


Harnessing the Potential of Antibacterial and Antibiofilm Phytochemicals in the Combat Against Superbugs: A One Health Perspective

 , , and
, , and 
Abstract
1. Introduction
2. Threat Posed by Drug-Resistant Bacteria
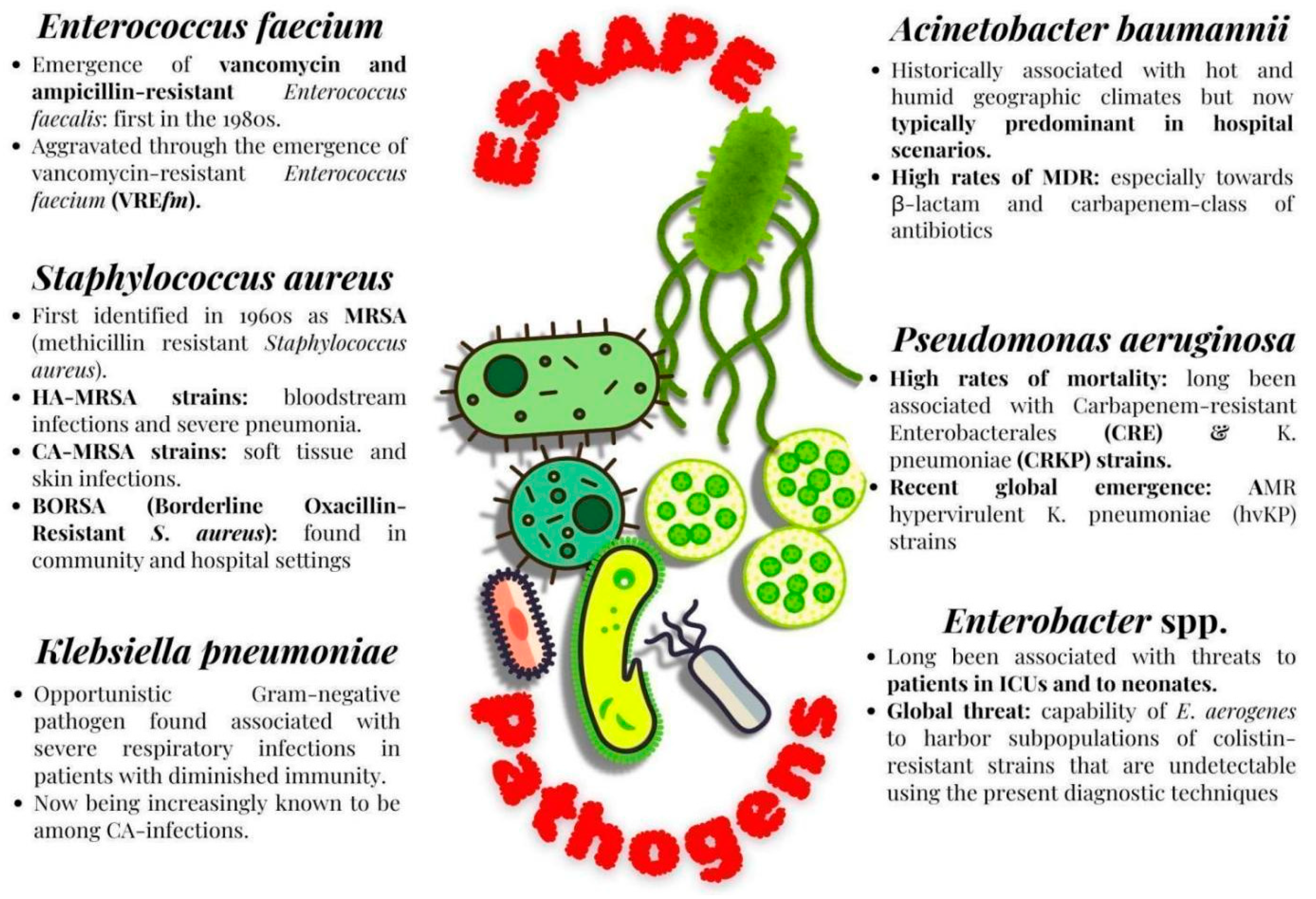
2.1. Characteristic Features of ESKAPE Pathogens
2.1.1. Enterococcus Faecium
2.1.2. Methicillin-Resistant Staphylococcus aureus (MRSA)
2.1.3. Klebsiella pneumoniae
2.1.4. Acinetobacter baumannii
2.1.5. Pseudomonas aeruginosa
2.1.6. Enterobacter Species
3. Survival Strategies of Bacteria
4. Phytochemicals with Antibacterial Properties
4.1. Phytochemicals Inhibiting Gram-Negative Bacteria
4.2. Phytochemicals Inhibiting Gram-Positive Bacteria
4.3. Phytochemicals Inhibiting Mycobacterium Tuberculosis
5. Phytochemicals with Antibiofilm Properties
5.1. Alkaloids
5.2. Flavonoids
5.3. Saponins
5.4. Tannins
5.5. Phenolics
6. Mechanisms Disrupting Bacterial Growth and Biofilm Formation
7. Limitations of Current Antibacterial Strategies
8. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, S.; Barbosa, C.; Woods, R. Why is Preventing Antibiotic Resistance So Hard? Analysis of Failed Resistance Management. Evol. Med. Public Health 2020, 2020, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Zeiler, M.; Melander, R.; Melander, C. Second-Generation Meridianin Analogues Inhibit the Formation of Mycobacterium smegmatis Biofilms and Sensitize Polymyxin-Resistant Gram-Negative Bacteria to Colistin. ChemMedChem 2020, 15, 1672–1679. [Google Scholar] [CrossRef]
- Wang, Z.; Tian, W.; Sun, S.; Chen, X.; Wang, H. Genomic and Proteomic Analysis of Pseudomonas Aeruginosa Isolated from Industrial Wastewater to Assess its Resistance to Antibiotics. Separations 2023, 10, 549. [Google Scholar] [CrossRef]
- Li, J.; Li, Y.; Cao, X.; Zheng, J.; Zhang, Y.; Xie, H.; Li, C.; Liu, C.; Shen, H. Genome-Wide Identification and Oxacillinase OXA Distribution Characteristics of Acinetobacter spp. Based on a Global Database. Front. Microbiol. 2023, 14, 1174200. [Google Scholar] [CrossRef]
- Alsubaie, M.; Alsuheili, A.; Aljehani, M.; Alothman, A.; Alzahrani, A.; Mohammedfadel, H.; Alnajjar, A. Antibiotic Resistance Patterns of Pediatric Community-Acquired Urinary Tract Infections in a Tertiary Care Center in Jeddah, Saudi Arabia. J. Infect. Dev. Ctries. 2023, 17, 1430–1435. [Google Scholar] [CrossRef]
- Zhao, A.; Sun, J.; Liu, Y. Understanding Bacterial Biofilms: From Definition to Treatment Strategies. Front. Cell. Infect. Microbiol. 2023, 13, 1137947. [Google Scholar] [CrossRef]
- Mukhopadhyay, S.; Singh, M.; Ghosh, M.M.; Chakrabarti, S.; Ganguli, S. Comparative Genomics and Characterization of Shigella flexneri Isolated from Urban Wastewater. Microbes Environ. 2024, 39, ME23105. [Google Scholar] [CrossRef]
- Wójcicki, M.; Świder, O.; Daniluk, K.J.; Średnicka, P.; Kowalczyk, M.; Roszko, M.; Juszczuk-Kubiak, E. Transcriptional Regulation of the Multiple Resistance Mechanisms in Salmonella—A Review. Pathogens 2021, 10, 801. [Google Scholar] [CrossRef]
- Li, W.; Hu, J.; Li, L.; Zhang, M.; Cui, Q.; Ma, Y.; Wang, M. New Mutations in cls Lead to Daptomycin Resistance in a Clinical Vancomycin- and Daptomycin-Resistant Enterococcus faecium Strain. Front. Microbiol. 2022, 13, 896916. [Google Scholar] [CrossRef]
- Banerjee, D.K.; Biswas, P.; Mazumder, K.; Palai, S.; Hossain, C.M.; Karmakar, S.; Biswas, K. Exploration of Phytochemicals as Anti-Biofilm Agents Against Pathogenic Bacteria: Their Potential and Challenges. Infect. Disord. Drug Targets 2024, 25, 1204182204. [Google Scholar] [CrossRef]
- Kanwar, K.; Thakur, P.; Azmi, W. Use of Phytochemicals as Emerging Strategy for Control of Biofilm Formed by Pathogens. Ann. Phytomed. 2018, 7, 25–37. [Google Scholar] [CrossRef]
- Hannan, A.; Du, X.X.; Maqbool, B.; Khan, A. Nanoparticles as potent allies in combating antibiotic resistance: A promising frontier in antimicrobial therapy. Pak. Vet. J. 2024, 44, 957–967. [Google Scholar] [CrossRef]
- Kinsella, R.L.; Kimmey, J.M.; Smirnov, A.; Woodson, R.; Gaggioli, M.R.; Chavez, S.M.; Kreamalmeyer, D.; Stallings, C.L. Autophagy Prevents Early Proinflammatory Responses and Neutrophil Recruitment during Mycobacterium tuberculosis Infection Without Affecting Pathogen Burden in Macrophages. PLoS Biol. 2023, 21, e3002159. [Google Scholar] [CrossRef]
- Saad, N.; El-Abasy, M.A.; El-Khayat, F.; Ali, N.G.; Ismail, M.M. Efficacy of Chitosan Nanoparticles as a Natural Antibacterial Agent Against Pathogenic Bacteria Causing Omphalitis in Poultry. Pak. Vet. J. 2023, 43, 573–578. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Leiknes, T. Quorum-Quenching Bacteria Isolated from Red Sea Sediments Reduce Biofilm Formation by Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 1354. [Google Scholar] [CrossRef]
- Balloux, F.; van Dorp, L. Q&A: What are Pathogens, and What Have They Done to and for us? BMC Biol. 2017, 15, 91. [Google Scholar] [CrossRef]
- Reeves, A.B.; Ramey, A.M.; Koch, J.C.; Poulson, R.L.; Stallknecht, D.E. Field-Based Method for Assessing Duration of Infectivity for Influenza A Viruses in the Environment. J. Virol. Methods 2020, 277, 113818. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Bartlett, J.; Wunderink, R.; Gilbert, D.N. Novel Approaches Are Needed to Develop Tomorrow’s Antibacterial Therapies. Am. J. Respir. Crit. Care Med. 2015, 191, 135–140. [Google Scholar] [CrossRef] [PubMed]
