
Four Weeks of Probiotic Supplementation Alters the Metabolic Perturbations Induced by Marathon Running: Insight from Metabolomics

 , , and
, , and
Abstract
:1. Introduction
2. Results
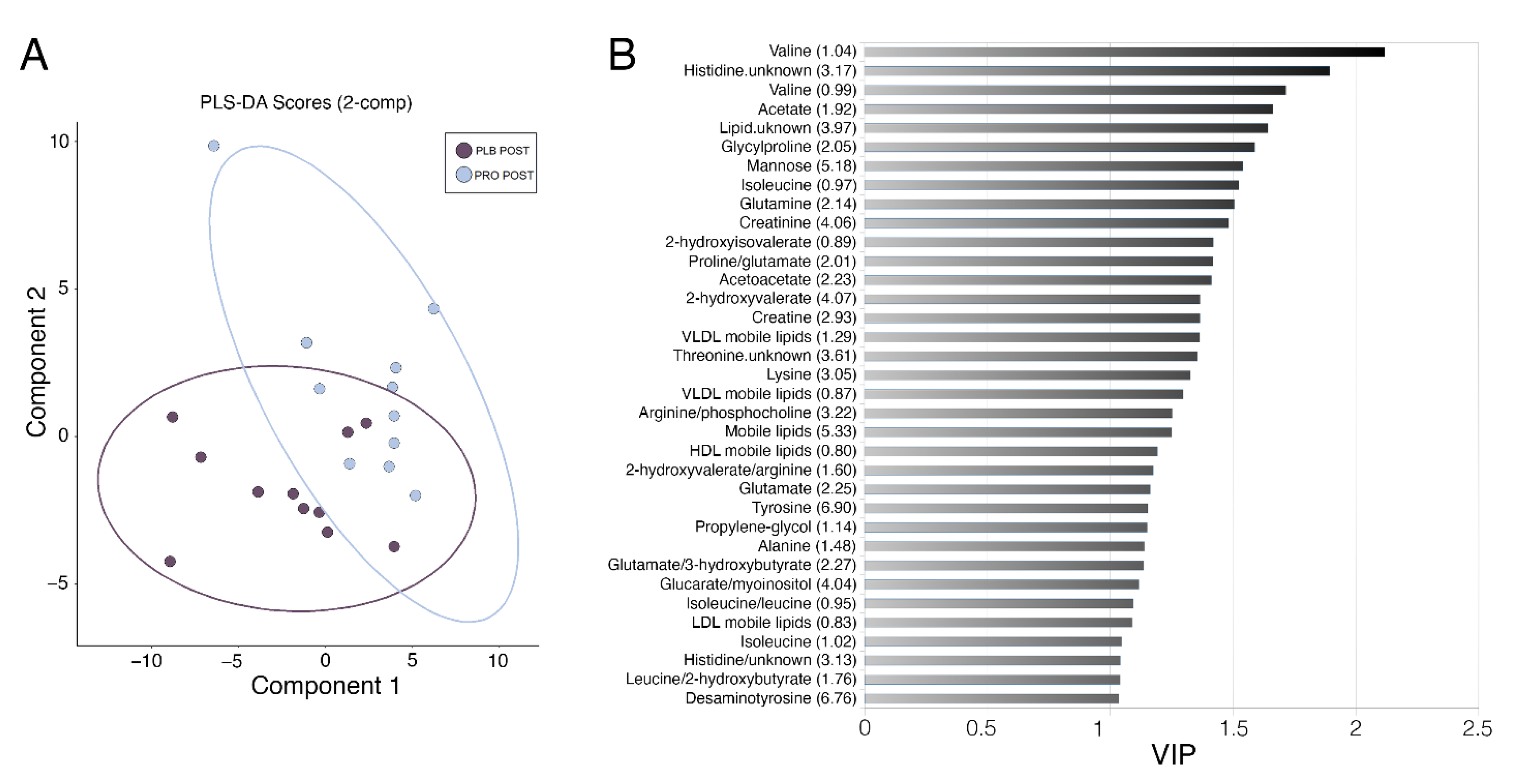
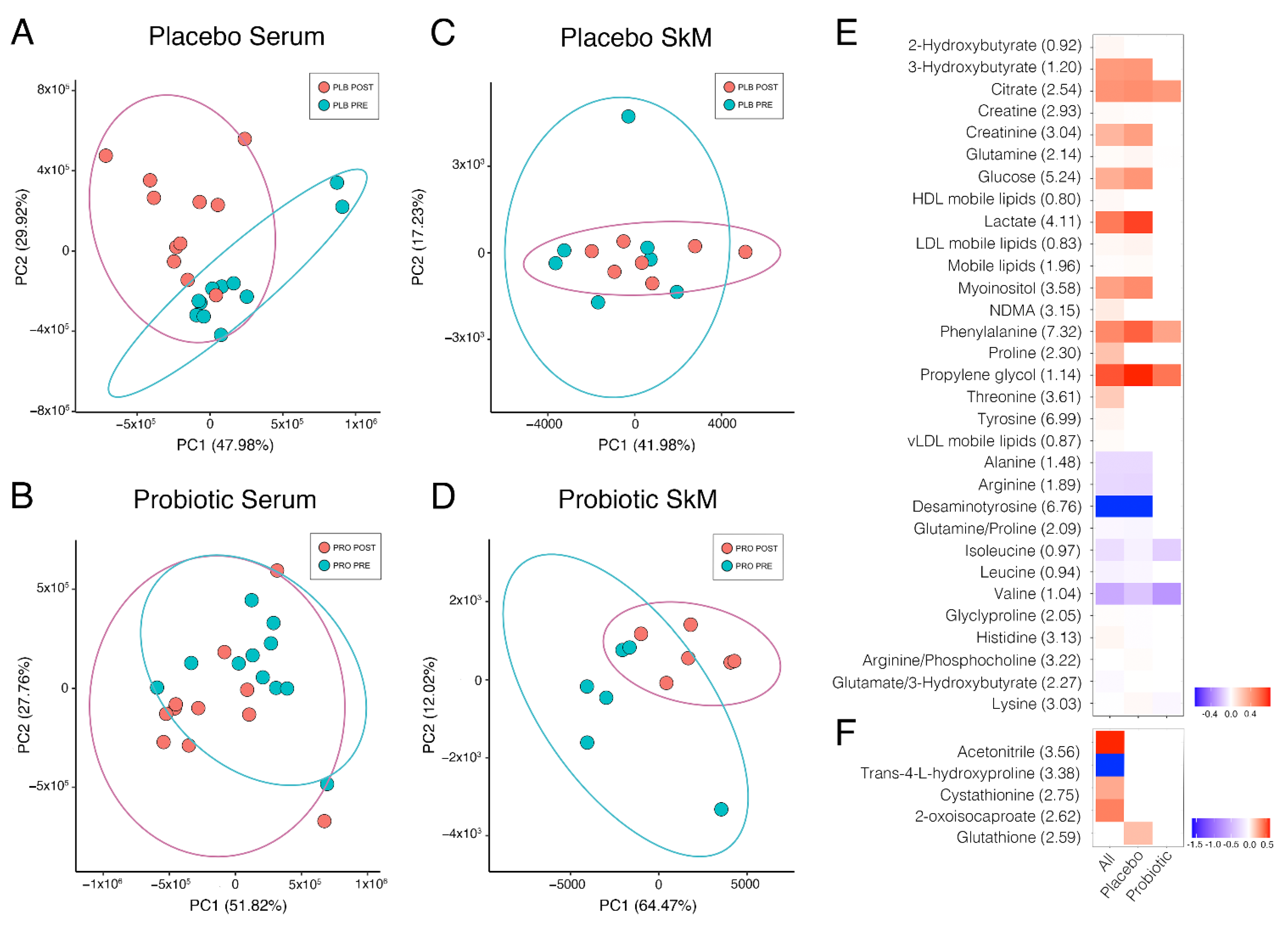
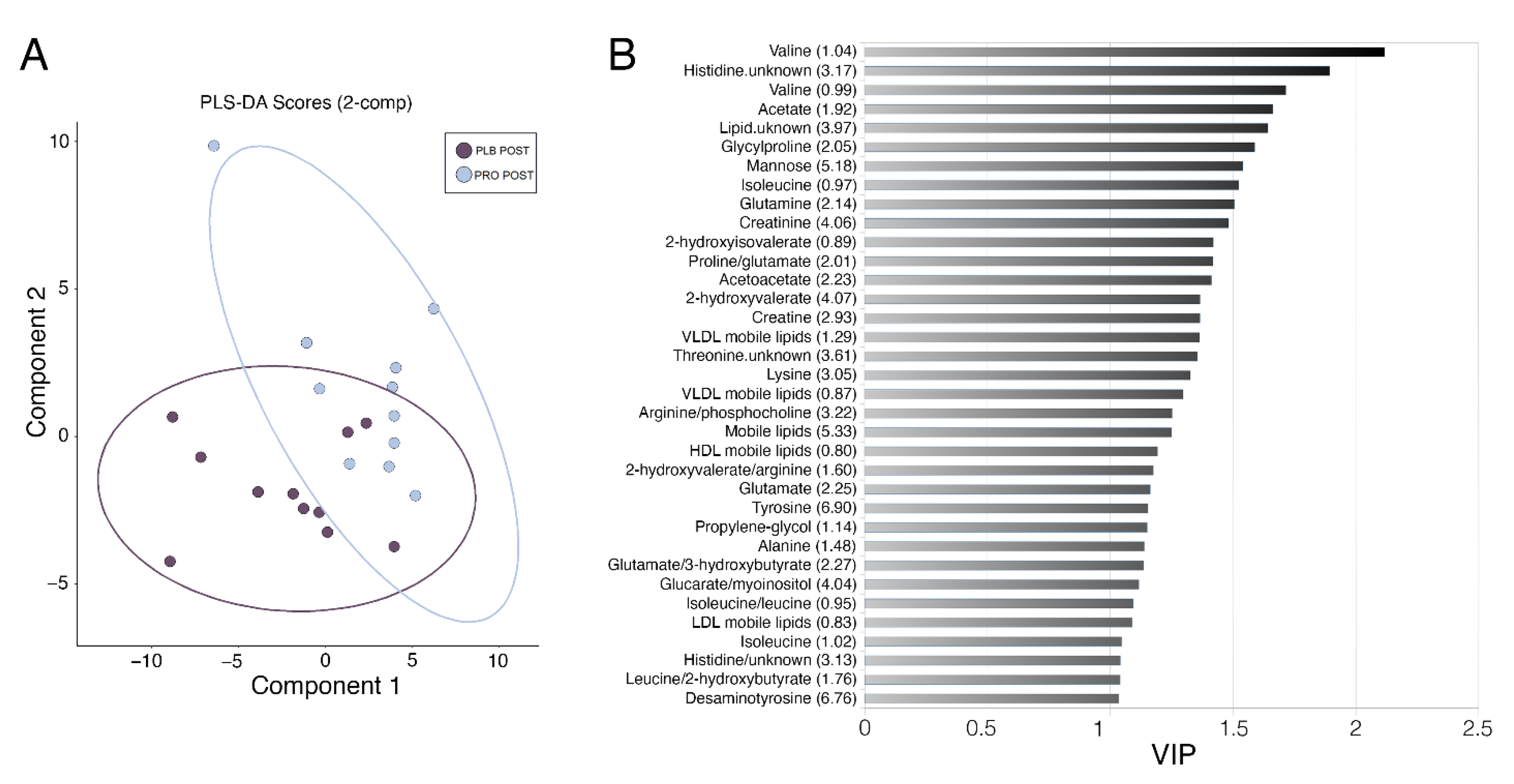
2.1. Effects of Multi-Strain Probiotic Supplementation on the Alterations to the Serum Metabolome Induced by Marathon Running
2.2. Effects of Marathon Running and Probiotic Supplementation on the Skeletal Muscle Metabolome
3. Discussion
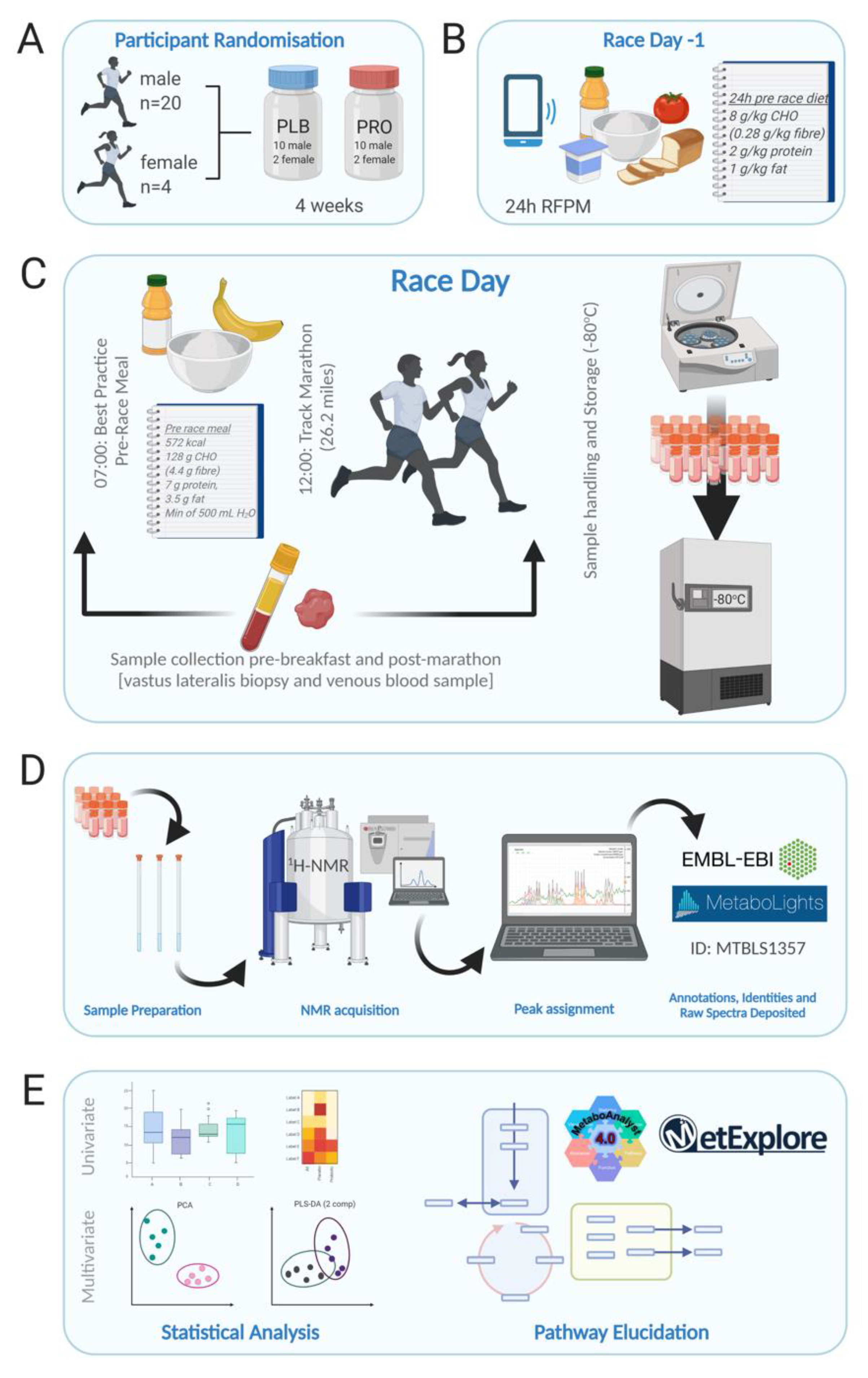
4. Materials and Methods
4.1. Participants
4.2. Experimental Design
4.3. Serum Sample Collection
4.4. Tissue Sample Collection and Extraction
4.5. Sample Preparation for NMR
4.6. NMR Set-Up and Acquisition
4.7. Spectra Processing and Quality Control
4.8. Metabolite Annotation and Identification
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pugh, J.N.; Kirk, B.; Fearn, R.; Morton, J.P.; Close, G.L. Prevalence, Severity and Potential Nutritional Causes of Gastrointestinal Symptoms during a Marathon in Recreational Runners. Nutrients 2018, 10, 811. [Google Scholar] [CrossRef] [Green Version]
- Keeffe, E.B.; Lowe, D.K.; Goss, J.R.; Wayne, R. Gastrointestinal symptoms of marathon runners. West J. Med. 1984, 141, 481–484. [Google Scholar] [PubMed]
- De Oliveira, E.P.; Burini, R.C. Carbohydrate-dependent, exercise-induced gastrointestinal distress. Nutrients 2014, 6, 4191–4199. [Google Scholar] [CrossRef] [Green Version]
- Van Wijck, K.; Lenaerts, K.; van Loon, L.J.; Peters, W.H.; Buurman, W.A.; Dejong, C.H. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 2011, 6, e22366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currell, K.; Jeukendrup, A.E. Superior endurance performance with ingestion of multiple transportable carbohydrates. Med. Sci. Sports Exerc. 2008, 40, 275–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coyle, E.F.; Hagberg, J.M.; Hurley, B.F.; Martin, W.H.; Ehsani, A.A.; Holloszy, J.O. Carbohydrate feeding during prolonged strenuous exercise can delay fatigue. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Valdes, A.M.; Walter, J.; Segal, E.; Spector, T.D. Role of the gut microbiota in nutrition and health. BMJ 2018, 361, k2179. [Google Scholar] [CrossRef] [Green Version]
