

Antimicrobial Activity of Greek Native Essential Oils Against Escherichia coli O157:H7 and Antibiotic Resistance Strains Harboring pNorm Plasmid, mecA, mcr-1 and blaOXA Genes


Abstract
1. Introduction
2. Results
2.1. Disk Diffusion Assay
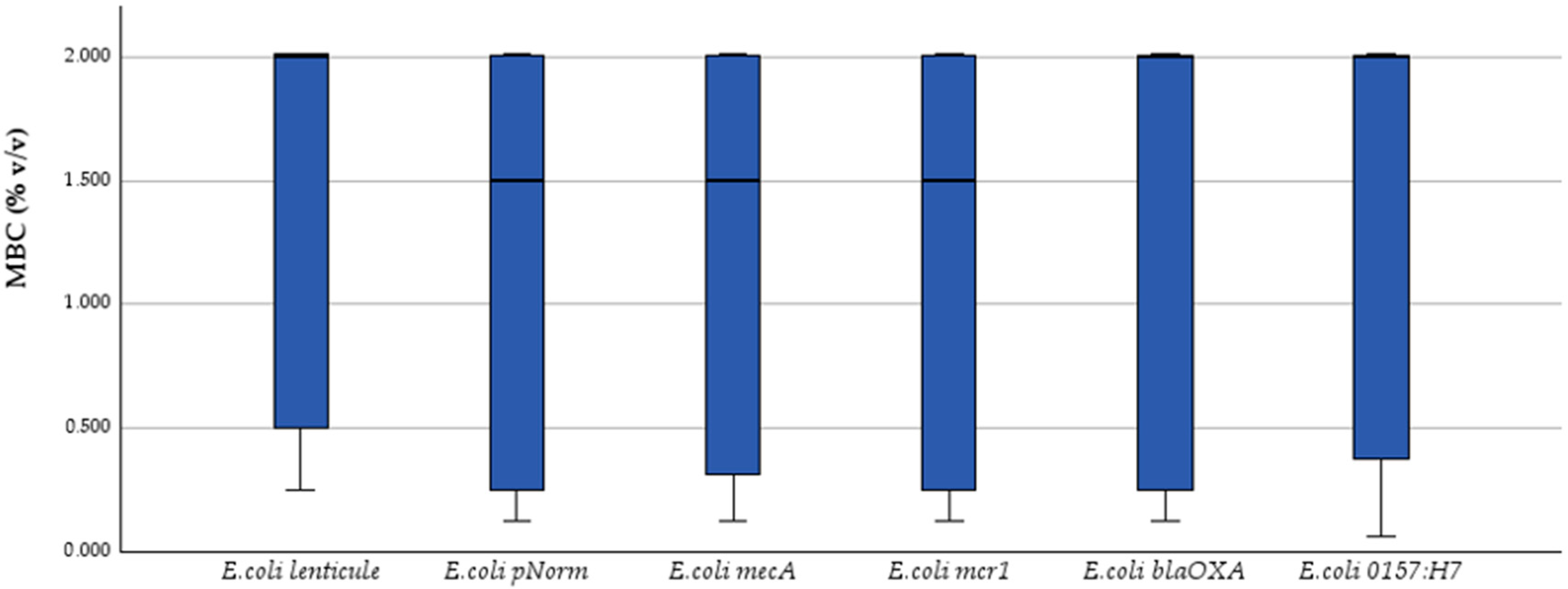
2.2. MIC and MBC
2.3. Statistical Analysis
3. Discussion
4. Materials and Methods
4.1. Essential Oils
4.2. GC–MS Chemical Profiling of Essential Oils
4.3. Bacterial Strains
4.4. Disk Diffusion Method
4.5. MIC and MBC Determination
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMR | Antimicrobial Resistance |
| EO | Essential Oil |
| MIC | Minimum Inhibitory Concentration |
| MBC | Minimum Bactericidal Concentration |
| CC-MS | Gass Chromatography–Mass Spectrometry |
| LPS | Lipopolysaccharide |
| GRAS | Generally Recognized As Safe |
| FDA | Food and Drug Administration |
| M.H | Monoterpene Hydrocarbons |
| O.M | Oxygenated Monoterpenes |
| S.H | Sesquiterpene Hydrocarbons |
| O.S | Oxygenated Sesquiterpenes |
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Robles Aguilar, G.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Ahmed, S.K.; Hussein, S.; Qurbani, K.; Ibrahim, R.H.; Fareeq, A.; Mahmood, K.A.; Mohamed, M.G. Antimicrobial resistance: Impacts, challenges, and future prospects. J. Med. Surg. Public Health 2024, 2, 100081. [Google Scholar] [CrossRef]
- Ferraz, M.P. Antimicrobial Resistance: The Impact from and on Society According to One Health Approach. Societies 2024, 14, 187. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Li, M.; Jing, Y.; Liu, K.; Wu, Y.; Peng, Z. What Are the Drivers Triggering Antimicrobial Resistance Emergence and Spread? Outlook from a One Health Perspective. Antibiotics 2025, 14, 543. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Resistance in the Environment and the Food Supply: Causes and How It Spreads|Antimicrobial Resistance|CDC. Available online: https://www.cdc.gov/antimicrobial-resistance/causes/environmental-food.html (accessed on 27 June 2025).
- Antimicrobial Resistance, Food, and Food Animals|Food Safety|CDC. Available online: https://www.cdc.gov/food-safety/foods/antimicrobial-resistance.html (accessed on 27 June 2025).