- McCann, C.M.; Christgen, B.; Roberts, J.A.; Su, J.Q.; Arnold, K.E.; Gray, N.D.; Graham, D.W. Understanding Drivers of Antibiotic Resistance Genes in High Arctic Soil Ecosystems. Environ. Int. 2019, 125, 497–504. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Kafil, H.S. Molecular Mechanisms Related to Colistin Resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC); National Center for Emerging and Zoonotic Infectious Diseases (NCEZID); Division of Healthcare Quality Promotion (DHQP). 2019 AR Threats Report; U.S. Department of Health & Human Services: Washington, DC, USA, 2020.
- World Health Organization. WHO Bacterial Priority Pathogens List, 2024: Bacterial Pathogens of Public Health Importance, to Guide Research, Development, and Strategies to Prevent and Control Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2024; Available online: https://www.who.int/publications/i/item/9789240093461 (accessed on 22 May 2025).
- World Health Organization. Global Antimicrobial Resistance and Use Surveillance System (GLASS) Report 2022; World Health Organization: Geneva, Switzerland, 2022; Available online: https://iris.who.int/handle/10665/364996 (accessed on 22 May 2025).
- Flemming, H.C.; Wuertz, S. Bacteria and Archaea on Earth and Their Abundance in Biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Meesilp, N.; Mesil, N. Effect of Microbial Sanitizers for Reducing Biofilm Formation of Staphylococcus aureus and Pseudomonas aeruginosa on Stainless Steel by Cultivation with UHT Milk. Food Sci. Biotechnol. 2019, 28, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Carrascosa, C.; Raheem, D.; Ramos, F.; Saraiva, A.; Raposo, A. Microbial Biofilms in the Food Industry—A Comprehensive Review. Int. J. Environ. Res. Public Health 2021, 18, 2014. [Google Scholar] [CrossRef]
- Sharahi, J.Y.; Azimi, T.; Shariati, A.; Safari, H.; Tehrani, M.K.; Hashemi, A. Advanced Strategies for Combating Bacterial Biofilms. J. Cell. Physiol. 2019, 234, 14689–14708. [Google Scholar] [CrossRef] [PubMed]
- Velazquez-Meza, M.E.; Galarde-López, M.; Carrillo-Quiróz, B.; Alpuche-Aranda, C.M. Antimicrobial resistance: One health approach. Vet. World 2022, 15, 743. [Google Scholar] [CrossRef]
- Zelaya, C.A.; Arriagada, G.; Medina, R.; Escobar, B.; Sánchez, F.; Galarce, N.; Lapierre, L. The Risk Factors Associated with the Carriage to Critical Antimicrobial-Resistant Escherichia coli in Healthy Household Dogs: A One Health Perspective. Animals 2025, 15, 1357. [Google Scholar] [CrossRef]
- Salvarani, F.M.; Oliveira, H.G.d.S.; Uzal, F.A. Clostridioides difficile in Animal Inflammatory Bowel Disease: A One Health Perspective on Emerging Zoonotic Threats. Microorganisms 2025, 13, 1233. [Google Scholar] [CrossRef]
- Kim, D.W.; Cha, C.J. Antibiotic resistome from the One-Health perspective: Understanding and controlling antimicrobial resistance transmission. Exp. Mol. Med. 2021, 53, 301–309. [Google Scholar] [CrossRef]
- Tacconelli, E. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Beatson, S.A.; Walker, M.J. Tracking Antibiotic Resistance. Science 2014, 345, 1454–1455. [Google Scholar]
- Naylor, N.R.; Atun, R.; Zhu, N.; Kulasabanathan, K.; Silva, S.; Chatterjee, A.; Robotham, J.V. Estimating the Burden of Antimicrobial Resistance: A Systematic Literature Review. Antimicrob. Resist. Infect. Control 2018, 7, 129. [Google Scholar] [CrossRef]
- Murray, B.E. The Life and Times of the Enterococcus. Clin. Microbiol. Rev. 1990, 3, 46–65. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.; Pang, S.; Abraham, S.; Coombs, G.W. Antimicrobial-Resistant CC17 Enterococcus faecium: The Past, the Present and the Future. J. Glob. Antimicrob. Resist. 2019, 16, 36–47. [Google Scholar] [CrossRef]
- Satilmis, L.; Vanhems, P.; Benet, T. Outbreaks of Vancomycin-Resistant Enterococci in Hospital Settings: A Systematic Review and Calculation of the Basic Reproductive Number. Infect. Control Hosp. Epidemiol. 2016, 37, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, K.R. “Celbenin”-Resistant Staphylococci. Ugeskr. Laeger 1961, 123, 384–386. [Google Scholar] [PubMed]
- DeLeo, F.R.; Otto, M.; Kreiswirth, B.N.; Chambers, H.F. Community-Associated Meticillin-Resistant Staphylococcus aureus. Lancet 2010, 375, 1557–1568. [Google Scholar] [CrossRef]
- Skinner, S.; Murray, M.; Walus, T.; Karlowsky, J.A. Failure of Cloxacillin in Treatment of a Patient with Borderline Oxacillin-Resistant Staphylococcus aureus Endocarditis. J. Clin. Microbiol. 2009, 47, 859–861. [Google Scholar] [CrossRef]
- Tzouvelekis, L.S.; Markogiannakis, A.; Psichogiou, M.; Tassios, P.T.; Daikos, G.L. Carbapenemases in Klebsiella pneumoniae and Other Enterobacteriaceae: An Evolving Crisis of Global Dimensions. Clin. Microbiol. Rev. 2012, 25, 682–707. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services; Centers for Disease Control and Prevention. Antibiotic Resistance Threats in the United States; U.S. Department of Health and Human Services: Washington, DC, USA, 2013.
