. Ther. 2019, 10, 336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. Off. Organ Wroclaw Med. Univ. 2017, 26, 865–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lock, M.C.; Tellam, R.L.; Botting, K.J.; Wang, K.C.W.; Selvanayagam, J.B.; Brooks, D.A.; Seed, M.; Morrison, J.L. The role of MiRNA regulation in fetal cardiomyocytes, cardiac maturation and the risk of heart disease in adults. J. Physiol. 2018, 596, 5625–5640. [Google Scholar] [CrossRef]
- Zhou, S.; Jin, J.; Wang, J.; Zhang, Z.; Freedman, J.H.; Zheng, Y.; Cai, L. MiRNAS in cardiovascular diseases: Potential biomarkers, therapeutic targets and challenges. Acta Pharmacol. Sin. 2018, 39, 1073–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Rooij, E.; Sutherland, L.B.; Liu, N.; Williams, A.H.; McAnally, J.; Gerard, R.D.; Richardson, J.A.; Olson, E.N. A signature pattern of stress-responsive MicroRNAs that can evoke cardiac hypertrophy and heart failure. Proc. Natl. Acad. Sci. USA 2006, 103, 18255–18260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ucar, A.; Gupta, S.K.; Fiedler, J.; Erikci, E.; Kardasinski, M.; Batkai, S.; Dangwal, S.; Kumarswamy, R.; Bang, C.; Holzmann, A.; et al. The MiRNA-212/132 family regulates both cardiac hypertrophy and cardiomyocyte autophagy. Nat. Commun. 2012, 3, 1078. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.K.; Sahu, A.; Sun, Y.; Mohan, M.L.; Kumar, A.; Zalavadia, A.; Wang, X.; Martelli, E.E.; Stenson, K.; Witherow, C.P.; et al. Cardiac expression of MicroRNA-7 is associated with adverse cardiac remodeling. Sci. Rep. 2021, 11, 22018. [Google Scholar] [CrossRef] [PubMed]
- Hullinger, T.G.; Montgomery, R.L.; Seto, A.G.; Dickinson, B.A.; Semus, H.M.; Lynch, J.M.; Dalby, C.M.; Robinson, K.; Stack, C.; Latimer, P.A.; et al. Inhibition of MiR-15 Protects against Cardiac Ischemic Injury. Circ. Res. 2012, 110, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Van Rooij, E.; Sutherland, L.B.; Thatcher, J.E.; DiMaio, J.M.; Naseem, R.H.; Marshall, W.S.; Hill, J.A.; Olson, E.N. Dysregulation of MicroRNAs after Myocardial Infarction Reveals a Role of MiR-29 in Cardiac Fibrosis. Proc. Natl. Acad. Sci. USA 2008, 105, 13027–13032. [Google Scholar] [CrossRef] [Green Version]
- Van Rooij, E.; Sutherland, L.B.; Qi, X.; Richardson, J.A.; Hill, J.; Olson, E.N. Control of stress-dependent cardiac growth and gene expression by a MicroRNA. Science 2007, 316, 575–579. [Google Scholar] [CrossRef] [Green Version]
- Thum, T.; Gross, C.; Fiedler, J.; Fischer, T.; Kissler, S.; Bussen, M.; Galuppo, P.; Just, S.; Rottbauer, W.; Frantz, S.; et al. MicroRNA-21 Contributes to Myocardial Disease by Stimulating MAP Kinase Signalling in Fibroblasts. Nature 2008, 456, 980–984. [Google Scholar] [CrossRef]
- Carè, A.; Catalucci, D.; Felicetti, F.; Bonci, D.; Addario, A.; Gallo, P.; Bang, M.-L.; Segnalini, P.; Gu, Y.; Dalton, N.D.; et al. MicroRNA-133 Controls Cardiac Hypertrophy. Nat. Med. 2007, 13, 613–618. [Google Scholar] [CrossRef]
- Lin, Z.; Murtaza, I.; Wang, K.; Jiao, J.; Gao, J.; Li, P.-F. MiR-23a functions downstream of NFATc3 to regulate Cardiac Hypertrophy. Proc. Natl. Acad. Sci. USA 2009, 106, 12103–12108. [Google Scholar] [CrossRef] [Green Version]