- Wosinska, L.; Cotter, P.D.; O’Sullivan, O.; Guinane, C. The Potential Impact of Probiotics on the Gut Microbiome of Athletes. Nutrients 2019, 11, 2270. [Google Scholar] [CrossRef] [Green Version]
- Jager, R.; Mohr, A.E.; Carpenter, K.C.; Kerksick, C.M.; Purpura, M.; Moussa, A.; Townsend, J.R.; Lamprecht, M.; West, N.P.; Black, K.; et al. International Society of Sports Nutrition Position Stand: Probiotics. J. Int. Soc. Sports Nutr. 2019, 16, 62. [Google Scholar] [CrossRef] [Green Version]
- Pugh, J.N.; Sparks, A.S.; Doran, D.A.; Fleming, S.C.; Langan-Evans, C.; Kirk, B.; Fearn, R.; Morton, J.P.; Close, G.L. Four weeks of probiotic supplementation reduces GI symptoms during a marathon race. Eur. J. Appl. Physiol. 2019, 119, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kekkonen, R.A.; Vasankari, T.J.; Vuorimaa, T.; Haahtela, T.; Julkunen, I.; Korpela, R. The effect of probiotics on respiratory infections and gastrointestinal symptoms during training in marathon runners. Int. J. Sport Nutr. Exerc. Metab. 2007, 17, 352–363. [Google Scholar] [CrossRef]
- Roberts, J.D.; Suckling, C.A.; Peedle, G.Y.; Murphy, J.A.; Dawkins, T.G.; Roberts, M.G. An Exploratory Investigation of Endotoxin Levels in Novice Long Distance Triathletes, and the Effects of a Multi-Strain Probiotic/Prebiotic, Antioxidant Intervention. Nutrients 2016, 8, 733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikoff, W.R.; Anfora, A.T.; Liu, J.; Schultz, P.G.; Lesley, S.A.; Peters, E.C.; Siuzdak, G. Metabolomics analysis reveals large effects of gut microflora on mammalian blood metabolites. Proc. Natl. Acad. Sci. USA 2009, 106, 3698–3703. [Google Scholar] [CrossRef] [Green Version]
- Stander, Z.; Luies, L.; Mienie, L.J.; Keane, K.M.; Howatson, G.; Clifford, T.; Stevenson, E.J.; Loots, D.T. The altered human serum metabolome induced by a marathon. Metabolomics 2018, 14, 150. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.K.; Correa, J.B.; Han, H.; Allen, H.R.; Rood, J.C.; Champagne, C.M.; Gunturk, B.K.; Bray, G.A. Validity of the Remote Food Photography Method (RFPM) for estimating energy and nutrient intake in near real-time. Obesity 2012, 20, 891–899. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.K.; Samak, G. Protection and Restitution of Gut Barrier by Probiotics: Nutritional and Clinical Implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Y.; Gao, J.; Kou, Y.; Liu, M.; Meng, L.; Zheng, X.; Xu, S.; Liang, M.; Sun, H.; Liu, Z.; et al. The intestinal microbial metabolite desaminotyrosine is an anti-inflammatory molecule that modulates local and systemic immune homeostasis. FASEB J. 2020, 34, 16117–16128. [Google Scholar] [CrossRef]
- Palsson-McDermott, E.M.; O’Neill, L.A.J. Targeting immunometabolism as an anti-inflammatory strategy. Cell Res. 2020, 30, 300–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2016, 120, 692–701. [Google Scholar] [CrossRef]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [Green Version]
- Seth, A.; Yan, F.; Polk, D.B.; Rao, R.K. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier disruption by a PKC- and MAP kinase-dependent mechanism. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1060–G1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borsheim, E.; Cree, M.G.; Tipton, K.D.; Elliott, T.A.; Aarsland, A.; Wolfe, R.R. Effect of carbohydrate intake on net muscle protein synthesis during recovery from resistance exercise. J. Appl. Physiol. 2004, 96, 674–678. [Google Scholar] [CrossRef] [Green Version]
- Coyle, E.F.; Jeukendrup, A.E.; Wagenmakers, A.J.; Saris, W.H. Fatty acid oxidation is directly regulated by carbohydrate metabolism during exercise. Am. J. Physiol. 1997, 273, E268–E275. [Google Scholar] [CrossRef]
- Jeukendrup, A.E.; Moseley, L. Multiple transportable carbohydrates enhance gastric emptying and fluid delivery. Scand. J. Med. Sci. Sports 2010, 20, 112–121. [Google Scholar] [CrossRef]
- Pugh, J.N.; Wagenmakers, A.J.M.; Doran, D.A.; Fleming, S.C.; Fielding, B.A.; Morton, J.P.; Close, G.L. Probiotic supplementation increases carbohydrate metabolism in trained male cyclists: A randomized, double-blind, placebo-controlled crossover trial. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E504–E513. [Google Scholar] [CrossRef] [Green Version]