- Ramos, S.; Silva, V.; de Lurdes Enes Dapkevicius, M.; Caniça, M.; Tejedor-Junco, M.T.; Igrejas, G.; Poeta, P. Escherichia coli as Commensal and Pathogenic Bacteria among Food-Producing Animals: Health Implications of Extended Spectrum β-Lactamase (ESBL) Production. Animals 2020, 10, 2239. [Google Scholar] [CrossRef] [PubMed]
- Hembach, N.; Schmid, F.; Alexander, J.; Hiller, C.; Rogall, E.T.; Schwartz, T. Occurrence of the mcr-1 colistin resistance gene and other clinically relevant antibiotic resistance genes in microbial populations at different municipal wastewater treatment plants in Germany. Front. Microbiol. 2017, 8, 1282. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Liang, B.; Xu, X.; Yang, L.; Li, H.; Li, P.; Qiu, S.; Song, H. Identification of mcr-1-positive multidrug-resistant Escherichia coli isolates from clinical samples in Shanghai, China. J. Glob. Antimicrob. Resist. 2022, 29, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Loucif, L.; Chelaghma, W.; Bendjama, E.; Cherak, Z.; Khellaf, M.; Khemri, A.; Rolain, J.M. Detection of blaOXA-48 and mcr-1 Genes in Escherichia coli Isolates from Pigeon (Columba livia) in Algeria. Microorganisms 2022, 10, 975. [Google Scholar] [CrossRef] [PubMed]
- Tyasningsih, W.; Ramandinianto, S.C.; Ansharieta, R.; Witaningrum, A.M.; Permatasari, D.A.; Wardhana, D.K.; Effendi, M.H.; Ugbo, E.N. Prevalence and antibiotic resistance of Staphylococcus aureus and Escherichia coli isolated from raw milk in East Java, Indonesia. Vet. World 2022, 15, 2021. [Google Scholar] [CrossRef] [PubMed]
- Jangid, H.; Kumar, D.; Kumar, G.; Kumar, R.; Mamidi, N. An Emerging Foodborne Pathogen Spotlight: A Bibliometric Analysis and Scholarly Review of Escherichia coli O157 Research. Antibiotics 2024, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Yammine, J.; Chihib, N.E.; Gharsallaoui, A.; Dumas, E.; Ismail, A.; Karam, L. Essential oils and their active components applied as: Free, encapsulated and in hurdle technology to fight microbial contaminations. A review. Heliyon 2022, 8, e12472. [Google Scholar] [CrossRef] [PubMed]
- Maggio, F.; Rossi, C.; Serio, A.; Chaves-Lopez, C.; Casaccia, M.; Paparella, A. Anti-biofilm mechanisms of action of essential oils by targeting genes involved in quorum sensing, motility, adhesion, and virulence: A review. Int. J. Food Microbiol. 2025, 426, 110874. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, Z.; Čabarkapa, I.; Šarić, L.; Pajić, M.; Tomanić, D.; Kokić, B.; Božić, D.D. Natural Solutions to Antimicrobial Resistance: The Role of Essential Oils in Poultry Meat Preservation with Focus on Gram-Negative Bacteria. Foods 2024, 13, 3905. [Google Scholar] [CrossRef] [PubMed]
- Bibow, A.; Oleszek, W. Essential Oils as Potential Natural Antioxidants, Antimicrobial, and Antifungal Agents in Active Food Packaging. Antibiotics 2024, 13, 1168. [Google Scholar] [CrossRef] [PubMed]
- Alexopoulos, A.; Kimbaris, A.C.; Plessas, S.; Mantzourani, I.; Theodoridou, I.; Stavropoulou, E.; Polissiou, M.G.; Bezirtzoglou, E. Antibacterial activities of essential oils from eight Greek aromatic plants against clinical isolates of Staphylococcus aureus. Anaerobe 2011, 17, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, H.; Papadopoulou, C. Antimicrobial activity of basil, oregano, and thyme essential oils. J. Microbiol. Biotechnol. 2017, 27, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Moussa, M.M.; Mazrou, R.M.; Hassan, F.A.S. How Sage and Rosemary Essential Oils Regulate Postharvest Senescence and Extend the Vase Life of Cut Gladiolus Spikes. Horticulturae 2024, 10, 638. [Google Scholar] [CrossRef]
- Fokas, R.; Koukousianou, V.; Strataki, A.; Sinou, P.S.; Lamari, F.N.; Vantarakis, A. Comparative invitro evaluation of the antimicrobial properties of essential oils from Lamiaceae, Cistaceae, and Asteraceae families against Enterococcus faecalis. J. Food Sci. Nutr. Res. 2024, 7, 188–197. [Google Scholar] [CrossRef]
- Soliman, A.M.; Shaaban, M.T.; Dawood, A.S.; Shaheen, M.N.; Salama, H.S. Microbiological studies on Escherichia coli and Klebsiella pneumoniae causing vaginal and urinary tract inflammation; prevalence, antibiotics-resistance and natural products susceptibility. Microb. Pathog. 2024, 196, 107000. [Google Scholar] [CrossRef] [PubMed]
- Sobhy, M.; Abdelkarim, E.A.; Hussein, M.A.; Aziz, T.; Al-Asmari, F.; Alabbosh, K.F.; Cui, H.; Lin, L. Essential oils as antibacterials against multidrug-resistant foodborne pathogens: Mechanisms, recent advances, and legal considerations. Food Biosci. 2025, 64, 105937. [Google Scholar] [CrossRef]