- Krapp, F.; Morris, A.R.; Ozer, E.A.; Hauser, A.R. Virulence Characteristics of Carbapenem-Resistant Klebsiella pneumoniae Strains from Patients with Necrotizing Skin and Soft Tissue Infections. Sci. Rep. 2017, 7, 13533. [Google Scholar] [CrossRef]
- Li, W.; Sun, G.; Yu, Y.; Li, N.; Chen, M.; Jin, R.; Wu, H. Increasing Occurrence of Antimicrobial-Resistant Hypervirulent (Hypermucoviscous) Klebsiella pneumoniae Isolates in China. Clin. Infect. Dis. 2014, 58, 225–232. [Google Scholar] [CrossRef]
- Ibrahim, M.E. Prevalence of Acinetobacter baumannii in Saudi Arabia: Risk Factors, Antimicrobial Resistance Patterns and Mechanisms of Carbapenem Resistance. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 1. [Google Scholar] [CrossRef]
- Xie, R.; Zhang, X.D.; Zhao, Q.; Peng, B.; Zheng, J. Analysis of Global Prevalence of Antibiotic Resistance in Acinetobacter baumannii Infections Disclosed a Faster Increase in OECD Countries. Emerg. Microbes Infect. 2018, 7, 31. [Google Scholar] [CrossRef]
- Gellatly, S.L.; Hancock, R.E.W. Pseudomonas aeruginosa: New Insights into Pathogenesis and Host Defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Davin-Regli, A.; Pagès, J.M. Enterobacter aerogenes and Enterobacter cloacae; Versatile Bacterial Pathogens Confronting Antibiotic Treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef]
- Band, V.I.; Crispell, E.K.; Napier, B.A.; Herrera, C.M.; Tharp, G.K.; Vavikolanu, K.; Weiss, D.S. Antibiotic Failure Mediated by a Resistant Subpopulation in Enterobacter cloacae. Nat. Microbiol. 2016, 1, 16053. [Google Scholar] [CrossRef]
- Baker, S.J.; Payne, D.J.; Rappuoli, R.; De Gregorio, E. Technologies to Address Antimicrobial Resistance. Proc. Natl. Acad. Sci. USA 2018, 115, 12887–12895. [Google Scholar] [CrossRef] [PubMed]
- Munita, J.M.; Arias, C.A. Mechanisms of Antibiotic Resistance. In Virulence Mechanisms of Bacterial Pathogens; ASM Press: Washington, DC, USA, 2016; pp. 481–511. [Google Scholar]
- Lerminiaux, N.A.; Cameron, A.D.S. Horizontal Transfer of Antibiotic Resistance Genes in Clinical Environments. Can. J. Microbiol. 2019, 65, 34–44. [Google Scholar] [CrossRef]
- Pacios, O.; Blasco, L.; Bleriot, I.; Fernandez-Garcia, L.; González Bardanca, M.; Ambroa, A.; Tomás, M. Strategies to Combat Multidrug-Resistant and Persistent Infectious Diseases. Antibiotics 2020, 9, 65. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, C.H.; Li, X.; Li, F.; Jiang, M.L.; Liu, Z.K.; Li, J.Y. Characteristics of The Plasmid-Mediated Colistin-Resistance Gene Mcr-1 In Escherichia Coli Isolated from Pig Farm in Jiangxi. Pak. Vet. J. 2024, 44, 1303–1307. [Google Scholar] [CrossRef]
- Shafiq, M.; Zeng, M.; Permana, B.; Bilal, H.; Huang, J.; Yao, F.; Jiao, X. Coexistence of blaNDM–5 and tet(X4) in International High-Risk Escherichia coli Clone ST648 of Human Origin in China. Front. Microbiol. 2022, 13, 1031688. [Google Scholar] [CrossRef]
- Van den Bergh, B.; Fauvart, M.; Michiels, J. Formation, Physiology, Ecology, Evolution and Clinical Importance of Bacterial Persisters. FEMS Microbiol. Rev. 2017, 41, 219–251. [Google Scholar] [CrossRef]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing Between Resistance, Tolerance and Persistence to Antibiotic Treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef]
- Fisher, R.A.; Gollan, B.; Helaine, S. Persistent Bacterial Infections and Persister Cells. Nat. Rev. Microbiol. 2017, 15, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Zinkernagel, A. Definitions and Guidelines for Research on Antibiotic Persistence. Nat. Rev. Microbiol. 2019, 17, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Ayrapetyan, M.; Williams, T.; Oliver, J.D. Relationship Between the Viable but Nonculturable State and Antibiotic Persister Cells. J. Bacteriol. 2018, 200, e00249-18. [Google Scholar] [CrossRef]
- Bodor, A.; Bounedjoum, N.; Vincze, G.E.; Erdeiné Kis, Á.; Laczi, K.; Bende, G.; Rákhely, G. Challenges of Unculturable Bacteria: Environmental Perspectives. Rev. Environ. Sci. Biotechnol. 2020, 19, 1–22. [Google Scholar] [CrossRef]
- Chebotar, I.V.; Emelyanova, M.A.; Bocharova, J.A.; Mayansky, N.A.; Kopantseva, E.E.; Mikhailovich, V.M. The Classification of Bacterial Survival Strategies in the Presence of Antimicrobials. Microb. Pathog. 2021, 155, 104901. [Google Scholar] [CrossRef]
- Pangeni, B.; Bhattarai, S.; Paudyal, H.; Chaudhary, R.P. Antibacterial Activity of Selected Ethnomedicinal Plants Popular in Magar Ethnic Community of Palpa District, Western Nepal. Arch. Ecotoxicol. 2021, 3, 13–19. [Google Scholar] [CrossRef]
- Neagu, R.; Popovici, V.; Ionescu, L.E.; Ordeanu, V.; Biță, A.; Popescu, D.M.; Gîrd, C.E. Phytochemical screening and antibacterial activity of commercially available essential oils combinations with conventional antibiotics against gram-positive and gram-negative bacteria. Antibiotics 2024, 13, 478. [Google Scholar] [CrossRef]