- Jang, C.; Chen, L.; Rabinowitz, J.D. Metabolomics and Isotope Tracing. Cell 2018, 173, 822–837. [Google Scholar] [CrossRef] [Green Version]
- Carraro, F.; Hartl, W.H.; Stuart, C.A.; Layman, D.K.; Jahoor, F.; Wolfe, R.R. Whole body and plasma protein synthesis in exercise and recovery in human subjects. Am. J. Physiol. 1990, 258, E821–E831. [Google Scholar] [CrossRef] [PubMed]
- Tipton, K.D.; Ferrando, A.A.; Williams, B.D.; Wolfe, R.R. Muscle protein metabolism in female swimmers after a combination of resistance and endurance exercise. J. Appl. Physiol. 1996, 81, 2034–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harber, M.P.; Konopka, A.R.; Douglass, M.D.; Minchev, K.; Kaminsky, L.A.; Trappe, T.A.; Trappe, S. Aerobic exercise training improves whole muscle and single myofiber size and function in older women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R1452–R1459. [Google Scholar] [CrossRef] [PubMed]
- Short, K.R.; Vittone, J.L.; Bigelow, M.L.; Proctor, D.N.; Nair, K.S. Age and aerobic exercise training effects on whole body and muscle protein metabolism. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E92–E101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pikosky, M.A.; Gaine, P.C.; Martin, W.F.; Grabarz, K.C.; Ferrando, A.A.; Wolfe, R.R.; Rodriguez, N.R. Aerobic exercise training increases skeletal muscle protein turnover in healthy adults at rest. J. Nutr. 2006, 136, 379–383. [Google Scholar] [CrossRef] [Green Version]
- Weight, L.M.; Alexander, D.; Jacobs, P. Strenuous exercise: Analogous to the acute-phase response? Clin. Sci. 1991, 81, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Starkie, R.L.; Rolland, J.; Angus, D.J.; Anderson, M.J.; Febbraio, M.A. Circulating monocytes are not the source of elevations in plasma IL-6 and TNF-alpha levels after prolonged running. Am. J. Physiol. Cell Physiol. 2001, 280, C769–C774. [Google Scholar] [CrossRef]