- Lupia, C.; Castagna, F.; Bava, R.; Naturale, M.D.; Zicarelli, L.; Marrelli, M.; Statti, G.; Tilocca, B.; Roncada, P.; Britti, D.; et al. Use of Essential Oils to Counteract the Phenomena of Antimicrobial Resistance in Livestock Species. Antibiotics 2024, 13, 163. [Google Scholar] [CrossRef] [PubMed]
- Fournomiti, M.; Kimbaris, A.; Mantzourani, I.; Plessas, S.; Theodoridou, I.; Papaemmanouil, V.; Kapsiotis, I.; Panopoulou, M.; Stavropoulou, E.; Bezirtzoglou, E.E.; et al. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (Salvia officinalis), and thyme (Thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microb. Ecol. Health Dis. 2015, 26, 23289. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, S.; Tsvetkova, A.; Stamova, S.; Ermenlieva, N.; Tsankova, G.; Georgieva, E.; Peycheva, K.; Panayotova, V.; Voynikov, Y. Antibacterial Effects of Bulgarian Oregano and Thyme Essential Oils Alone and in Combination with Antibiotics Against Klebsiella pneumoniae and Pseudomonas aeruginosa. Microorganisms 2025, 13, 843. [Google Scholar] [CrossRef] [PubMed]
- Santurio, F.; de Jesus, K.; Pantella, F.; Adriel, R.; Schlemmer, B.; Fries, M.; Lucy, L.; Scientiae, A.; Flores Santurio, D.; Pantella Kunz de Jesus, F.; et al. Sistema de Información Científica Antimicrobial Activity of the Essential Oil of Thyme and of Thymol against Escherichia coli Strains. Acta Sci. Vet. 2014, 42, 1–4. [Google Scholar]
- Mitropoulou, G.; Fitsiou, E.; Stavropoulou, E.; Papavassilopoulou, E.; Vamvakias, M.; Pappa, A.; Oreopoulou, A.; Kourkoutas, Y. Composition, antimicrobial, antioxidant, and antiproliferative activity of Origanum dictamnus (dittany) essential oil. Microb. Ecol. Health Dis. 2015, 26, 26543. [Google Scholar] [CrossRef] [PubMed]
- El Maimouni, M.A.; El Amrani, S.; Lairini, S.; Bouslamti, R.; Er-Rajy, M.; Mujwar, S.; Chalkha, M.; Chebbac, K.; El Akhal, F.; El Ouali Lalami, A. Gas Chromatography-Mass Spectrometry Profiling, Antioxidant and Antibacterial Evaluations of Essential Oils Extracted from Mentha piperita and Mentha pulegium. Trop. J. Nat. Prod. Res. 2024, 8, 9386–9398. [Google Scholar] [CrossRef]
- Puvača, N.; Milenković, J.; Galonja Coghill, T.; Bursić, V.; Petrović, A.; Tanasković, S.; Pelić, M.; Ljubojević Pelić, D.; Miljković, T. Antimicrobial Activity of Selected Essential Oils against Selected Pathogenic Bacteria: In Vitro Study. Antibiotics 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Kabotso, D.E.K.; Neglo, D.; Gaba, S.E.; Danyo, E.K.; Dayie, A.D.; Asantewaa, A.A.; Kotey, F.C.N.; Dayie, N.T.K.D. In Vitro Evaluation of Rosemary Essential Oil: GC-MS Profiling, Antibacterial Synergy, and Biofilm Inhibition. Pharmaceuticals 2024, 17, 1653. [Google Scholar] [CrossRef] [PubMed]
- Zych, S.; Adaszyńska-Skwirzyńska, M.; Szewczuk, M.A.; Szczerbińska, D. Interaction between Enrofloxacin and Three Essential Oils (Cinnamon Bark, Clove Bud and Lavender Flower)—A Study on Multidrug-Resistant Escherichia coli Strains Isolated from 1-Day-Old Broiler Chickens. Int. J. Mol. Sci. 2024, 25, 5220. [Google Scholar] [CrossRef] [PubMed]
- Chtibi, H.; Harboul, K.; Benali, T.; Bouyahya, A.; Abdelamjid, K.; Louahlia, S.; Chew, J.; Ming, L.C.; Hammani, K. Comparative Study of Antibacterial Activity of Cistus ladanifer L. Leaves Extracted by Ultrasound-Assisted Extraction and Maceration. Prog. Microbes Mol. Biol. 2023, 6, a0000394. [Google Scholar] [CrossRef]
- Djihane, B.; Wafa, N.; Elkhamssa, S.; Pedro, D.H.J.; Maria, A.E.; Mohamed Mihoub, Z. Chemical constituents of Helichrysum italicum (Roth) G. Don essential oil and their antimicrobial activity against Gram-positive and Gram-negative bacteria, filamentous fungi and Candida albicans. Saudi Pharm. J. 2017, 25, 780–787. [Google Scholar] [CrossRef] [PubMed]
- Bhavaniramya, S.; Vishnupriya, S.; Al-Aboody, M.S.; Vijayakumar, R.; Baskaran, D. Role of essential oils in food safety: Antimicrobial and antioxidant applications. Grain Oil Sci. Technol. 2019, 2, 49–55. [Google Scholar] [CrossRef]
- Kachur, K.; Suntres, Z. The antibacterial properties of phenolic isomers, carvacrol and thymol. Crit. Rev. Food Sci. Nutr. 2020, 60, 3042–3053. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; You, Y.; Wang, N.; Wang, M.; Song, T.; He, Y.; Zou, Y.; He, Y.; Peng, T.; Mei, L. Development of amphipathic derivatives of thymol and carvacrol as potent broad-spectrum antibacterial agents. Eur. J. Med. Chem. 2024, 276, 116716. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; Von Wright, A. Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Lu, M.; Dai, T.; Murray, C.K.; Wu, M.X. Bactericidal Property of Oregano Oil Against Multidrug-Resistant Clinical Isolates. Front. Microbiol. 2018, 9, 2329. [Google Scholar] [CrossRef] [PubMed]