- Gupta, V.; Kumar, R.; Chaudhary, D.; Yadav, N. In-Vitro Analysis of Potential Antibacterial Activity of Three Medicinal Plants. J. Appl. Nat. Sci. 2016, 8, 1497–1500. [Google Scholar] [CrossRef]
- Rauf, A.; Abu-Izneid, T.; Rashid, U.; Alhumaydhi, F.A.; Bawazeer, S.; Khalil, A.A.; Ntsefong, G.N. Anti-Inflammatory, Antibacterial, Toxicological Profile, and In Silico Studies of Dimeric Naphthoquinones from Diospyros lotus. BioMed Res. Int. 2020, 2020, 7942549. [Google Scholar] [CrossRef]
- Mogana, R.; Teng-Jin, K.; Wiart, C. In vitro antimicrobial, antioxidant activities and phytochemical analysis of Canarium patentinervium Miq. from Malaysia. Biotechnol. Res. Int. 2011, 2011, 768673. [Google Scholar] [CrossRef]
- Elbalola, A.A.; Abbas, Z.K. Phytochemical Diversity, Classification and Antibacterial Activity of Some Medicinal Plant Species from Tabuk (Saudi Arabia). Chem. Biodivers. 2023, 20, e202300545. [Google Scholar] [CrossRef]
- Kakaraparthy, R.; Sruthi, K.S.; Banerjee, M. Evaluation of Antibacterial and Wound Healing Activities of Ethanolic Extract of Linum usitatissimum (Flax Seed) on Rabbits. J. Clin. Pharm. Res. 2023, 3, 13–21. [Google Scholar] [CrossRef]
- Shiferaw, Z.; Sasikumar, J.M.; Kebede, A.; Teju, E. Antibacterial Effects of Extracts from Tanacetum parthenium L. Leaves. Bact. Emp. 2022, 5, e394. [Google Scholar] [CrossRef]
- Atta, S.; Waseem, D.; Fatima, H.; Naz, I.; Rasheed, F.; Kanwal, N. Antibacterial potential and synergistic interaction between natural polyphenolic extracts and synthetic antibiotic on clinical isolates. Saudi J. Biol. Sci. 2023, 30, 103576. [Google Scholar] [CrossRef]
- Istaufa, F.; Subagio, Y.; Suswati, I. Ricinus communis L. Leaf Extract as Potential Antibacterial Against the Growth of Mycobacterium tuberculosis. Folia Med. Indones. 2022, 58, 371–376. [Google Scholar] [CrossRef]
- Puspaningtyas, A.R.; Nugraha, A.S.; Retnaningtyas, Y.; Zulaiha, S. Anti-Tuberculosis Study of Mycobacterium tuberculosis H37Rv of Aspilia pluriceta Extract and Fractions. Trop. J. Nat. Prod. Res. 2024, 8, 6516–6518. [Google Scholar]
- Sarkar, S.; Singh, R.P. Nyctanthes arbortristis L.: Perspective of Phytochemical-Based Inhibition of Fatty Acid Biosynthesis in Mycobacterium tuberculosis. Int. J. Plant Based Pharm. 2022, 2, 166–175. [Google Scholar] [CrossRef]
- Moni, E.D.F.N.; Betote, P.H.D.; Kom, C.W.; Benga, C.F.M.; Tchamgoue, A.D.; Nyegue, M.A. Inhibitory Effects of Hydroethanolic Extracts from Three Cameroonian Medicinal Plants on Proteins Inflammation and Growth of Multi-Resistant Strains of Mycobacterium tuberculosis. J. Drug Deliv. Ther. 2021, 11, 15–21. [Google Scholar] [CrossRef]
- Issa, N.A. Evaluation the Antimicrobial Activity of Essential Oils Against Veterinary Pathogens, Multidrug-resistant Bacteria and Dermatophytes. Pak. Vet. J. 2024, 44, 260–265. [Google Scholar] [CrossRef]
- Aljohani, A.S.M. Botanical compounds: A promising approach to control Mycobacterium species of veterinary and zoonotic importance. Pak. Vet. J. 2023, 43, 633–642. [Google Scholar] [CrossRef]
- Qian, W.; Liu, M.; Fu, Y.; Zhang, J.; Liu, W.; Li, J.; Li, X.; Li, Y.; Wang, T. Antimicrobial mechanism of luteolin against Staphylococcus aureus and Listeria monocytogenes and its antibiofilm properties. Microb. Pathog. 2020, 142, 104056. [Google Scholar] [CrossRef]
- Zhong, Z.X.; Zhou, S.; Liang, Y.J.; Wei, Y.Y.; Li, Y.; Long, T.F.; Sun, J. Natural flavonoids disrupt bacterial iron homeostasis to potentiate colistin efficacy. Sci. Adv. 2023, 9, eadg4205. [Google Scholar] [CrossRef]
- Osagie, E.; Erhauyi, O.; Udogadi, N.; Olalekan, S. Action of Fractionated Moringa oleifera Lam Leaf Extracts on Multidrug Resistant Pseudomonas aeruginosa Strains. Int. J. Clin. Exp. Med. Res. 2021, 5, 316–329. [Google Scholar] [CrossRef]
- Musuasua, M.; Kabena, O.; Kalanda, L.; Kangudia, B.; Mutembue, D.; Masens, D.; Mpiana, P. Phytochemical Screening and In Vitro Antibacterial Activity of Aqueous Extracts of Phyllanthus muellerianus (Kuntze) Exell from Kasaï Oriental (DRC) on a Few Bacterial Strains. Int. J. Pathog. Res. 2022, 9, 22–33. [Google Scholar] [CrossRef]
- Semwal, P.; Painuli, S.; Badoni, H.; Bacheti, R. Screening of Phytoconstituents and Antibacterial Activity of Leaves and Bark of Quercus leucotrichophora A. Camus from Uttarakhand Himalaya. Clin. Phytosci. 2018, 4, 30. [Google Scholar] [CrossRef]
- Silva Leandro, M.K.D.N.; Rocha, J.E.; Bezerra, C.F.; Freitas, P.R.; Feitosa, J.H.F.; Bezerra, V.B.; Barros, R.d.O.; Leandro, L.M.G.; Aguiar, J.J.d.S.; Pereira, P.S.; et al. Modulation of Antibiotic Resistance by the Essential Oil of Ocimum gratissimum L. in Association with Light-Emitting Diodes (LED) Lights. Z. Naturforsch. C 2020, 75, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Kumari, B.; Raveesha, H. Phytochemical Analysis and Antibacterial Activity of Andrographis lineata Nees (Acanthaceae). Curr. Bot. 2024, 15, 1–6. [Google Scholar] [CrossRef]
