Antimicrobial Effect of Visible Light—Photoinactivation of Legionella rubrilucens by Irradiation at 450, 470, and 620 nm


Abstract
1. Introduction
2. Results
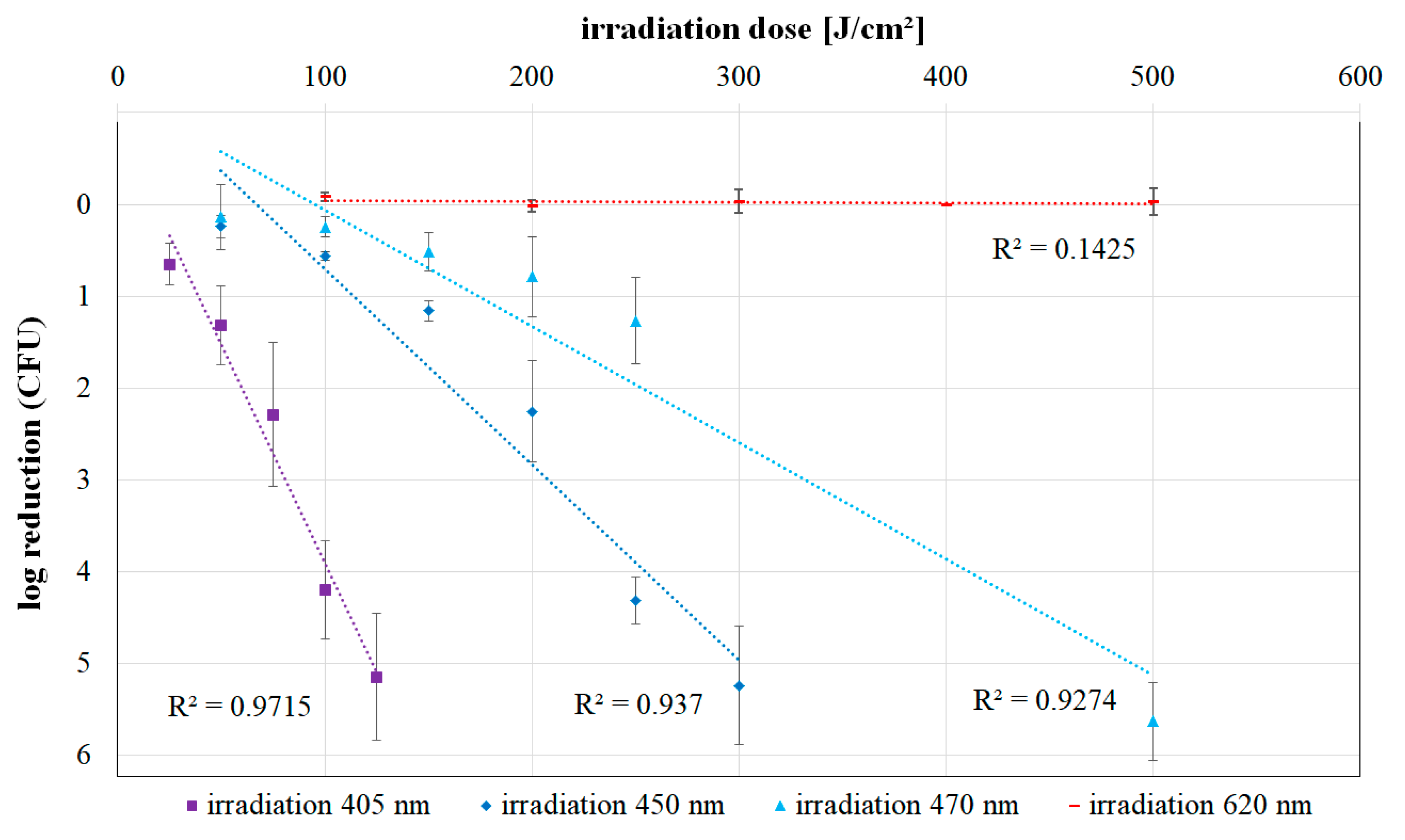
2.1. Irradiation Experiments
2.2. Colony Quantification
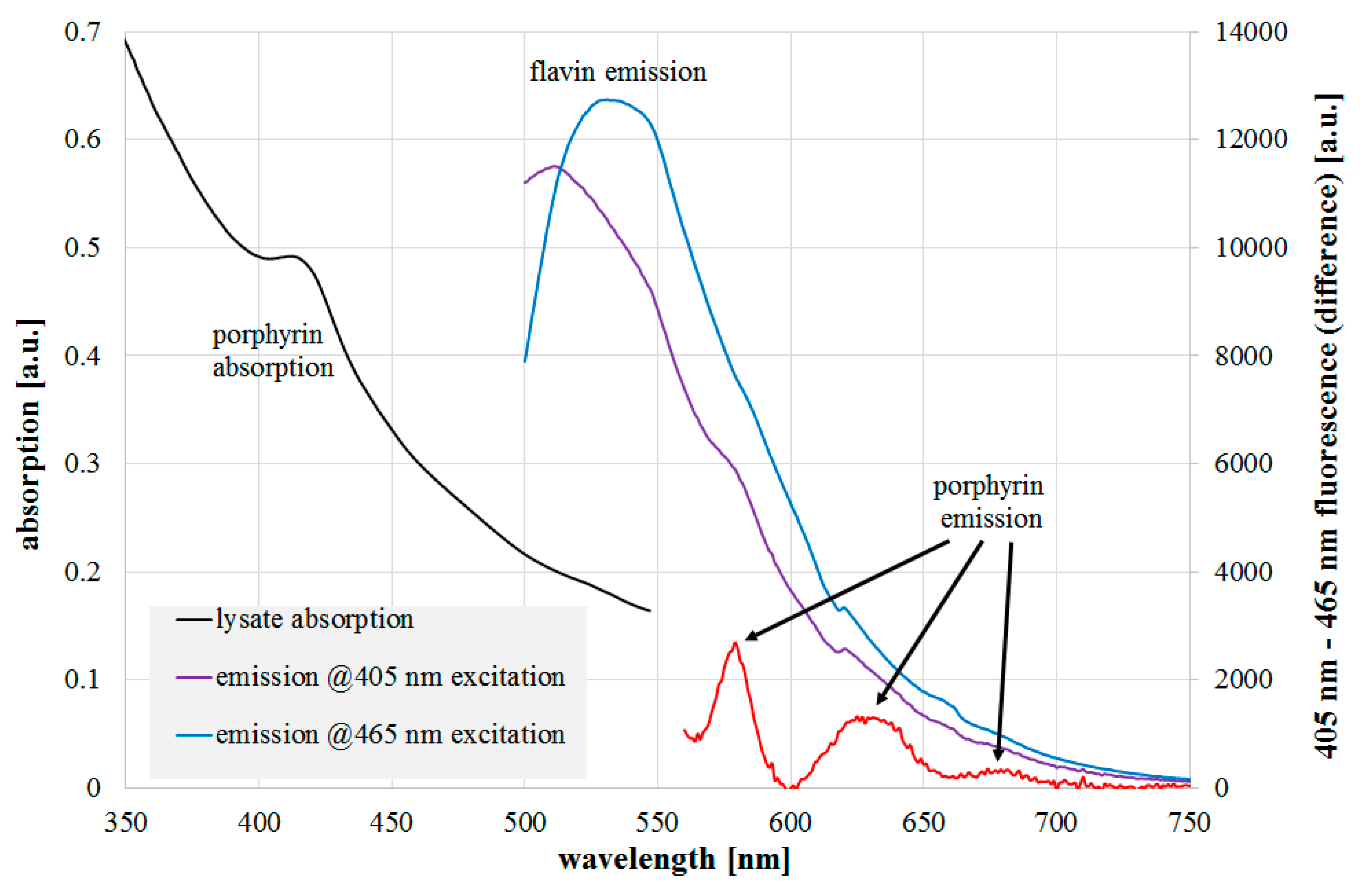
2.3. Spectroscopic Investigations
3. Discussion and Conclusions
4. Materials and Methods
4.1. Bacterial Preparation
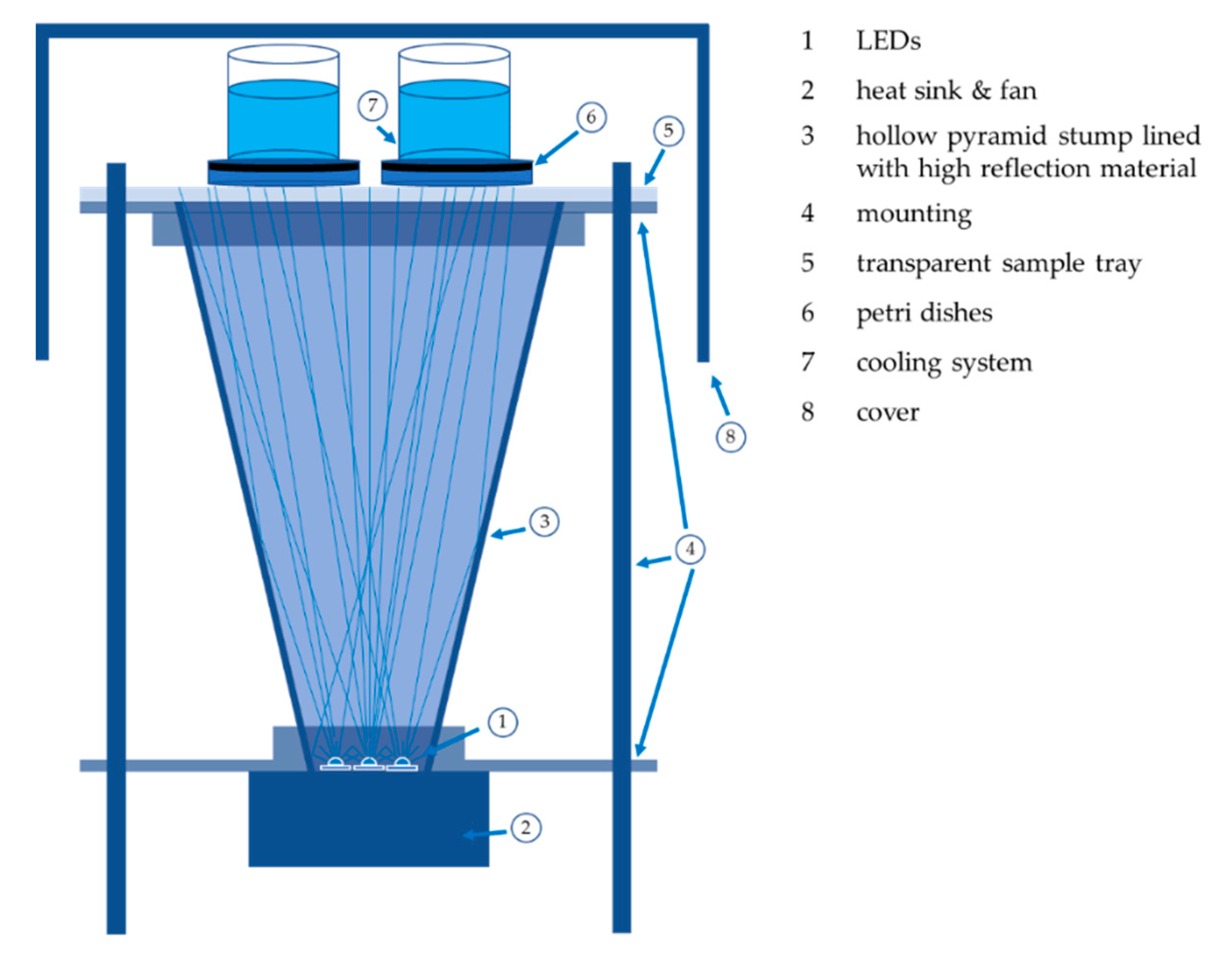
4.2. Irradiation Experiments
4.3. Colony Quantification
4.4. Spectroscopic Investigations
Author Contributions
Funding
Conflicts of Interest
References
- Atlas, R.M. Legionella: From environmental habitats to disease pathology, detection and control. Environ. Microbiol. 1999, 1, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Patterson, W.J.; Hay, J.; Seal, D.V.; McLuckie, J.C. Colonization of transplant unit water supplies with Legionella and protozoa: Precautions required to reduce the risk of legionellosis. J. Hosp. Infect. 1997, 37, 7–17. [Google Scholar] [CrossRef]
- von Baum, H.; Luck, C. Ambulant erworbene Legionellenpneumonie: Aktuelle Daten aus dem CAPNETZ. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2011, 54, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Hessling, M.; Feiertag, J.; Hönes, K. Pathogens provoking most deaths worldwide. Biosci. Biotechnol. Res. Commun. 2017, 10, 1–7. [Google Scholar]
- Hamilton, K.A.; Hamilton, M.T.; Johnson, W.; Jjemba, P.; Bukhari, Z.; LeChevallier, M.; Haas, C.N. Health risks from exposure to Legionella in reclaimed water aerosols: Toilet flushing, spray irrigation, and cooling towers. Water Res. 2018, 134, 261–279. [Google Scholar] [CrossRef] [PubMed]
- Sedlata Juraskova, E.; Sedlackova, H.; Janska, J.; Holy, O.; Lalova, I.; Matouskova, I. Legionella spp. in dental unit waterlines. Bratisl. Lek. Listy 2017, 118, 310–314. [Google Scholar] [CrossRef] [PubMed]