- Kratz, A.; Lewandrowski, K.B.; Siegel, A.J.; Chun, K.Y.; Flood, J.G.; Van Cott, E.M.; Lee-Lewandrowski, E. Effect of marathon running on hematologic and biochemical laboratory parameters, including cardiac markers. Am. J. Clin. Pathol. 2002, 118, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.E.; Garbutt, G.; Lopes, P.; Tunstall Pedoe, D. Effects of prolonged strenuous exercise (marathon running) on biochemical and haematological markers used in the investigation of patients in the emergency department. Br. J. Sports Med. 2004, 38, 292–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Peake, J.; Nosaka, K.; Okutsu, M.; Abbiss, C.R.; Surriano, R.; Bishop, D.; Quod, M.J.; Lee, H.; Martin, D.T.; et al. Changes in markers of muscle damage, inflammation and HSP70 after an Ironman Triathlon race. Eur. J. Appl. Physiol. 2006, 98, 525–534. [Google Scholar] [CrossRef]
- Kato, H.; Suzuki, K.; Bannai, M.; Moore, D.R. Protein Requirements Are Elevated in Endurance Athletes after Exercise as Determined by the Indicator Amino Acid Oxidation Method. PLoS ONE 2016, 11, e0157406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sen, C.K.; Atalay, M.; Hanninen, O. Exercise-induced oxidative stress: Glutathione supplementation and deficiency. J. Appl. Physiol. 1994, 77, 2177–2187. [Google Scholar] [CrossRef]
- Leeuwenburgh, C.; Hollander, J.; Leichtweis, S.; Griffiths, M.; Gore, M.; Ji, L.L. Adaptations of glutathione antioxidant system to endurance training are tissue and muscle fiber specific. Am. J. Physiol. 1997, 272, R363–R369. [Google Scholar] [CrossRef] [PubMed]
- Heshmati, J.; Farsi, F.; Shokri, F.; Rezaeinejad, M.; Almasi-Hashiani, A.; Vesali, S.; Sepidarkish, M. A systematic review and meta-analysis of the probiotics and synbiotics effects on oxidative stress. J. Funct. Foods 2018, 46, 66–84. [Google Scholar] [CrossRef]
- Clark, A.; Mach, N. The Crosstalk between the Gut Microbiota and Mitochondria during Exercise. Front. Physiol. 2017, 8, 319. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, J.T.; Lerin, C.; Haas, W.; Gygi, S.P.; Spiegelman, B.M.; Puigserver, P. Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature 2005, 434, 113–118. [Google Scholar] [CrossRef]
- Wishart, D.S. Advances in metabolite identification. Bioanalysis 2011, 3, 1769–1782. [Google Scholar] [CrossRef]