- Mambe, F.; Na-Iya, J.; Fotso, G.; Ashu, F.; Ngameni, B.; Ngadjui, B.; Kuete, V. Antibacterial and Antibiotic Modifying Potential of Crude Extracts, Fractions, and Compounds from Acacia polyacantha Willd. Against MDR Gram-Negative Bacteria. Evid. Based Complement. Alternat. Med. 2019, 2019, 7507549. [Google Scholar] [CrossRef]
- Gangwar, B.; Kumar, S.; Darokar, M. Antioxidant Phytochemicals as Novel Therapeutic Strategies Against Drug-Resistant Bacteria; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Borges, A.; Abreu, A.; Dias, C.; Saavedra, M.; Borges, F.; Simões, M. New Perspectives on the Use of Phytochemicals as an Emergent Strategy to Control Bacterial Infections Including Biofilms. Molecules 2016, 21, 877. [Google Scholar] [CrossRef]
- Barati, M.; Chahardehi, A.M.; Barati, M.; Chahardehi, A.M. Alkaloids: The Potential of Their Antimicrobial Activities of Medicinal Plants. In Medicinal Plants—Chemical, Biochemical, and Pharmacological Approaches; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Parai, D.; Banerjee, M.; Dey, P.; Mukherjee, S.K. Reserpine Attenuates Biofilm Formation and Virulence of Staphylococcus aureus. Microb. Pathog. 2020, 138, 103790. [Google Scholar] [CrossRef] [PubMed]
- Saha, S.; Do, T.; Maycock, J.; Wood, S.; Boesch, C. Antibiofilm Efficacies of Flavonoid-Rich Sweet Orange Waste Extract Against Dual-Species Biofilms. Pathogens 2023, 12, 657. [Google Scholar] [CrossRef]
- Faleye, O.S.; Lee, J.-H.; Lee, J. Selected Flavonoids Exhibit Antibiofilm and Antibacterial Effects Against Vibrio by Disrupting Membrane Integrity, Virulence and Metabolic Activities. Biofilm 2023, 6, 100165. [Google Scholar] [CrossRef]
- Raorane, C.J.; Lee, J.-H.; Kim, Y.-G.; Rajasekharan, S.K.; García-Contreras, R.; Lee, J. Antibiofilm and Antivirulence Efficacies of Flavonoids and Curcumin Against Acinetobacter baumannii. Front. Microbiol. 2019, 10, 990. [Google Scholar] [CrossRef]
- Shang, F.; Wang, H.; Xue, T. Anti-Biofilm Effect of Tea Saponin on a Streptococcus agalactiae Strain Isolated from Bovine Mastitis. Animals 2020, 10, 1713. [Google Scholar] [CrossRef]
- Li, L.; Wei, M.; Yu, H.; Xie, Y.; Guo, Y.; Cheng, Y.; Yao, W. Antifungal Activity of Sapindus Saponins Against Candida albicans: Interruption of Biofilm Formation. J. Herb. Med. 2023, 42, 100776. [Google Scholar] [CrossRef]
- Fink, R.; Filip, S. Surface-Active Natural Saponins: Properties, Safety, and Efficacy. Int. J. Environ. Health Res. 2023, 33, 639–648. [Google Scholar] [CrossRef]
- Adnan, M.; Siddiqui, A.J.; Ashraf, S.A.; Ashraf, M.S.; Alomrani, S.O.; Alreshidi, M.; Tepe, B.; Sachidanandan, M.; Danciu, C.; Patel, M. Saponin-Derived Silver Nanoparticles from Phoenix dactylifera (Ajwa Dates) Exhibit Broad-Spectrum Bioactivities Combating Bacterial Infections. Antibiotics 2023, 12, 1415. [Google Scholar] [CrossRef]
- Villanueva, X.; Zhen, L.; Ares, J.N.; Vackier, T.; Lange, H.; Crestini, C.; Steenackers, H.P. Effect of Chemical Modifications of Tannins on Their Antimicrobial and Antibiofilm Effect Against Gram-Negative and Gram-Positive Bacteria. Front. Microbiol. 2023, 13, 987164. [Google Scholar] [CrossRef]
- Qin, J.; Yu, L.; Peng, F.; Ye, X.; Li, G.; Sun, C.; Cheng, F.; Peng, C.; Xie, X. Tannin Extracted from Penthorum chinense Pursh, a Potential Drug with Antimicrobial and Antibiofilm Effects Against Methicillin-Sensitive Staphylococcus aureus and Methicillin-Resistant Staphylococcus aureus. Front. Microbiol. 2023, 14, 1134207. [Google Scholar] [CrossRef]
- Alejo-Armijo, A.; Cobo, A.; Alejo-Armijo, A.; Altarejos, J.; Salido, S.; Ortega-Morente, E. Evaluation of Antibacterial and Antibiofilm Properties of Phenolics with Coumarin, Naphthoquinone and Pyranone Moieties Against Foodborne Microorganisms. Molecules 2025, 30, 944. [Google Scholar] [CrossRef] [PubMed]
- Glamočlija, J.; Ćirić, A.; Nikolić, M.; Fernandes, Â.; Barros, L.; Calhelha, R.C.; Ferreira, I.C.F.R.; Soković, M.; van Griensven, L.J.L.D. Chemical Characterization and Biological Activity of Chaga (Inonotus obliquus), a Medicinal “Mushroom”. J. Ethnopharmacol. 2015, 162, 323–332. [Google Scholar] [CrossRef]
- Slobodníková, L.; Fialová, S.; Rendeková, K.; Kováč, J.; Mučaji, P. Antibiofilm Activity of Plant Polyphenols. Molecules 2016, 21, 1717. [Google Scholar] [CrossRef] [PubMed]
- Fydrych, D.; Jeziurska, J.; Wełna, J.; Kwiecińska-Piróg, J. Potential Use of Selected Natural Compounds with Anti-Biofilm Activity. Int. J. Mol. Sci. 2025, 26, 607. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Z.; Pan, Q.; Fan, L.; Pan, T.; Zhu, F.; Pan, Q.; Shan, L.; Zhao, L. Berberine at Sub-Inhibitory Concentration Inhibits Biofilm Dispersal in Staphylococcus aureus. Microbiology 2022, 168, 001243. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, W.; Liu, M.; Zhang, J.; Yang, M.; Wang, T.; Qian, W. In vitro anti-biofilm efficacy of sanguinarine against carbapenem-resistant Serratia marcescens. Biofouling 2021, 37, 341–351. [Google Scholar] [CrossRef]