- Scandorieiro, S.; de Camargo, L.C.; Lancheros, C.A.C.; Yamada-Ogatta, S.F.; Nakamura, C.V.; de Oliveira, A.G.; Andrade, C.G.T.J.; Duran, N.; Nakazato, G.; Kobayashi, R.K.T. Synergistic and additive effect of oregano essential oil and biological silver nanoparticles against multidrug-resistant bacterial strains. Front. Microbiol. 2016, 7, 181296. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Wang, J.; Ahn, J. Advances in the Discovery of Efflux Pump Inhibitors as Novel Potentiators to Control Antimicrobial-Resistant Pathogens. Antibiotics 2023, 12, 1417. [Google Scholar] [CrossRef] [PubMed]
- Marchese, A.; Arciola, C.R.; Barbieri, R.; Silva, A.S.; Nabavi, S.F.; Sokeng, A.J.T.; Izadi, M.; Jafari, N.J.; Suntar, I.; Daglia, M.; et al. Update on Monoterpenes as Antimicrobial Agents: A Particular Focus on p-Cymene. Materials 2017, 10, 947. [Google Scholar] [CrossRef] [PubMed]
- Bairamis, A.; Sotiropoulou, N.S.D.; Tsadila, C.; Tarantilis, P.; Mossialos, D. Chemical Composition and Antimicrobial Activity of Essential Oils and Hydrosols from Oregano, Sage and Pennyroyal against Oral Pathogens. Appl. Sci. 2024, 14, 3238. [Google Scholar] [CrossRef]
- Tzortzakis, N. Origanum dictamnus Essential Oil in Vapour or Aqueous Solution Application for Pepper Fruit Preservation against Botrytis cinerea. Agronomy 2024, 14, 257. [Google Scholar] [CrossRef]
- Ferreira, A.P.; Almeida-Aguiar, C.; Costa, S.P.G.; Neves, I.C. Essential Oils Encapsulated in Zeolite Structures as Delivery Systems (EODS): An Overview. Molecules 2022, 27, 8525. [Google Scholar] [CrossRef] [PubMed]
- Cho, T.J.; Park, S.M.; Yu, H.; Seo, G.H.; Kim, H.W.; Kim, S.A.; Rhee, M.S. Recent advances in the application of antibacterial complexes using essential oils. Molecules 2020, 25, 1752. [Google Scholar] [CrossRef] [PubMed]
- Beigi, M.; Torki-Harchegani, M.; Pirbalouti, A.G. Quantity and chemical composition of essential oil of peppermint (Mentha × piperita l.) leaves under different drying methods. Int. J. Food Prop. 2018, 21, 267–276. [Google Scholar] [CrossRef]
- Khatri, P.K.; Paolini, M.; Larcher, R.; Ziller, L.; Magdas, D.A.; Marincas, O.; Roncone, A.; Bontempo, L. Botanical characterization and authentication of lavender essential oil using its volatile organic compounds and compound-specific carbon and hydrogen isotope ratio analysis. Food Control 2023, 154, 110002. [Google Scholar] [CrossRef]
- Aziz, E.; Batool, R.; Akhtar, W.; Shahzad, T.; Malik, A.; Shah, M.A.; Iqbal, S.; Rauf, A.; Zengin, G.; Bouyahya, A.; et al. Rosemary species: A review of phytochemicals, bioactivities and industrial applications. S. Afr. J. Bot. 2022, 151, 3–18. [Google Scholar] [CrossRef]
- Boy, F.R.; Benito, M.J.; Córdoba, M.D.G.; Rodríguez, A.; Casquete, R. Antimicrobial Properties of Essential Oils Obtained from Autochthonous Aromatic Plants. Int. J. Environ. Res. Public Health 2023, 20, 1657. [Google Scholar] [CrossRef] [PubMed]
- Furlan, V.; Bren, U. Helichrysum italicum: From Extraction, Distillation, and Encapsulation Techniques to Beneficial Health Effects. Foods 2023, 12, 802. [Google Scholar] [CrossRef] [PubMed]
- Ergüden, B. Phenol group of terpenoids is crucial for antibacterial activity upon ion leakage. Lett. Appl. Microbiol. 2021, 73, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Snyder, D.S.; McIntosh, T.J. The lipopolysaccharide barrier: Correlation of antibiotic susceptibility with antibiotic permeability and fluorescent probe binding kinetics. Biochemistry 2000, 39, 11777–11787. [Google Scholar] [CrossRef] [PubMed]
- Investigation Update: E. coli Outbreak, Onions Served at McDonald’s-November 13, 2024|E. coli Infection|CDC. Available online: https://www.cdc.gov/ecoli/outbreaks/investigation-update-e-coli-o157-2024.html (accessed on 29 June 2025).
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451. [Google Scholar] [CrossRef] [PubMed]
- Foda, A.M.; Kalaba, M.H.; El-Sherbiny, G.M.; Moghannem, S.A.; El-Fakharany, E.M. Antibacterial activity of essential oils for combating colistin-resistant bacteria. Expert Rev. Anti-Infect. Ther. 2022, 20, 1351–1364. [Google Scholar] [CrossRef] [PubMed]
- Ben Selma, W.; Farouk, A.; Ban, Z.; Ferjeni, M.; Alsulami, T.; Ali, H.; Boukadida, J. Thymus algeriensis essential oil: Phytochemical investigation, bactericidal activity, synergistic effect with colistin, molecular docking, and dynamics analysis against Gram-negative bacteria resistant to colistin. Heliyon 2024, 10, e38281. [Google Scholar] [CrossRef] [PubMed]