- Dona, A.C.; Jimenez, B.; Schafer, H.; Humpfer, E.; Spraul, M.; Lewis, M.R.; Pearce, J.T.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Precision high-throughput proton NMR spectroscopy of human urine, serum, and plasma for large-scale metabolic phenotyping. Anal. Chem. 2014, 86, 9887–9894. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, B.; Holmes, E.; Heude, C.; Tolson, R.F.; Harvey, N.; Lodge, S.L.; Chetwynd, A.J.; Cannet, C.; Fang, F.; Pearce, J.T.M.; et al. Quantitative Lipoprotein Subclass and Low Molecular Weight Metabolite Analysis in Human Serum and Plasma by (1)H NMR Spectroscopy in a Multilaboratory Trial. Anal. Chem. 2018, 90, 11962–11971. [Google Scholar] [CrossRef] [PubMed]
- Psychogios, N.; Hau, D.D.; Peng, J.; Guo, A.C.; Mandal, R.; Bouatra, S.; Sinelnikov, I.; Krishnamurthy, R.; Eisner, R.; Gautam, B.; et al. The human serum metabolome. PLoS ONE 2011, 6, e16957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viant, M.R.; Kurland, I.J.; Jones, M.R.; Dunn, W.B. How close are we to complete annotation of metabolomes? Curr. Opin. Chem. Biol. 2017, 36, 64–69. [Google Scholar] [CrossRef]
- Beckonert, O.; Keun, H.C.; Ebbels, T.M.; Bundy, J.; Holmes, E.; Lindon, J.C.; Nicholson, J.K. Metabolic profiling, metabolomic and metabonomic procedures for NMR spectroscopy of urine, plasma, serum and tissue extracts. Nat. Protoc. 2007, 2, 2692–2703. [Google Scholar] [CrossRef]
- Haug, K.; Salek, R.M.; Conesa, P.; Hastings, J.; de Matos, P.; Rijnbeek, M.; Mahendraker, T.; Williams, M.; Neumann, S.; Rocca-Serra, P.; et al. MetaboLights--an open-access general-purpose repository for metabolomics studies and associated meta-data. Nucleic Acids Res. 2013, 41, D781–D786. [Google Scholar] [CrossRef]
- Salek, R.M.; Steinbeck, C.; Viant, M.R.; Goodacre, R.; Dunn, W.B. The role of reporting standards for metabolite annotation and identification in metabolomic studies. Gigascience 2013, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kohl, S.M.; Klein, M.S.; Hochrein, J.; Oefner, P.J.; Spang, R.; Gronwald, W. State-of-the art data normalization methods improve NMR-based metabolomic analysis. Metabolomics 2012, 8, 146–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 21 November 2018).