- Das, S.; Malik, M.; Dastidar, D.G.; Roy, R.; Paul, P.; Sarkar, S.; Chakraborty, P.; Maity, A.; Dasgupta, M.; Tribedi, P.; et al. Piperine, a phytochemical prevents the biofilm city of methicillin-resistant Staphylococcus aureus: A biochemical approach to understand the underlying mechanism. Microb. Pathog. 2024, 189, 106601. [Google Scholar] [CrossRef] [PubMed]
- Matilla-Cuenca, L.; Gil, C.; Cuesta, S.; Rapún-Araiz, B.; Žiemytė, M.; Mira, A.; Lasa, I.; Valle, J. Antibiofilm Activity of Flavonoids on Staphylococcal Biofilms through Targeting BAP Amyloids. Sci. Rep. 2020, 10, 18968. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, Y.; Ma, R.; Sun, W.; Ji, Z. Antibacterial Activity of Epigallocatechin Gallate (EGCG) Against Shigella flexneri. Int. J. Environ. Res. Public Health 2023, 20, 4676. [Google Scholar] [CrossRef]
- Paluch, E.; Bortkiewicz, O.; Widelski, J.; Duda-Madej, A.; Gleńsk, M.; Nawrot, U.; Lamch, Ł.; Długowska, D.; Sobieszczańska, B.; Wilk, K.A. A Combination of β-Aescin and Newly Synthesized Alkylamidobetaines as Modern Components Eradicating the Biofilms of Multidrug-Resistant Clinical Strains of Candida glabrata. Int. J. Mol. Sci. 2024, 25, 2541. [Google Scholar] [CrossRef]
- Antolak, H.; Mizerska, U.; Berłowska, J.; Otlewska, A.; Kręgiel, D. Quillaja saponaria Saponins with Potential to Enhance the Effectiveness of Disinfection Processes in the Beverage Industry. Appl. Sci. 2018, 8, 368. [Google Scholar] [CrossRef]
- Ratti, A.; Fassi, E.M.A.; Forlani, F.; Mori, M.; Villa, F.; Cappitelli, F.; Sgrignani, J.; Roda, G.; Cavalli, A.; Villa, S.; et al. Mechanistic Insights into the Antibiofilm Mode of Action of Ellagic Acid. Pharmaceutics 2023, 15, 1757. [Google Scholar] [CrossRef]
- Ulrey, R.K.; Barksdale, S.M.; Zhou, W.; van Hoek, M.L. Cranberry Proanthocyanidins Have Anti-Biofilm Properties Against Pseudomonas aeruginosa. BMC Complement. Altern. Med. 2014, 14, 499. [Google Scholar] [CrossRef]
- Qin, T.; Chen, K.; Xi, B.; Pan, L.; Xie, J.; Lu, L.; Liu, K. In Vitro Antibiofilm Activity of Resveratrol Against Aeromonas hydrophila. Antibiotics 2023, 12, 686. [Google Scholar] [CrossRef]
- Asadi, S.; Nayeri-Fasaei, B.; Zahraei-Salehi, T.; Yahya-Rayat, R.; Shams, N.; Sharifi, A. Antibacterial and Anti-Biofilm Properties of Carvacrol Alone and in Combination with Cefixime Against Escherichia coli. BMC Microbiol. 2023, 23, 55. [Google Scholar] [CrossRef]
- Bisso Ndezo, B.; Tokam Kuaté, C.R.; Dzoyem, J.P. Synergistic Antibiofilm Efficacy of Thymol and Piperine in Combination with Three Aminoglycoside Antibiotics Against Klebsiella pneumoniae Biofilms. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 7029944. [Google Scholar] [CrossRef]
- Albano, C.; Nabawy, A.; Tran, W.C.; Prithviraj, M.; Kado, T.; Hassan, M.A.; Makabenta, J.M.V.; Rotello, V.M.; Morita, Y.S.; Neyrolles, O. Effective killing of Mycobacterium abscessus biofilm by nanoemulsion delivery of plant phytochemicals. Microbiology Spectrum. 2025, 13, e02166-24. [Google Scholar] [CrossRef] [PubMed]
- Iskandar, K.; Murugaiyan, J.; Hammoudi Halat, D.; Hage, S.E.; Chibabhai, V.; Adukkadukkam, S.; Roques, C.; Molinier, L.; Salameh, P.; Van Dongen, M. Antibiotic Discovery and Resistance: The Chase and the Race. Antibiotics 2022, 11, 182. [Google Scholar] [CrossRef]
- Halawa, E.M.; Fadel, M.; Al-Rabia, M.W.; Behairy, A.; Nouh, N.A.; Abdo, M.; Olga, R.; Fericean, L.; Atwa, A.M.; El-Nablaway, M.; et al. Antibiotic Action and Resistance: Updated Review of Mechanisms, Spread, Influencing Factors, and Alternative Approaches for Combating Resistance. Front. Pharmacol. 2024, 14, 1305294. [Google Scholar] [CrossRef]
- Manner, S.; Fallarero, A. Screening of Natural Product Derivatives Identifies Two Structurally Related Flavonoids as Potent Quorum Sensing Inhibitors Against Gram-Negative Bacteria. Int. J. Mol. Sci. 2018, 19, 1346. [Google Scholar] [CrossRef]
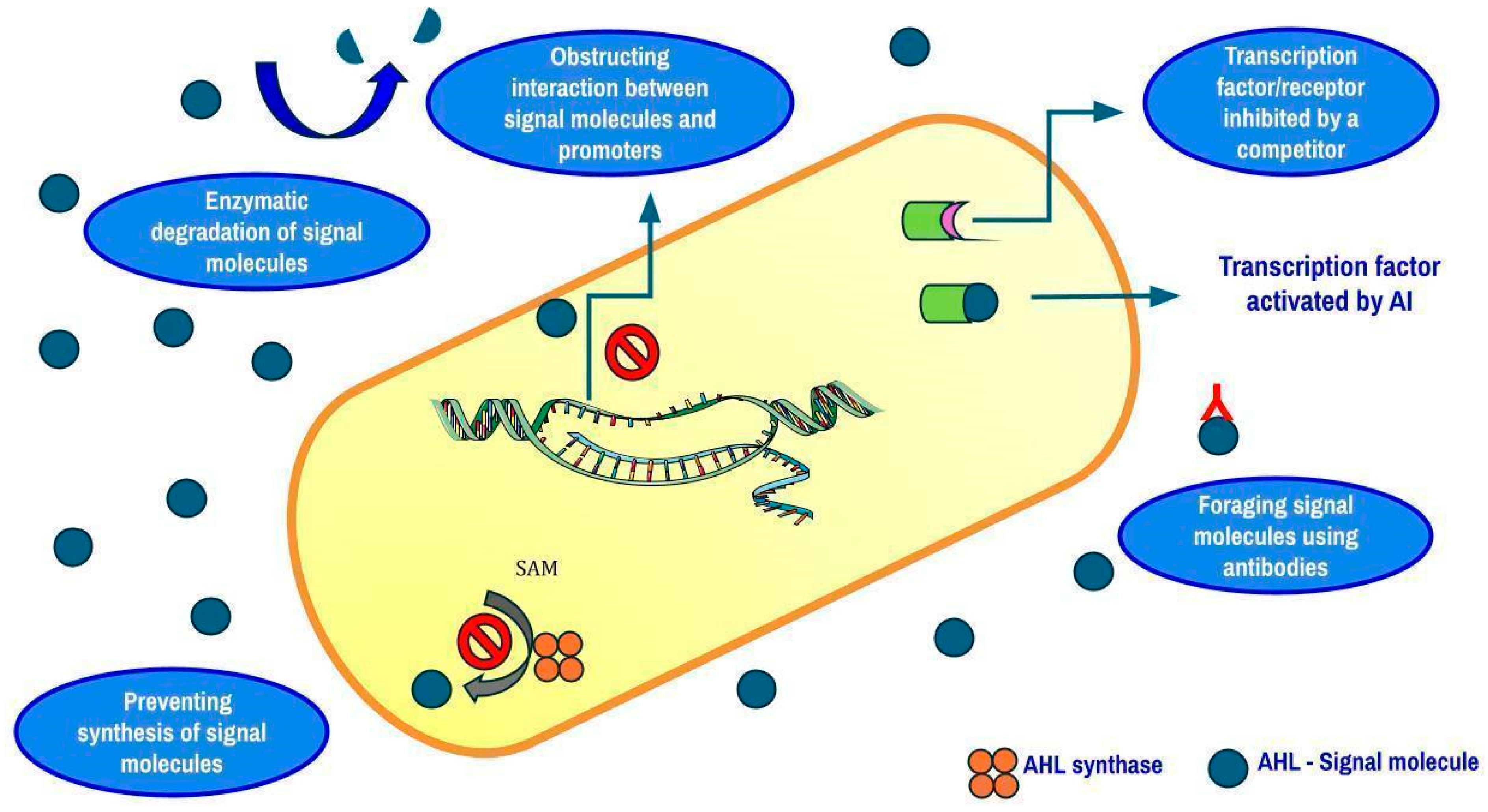
- Sikdar, R.; Elias, M. Quorum Quenching Enzymes and Their Effects on Virulence, Biofilm and Microbiomes: A Review of Recent Advances. Expert Rev. Anti Infect. Ther. 2020, 18, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Rehman, Z.U.; Momin, A.A.; Aldehaiman, A.; Irum, T.; Grünberg, R.; Arold, S.T. The Exceptionally Efficient Quorum Quenching Enzyme LrsL Suppresses Pseudomonas aeruginosa Biofilm Production. Front. Microbiol. 2022, 13, 977673. [Google Scholar]
- Fong, J.; Zhang, C.; Yang, R.; Boo, Z.Z.; Tan, S.K.; Nielsen, T.E.; Givskov, M.; Liu, X.-W.; Bin, W.; Su, H.; et al. Combination Therapy Strategy of Quorum Quenching Enzyme and Quorum Sensing Inhibitor in Suppressing Multiple Quorum Sensing Pathways of P. aeruginosa. Sci. Rep. 2018, 8, 1155. [Google Scholar] [CrossRef]
- Naga, N.G.; Zaki, A.A.; El-Badan, D.E.; Rateb, H.S.; Ghanem, K.M.; Shaaban, M.I. Inhibition of Pseudomonas aeruginosa Quorum Sensing by Methyl Gallate from Mangifera indica. Sci. Rep. 2023, 13, 17942. [Google Scholar] [CrossRef]