- Chevrefils, G.; Caron, E. UV Dose Required to Achieve Incremental Log Inactivation of Bacteria, Protozoa and Viruses1. IUVA News 2006, 8, 38–45. [Google Scholar]
- Kleinpenning, M.M.; Smits, T.; Frunt, M.H.A.; van Erp, P.E.J.; van de Kerkhof, P.C.M.; Gerritsen, R.M.J.P. Clinical and histological effects of blue light on normal skin. Photodermatol. Photoimmunol. Photomed. 2010, 26, 16–21. [Google Scholar] [CrossRef]
- McDonald, R.S.; Gupta, S.; Maclean, M.; Ramakrishnan, P.; Anderson, J.G.; MacGregor, S.J.; Meek, R.M.D.; Grant, M.H. 405 nm Light exposure of osteoblasts and inactivation of bacterial isolates from arthroplasty patients: Potential for new disinfection applications? Eur. Cell. Mater. 2013, 25, 204–214. [Google Scholar] [CrossRef]
- Ramakrishnan, P.; Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Grant, M.H. Differential sensitivity of osteoblasts and bacterial pathogens to 405 nm light highlighting potential for decontamination applications in orthopedic surgery. J. Biomed. Opt. 2014, 19, 105001. [Google Scholar] [CrossRef]
- Ramakrishnan, P.; Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Grant, M.H. Cytotoxic responses to 405 nm light exposure in mammalian and bacterial cells: Involvement of reactive oxygen species. Toxicol. Vitr. 2016, 33, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Ashkenazi, H.; Malik, Z.; Harth, Y.; Nitzan, Y. Eradication of propionibacterium acnes by its endogenic porphyrins after illumination with high intensity blue light. FEMS Immunol. Med. Microbiol. 2003, 35, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Feuerstein, O.; Dayan, E.; Ginsburg, I.; Veler, D.; Weiss, E.I. Mechanism of visible light phototoxicity on porphyromonas gingivalis and fusobacterium nucleatum. Photochem. Photobiol. 2005, 81, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Hessling, M.; Spellerberg, B.; Hoenes, K. Photoinactivation of bacteria by endogenous photosensitizers and exposure to visible light of different wavelengths-a review on existing data. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G. High-intensity narrow-spectrum light inactivation and wavelength sensitivity of staphylococcus aureus. FEMS Microbiol. Lett. 2008, 285, 227–232. [Google Scholar] [CrossRef]
- Maclean, M.; MacGregor, S.J.; Anderson, J.G.; Woolsey, G. Inactivation of bacterial pathogens following exposure to light from a 405 nm light-emitting diode array. Appl. Environ. Microbiol. 2009, 75, 1932–1937. [Google Scholar] [CrossRef]
- Tomb, R.M.; White, T.A.; Coia, J.E.; Anderson, J.G.; MacGregor, S.J.; Maclean, M. Review of the comparative susceptibility of microbial species to photoinactivation using 380–480 nm Violet-Blue Light. Photochem. Photobiol. 2018, 94, 445–458. [Google Scholar] [CrossRef]
- Amin, R.M.; Bhayana, B.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of pseudomonas aeruginosa by photo-excitation of endogenous porphyrins: In vitro and in vivo studies. Lasers Surg. Med. 2016, 48, 562–568. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Viveiros, J.; Yang, C.; Ahmadi, A.; Ganz, R.A.; Tolkoff, M.J. Helicobacter pylori accumulates photoactive porphyrins and is killed by visible light. Antimicrob. Agents Chemother. 2005, 49, 2822–2827. [Google Scholar] [CrossRef]
- Dai, T.; Gupta, A.; Murray, C.K.; Vrahas, M.S.; Tegos, G.P.; Hamblin, M.R. Blue light for infectious diseases: Propionibacterium acnes, helicobacter pylori, and beyond? Drug Resist. Updat. 2012, 15, 223–236. [Google Scholar] [CrossRef]
- Hönes, K.; Hess, M.; Vatter, P.; Spellerberg, B.; Hessling, M. 405 nm and 450 nm photoinactivation of saccharomyces cerevisiae. Eur. J. Microbiol. Immunol. 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Plavskii, V.Y.; Mikulich, A.V.; Tretyakova, A.I.; Leusenka, I.A.; Plavskaya, L.G.; Kazyuchits, O.A.; Dobysh, I.I.; Krasnenkova, T.P. Porphyrins and flavins as endogenous acceptors of optical radiation of blue spectral region determining photoinactivation of microbial cells. J. Photochem. Photobiol. B 2018, 183, 172–183. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Hoenes, K.; Rath, M.; Vatter, P.; Spellerberg, B.; Hessling, M. Photoinactivation of legionella rubrilucens by visible light. Eur. J. Microbiol. Immunol. 2017, 7, 146–149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, Y.; Chen, J.; Wang, Y.; Sherwood, M.E.; Murray, C.K.; Vrahas, M.S.; Hooper, D.C.; Hamblin, M.R.; Dai, T. Antimicrobial blue light inactivation of candida albicans: in vitro and in vivo studies. Virulence 2016, 7, 536–545. [Google Scholar] [CrossRef]
- Cieplik, F.; Späth, A.; Leibl, C.; Gollmer, A.; Regensburger, J.; Tabenski, L.; Hiller, K.A.; Maisch, T.; Schmalz, G. Blue light kills aggregatibacter actinomycetemcomitans due to its endogenous photosensitizers. Clin. Oral Investig. 2014, 18, 1763–1769. [Google Scholar] [CrossRef]
- Hönes, K.; Spellerberg, B.; Hessling, M. Photoinactivation sensitivity of staphylococcus carnosus to visible light irradiation as a function of wavelength. Photochem. Photobiol. 2019. [Google Scholar] [CrossRef]
- Matsui, M.; Fujii, S.; Shiroiwa, R.; Amemura-Maekawa, J.; Chang, B.; Kura, F.; Yamauchi, K. Isolation of Legionella rubrilucens from a pneumonia patient co-infected with legionella pneumophila. J. Med. Microbiol. 2010, 59, 1242–1246. [Google Scholar] [CrossRef]
- Lipovsky, A.; Nitzan, Y.; Friedmann, H.; Lubart, R. Sensitivity of staphylococcus aureus strains to broadband visible light. Photochem. Photobiol. 2009, 85, 255–260. [Google Scholar] [CrossRef]
- Wang, Y.; Ferrer-Espada, R.; Baglo, Y.; Gu, Y.; Dai, T. Antimicrobial blue light inactivation of neisseria gonorrhoeae: Roles of wavelength, endogenous photosensitizer, oxygen, and reactive oxygen species. Lasers Surg. Med. 2019. [CrossRef]