- Le Cao, K.-A.; Rohart, F.; Gonzalez, I.; Dejean, S. mixOmics: Omics Data Integration Project, R package version 6.1.3; 2017.
- Xia, J.; Sinelnikov, I.V.; Han, B.; Wishart, D.S. MetaboAnalyst 3.0--making metabolomics more meaningful. Nucleic Acids Res. 2015, 43, W251–W257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cottret, L.; Frainay, C.; Chazalviel, M.; Cabanettes, F.; Gloaguen, Y.; Camenen, E.; Merlet, B.; Heux, S.; Portais, J.C.; Poupin, N.; et al. MetExplore: Collaborative edition and exploration of metabolic networks. Nucleic Acids Res. 2018, 46, W495–W502. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Biological Process | Total | Expected | Hits | Raw p-Value | Holm Adjusted p-Value | FDR |
|---|---|---|---|---|---|---|
| Aminoacyl-tRNA biosynthesis | 48 | 0.778 | 12 | 7.53 × 10−13 | 6.32 × 10−11 | 6.32 × 10−11 |
| Valine, leucine and isoleucine biosynthesis | 8 | 0.13 | 4 | 3.61 × 10−6 | 3.00 × 10−4 | 0.000152 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | 4 | 0.0649 | 2 | 0.00149 | 0.122 | 0.0416 |
| Phenylalanine metabolism | 10 | 0.162 | 2 | 0.0105 | 0.85 | 0.22 |
| Arginine biosynthesis | 14 | 0.227 | 2 | 0.0204 | 1 | 0.301 |
| Arginine and proline metabolism | 38 | 0.616 | 3 | 0.0218 | 1 | 0.301 |
| Valine, leucine and isoleucine degradation | 40 | 0.649 | 3 | 0.025 | 1 | 0.301 |
| Neomycin, kanamycin and gentamicin biosynthesis | 2 | 0.0324 | 1 | 0.0322 | 1 | 0.338 |
| Galactose metabolism | 27 | 0.438 | 2 | 0.0692 | 1 | 0.62 |
| Biological Process | Total | Expected | Hits | Raw p-Value | Holm Adjusted p-Value | FDR |
|---|---|---|---|---|---|---|
| Aminoacyl-tRNA biosynthesis | 48 | 0.725 | 10 | 3.10 × 10−10 | 2.61 × 10−8 | 2.61 × 10−8 |
| d-Glutamine and d-glutamate metabolism | 6 | 0.0907 | 4 | 5.85 × 10−7 | 4.85 × 10−5 | 2.46 × 10−5 |
| Valine, leucine and isoleucine biosynthesis | 8 | 0.121 | 3 | 0.000161 | 0.0132 | 0.00451 |
| Arginine biosynthesis | 14 | 0.212 | 3 | 0.000986 | 0.0798 | 0.0207 |
| Valine, leucine and isoleucine degradation | 40 | 0.604 | 4 | 0.00253 | 0.202 | 0.0425 |
| Nitrogen metabolism | 6 | 0.0907 | 2 | 0.00316 | 0.25 | 0.0442 |