- Hetta, H.F.; Ramadan, Y.N.; Rashed, Z.I.; Alharbi, A.A.; Alsharef, S.; Alkindy, T.T.; Alkhamali, A.; Albalawi, A.S.; Battah, B.; Donadu, M.G. Quorum Sensing Inhibitors: An Alternative Strategy to Win the Battle Against Multidrug-Resistant (MDR) Bacteria. Molecules. 2024, 29, 3466. [Google Scholar] [CrossRef]
- Mitropoulou, G.; Karapantzou, I.; Tsimogiannis, D.; Oreopoulou, V.; Lazăr, V.; Kourkoutas, Y. Inhibitory Effects of Essential Oils and Extracts of the Water-Steam Distillation Residues from Greek Herbs on Adherent Biofilm Formation by Common Pathogens. Appl. Sci. 2025, 15, 3385. [Google Scholar] [CrossRef]
- Chimi, L.Y.; Bisso, B.N.; Njateng, G.S.S.; Dzoyem, J.P. Antibiotic-Potentiating Effect of Some Bioactive Natural Products Against Planktonic Cells, Biofilms, and Virulence Factors of Pseudomonas aeruginosa. BioMed. Res. Int. 2023, 2023, 9410609. [Google Scholar] [CrossRef]
- Getahun, M.; Nesru, Y.; Ahmed, M.; Satapathy, S.; Shenkute, K.; Gupta, N.; Naimuddin, M. Phytochemical Composition, Antioxidant, Antimicrobial, Antibiofilm, and Antiquorum Sensing Potential of Methaol Extract and Essential Oil from Acanthus polystachyus Delile (Acanthaceae). ACS Omega 2023, 8, 43024–43036. [Google Scholar] [CrossRef]
- Nikolic, I.; Aleksic Sabo, V.; Gavric, D.; Knezevic, P. Anti-Staphylococcus aureus Activity of Volatile Phytochemicals and Their Combinations with Conventional Antibiotics Against Methicillin-Susceptible S. aureus (MSSA) and Methicillin-Resistant S. aureus (MRSA) Strains. Antibiotics 2024, 13, 1030. [Google Scholar] [CrossRef]
- Boriollo, M.F.G.; Marques, M.B.; da Silva, T.A.; Da Silva, J.J.; Dias, R.A.; Silva Filho, T.H.N.; Melo, I.L.R.; dos Santos Dias, C.T.; Bernardo, W.L.D.C.; de Mello Silva Oliveira, N.; et al. Antimicrobial potential, phytochemical profile, cytotoxic and genotoxic screening of Sedum praealtum A. DC. (balsam). BMC Complement. Med. Ther. 2020, 20, 133. [Google Scholar] [CrossRef]
- Anywar, G.U.; Kakudidi, E.; Oryem-Origa, H.; Schubert, A.; Jassoy, C. Cytotoxicity of Medicinal Plant Species Used by Traditional Healers in Treating People Suffering from HIV/AIDS in Uganda. Front. Toxicol. 2022, 4, 832780. [Google Scholar] [CrossRef]
- Sun, Y.; Mao, W.; Cao, J.; Hao Pgao Jianguo, S.; Yin, K.; Gu, K.; Zhao, H. Chinese Medicine Monomers Inhibit Biofilm Formation in Multidrug-Resistant Pasteurella multocida Isolated from Cattle Respiratory Infections. Pak. Vet. J. 2024, 44, 1095–1104. [Google Scholar] [CrossRef]
- Alves-Barroco, C.; Botelho, A.M.N.; Américo, M.A.; Fracalanzza, S.E.L.; de Matos, A.P.A.; Guimaraes, M.A.; Ferreira-Carvalho, B.T.; Figueiredo, A.M.S.; Fernandes, A.R. Assessing in vivo and in vitro biofilm development by Streptococcus dysgalactiae subsp. Dysgalactiae using a murine model of catheter-associated biofilm and human keratinocyte cell. Front. Cell. Infect. Microbiol. 2022, 12, 874694. [Google Scholar] [CrossRef]
- Santhosh, S.K.; Sarojini, S. Antibiofilm and antiquorum properties of ethanolic leaf extracts of Syzygium jambos and Psidium guajava and their gel formulation for wound healing applications. Plant Sci. Today 2025, 12, 2. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phytochemicals | Target Bacteria | Mode of Action | References |
|---|---|---|---|
| Luteolin (bioflavonoid) | Staphylococcus aureus | Kills both planktonic and biofilm-forming bacteria by disrupting the cell membrane and inhibiting biofilm formation | [78] |
| Catechols (7,8-dihydroxyflavone, myricetin, and luteolin) | Klebsiella pneumoniae | Disrupt iron homeostasis in bacteria—ferric ions were reduced to ferrous form—and increases ROS production, leading to cell ferroptosis | [79] |
| Flavonoids, phenols, saponins, steroids, terpenoids | Pseudomonas aeruginosa strain NAPCC-1 and strain KAR12. | Cause membrane destabilization and interrupts the life cycle by targeting important enzymes. | [80] |
| Alkaloids, quinones, steroids, terpenes, saponins, flavonoids, tannins | 20 bacterial strains representing 13 species | Disrupt bacterial membranes, leading to leakage of cellular contents and inhibition of essential cellular functions. | [81] |
| Linoleic acid, Lupeol, Epi-psi-Taraxastanonol | Staphylococcus aureus, Pseudomonas aeruginosa | Disturb bacterial cell membrane integrity, causing the leakage of cellular contents. | [82] |
| Eugenol, essential oils | Staphylococcus aureus (multi-resistant strain) Escherichia coli (multi-resistant strain) | Inhibit protein biosynthesis and disturbs the bacterial cell membrane. | [83] |
| Alkaloids, flavonoids, phenols, tannins, terpenoids | Enterobacter faecalis, Escherichia coli, Pseudomonas aeruginosa, Klebsiella pneumoniae | Breach the outer membrane to interfere with essential intracellular processes. | [84] |