- Siede, D.; Rapti, K.; Gorska, A.A.; Katus, H.A.; Altmüller, J.; Boeckel, J.N.; Meder, B.; Maack, C.; Völkers, M.; Müller, O.J.; et al. Identification of circular RNAs with host gene-independent expression in human model systems for cardiac differentiation and disease. J. Mol. Cell. Cardiol. 2017, 109, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Howe, K.L.; Achuthan, P.; Allen, J.; Alvarez-Jarreta, J.; Amode, M.R.; Armean, I.M.; Azov, A.G.; Bennett, R.; Bhai, J. Ensembl 2021. Nucleic Acids Res. 2021, 49, D884–D891. [Google Scholar] [CrossRef] [PubMed]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinforma. Oxf. Engl. 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, D.J.; Chen, Y.; Smyth, G.K. Differential expression analysis of multifactor RNA-Seq experiments with respect to biological variation. Nucleic Acids Res. 2012, 40, 4288–4297. [Google Scholar] [CrossRef] [Green Version]
- Goeman, J.J.; van de Geer, S.A.; de Kort, F.; van Houwelingen, H.C. A global test for groups of genes: Testing association with a clinical outcome. Bioinformatics 2004, 20, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Iterson, M.; Bervoets, S.; de Meijer, E.J.; Buermans, H.P.; ’t Hoen, P.A.C.; Menezes, R.X.; Boer, J.M. Integrated analysis of MicroRNA and MRNA expression: Adding biological significance to MicroRNA target predictions. Nucleic Acids Res. 2013, 41, e146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. stringtie enables improved reconstruction of a transcriptome from RNA-Seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-Seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective MicroRNA target sites in mammalian MRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, J.; Chen, Y.; Wang, C.; Wu, E.; Fu, L.; Xie, C. Quantification of distinct Let-7 MicroRNA family members by a modified stem-loop RT-QPCR. Mol. Med. Rep. 2018, 17, 3690–3696. [Google Scholar] [CrossRef] [Green Version]
- Winer, J.; Jung, C.K.S.; Shackel, I.; Williams, P.M. Development and validation of real-time quantitative reverse transcriptase–Polymerase chain reaction for monitoring gene expression in cardiac myocytesin vitro. Anal. Biochem. 1999, 270, 41–49. [Google Scholar] [CrossRef]
- Boon, R.A.; Dimmeler, S. MicroRNAs in Myocardial Infarction. Nat. Rev. Cardiol. 2015, 12, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Kalayinia, S.; Arjmand, F.; Maleki, M.; Malakootian, M.; Singh, C.P. MicroRNAs: Roles in cardiovascular development and disease. Cardiovasc. Pathol. Off. J. Soc. Cardiovasc. Pathol. 2021, 50, 107296. [Google Scholar] [CrossRef]
- Coppola, A.; Romito, A.; Borel, C.; Gehrig, C.; Gagnebin, M.; Falconnet, E.; Izzo, A.; Altucci, L.; Banfi, S.; Antonarakis, S.E.; et al. Cardiomyogenesis is controlled by the MiR-99a/Let-7c cluster and epigenetic modifications. Stem Cell Res. 2014, 12, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.-F.; Lin, J.-F.; Chou, K.-Y.; Lin, Y.-C.; Chen, H.-E.; Hwang, T.I.-S. MiR-99a-5p acts as tumor suppressor via targeting to MTOR and enhances RAD001-induced apoptosis in human urinary bladder urothelial carcinoma cells. OncoTargets Ther. 2018, 11, 239–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warth, S.C.; Hoefig, K.P.; Hiekel, A.; Schallenberg, S.; Jovanovic, K.; Klein, L.; Kretschmer, K.; Ansel, K.M.; Heissmeyer, V. Induced MiR-99a expression represses mtor cooperatively with MiR-150 to promote regulatory T-cell differentiation. EMBO J. 2015, 34, 1195–1213. [Google Scholar] [CrossRef] [Green Version]