- Avci, F.G.; Tastekil, I.; Jaisi, A.; Ozbek Sarica, P.; Sariyar Akbulut, B. A review on the mechanistic details of OXA enzymes of ESKAPE pathogens. Pathog. Glob. Health 2022, 117, 219. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.S.X.; Yiap, B.C.; Ping, H.C.; Lim, S.H.E. Essential Oils, A New Horizon in Combating Bacterial Antibiotic Resistance. Open Microbiol. J. 2014, 8, 6. [Google Scholar] [CrossRef] [PubMed]
- Pournajaf, A.; Ardebili, A.; Goudarzi, L.; Khodabandeh, M.; Narimani, T.; Abbaszadeh, H. PCR-based identification of methicillin-resistant Staphylococcus aureus strains and their antibiotic resistance profiles. Asian Pac. J. Trop. Biomed. 2014, 4, S293–S297. [Google Scholar] [CrossRef] [PubMed]
- Kassem, I.I.; Esseili, M.A.; Sigler, V. Occurrence of mecA in nonstaphylococcal pathogens in surface waters. J. Clin. Microbiol. 2008, 46, 3868–3869. [Google Scholar] [CrossRef] [PubMed]
- Miragaia, M. Factors contributing to the evolution of Meca-mediated β-lactam resistance in staphylococci: Update and new insights from whole genome sequencing (WGS). Front. Microbiol. 2018, 9, 396897. [Google Scholar] [CrossRef] [PubMed]
- Visan, A.I.; Negut, I. Coatings Based on Essential Oils for Combating Antibiotic Resistance. Antibiotics 2024, 13, 625. [Google Scholar] [CrossRef] [PubMed]
- Selim, S. Antimicrobial Activity of Essential Oils Against Vancomycin-Resistant Enterococci (Vre) and Escherichia coli O157:H7 in Feta Soft Cheese and Minced Beef Meat. Braz. J. Microbiol. 2011, 42, 187. [Google Scholar] [CrossRef]
- Santiesteban-López, N.A.; Gómez-Salazar, J.A.; Santos, E.M.; Campagnol, P.C.B.; Teixeira, A.; Lorenzo, J.M.; Sosa-Morales, M.E.; Domínguez, R. Natural Antimicrobials: A Clean Label Strategy to Improve the Shelf Life and Safety of Reformulated Meat Products. Foods 2022, 11, 2613. [Google Scholar] [CrossRef] [PubMed]
- Asioli, D.; Aschemann-Witzel, J.; Caputo, V.; Vecchio, R.; Annunziata, A.; Næs, T.; Varela, P. Making sense of the “clean label” trends: A review of consumer food choice behavior and discussion of industry implications. Food Res. Int. 2017, 99, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Gurtler, J.B.; Garner, C.M. A Review of Essential Oils as Antimicrobials in Foods with Special Emphasis on Fresh Produce. J. Food Prot. 2022, 85, 1300–1319. [Google Scholar] [CrossRef] [PubMed]
- Gaba, A.B.M.; Hassan, M.A.; Abd El-Tawab, A.A.; Abdelmonem, M.A.; Morsy, M.K. Protective Impact of Chitosan Film Loaded Oregano and Thyme Essential Oil on the Microbial Profile and Quality Attributes of Beef Meat. Antibiotics 2022, 11, 583. [Google Scholar] [CrossRef] [PubMed]
- Sarengaowa; Hu, W.; Feng, K.; Xiu, Z.; Jiang, A.; Lao, Y. Thyme oil alginate-based edible coatings inhibit growth of pathogenic microorganisms spoiling fresh-cut cantaloupe. Food Biosci. 2019, 32, 100467. [Google Scholar] [CrossRef]
- Pires, A.; Pietruszka, H.; Bożek, A.; Szkolnicka, K.; Gomes, D.; Díaz, O.; Cobos, A.; Pereira, C. Sheep’s Second Cheese Whey Edible Coatings with Oregano and Clary Sage Essential Oils Used as Sustainable Packaging Material in Cheese. Foods 2024, 13, 674. [Google Scholar] [CrossRef] [PubMed]
- Casalini, S.; Baschetti, M.G.; Cappelletti, M.; Guerreiro, A.C.; Gago, C.M.; Nici, S.; Antunes, M.D. Antimicrobial activity of different nanocellulose films embedded with thyme, cinnamon, and oregano essential oils for active packaging application on raspberries. Front. Sustain. Food Syst. 2023, 7, 1190979. [Google Scholar] [CrossRef]
- Fokas, R.; Giormezis, N.; Vantarakis, A. Synergistic Approaches to Foodborne Pathogen Control: A Narrative Review of Essential Oils and Bacteriophages. Foods 2025, 14, 1508. [Google Scholar] [CrossRef] [PubMed]
- Fokas, R.; Kotsiri, Z.; Vantarakis, A. Can Bacteriophages Be Effectively Utilized for Disinfection in Animal-Derived Food Products? A Systematic Review. Pathogens 2025, 14, 291. [Google Scholar] [CrossRef] [PubMed]
- Yap, P.S.X.; Yusoff, K.; Lim, S.H.E.; Chong, C.M.; Lai, K.S. Membrane Disruption Properties of Essential Oils—A Double-Edged Sword? Processes 2021, 9, 595. [Google Scholar] [CrossRef]
- Cacace, D.; Fatta-Kassinos, D.; Manaia, C.M.; Cytryn, E.; Kreuzinger, N.; Rizzo, L.; Karaolia, P.; Schwartz, T.; Alexander, J.; Merlin, C.; et al. Antibiotic resistance genes in treated wastewater and in the receiving water bodies: A pan-European survey of urban settings. Water Res. 2019, 162, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Eucast: Disk Diffusion Methodology. Available online: https://www.eucast.org/ast_of_bacteria/disk_diffusion_methodology (accessed on 27 June 2025).