| Stigmasterol, β-amyrin, 3-O-β-D-glucopyranosylstigmasterol, 3-O-β-galactopyranosyl-(1→4)-β-D-galactopyranosyl-oleanolic acid | Klebsiella pneumoniae, Pseudomonas aeruginosa and Escherichia coli | Inhibit bacterial efflux pumps, increasing intracellular antibiotic levels and combating multidrug resistance | [85] |
| Alcarinol, spathulenol, and phytol | Vancomycin-resistant Enterococci, M. tuberculosis Methicillin-resistant Staphylococcus aureus | Disrupt bacterial membranes and inhibit enzymes, causing cell death | [86] |
| Phytochemicals | Compounds | Mode of Action | Reference |
|---|---|---|---|
| Alkaloids | Berberine | Decreases the transcription of AgrA and thereby targets biofilm dispersal in S. aureus | [103] |
| Sanguinarine | Inhibits the biofilm formation in carbapenem-resistant Serratia marcescens (CRSM) by disrupting the cell-membrane integrity. | [104] | |
| Piperine | Downregulates icaA gene, resulting in a reduction in extracellular polysaccharide production, a reduction in cell-surface hydrophobicity, and the regulation of AgrA protein, resulting in interference with quorum-sensing and microbial motility components in Methicillin-resistant Staphylococcus aureus (MRSA) | [105] | |
| Flavonoids | Quercetin Myricetin Scutellarein | Inhibits the assembly of Bap-related amyloid-like structures in S. aureus and thus prevents biofilm formation | [106] |
| Epigallocatechin gallate | Prevents the formation of extracellular polysaccharide | [107] | |
| Saponins | β-Aescin | Alters the hydrophobicity of the cell surface and thereby eradicates biofilms | [108] |
| Quillaja saponaria saponins | Acts as a natural detergent to eradicate biofilms and thus is used in wastewater treatment | [109] | |
| Tannins | Ellagic acid | Disrupts bacterial redox balance via WrbA, thereby impairing biofilm formation. | [110] |
| Proanthocyanidin | Chelates iron, thereby preventing the biofilm from maturing completely or allowing only a thin layer of biofilm to form that is not very resistant to antibiotics | [111] | |
| Phenolics | Curcumin | Inhibits pellicle and pili formation and restricts motility | [102] |
| Resveratrol | Inhibits motility and quorum sensing in Aeromonas hydrophila | [112] | |
| Terpenes | Carvacrol | Inhibits quorum sensing in E. coli | [113] |
| Thymol | Inhibits quorum sensing and production of PIA | [114] |
| Pathogens | Natural Compounds | MIC | MBIC | MBEC | Reference |
|---|---|---|---|---|---|
| E. coli | Thymol | 2.4 mg/mL | 0.3 mg/mL | 1.2 mg/mL | [124] |
| Carvacrol | 0.6 mg/mL | 0.6 mg/mL | 2.4 mg/mL | ||
| p-Cymene | 0.7 mg/mL | 0.1 mg/mL | 0.5 mg/mL | ||
| γ-Terpinine | 2.2 mg/mL | 0.5 mg/mL | 2.1 mg/mL | ||
| P. aeruginosa | Curcumin | 417.68 μg/mL | 565.89 μg/mL | >2048 μg/mL | [125] |
| Piperine | >1024 μg/mL | >2048 μg/mL | >2048 μg/mL | ||
| Quercetin | 77.47 μg/mL | 52.21 μg/mL | 195.37 μg/mL | ||
| Plumbagin | 644.92 μg/mL | 1024–2048 μg/mL | >2048 μg/mL | ||
| A. baumannii | Methanol Extract from Acanthus polystachyus Delile | 0.5 mg/mL | 0.5 mg/mL | - | [126] |
| Essential Oil from Acanthus polystachyus Delile | 0.31 mg/mL | 0.31 mg/mL | - | ||
| Methicillin-Susceptible Staphylococcus aureus (MSSA) and Methicillin-Resistant Staphylococcus aureus (MRSA) strains | Volatile phytochemicals Carvacrol (CAR) and Thymol (THY) | CAR: 128–203.2 μg/mL for MSSA and 362–1024 μg/mL for MRSA THY: 256–724.01 μg/mL for MSSA and 512.0 ≥ 2048 μg/mL for MRSA | - | - | [127] |
| K. pneumoniae | Thyme essential oils | 6 μL/mL | - | - | [76] |
| Lavender essential oils | 6 μL/mL | - | - | ||
| Mint essential oils | 6 μL/mL | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sarojini, S.; Jayaram, S.; Kalathilparambil Santhosh, S.; Priyadarshini, P.; Pappuswamy, M.; Balasubramanian, B. Harnessing the Potential of Antibacterial and Antibiofilm Phytochemicals in the Combat Against Superbugs: A One Health Perspective. Antibiotics 2025, 14, 692. https://doi.org/10.3390/antibiotics14070692
Sarojini S, Jayaram S, Kalathilparambil Santhosh S, Priyadarshini P, Pappuswamy M, Balasubramanian B. Harnessing the Potential of Antibacterial and Antibiofilm Phytochemicals in the Combat Against Superbugs: A One Health Perspective. Antibiotics. 2025; 14(7):692. https://doi.org/10.3390/antibiotics14070692
Chicago/Turabian StyleSarojini, Suma, Saranya Jayaram, Sandhya Kalathilparambil Santhosh, Pragyan Priyadarshini, Manikantan Pappuswamy, and Balamuralikrishnan Balasubramanian. 2025. "Harnessing the Potential of Antibacterial and Antibiofilm Phytochemicals in the Combat Against Superbugs: A One Health Perspective" Antibiotics 14, no. 7: 692. https://doi.org/10.3390/antibiotics14070692
APA StyleSarojini, S., Jayaram, S., Kalathilparambil Santhosh, S., Priyadarshini, P., Pappuswamy, M., & Balasubramanian, B. (2025). Harnessing the Potential of Antibacterial and Antibiofilm Phytochemicals in the Combat Against Superbugs: A One Health Perspective. Antibiotics, 14(7), 692. https://doi.org/10.3390/antibiotics14070692