| Ascorbate and aldarate metabolism | 8 | 0.121 | 2 | 0.00579 | 0.452 | 0.0695 |
| Alanine, aspartate and glutamate metabolism | 28 | 0.423 | 3 | 0.00772 | 0.595 | 0.0811 |
| Glyoxylate and dicarboxylate metabolism | 32 | 0.484 | 3 | 0.0112 | 0.854 | 0.105 |
| Arginine and proline metabolism | 38 | 0.574 | 3 | 0.018 | 1 | 0.151 |
| Butanoate metabolism | 15 | 0.227 | 2 | 0.0204 | 1 | 0.155 |
| Histidine metabolism | 16 | 0.242 | 2 | 0.0231 | 1 | 0.161 |
| PLB | PRO | |||
|---|---|---|---|---|
| Mean ± SD | Range | Mean ± SD | Range | |
| Age (years) Height (m) Body Mass (kg) VO2peak (mL·kg·min−1) LT (km·h−1) Most recent Marathon time (min) | 36.1 ± 7.5 1.75 ± 1.11 73.5 ± 11.3 56.4 ± 8.6 11.9 ± 1.9 220 ± 40 | 29–50 1.52–1.86 48–95 47.2–70.0 9–16.0 150–283 | 34.8 ± 6.9 1.79 ± 0.6 76.5 ± 9.4 57.6 ± 8.0 12.3 ± 1.9 222 ± 46 | 22–43 1.68–1.90 61–92 48.1–66.7 10–15.5 152–315 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pugh, J.N.; Phelan, M.M.; Caamaño-Gutiérrez, E.; Sparks, S.A.; Morton, J.P.; Close, G.L.; Owens, D.J. Four Weeks of Probiotic Supplementation Alters the Metabolic Perturbations Induced by Marathon Running: Insight from Metabolomics. Metabolites 2021, 11, 535. https://doi.org/10.3390/metabo11080535
Pugh JN, Phelan MM, Caamaño-Gutiérrez E, Sparks SA, Morton JP, Close GL, Owens DJ. Four Weeks of Probiotic Supplementation Alters the Metabolic Perturbations Induced by Marathon Running: Insight from Metabolomics. Metabolites. 2021; 11(8):535. https://doi.org/10.3390/metabo11080535
Chicago/Turabian StylePugh, Jamie N., Marie M. Phelan, Eva Caamaño-Gutiérrez, S. Andy Sparks, James P. Morton, Graeme L. Close, and Daniel J. Owens. 2021. "Four Weeks of Probiotic Supplementation Alters the Metabolic Perturbations Induced by Marathon Running: Insight from Metabolomics" Metabolites 11, no. 8: 535. https://doi.org/10.3390/metabo11080535
APA StylePugh, J. N., Phelan, M. M., Caamaño-Gutiérrez, E., Sparks, S. A., Morton, J. P., Close, G. L., & Owens, D. J. (2021). Four Weeks of Probiotic Supplementation Alters the Metabolic Perturbations Induced by Marathon Running: Insight from Metabolomics. Metabolites, 11(8), 535. https://doi.org/10.3390/metabo11080535