- Hirt, M.N.; Werner, T.R.; Indenbirken, D.; Alawi, M.; Demin, P.; Kunze, A.-C.; Stenzig, J.; Starbatty, J.; Hansen, A.; Fiedler, J.; et al. Deciphering the MicroRNA Signature of Pathological Cardiac Hypertrophy by Engineered Heart Tissue- and Sequencing-Technology. J. Mol. Cell. Cardiol. 2015, 81, 1–9. [Google Scholar] [CrossRef]
- Leonov, G.; Shah, K.; Yee, D.; Timmis, J.; Sharp, T.V.; Lagos, D. Suppression of AGO2 by MiR-132 as a determinant of MiRNA-mediated silencing in human primary endothelial cells. Int. J. Biochem. Cell Biol. 2015, 69, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Cui, R.; Liu, Y. MicroRNA-212-3p Inhibits Paclitaxel Resistance through Regulating Epithelial-mesenchymal Transition, Migration and Invasion by Targeting ZEB2 in Human Hepatocellular Carcinoma. Oncol. Lett. 2020, 20, 23. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-M.; Martinez, N.J.; Thornton, J.E.; Hagan, J.P.; Nguyen, K.D.; Gregory, R.I. Trim71 Cooperates with MicroRNAs to Repress Cdkn1a Expression and Promote Embryonic Stem Cell Proliferation. Nat. Commun. 2012, 3, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worringer, K.A.; Rand, T.A.; Hayashi, Y.; Sami, S.; Takahashi, K.; Tanabe, K.; Narita, M.; Srivastava, D.; Yamanaka, S. The Let-7/LIN-41 Pathway Regulates Reprogramming to Human Induced Pluripotent Stem Cells by Controlling Expression of Prodifferentiation Genes. Cell Stem Cell 2014, 14, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Torres Fernández, L.A.; Mitschka, S.; Ulas, T.; Weise, S.; Dahm, K.; Becker, M.; Händler, K.; Beyer, M.; Windhausen, J.; Schultze, J.L.; et al. The stem cell-specific protein TRIM71 inhibits maturation and activity of the pro-differentiation MiRNA Let-7 via two independent molecular mechanisms. RNA 2021, 27, 805–828. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, A.; Montserrat, N.; Zacchigna, S.; Nivet, E.; Hishida, T.; Krause, M.N.; Kurian, L.; Ocampo, A.; Vazquez-Ferrer, E.; Rodriguez-Esteban, C.; et al. In vivo activation of a conserved MicroRNA program induces mammalian heart regeneration. Cell Stem Cell 2014, 15, 589–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiti, E.; Paolo, M.D.; Turillazzi, E.; Rocchi, A. MicroRNAs in Hypertrophic, Arrhythmogenic and Dilated Cardiomyopathy. Diagnostics 2021, 11, 1720. [Google Scholar] [CrossRef]
- Colpaert, R.M.W.; Calore, M. MicroRNAs in cardiac diseases. Cells 2019, 8, 737. [Google Scholar] [CrossRef] [Green Version]
- Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and intercellular transfer of MicroRNAs in living cells. J. Biol. Chem. 2010, 285, 17442–17452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caby, M.-P.; Lankar, D.; Vincendeau-Scherrer, C.; Raposo, G.; Bonnerot, C. Exosomal-like vesicles are present in human blood plasma. Int. Immunol. 2005, 17, 879–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanz-Rubio, D.; Martin-Burriel, I.; Gil, A.; Cubero, P.; Forner, M.; Khalyfa, A.; Marin, J.M. Stability of circulating exosomal MiRNAs in healthy subjects. Sci. Rep. 2018, 8, 10306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, S.; Wu, Y.; Yan, Y.; Shao, S.; Zhang, J.; Liu, J.; Hui, B.; Liu, R.; Ma, H.; Zhang, X.; et al. Construct a CircRNA/MiRNA/MRNA regulatory network to explore potential pathogenesis and therapy options of clear cell renal cell carcinoma. Sci. Rep. 2020, 10, 13659. [Google Scholar] [CrossRef]
- Chen, X.; Xie, D.; Zhao, Q.; You, Z.-H. MicroRNAs and complex diseases: From experimental results to computational models. Brief. Bioinform. 2019, 20, 515–539. [Google Scholar] [CrossRef] [PubMed]
- Krützfeldt, J.; Rajewsky, N.; Braich, R.; Rajeev, K.G.; Tuschl, T.; Manoharan, M.; Stoffel, M. Silencing of MicroRNAs in vivo with ‘Antagomirs’. Nature 2005, 438, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Roncarati, R.; Viviani Anselmi, C.; Losi, M.A.; Papa, L.; Cavarretta, E.; Da Costa Martins, P.; Contaldi, C.; Saccani Jotti, G.; Franzone, A.; Galastri, L.; et al. Circulating MiR-29a, among other up-regulated MicroRNAs, is the only biomarker for both hypertrophy and fibrosis in patients with hypertrophic cardiomyopathy. J. Am. Coll. Cardiol. 2014, 63, 920–927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, L.; Ellims, A.H.; Moore, X.; White, D.A.; Taylor, A.J.; Chin-Dusting, J.; Dart, A.M. Circulating MicroRNAs as biomarkers for diffuse myocardial fibrosis in patients with hypertrophic cardiomyopathy. J. Transl. Med. 2015, 13, 314. [Google Scholar] [CrossRef]
- Song, L.; Su, M.; Wang, S.; Zou, Y.; Wang, X.; Wang, Y.; Cui, H.; Zhao, P.; Hui, R.; Wang, J. MiR-451 is decreased in hypertrophic cardiomyopathy and regulates autophagy by targeting TSC1. J. Cell. Mol. Med. 2014, 18, 2266–2274. [Google Scholar] [CrossRef]
- Patrick, D.M.; Montgomery, R.L.; Qi, X.; Obad, S.; Kauppinen, S.; Hill, J.A.; van Rooij, E.; Olson, E.N. Stress-dependent cardiac remodeling occurs in the absence of MicroRNA-21 in mice. J. Clin. Investig. 2010, 120, 3912–3916. [Google Scholar] [CrossRef] [Green Version]
- Foinquinos, A.; Batkai, S.; Genschel, C.; Viereck, J.; Rump, S.; Gyöngyösi, M.; Traxler, D.; Riesenhuber, M.; Spannbauer, A.; Lukovic, D.; et al. Preclinical development of a MiR-132 inhibitor for heart failure treatment. Nat. Commun. 2020, 11, 633. [Google Scholar] [CrossRef]
- Täubel, J.; Hauke, W.; Rump, S.; Viereck, J.; Batkai, S.; Poetzsch, J.; Rode, L.; Weigt, H.; Genschel, C.; Lorch, U.; et al. Novel antisense therapy targeting MicroRNA-132 in patients with heart failure: Results of a first-in-human phase 1b randomized, double-blind, placebo-controlled study. Eur. Heart J. 2021, 42, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Gallant-Behm, C.L.; Piper, J.; Dickinson, B.A.; Dalby, C.M.; Pestano, L.A.; Jackson, A.L. A synthetic MicroRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. Off. Publ. Wound Heal. Soc. Eur. Tissue Repair Soc. 2018, 26, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Jopling, C.L.; Yi, M.; Lancaster, A.M.; Lemon, S.M.; Sarnow, P. Modulation of Hepatitis C Virus RNA Abundance by a Liver-Specific MicroRNA. Science 2005, 309, 1577–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | p-Value | Number of mRNA Targets |