{kind=link}
{kind=link}
{kind=link}
| Essential Oil | E. coli (lenticule) | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 10.0 ± 0.0 | 9.3 ± 1.5 | 7.5 ± 0.5 | 7.0 ± 0.0 | 6.5 ± 0.5 | 0.0 ± 0.0 | 9.6 ± 0.4 | 9.0 ± 0.0 | 7.7 ± 0.3 | 7.2 ± 0.2 | 6.3 ± 0.3 | 6.1 ± 0.6 |
| Oregano | 22.1 ± 0.9 | 19.4 ± 0.5 | 18.4 ± 0.5 | 11.2 ± 1.1 | 9.2 ± 1.3 | 8.4 ± 0.5 | 21.4 ± 0.5 | 19.1 ± 0.9 | 17.5 ± 1.7 | 11.1 ± 1.0 | 9.1 ± 1.2 | 8.2 ± 0.8 |
| Dittany | 28.3 ± 0.6 | 24.0 ± 1.7 | 19.0 ± 0.0 | 12.7 ± 1.2 | 9.7 ± 1.5 | 9.3 ± 0.6 | 27.7 ± 0.6 | 21.7 ± 2.9 | 15.0 ± 2.0 | 12.0 ± 1.7 | 10.0 ± 1.0 | 9.7 ± 1.5 |
| Rosemary | 11.0 ± 1.0 | 11.7 ± 2.1 | 8.7 ± 0.6 | 6.3 ± 0.6 | 7.3 ± 0.6 | 0.0 ± 0.0 | 10.7 ± 0.6 | 10.0 ± 1.0 | 9.3 ± 0.6 | 7.3 ± 0.4 | 7.0 ± 0.6 | 0.0 ± 0.0 |
| Thyme | 24.0 ± 3.6 | 17.7 ± 1.2 | 15.0 ± 0.6 | 11.0 ± 0.6 | 8.3 ± 0.6 | 7.5 ± 1.3 | 23.3 ± 2.9 | 15.7 ± 1.2 | 14.3 ± 2.5 | 11.7 ± 2.3 | 8.0 ± 0.0 | 7.5 ± 1.3 |
| Lavender | 10.0 ± 0.0 | 10.0 ± 1.7 | 7.3 ± 0.6 | 7.0 ± 0.6 | 0.0 ± 0.0 | 0.0 ± 0.0 | 9.3 ± 0.6 | 9.0 ± 1.0 | 7.7 ± 0.6 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Cistus | 12.7 ± 0.6 | 11.5 ± 2.1 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 10.7 ± 2.9 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 7.0 ± 0.0 | 6.5 ± 0.7 | 6.5 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Essential Oil | E. coli (pNorm) | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 8.3 ± 0.6 | 7.3 ± 0.6 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 8 ± | 7.3 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Oregano | OMZ | 29.7 ± 0.6 | 26.7 ± 2.9 | 17.7 ± 0.6 | 14 ± 1.0 | 10.7 ± 0.6 | OMZ | 29.3 ± 1.2 | 24.3 ± 0.6 | 17 ± 0.0 | 12.7 ± 0.6 | 10 ± 0.0 |
| Dittany | 31.7 ± 2.9 | 28.3 ± 1.5 | 14.3 ± 0.6 | 11.3 ± 0.6 | 10 ± 0.0 | 8.3 ± 0.6 | 29.3 ± 1.2 | 27.3 ± 2.5 | 13.7 ± 0.6 | 11.3 ± 0.6 | 9.7 ± 0.6 | 8.7 ± 0.6 |
| Rosemary | 11 ± 1.7 | 8 ± 0.6 | 7.3 ± 0.6 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 11 ± 1.7 | 8.3 ± 0.6 | 7 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Thyme | OMZ | 36.7 ± 4.2 | 17.7 ± 2.5 | 12 ± 0.0 | 10 ± 0.0 | 8 ± 0.0 | OMZ | 36.7 ± 4.2 | 17.7 ± 2.5 | 12 ± 0.0 | 10 ± 0.0 | 8 ± 0.0 |
| Lavender | 9.7 ± 1.2 | 8 ± 0.0 | 7.0 ± 0.0 | 7.0 ± 0.0 | 7.0 ± 0.0 | 0.0 ± 0.0 | 9.7 ± 1.2 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 |
| Cistus | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7.0 ± 0.0 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7.0 ± 0.0 | 7.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Essential Oil | E. coli (mecA) | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 10 ± 0.0 | 10.7 ± 1.2 | 9.3 ± 1.2 | 7.7 ± 1.2 | 7.7 ± 1.5 | 7.5 ± 0.0 | 10 ± 0.0 | 10.7 ± 1.2 | 8.3 ± 0.6 | 7.7 ± 1.2 | 8 ± 0.0 | 7 ± 0.0 |
| Oregano | 26.3 ± 3.2 | 23 ± 1.0 | 20 ± 0.0 | 15.7 ± 2.1 | 12.3 ± 1.5 | 10 ± 0.0 | 26.7 ± 2.9 | 23 ± 2.0 | 20.7 ± 3.1 | 15 ± 1.0 | 12 ± 0.0 | 10 ± 0.0 |
| Dittany | 21.7 ± 1.5 | 18.3 ± 1.5 | 14.7 ± 0.6 | 11.3 ± 1.2 | 8.7 ± 0.6 | 7 ± 0.0 | 21 ± 1.7 | 17.3 ± 2.5 | 13.3 ± 0.6 | 10.3 ± 0.6 | 8 ± 1.0 | 7 ± 0.0 |
| Rosemary | 9 ± 1.0 | 8.3 ± 0.6 | 7.7 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 8 ± 0.6 | 8.3 ± 1.5 | 7.3 ± 0.6 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Thyme | 23.3 ± 2.9 | 19.3 ± 1.2 | 13.7 ± 1.2 | 11.3 ± 0.6 | 10 ± 0.0 | 7.7 ± 0.6 | 20.3 ± 0.6 | 17.3 ± 2.5 | 12.7 ± 1.5 | 10.7 ± 0.6 | 10 ± 0.0 | 8 ± 0.0 |
| Lavender | 10 ± 0.0 | 9.3 ± 1.2 | 8 ± 1.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 9.7 ± 0.6 | 9.3 ± 0.6 | 8.3 ± 1.5 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 |
| Cistus | 10.3 ± 2.5 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 10.7 ± 3.8 | 8 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0. | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Essential Oil | E. coli (mcr-1) | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 10.3 ± 0.6 | 9.3 ± 0.6 | 8 ± 1.0 | 7.7 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 | 9.7 ± 0.6 | 8.7 ± 0.6 | 7.7 ± 0.6 | 7.7 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 |
| Oregano | 24 ± 1.0 | 22 ± 1.0 | 20.7 ± 1.2 | 17.3 ± 0.6 | 13.3 ± 0.6 | 10.3 ± 0.6 | 24.7 ± 0.6 | 21 ± 1.0 | 20 ± 0.0 | 16 ± 1.7 | 12.7 ± 0.6 | 10 ± 0.0 |