- Webb, R.B.; Brown, M.S. Sensitivity of strains of escherichia coli differing in repair capability to far UV, near UV and visible radiations. Photochem. Photobiol. 1976, 24, 425–432. [Google Scholar] [CrossRef]
- Sailer, R.; Strauss, W.S.L.; König, K.; Rück, A.; Steiner, R. Correlation between porphyrin biosynthesis and photodynamic inactivation of pseudomonas aeruginosa after incubation with 5-aminolaevulinic amid. J. Photochem. Photobiol. B 1997, 39, 236–242. [Google Scholar] [CrossRef]
- Gourmelon, M.; Cillard, J.; Pommepuy, M. Visible light damage to escherichia coli in seawater: Oxidative stress hypothesis. J. Appl. Bacteriol. 1994, 77, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Fila, G.; Kawiak, A.; Grinholc, M.S. Blue light treatment of pseudomonas aeruginosa: Strong bactericidal activity, synergism with antibiotics and inactivation of virulence factors. Virulence 2017, 8, 938–958. [Google Scholar] [CrossRef] [PubMed]
- Waterworth, P.M. The action of light on culture media. J. Clin. Pathol. 1969, 22, 273–277. [Google Scholar] [CrossRef]
- Xu, J.; Hurlbert, R.E. Toxicity of irradiated media for xenorhabdus spp. Appl. Environ. Microbiol. 1990, 56, 815–818. [Google Scholar]
- Edwards, A.M.; Silva, E. Effect of visible light on selected enzymes, vitamins and amino acids. J. Photochem. Photobiol. B 2001, 63, 126–131. [Google Scholar] [CrossRef]
- Grzelak, A.; Rychlik, B.; Bartosz, G. Light-Dependent generation of reactive oxygen species in cell culture media. Free Radic. Biol. Med. 2001, 30, 1418–1425. [Google Scholar] [CrossRef]
- Trzaska, W.J.; Wrigley, H.E.; Thwaite, J.E.; May, R.C. Species-Specific antifungal activity of blue light. Sci. Rep. 2017, 7, 4605. [Google Scholar] [CrossRef]
- Hoffman, P.S.; Pine, L.; Bell, S. Production of superoxide and hydrogen peroxide in medium used to culture legionella pneumophila: Catalytic decomposition by charcoal. Appl. Environ. Microbiol. 1983, 45, 784–791. [Google Scholar]