|---|---|---|
| hsa-let-7c-5p | 4.97331 × 10−12 | 6 |
| hsa-miR-31-5p | 3.70845 × 10−10 | 221 |
| hsa-miR-489-3p | 7.58567 × 10−10 | 162 |
| hsa-miR-490-3p | 1.08922 × 10−7 | 129 |
| hsa-miR-27b-3p | 2.98507 × 10−7 | 64 |
| hsa-miR-181d-5p | 3.3065 × 10−7 | 156 |
| hsa-miR-362-5p | 4.79262 × 10−7 | 72 |
| hsa-miR-28-3p | 5.71315 × 10−7 | 74 |
| hsa-miR-34c-5p | 7.34698 × 10−7 | 132 |
| hsa-miR-23b-3p | 2.08809 × 10−6 | 37 |
| hsa-miR-146a-5p | 2.1465 × 10−6 | 55 |
| hsa-miR-99b-5p | 2.89621 × 10−6 | 5 |
| hsa-miR-142-5p | 3.16525 × 10−6 | 238 |
| hsa-miR-302d-3p | 4.01342 × 10−6 | 2 |
| hsa-miR-99a-5p | 5.67015 × 10−6 | 17 |
| hsa-miR-449a | 8.3154 × 10−6 | 26 |
| hsa-miR-30a-5p | 8.33399 × 10−6 | 197 |
| hsa-miR-181c-5p | 1.49425 × 10−5 | 88 |
| hsa-miR-335-5p | 2.00942 × 10−5 | 156 |
| hsa-miR-30b-5p | 2.33941 × 10−5 | 317 |
| mRNA Target | p-Value | z-Score |
|---|---|---|
| TRIM71 | 1.86 × 10−6 | 8.7854311 |
| GMPS | 1.432 × 10−5 | 7.967437 |
| SLC44A1 | 0.0002668 | 6.3925893 |
| RAVER2 | 0.0172793 | 3.0612766 |
| ETV3 | 0.04815 | 2.0275677 |
| miRNA | p-Value | Number of mRNA Targets |
|---|---|---|
| hsa-miR-191-5p | 0.0013 | 34 |
| hsa-miR-503-5p | 0.0014 | 219 |
| hsa-miR-140-5p | 0.0022 | 238 |
| hsa-miR-370-5p | 0.0033 | 143 |
| hsa-miR-296-3p | 0.0043 | 41 |
| hsa-miR-208b-3p | 0.0056 | 67 |
| hsa-miR-212-3p | 0.0076 | 92 |
| hsa-miR-539-3p | 0.0106 | 223 |
| hsa-miR-30b-5p | 0.0124 | 302 |
| hsa-let-7d-5p | 0.0147 | 198 |
| hsa-miR-365a-3p | 0.0155 | 179 |
| hsa-miR-208a-3p | 0.0159 | 59 |
| hsa-miR-1271-5p | 0.0165 | 367 |
| hsa-miR-34a-5p | 0.0167 | 87 |
| hsa-miR-21-5p | 0.0173 | 194 |
| hsa-miR-99b-5p | 0.019 | 6 |
| hsa-miR-145-5p | 0.0221 | 10 |
| hsa-miR-125a-5p | 0.0258 | 166 |
| hsa-miR-20a-5p | 0.0271 | 192 |
| hsa-miR-17-5p | 0.0331 | 128 |
| mRNA Target | p-Value | z-Score |
|---|---|---|
| DAZAP2 | 0.0021 | 3.8486 |
| TLN2 | 0.0338 | 2.253 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bencun, M.; Britto-Borges, T.; Eschenbach, J.; Dieterich, C. New Tricks with Old Dogs: Computational Identification and Experimental Validation of New miRNA–mRNA Regulation in hiPSC-CMs. Biomedicines 2022, 10, 391. https://doi.org/10.3390/biomedicines10020391
Bencun M, Britto-Borges T, Eschenbach J, Dieterich C. New Tricks with Old Dogs: Computational Identification and Experimental Validation of New miRNA–mRNA Regulation in hiPSC-CMs. Biomedicines. 2022; 10(2):391. https://doi.org/10.3390/biomedicines10020391
Chicago/Turabian StyleBencun, Maja, Thiago Britto-Borges, Jessica Eschenbach, and Christoph Dieterich. 2022. "New Tricks with Old Dogs: Computational Identification and Experimental Validation of New miRNA–mRNA Regulation in hiPSC-CMs" Biomedicines 10, no. 2: 391. https://doi.org/10.3390/biomedicines10020391
APA StyleBencun, M., Britto-Borges, T., Eschenbach, J., & Dieterich, C. (2022). New Tricks with Old Dogs: Computational Identification and Experimental Validation of New miRNA–mRNA Regulation in hiPSC-CMs. Biomedicines, 10(2), 391. https://doi.org/10.3390/biomedicines10020391