| Dittany | 22 ± 1.0 | 21 ± 1.0 | 16 ± 1.7 | 10 ± 0.0 | 9 ± 0.0 | 8.3 ± 0.6 | 21 ± 1.0 | 20.3 ± 0.6 | 13.7 ± 0.6 | 9.3 ± 0.6 | 8 ± 0.0 | 8 ± 0.0 |
| Rosemary | 9 ± 0.0 | 8.7 ± 0.6 | 8 ± 0.0 | 7.3 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 8.7 ± 0.6 | 9 ± 1.0 | 8.3 ± 0.6 | 7.3 ± 0.6 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Thyme | 23.7 ± 0.6 | 19.3 ± 1.2 | 0.0 ± 0.0 | 10 ± 0.0 | 9.7 ± 0.6 | 8.3 ± 0.6 | 22.7 ± 1.5 | 15.7 ± 4.9 | 16 ± 1.7 | 10 ± 0.0 | 9.7 ± 0.6 | 8.7 ± 0.6 |
| Lavender | 10 ± 0.0 | 9 ± 0.0 | 8 ± 0.0 | 7.3 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 | 9.7 ± 0.6 | 9 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 |
| Cistus | 0.0 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Essential Oil | E. coli (blaOXA) | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 10 ± 1.0 | 8.3 ± 0.6 | 8 ± 0.0 | 8 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 8.7 ± 0.6 | 7.7 ± 0.6 | 7.3 ± 0.6 | 7.3 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 |
| Oregano | OMZ | 40.3 ± 1.5 | 30.7 ± 3.8 | 18.3 ± 0.6 | 13.7 ± 0.6 | 10 ± 0.0 | OMZ | 38.3 ± 2.9 | 32.3 ± 2.5 | 18.3 ± 0.6 | 13.3 ± 0.6 | 10.3 ± 0.6 |
| Dittany | 35.7 ± 3.5 | 31.7 ± 1.5 | 21.3 ± 3.1 | 11.7 ± 0.6 | 9.2 ± 0.6 | 8.3 ± 0.6 | 35 ± 1.0 | 31.7 ± 1.5 | 19 ± 1.7 | 10.7 ± 1.2 | 9.16 ± 0.6 | 8.3 ± 0.6 |
| Rosemary | 8.7 ± 0.6 | 8 ± 0.0 | 8 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 8.3 ± 0.6 | 8 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 |
| Thyme | OMZ | 37.7 ± 2.5 | 26.7 ± 0.6 | 14.7 ± 0.6 | 10.7 ± 0.6 | 9.7 ± 0.6 | OMZ | 35.7 ± 1.2 | 26 ± 3.6 | 13.7 ± 0.6 | 11.3 ± 0.6 | 9.3 ± 0.6 |
| Lavender | 12.7 ± 1.2 | 10.3 ± 0.6 | 8.7 ± 0.6 | 8.3 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 | 11 ± 0.0 | 8.7 ± 0.6 | 8 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 |
| Cistus | 9.7 ± 1.5 | 8 ± 0.0 | 8 ± 0.0 | 7.3 ± 0.6 | 7.3 ± 0.6 | 7 ± 0.0 | 9.3 ± 1.2 | 8 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 0.0 ± 0.0 | 8 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 7.7 ± 0.6 | 7.3 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 |
| Essential Oil | E. coli O157:H7 | |||||||||||
| 24 h | 48 h and 72 h | |||||||||||
| 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | 50% | 25% | 12.5% | 6.25% | 3.125% | 1.56% | |
| Peppermint | 0.0 ± 0.0 | 8.7 ± 0.6 | 8 ± 1.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 8.3 ± 0.6 | 8 ± 1.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 |
| Oregano | 28 ± 1.0 | 23.3 ± 1.2 | 21 ± 1.7 | 17.3 ± 0.6 | 10.3 ± 0.6 | 10 ± 0.0 | 24.7 ± 0.6 | 21.3 ± 1.2 | 20 ± 1.0 | 16 ± 1.0 | 10 ± 0.0 | 9.7 ± 0.6 |
| Dittany | 23.7 ± 4.7 | 19 ± 1.0 | 16.3 ± 1.5 | 12.3 ± 0.6 | 9 ± 1.0 | 7.7 ± 0.6 | 22.7 ± 3.8 | 18.3 ± 1.5 | 15.3 ± 1.5 | 11 ± 1.0 | 8.7 ± 0.6 | 8 ± 0.0 |
| Rosemary | 7.7 ± 0.6 | 8.3 ± 1.5 | 7.7 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 8 ± 0.0 | 8 ± 1.7 | 7.7 ± 0.6 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Thyme | 25.3 ± 1.5 | 22.3 ± 1.5 | 15.5 ± 2.1 | 13 ± 1.0 | 10 ± 0.0 | 8 ± 0.0 | 23.7 ± 1.2 | 21.7 ± 2.1 | 14.5 ± 0.7 | 12 ± 1.0 | 9.3 ± 0.6 | 8 ± 0.0 |
| Lavender | 11.3 ± 0.6 | 9.7 ± 1.2 | 8.7 ± 1.2 | 7.7 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 | 10.3 ± 0.6 | 9 ± 1.0 | 8.7 ± 0.6 | 7.7 ± 0.6 | 7 ± 0.0 | 7 ± 0.0 |
| Cistus | 7.5 ± 0.7 | 7 ± 0.0 | 7.3 ± 0.6 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7.3 ± 0.6 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Helichrysum | 0.0 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 7 ± 0.0 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| Strain (E. coli) | Ampicillin | Erythromycin | Oxytetracycline | |||
|---|---|---|---|---|---|---|
| Zones (mm) | Interpretation | Zones (mm) | Interpretation | Zones (mm) | Interpretation | |
| lenticule | 23.0 | S | 13.0 | ND | 27.0 | ND |
| pNorm | 14.0 | R | 15.0 | ND | 34.0 | ND |
| mecA | 0.0 | R | 0.0 | ND | 0.0 | ND |
| mcr-1 | 0.0 | R | 0.0 | ND | 0.0 | ND |
| blaOXA | 0.0 | R | 11.0 | ND | 33.0 | ND |
| O157:H7 | 20.0 | S | 16.0 | ND | 26.0 | ND |