- Deutsches Institut für Normung. Wasserbeschaffenheit—Zählung von Legionellen; Beuth-Verlag: Berlin, Germany, 2018. [Google Scholar]
- Buchrieser, C.; Hilbi, H. (Eds.) Legionella. Methods and Protocols; Humana Press: New York, NY, USA, 2013; ISBN 978-1-62703-160-8. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wavelength (nm) | Irradiation Dose (J cm−2) | Log Reduction | Standard Deviation of Log Reduction |
|---|---|---|---|
| 450 | 50 | 0.243 | 0.123 |
| 100 | 0.565 | 0.044 | |
| 150 | 1.162 | 0.107 | |
| 200 | 2.255 | 0.556 | |
| 250 | 4.317 | 0.258 | |
| 300 | 5.243 | 0.648 | |
| 470 | 50 | 0.137 | 0.353 |
| 100 | 0.249 | 0.110 | |
| 150 | 0.519 | 0.210 | |
| 200 | 0.788 | 0.434 | |
| 250 | 1.267 | 0.467 | |
| 500 | 5.633 | 0.425 | |
| 620 | 100 | −0.082 | 0.050 |
| 200 | 0.019 | 0.067 | |
| 300 | −0.034 | 0.133 | |
| 400 | 0.000 | 0.002 | |
| 500 | −0.027 | 0.144 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmid, J.; Hoenes, K.; Vatter, P.; Hessling, M. Antimicrobial Effect of Visible Light—Photoinactivation of Legionella rubrilucens by Irradiation at 450, 470, and 620 nm. Antibiotics 2019, 8, 187. https://doi.org/10.3390/antibiotics8040187
Schmid J, Hoenes K, Vatter P, Hessling M. Antimicrobial Effect of Visible Light—Photoinactivation of Legionella rubrilucens by Irradiation at 450, 470, and 620 nm. Antibiotics. 2019; 8(4):187. https://doi.org/10.3390/antibiotics8040187
Chicago/Turabian StyleSchmid, Julian, Katharina Hoenes, Petra Vatter, and Martin Hessling. 2019. "Antimicrobial Effect of Visible Light—Photoinactivation of Legionella rubrilucens by Irradiation at 450, 470, and 620 nm" Antibiotics 8, no. 4: 187. https://doi.org/10.3390/antibiotics8040187
APA StyleSchmid, J., Hoenes, K., Vatter, P., & Hessling, M. (2019). Antimicrobial Effect of Visible Light—Photoinactivation of Legionella rubrilucens by Irradiation at 450, 470, and 620 nm. Antibiotics, 8(4), 187. https://doi.org/10.3390/antibiotics8040187