| EOs | E. coli Lenticule | E. coli (pNorm) | E. coli (mecA) | E. coli (mcr-1) | E. coli (blaOXA) | E. coli O157:H7 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | |
| Peppermint | 1% | 2% | 1% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% |
| Oregano | 0.03125% | 0.25% | 0.0625% | 0.25% | 0.125% | 0.125% | 0.125% | 0.125% | 0.03125% | 0.125% | 0.0625% | 0.0625% |
| Dittany | 0.125% | 0.5% | 0.25% | 0.5% | 0.5% | 0.5% | 0.25% | 0.25% | 0.25% | 0.25% | 0.5% | 0.5% |
| Rosemary | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% | 2% |
| Thyme | 0.5% | 0.5% | 0.125% | 0.125% | 0.125% | 0.125% | 0.25% | 0.25% | 0.25% | 0.25% | 0.25% | 0.25% |
| Lavender | 2% | >2% | 0.5% | 1% | 1% | 1% | 1% | 1% | 1% | 2% | 2% | 2% |
| Cistus | 2% | >2% | 2% | >2% | 2% | >2% | 2% | >2% | 2% | >2% | 2% | >2% |
| Helichrysum | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | 2% | >2% |
| Essential Oil | Botanical Name | Plant Family | Harvest Region | Plant Part Used |
|---|---|---|---|---|
| Peppermint | Mentha × piperita | Lamiaceae | Greece | Aerial parts |
| Oregano | Origanum vulgare subsp. hirtum (91.3% carvacrol) | Lamiaceae | Greece | Aerial parts of flowering plant |
| Dittany | Origanum dictamnus | Lamiaceae | Crete, Greece | Aerial parts of flowering plant |
| Rosemary | Rosmarinus officinalis | Lamiaceae | Greece | Aerial parts |
| Thyme | Thymus vulgaris | Lamiaceae | Greece | Flowering tops |
| Lavender | Lavandula angustifolia cv. Hemus and Lavandula angustifolia cv. | Lamiaceae | Nea Tenedos, Northern Greece | Dried flowers and flowering tops |
| Cistus | Cistus ladanifer | Cistaceae | Nea Tenedos, Northern Greece | Leaves |
| Helichrysum | Helichrysum italicum | Asteraceae | Vavdos, Halkidiki, Central Macedonia | Flowering tops |
| EOs | Volatile Metabolites (%) | Total (%) | Density (g/mL) 25 °C |
|---|---|---|---|
| Peppermint | Menthol (46), Menthone (24.7), 1.8-Cineole (6.7), Menthyl Acetate (5.4) | 91 | 0.896 |
| M.H. 2.50%, O.M. 88.60% | |||
| Oregano | Carvacrol (91.30) | 97.29 | 0.95 |
| M.H. 2.38%, O.M. 93.44%, S.H. 0.96%, O.S. 0.44% | |||
| Dittany | Carvacrol (54.81), p-Cymene (13.99), γ-Terpinene (8.33) | 95.06 | 0.95 |
| M.H. 32.57%, O.M. 57.76%, S.H. 3.70% | |||
| Rosemary | Camphor (26.91), α-Pinene (14.76), 1,8-Cineole (13.41), Camphene (7.31), β-Myrcene (5.15) | 93.22 | 0.879 |
| M.H. 39.65%, O.M. 50.98%, S.H. 1.09% | |||
| Thyme | p-Cymene (33.53), Thymol (26.59), 1,8-Cineole (6.25), Limonene (5.32) | 90.59 | 0.9189 |
| M.H. 47.61%, O.M. 39.06%, S.H. 2.28% | |||
| Lavender | Linalyl Acetate (31.82), Linalool (29.27), Cis-β-Ocimene (6.47), Terpinen-4-ol (5.05) | 97.31% | 0.885–0.905 |
| M.H. 11.46%, O.M. 57.76%, S.H. 6.18% | |||
| Cistus | α-Pinene (38.94) | 71.74 | 0.962 |
| M.H. 52.88%, O.M. 12.82%, S.H. 0.64%, O.S. 2.67% | |||
| Helichrysum | γ-Curcumene (14.94), α-Pinene (14.94), Neryl acetate (14.14), β-Selinene (5.09) | 79.17 | 0.9 |
| M.H. 18.23%, O.M. 19.71%, S.H. 37.30% | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fokas, R.; Anastopoulou, Z.; Vantarakis, A. Antimicrobial Activity of Greek Native Essential Oils Against Escherichia coli O157:H7 and Antibiotic Resistance Strains Harboring pNorm Plasmid, mecA, mcr-1 and blaOXA Genes. Antibiotics 2025, 14, 741. https://doi.org/10.3390/antibiotics14080741
Fokas R, Anastopoulou Z, Vantarakis A. Antimicrobial Activity of Greek Native Essential Oils Against Escherichia coli O157:H7 and Antibiotic Resistance Strains Harboring pNorm Plasmid, mecA, mcr-1 and blaOXA Genes. Antibiotics. 2025; 14(8):741. https://doi.org/10.3390/antibiotics14080741
Chicago/Turabian StyleFokas, Rafail, Zoi Anastopoulou, and Apostolos Vantarakis. 2025. "Antimicrobial Activity of Greek Native Essential Oils Against Escherichia coli O157:H7 and Antibiotic Resistance Strains Harboring pNorm Plasmid, mecA, mcr-1 and blaOXA Genes" Antibiotics 14, no. 8: 741. https://doi.org/10.3390/antibiotics14080741
APA StyleFokas, R., Anastopoulou, Z., & Vantarakis, A. (2025). Antimicrobial Activity of Greek Native Essential Oils Against Escherichia coli O157:H7 and Antibiotic Resistance Strains Harboring pNorm Plasmid, mecA, mcr-1 and blaOXA Genes. Antibiotics, 14(8), 741. https://doi.org/10.3390/antibiotics14080741